



The Importance of Thermally Abnormal Waters for Bioinvasions—A Case Study of Pistia stratiotes



Abstract
:1. Introduction
2. Materials and Methods
2.1. Literature Review
2.2. Study Species
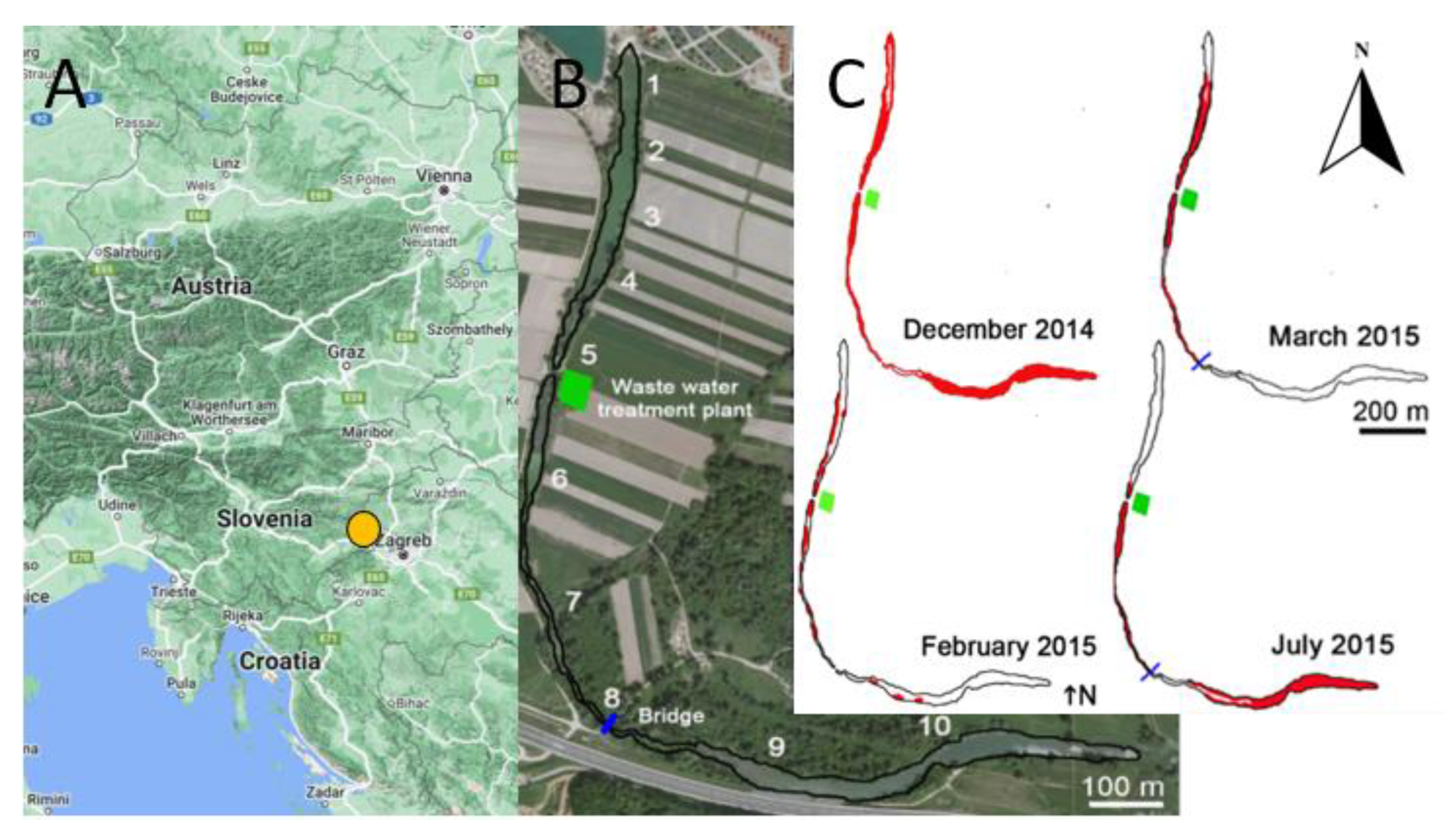
2.3. Study Site
2.4. Sampling Design
2.4.1. Abiotic Parameters
- Air temperature and air humidity 1.5 m above the water surface, using an aspiration psychrometer (Ahlborn FNAD 46, Holzkirchen, Germany);
- −
- Photosynthetically active radiation (PAR) on the water surface, using a quantum sensor (Quantum Meter; Apogee Instruments, Roseville, CA, USA);
- −
- Water temperature at 10 cm depth, using a digital thermometer (HI 98501; Hanna Instruments, Woonsocket, RI, USA);
- −
- Water temperature of the surface, using an infrared thermometer (572-2, Fluke, Washington, DC, USA).
2.4.2. Seasonal Surface Cover
2.4.3. Ecophysiological Measurements In Situ
2.4.4. Biomass Studies
2.5. Data Analyses
3. Results
3.1. Aquatic Plants Alien to Europe Associated with Thermally Abnormal Waters
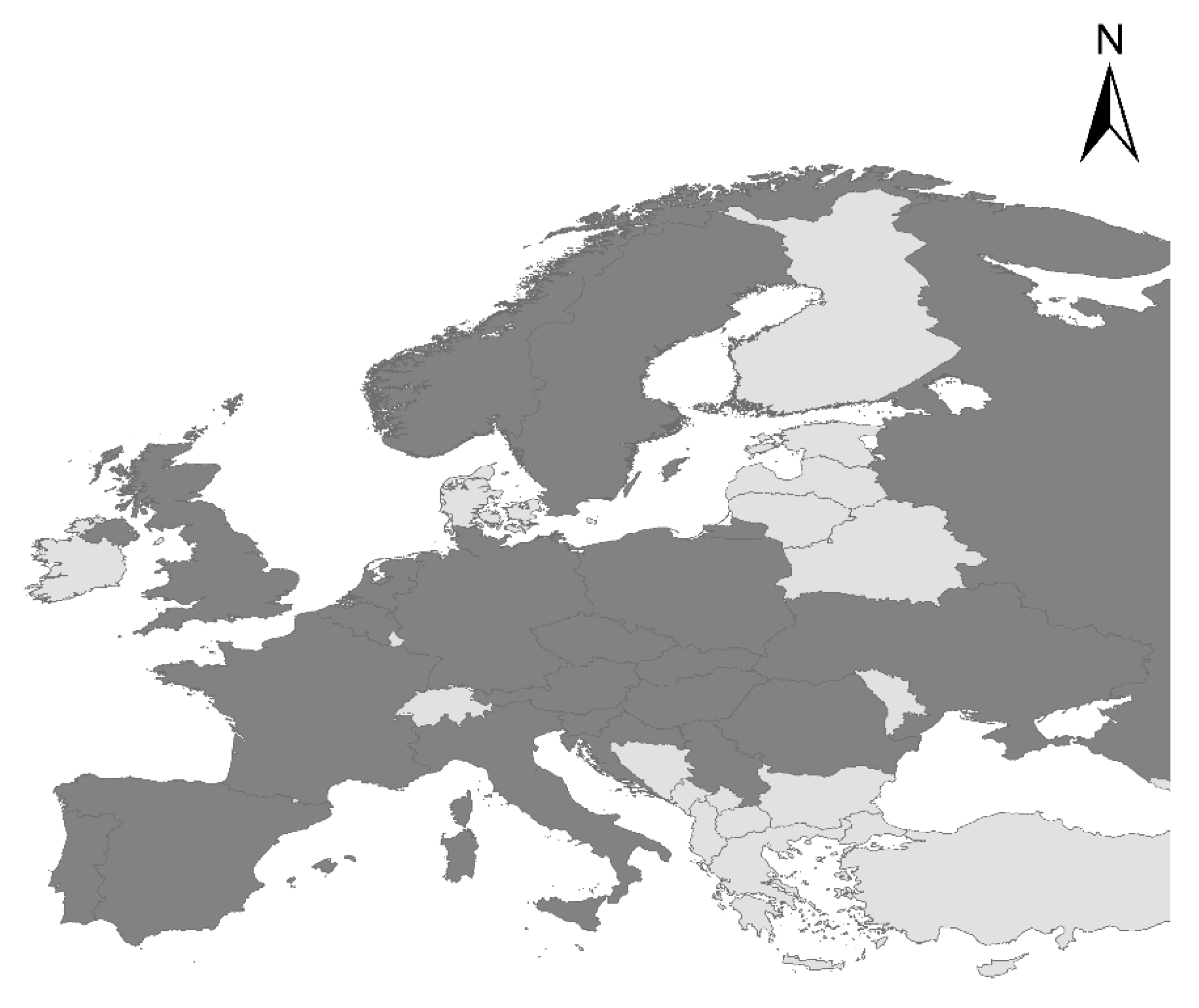
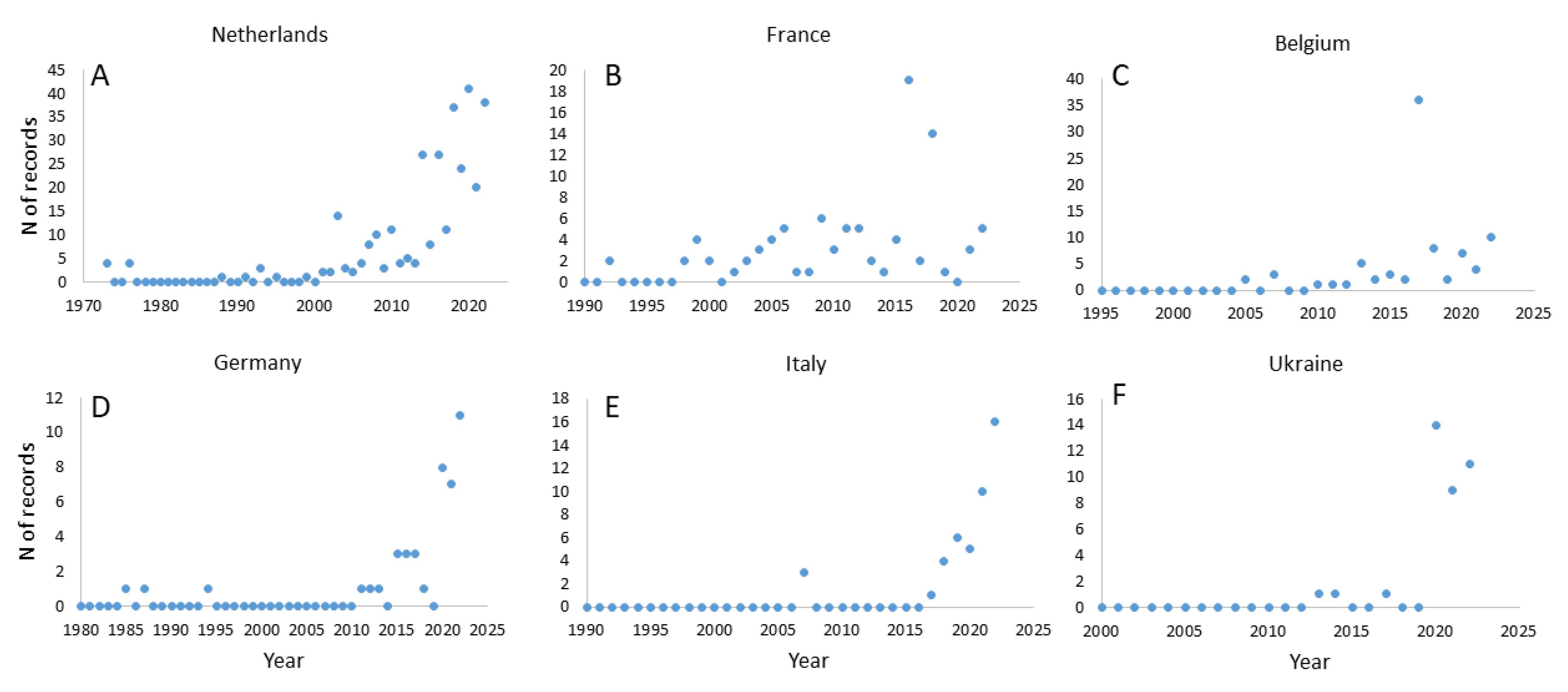
3.2. The Current Distribution Range of Tropical Macrophyte P. stratiotes in Europe
3.3. A Case Study from Thermal Stream Topla
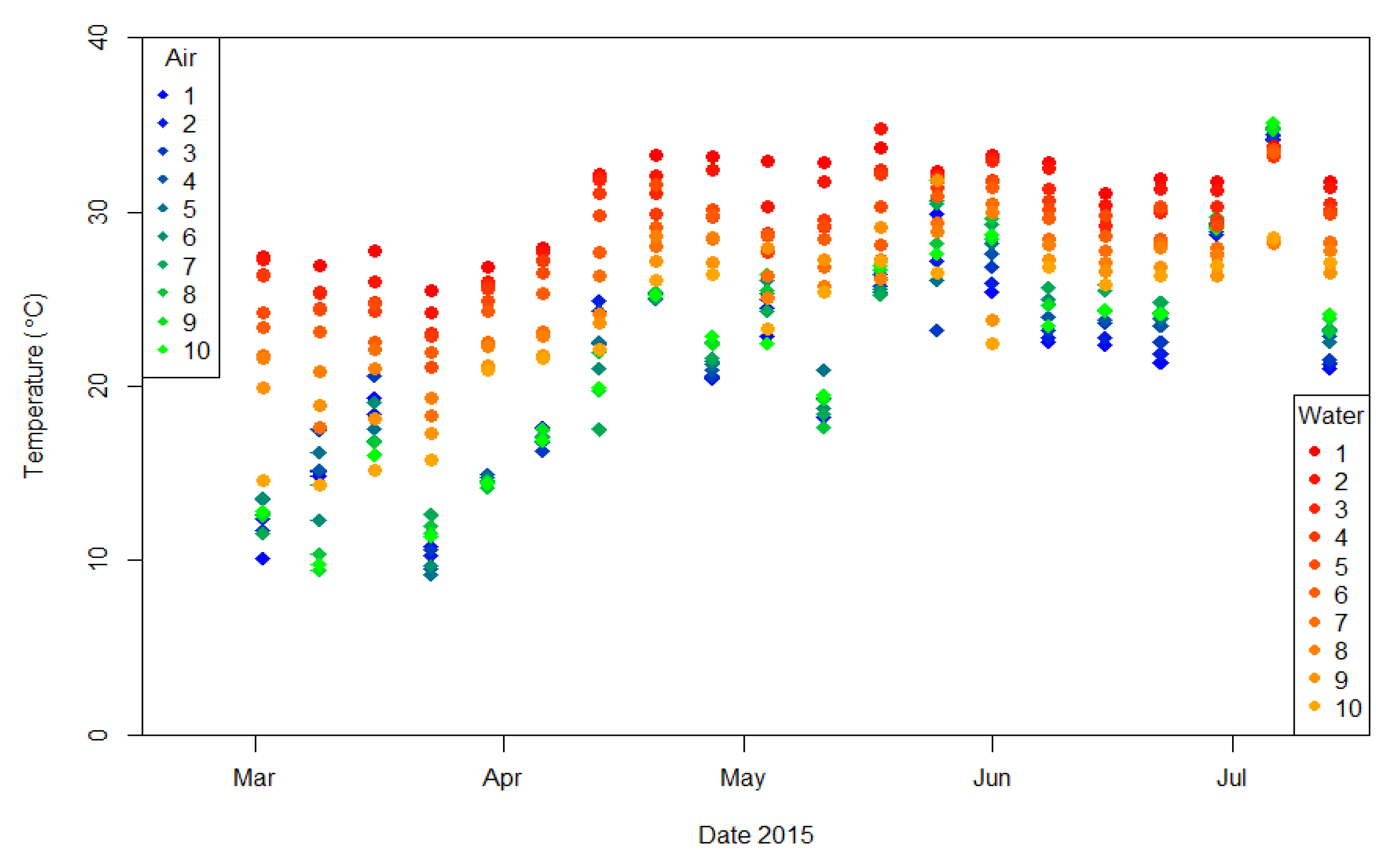
3.3.1. Physical and Chemical Parameters of the Environment
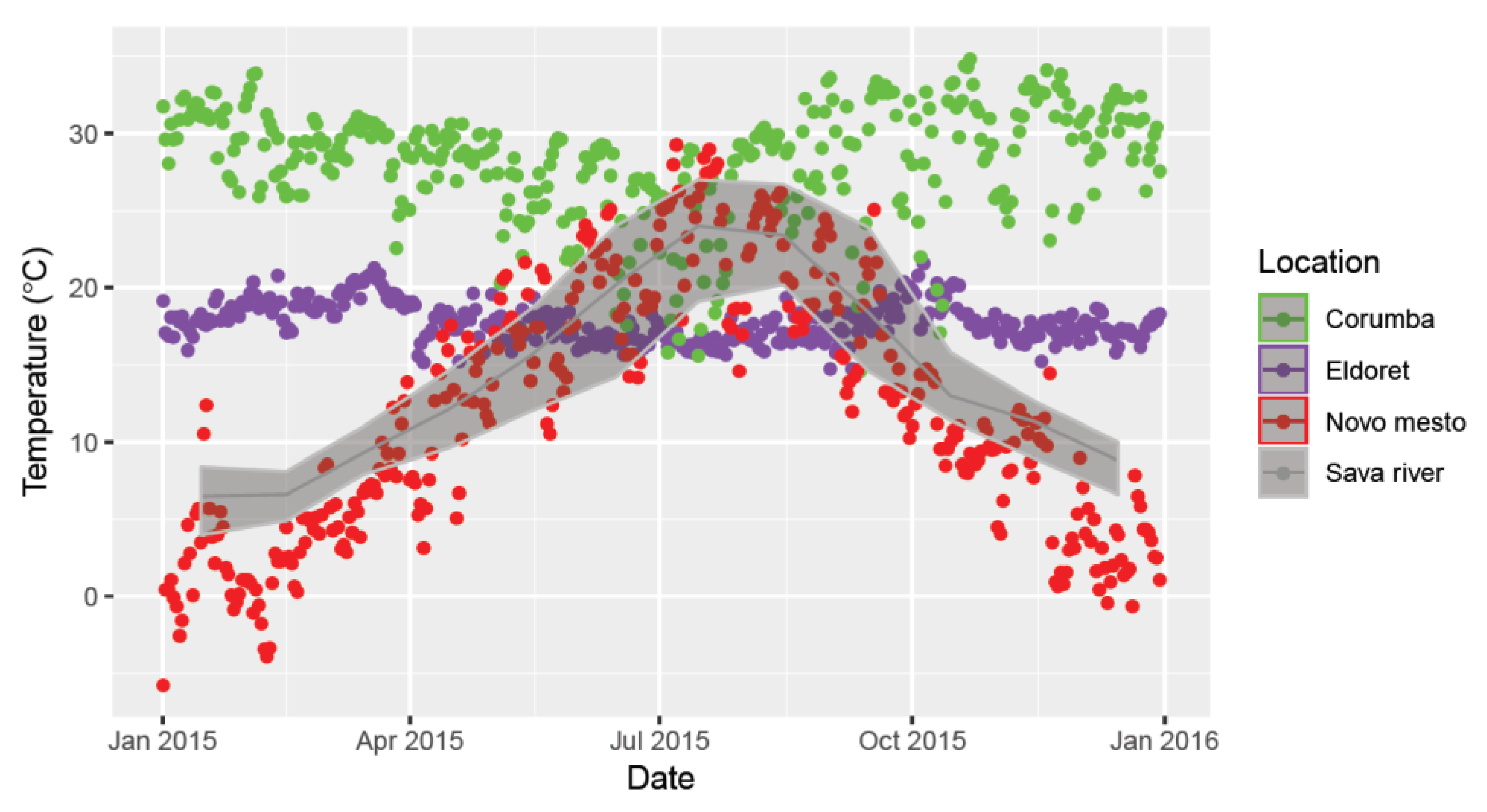
3.3.2. Thermal Conditions
3.3.3. Seasonal Surface Cover
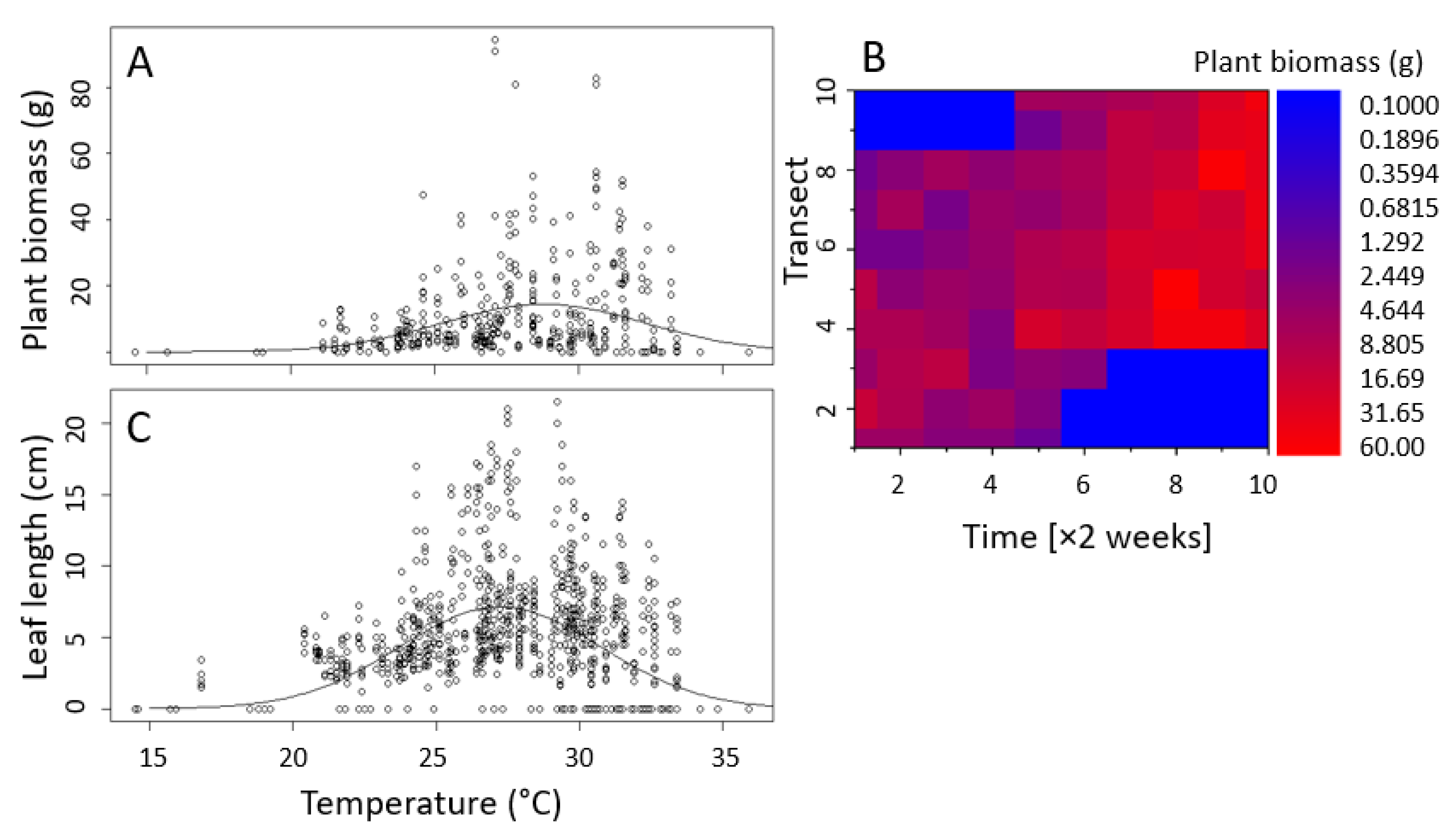
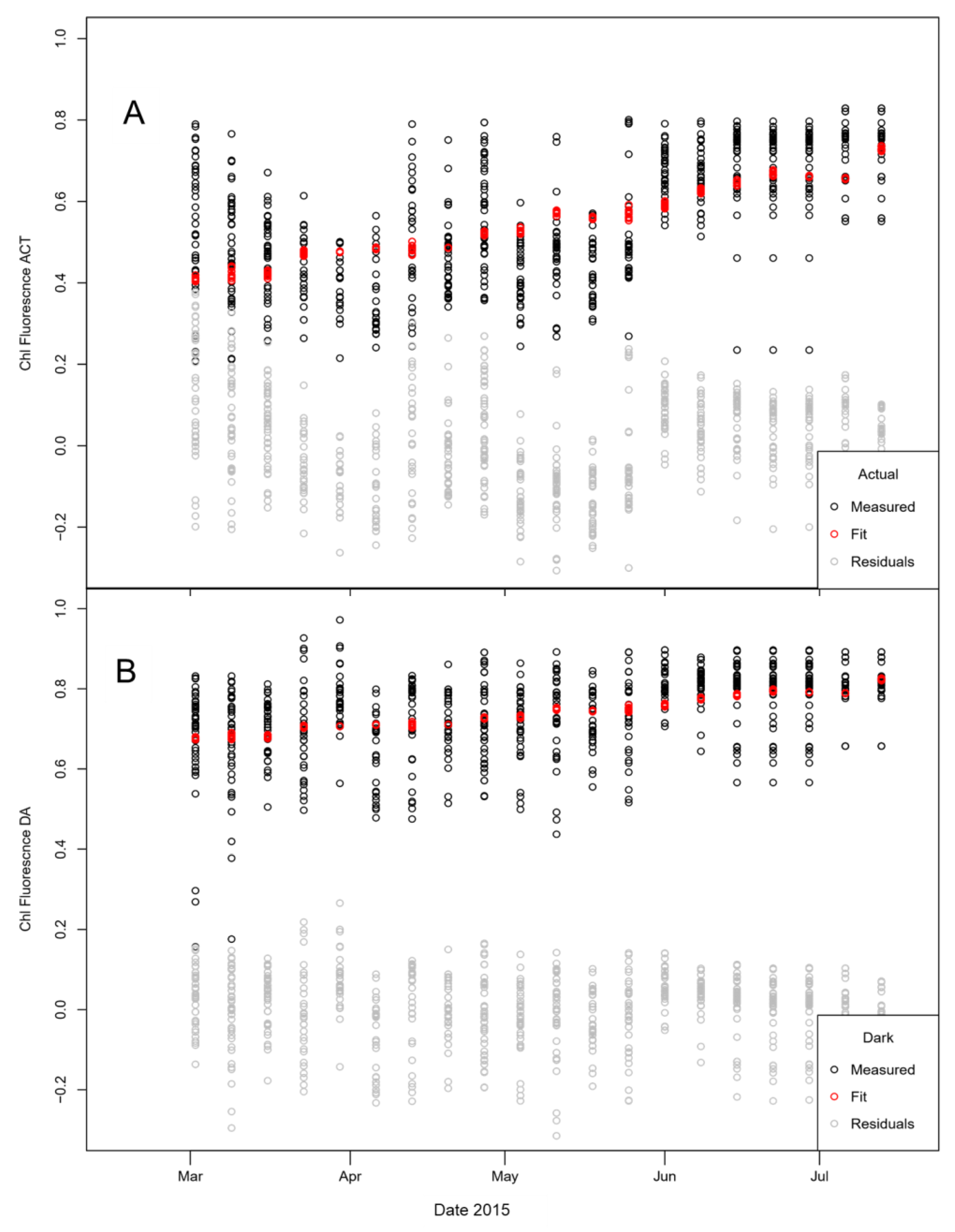
3.3.4. Effect of Temperature on Biomass and Chlorophyll Fluorescence
4. Discussion
4.1. Macrophytes in Thermally Abnormal European Waters
4.2. Pistia Stratiotes Is Expanding Introduced Distribution
4.3. Effect of Temperature on P. stratiotes Performance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Introduced Range (Year of the First Observation) | Native Distribution | Reference |
|---|---|---|---|
| Azolla filiculoides Lam. (Azollaceae); aquarium plant; free-floating fern | Germany (1991, River Erft—TP) Hungary (1940) | North, Central, and South America [9] | [6,29,73] |
| Bacopa caroliniana (Walt.) B.L. Robins (Plantaginaceae); aquarium plant; emersed | Hungary (2005) | Southern parts of North America [74] | [29] |
| Bacopa monnieri (L.) Wettst. (Plantaginaceae); aquarium plant; emersed | Hungary (2005) | Australia, Africa, Asia, South America, and southern parts of North America [74] | [29] |
| Cabomba caroliniana A. Gray (Cabombaceae); aquarium plant; submersed | Hungary (several localities; 1937, Héviz—G) European Russia (1990, backwater of the Moskva River—TP) Austria (2010 (20. cent.), thermal stream Warmbad Villach—D, G) Romania (2014 Peţa Lake—G) | South America [9] | [29,75,76,77,78] |
| Ceratopteris thalictroides (L.) Brongn. (Pteridaceae); aquarium plant; emersed fern | Hungary (1968) Austria (2013, thermal stream Warmbad Villach—D, G) Romania (2014, Peţa Lake—G) | Widespread in tropical regions [9] | [29,77,78] |
| Cryptocoryne crispatula subsp. balansae (Gagnep.) N.Jacobsen (Araceae); aquarium plant; submersed | Austria (2013, thermal stream Warmbad Villach—D, G) | N Vietnam, Thailand [77] | [77] |
| Cryptocoryne wendtii de Wit (Araceae); aquarium plant; submersed | Austria (2013, thermal stream Warmbad Villach—D, G) | Thailand [74] | [77] |
| Egeria densa Planch. (Hydrocharitaceae); aquarium plant; submersed | Germany (1980, River Erft—TP) Slovakia (1993) Hungary (1960) Russia (1983, Pekhorka River—TP) Iceland (2013, pond in Husavik—G; 2004, Opnur—G) Austria (1910, thermal stream Warmbad Villach—D, G) | South America [9] | [6,29,44,73,76,77,79] |
| Eichhornia crassipes (Mart.) Solms (Pontederiaceae); ornamental plant; free-floating; EPPO A2 | Hungary (1950) Germany (2005, River Erft—TP) | South America [9] | [6,29] |
| Eichhornia diversifolia (Vahl) Urb. (Pontederiaceae); aquarium plant; free-floating | Hungary (2005) | South America [74] | [29] |
| Gymnocoronis spilanthoides DC. (Asteraceae); aquarium and aquatic ornamental plant; emersed; EPPO A2 | Hungary (1988, Lake Héviz—G) | South America [74] | [29] |
| Heteranthera zosterifolia Mart. (Pontederiaceae); aquarium plant; submersed | Serbia (2016, Niška Banja—G) Austria (2013, thermal stream Warmbad Villach—D, G) | South America [74] | [77,80] |
| Houttuynia cordata Thumb. (Saururaceae); ornamental plant; emersed | Hungary (2005) | Southeast Asia [74] | [29] |
| Hydrilla verticillata (L. f.) Royle (Hydrocharitaceae); aquarium plant; submersed | Hungary (1980) Slovakia (1995) Austria (1907, thermal stream Warmbad Villach—D, G) | Asia [44] | [29,44,77] |
| Hydrocotyle ranunculoides Lf. (Apiaceae); ornamental pond plant; free-floating; EPPO A2 | Germany (2003, River Erft—TP) Hungary (2005) | North, Central, and South America [9] | [6,29] |
| Hygrophila corymbosa Lindau. (Acanthaceae); aquarium plant; emersed | Hungary (2005) | Southeast Asia [9] | [29] |
| Hygrophila difformis Blume (Acanthaceae); aquarium plant; emersed | Hungary (2005, thermal pond and parts of the Danube near the Lukács Budapest—G) | S Asia and SE Asia [9] | [29,81] |
| Hygrophila polysperma (Roxb.) T. Anderson (Acanthaceae); aquarium plant; emersed | Hungary (1958, thermal pond and parts of the Danube near the Lukács Budapest—G) Germany (2005, River Erft—TP) Poland (2008, lakes of the Konin Valley area—TP) Austria (2005, thermal stream Warmbad Villach—D, G) | India and Malaysia [9] | [6,29,77,81,82] |
| Lagarosiphon major (Ridl.) Moss (Hydrocharitaceae); aquarium plant; submersed | Hungary (2005) Austria (1966, thermal stream Warmbad Villach—D, G) | S Africa [9] | [29,77] |
| Lemna minuta Kunth (Lemnaceae); unintentional introduction; free-floating | Germany (1981, River Erft—TP) Serbia (2016) Russia (2008, Pekhorka River—TP) | North, Central, and South America [9] | [6,73,76,80] |
| Lemna aequinoctialis Welw. (Lemnaceae); unintentional introduction; free-floating | Germany (1982, River Erft—TP) | E Asia, southern hemisphere [74] | [35] |
| Limnophila heterophylla (Roxb.) Benth. (Plantaginaceae); aquarium plant; submersed | Romania (2014, Peţa Lake—G) Hungary (2018, thermal pond and parts of the Danube near the Lukács, Budapest—G) | India, SE China, Philippines [74] | [78,81] |
| Limnophila sessiliflora Blume (Plantaginaceae); aquarium plant; submersed | Hungary (1940) Slovakia (1993, Bojnice) | Asia [44] | [29,44] |
| Ludwigia sp.—hybrid (Onagraceae); aquarium plant; emersed | Hungary (2018, thermal pond and parts of the Danube near the Lukács, Budapest—G) | / | [81] |
| Ludwigia alternifolia L. (Onagraceae); ornamental pond plant; emersed | Hungary (1940) | E North America [74] | [29] |
| Ludwigia grandiflora (Michx.) Greuter & Burdet (Onagraceae); ornamental pond plant; emersed; EPPO A2 | Hungary (2005) | South America, parts of North America [74] | [29] |
| Ludwigia repens J.R. Forst. (Onagraceae); aquarium plant; emersed | Hungary (1924) Serbia (2011, Niška Banja—G) Slovakia (2017) | North and Central America [74] | [29,44,83] |
| Mimulus guttatus Fisch. ex DC. (Scrophulariaceae); ornamental pond plant; emersed | Hungary (1994) | North America [74] | [29] |
| Monochoria korsakowii Regel & Maack (Pontederiaceae); ornamental pond plant; emersed | Hungary (1988) | Ukraine, Caucasus, India, East Asia [74] | [29] |
| Myriophyllum aquaticum (Vell.) Verdc. (Haloragaceae); aquarium plant; submersed | Hungary (1968) Germany (2003, River Erft—TP) Romania (2014, Peţa Lake—G) Austria (1988, thermal stream Warmbad Villach—G, D) | South America [74] | [6,29,73,77,78] |
| Myriophyllum heterophyllum Michx. (Haloragaceae); aquarium plant, ornamental pond plant; submersed; EPPO A2 | Hungary (2006) | E-North America, Central America [74] | [29] |
| Myriophyllum tuberculatum Roxb. (Haloragaceae); aquarium plant; submersed | Hungary (2018, thermal pond and parts of the Danube near the Lukács, Budapest—G) | India to China, N Peninsula Malaysia [74] | [81] |
| Najas gracillima (A. Braun ex Engelm.) Magnus (Hydrocharitaceae); unintentional introduction; submersed | Hungary (2012) | Australasia, China, Eastern Asia, North America [74] | [29] |
| Najas guadalupensis (Spreng.) Magnus (Hydrocharitaceae); aquarium plant; submersed | Slovakia (1986) Hungary (2005) Romania (2014, Peţa Lake—G) | America [74] | [29,44,78] |
| Nelumbo nucifera Gaertn. (Nelumbonaceae); ornamental pond plant; leaf floating | Hungary (1955) | Ukraine to north Iran, Russian Far East to Tropical Asia, Australia [74] | [29] |
| Nuphar advena (Aiton) W.T. Aiton (Nymphaeaceae); ornamental pond plant; leaf floating | Hungary (1920) | North America [74] | [29] |
| Nymphaea “Blue Bird” (N. micrantha x N. capensis) (Nymphaeaceae); ornamental pond plant; leaf floating | Hungary (1900) | / | [29] |
| Nymphaea lotus var. thermalis L. (Nymphaeaceae); ornamental pond plant; leaf floating | Hungary (1842) | Endemic to the thermal water of the Peţa River in Romania [84] | [29] |
| Nymphaea nouchali var. caerulea (Savigny) Verdc. (Nymphaeaceae); ornamental pond plant; leaf floating | Hungary (1891, thermal pond and parts of the Danube near the Lukács, Budapest—G) | E Africa [74] | [29,81] |
| Nymphaea rubra Roxb. ex Andrews (Nymphaeaceae); ornamental pond plant; leaf floating | Hungary (1891) | Tropical Asia [74] | [29] |
| Pistia stratiotes L. (Araceae); aquarium plant, ornamental pond plant; free-floating; EPPO A2 | Hungary (1966) Germany (1981, River Erft—TP) Russia (1989, Volga—TP; 1998 Pekhorka River—TP) Serbia (1994, a thermal spring “Banjica”—G; 2005, Rgoška Banja spa—G) Slovenia (2001, stream Topla—G, D) Slovakia (2007) Ukraine (2011, Seversky Donets—TP) | Pantropical distribution, probably originating from South America [17] | [6,11,29,35,42,44,47,76,80] |
| Pontederia cordata L. (Pontederiaceae); ornamental pond plant; emersed | Hungary (2005) | North America, South America [74] | [29] |
| Rotala rotundifolia (Buch.-Ham. ex Roxb.) Koehne (Lythraceae); ornamental pond plant; emersed | Hungary (1998) Serbia (2016, Niška Banja—G) | Tropical Asia [74] | [29,80] |
| Sagittaria latifolia Willd. (Alismataceae); aquarium plant; emersed | Austria (1951, thermal stream Warmbad Villach—D, G | North America, Central America, N South America [74] | [77] |
| Sagittaria platyphylla (Engelm.) J.G. Sm. (Alismataceae); aquarium plant, ornamental pond plant; emersed | Russia (2002, Pekhorka River—TP) | S North America, Central America [74] | [76] |
| Sagittaria subulata (L.) Buchenau (Alismataceae); aquarium plant; submersed | Hungary (1965) Slovakia (1995) Germany (1984, Warme Wuhle—TP) | SE North America, N South America [74] | [29,44,85] |
| Salvinia auriculata Aubl. (Salviniaceae); aquarium plant; free-floating fern | Hungary (1964) | Central and South America [74] | [29] |
| Saururus cernuus L. (Saururaceae); ornamental plant; emersed | Hungary (2005) | E North America [74] | [29] |
| Shinnersia rivularis (A.Gray) R.M.King & H.Rob. (Asteraceae); aquarium plant; emersed | Germany (1992, River Erft—TPm) Slovakia (1998, 2002, a waste canal discharging thermal water from the bath house “Kalinka”, Bojnice—D, G) Hungary (1998, a thermal lake Hévíz—G) Austria (2000, thermal streams in Warmbad Villach—G) | Central America [9] | [29,44,86] |
| Utricularia gibba L. (Lentibulariaceae); aquarium plant; free-floating | Hungary (1936) Slovakia (1993) | America, Africa, Asia [74] | [29,44] |
| Vallisneria americana Michx. (Hydrocharitaceae); aquarium plant; submersed | Russia (2010, Pekhorka River—TP) | North America, Central America, N South America [74] | [76] |
| Vallisneria gigantea Graebn. (Hydrocharitaceae); aquarium plant; submersed | Hungary (1891) | SE Asia, Australia [74] | [29] |
| Vallisneria spiralis L. (Hydrocharitaceae); aquarium plant; submersed | Hungary (1808) Russia (1999, Desnogorsk Reservoir—TP; 1972, Belovskoe Reservoir—TP) Germany (2003, River Erft—TP; 2017, Reden—TP) Serbia (2011, Niška Banja—G) Poland (1993, Konin Lakes—TP) Iceland (2013, pond Husavik—G) Austria (1880, thermal streams in Warmbad Villach—D, G) | N Africa, Asia, S Europe [9] | [6,29,73,77,79,83,87,88,89,90,91] |
| Victoria amazonica Sowerby (Nymphaeaceae); ornamental pond plant; leaf floating | Slovakia (1998) | South America [74] | [44] |
| Wolffia globosa (Roxb.) Hartog & Plas (Araceae); unintentional introduction; free-floating | Russia (2002, Pekhorka River—TP) | Pakistan to Japan, Malaysia [74] | [76] |
References
- Strayer, D.L.; Dudgeon, D. Freshwater biodiversity conservation: Recent progress and future challenges. J. N. Am. Benthol. Soc. 2010, 29, 344–358. [Google Scholar] [CrossRef] [Green Version]
- Grzybowski, M.; Glińska-Lewczuk, K. Principal threats to the conservation of freshwater habitats in the continental biogeographical region of Central Europe. Biodivers. Conserv. 2019, 28, 4065–4097. [Google Scholar] [CrossRef] [Green Version]
- Díaz, S.; Settele, J.; Brondízio, E.; Ngo, H.; Guèze, M.; Agard, J.; Arneth, A.; Balvanera, P.; Brauman, K.; Butchart, S.; et al. Summary for Policymakers of the Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services. Project Report. Available online: https://uwe-repository.worktribe.com/output/1493508 (accessed on 5 January 2023).
- Vilà, M.; Basnou, C.; Pyšek, P.; Josefsson, M.; Genovesi, P.; Gollasch, S.; Nentwig, W.; Olenin, S.; Roques, A.; Roy, D.; et al. How well do we understand the impacts of alien species on ecosystem services? A pan-European, cross-taxa assessment. Front. Ecol. Environ. 2010, 8, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Hulme, P.E. Biological invasions in Europe: Drivers, pressures, states, impacts and responses. In Biodiversity under Threat; Hester, R., Harrison, R.M., Eds.; Cambridge University Press: Cambridge, UK, 2007; ISBN 978-0-85404-251-7. [Google Scholar]
- Hussner, A.; Heidbuechel, P.; Heiligtag, S. Vegetative overwintering and viable seed production explain the establishment of invasive Pistia stratiotes in the thermally abnormal Erft River (North Rhine-Westphalia, Germany). Aquat. Bot. 2014, 119, 28–32. [Google Scholar] [CrossRef]
- Jaklič, M.; Koren, Š.; Jogan, N. Alien water lettuce (Pistia stratiotes L.) outcompeted native macrophytes and altered the ecological conditions of a Sava oxbow lake (SE Slovenia). Acta Bot. Croat. 2020, 79, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Hussner, A.; Nehring, S.; Hilt, S. From first reports to successful control: A plea for improved management of alien aquatic plant species in Germany. Hydrobiologia 2013, 737, 321–331. [Google Scholar] [CrossRef]
- Hussner, A. Alien aquatic plant species in European countries. Weed Res. 2012, 52, 297–306. [Google Scholar] [CrossRef]
- Boršić, I.; Rubinić, T. First record of Pistia stratiotes L. (Araceae) in Croatia, with the consideration of possible introduction pathways. Period. Biol. 2021, 123, 35–39. [Google Scholar] [CrossRef]
- Šajna, N.; Haler, M.; Škornik, S.; Kaligarič, M. Survival and expansion of Pistia stratiotes L. in a thermal stream in Slovenia. Aquat. Bot. 2007, 87, 75–79. [Google Scholar] [CrossRef]
- Wilk-Woźniak, E.; Najberek, K. Towards clarifying the presence of alien algae in inland waters: Can we predict places of their occurrence? Biologia 2013, 68, 838–844. [Google Scholar] [CrossRef]
- Hussner, A.; van Dam, H.; Vermaat, J.E.; Hilt, S. Comparison of native and neophytic aquatic macrophyte developments in a geothermally warmed river and thermally normal channels. Fundam. Appl. Limnol. 2014, 185, 155–166. [Google Scholar] [CrossRef]
- Piria, M.; Radočaj, T.; Vilizzi, L.; Britvec, M. Climate change may exacerbate the risk of invasiveness of non-native aquatic plants: The case of the Pannonian and Mediterranean regions of Croatia. In Recent advancements in the Risk Screening of Freshwater and Terrestrial Non-Native Species; Giannetto, D., Piria, M., Tarkan, A.S., Zięba, G., Eds.; NeoBiota: Sofia, Bulgaria, 2022; pp. 25–52. [Google Scholar] [CrossRef]
- Ormerod, S.J.; Dobson, M.; Hildrew, A.G.; Townsend, C.R. Multiple stressors in freshwater ecosystems. Freshw. Biol. 2010, 55, 1–4. [Google Scholar] [CrossRef]
- Brundu, G.; Stinca, A.; Angius, L.; Bonanomi, G.; Celesti-Grapow, L.; D’Auria, G.; Griffo, R.; Migliozzi, A.; Motti, R.; Spigno, P. Pistia stratiotes L. and Eichhornia crassipes (Mart.) Solms.: Emerging invasive alien hydrophytes in Campania and Sardinia (Italy). Bull. OEPP 2012, 42, 568–579. [Google Scholar] [CrossRef]
- Neuenschwander, P.; Julien, M.H.; Center, T.D.; Hill, M.P. Pistia stratiotes L. (Araceae). In Biological Control of Tropical Weeds Using Arthropods; Muniappan, R., Reddy, G.V.P., Raman, A., Eds.; Cambridge University Press: London, UK, 2009; pp. 332–352. ISBN 978-0-521-87791-6. [Google Scholar]
- Cilliers, C.J. Biological control of water lettuce, Pistia stratiotes (Araceae), in South Africa. Agric. Ecosyst. Environ. 1991, 37, 225–229. [Google Scholar] [CrossRef]
- EPPO. Pistia stratiotes L. Datasheets on pests recommended for regulation. EPPO Bull. 2017, 47, 537–543. [CrossRef] [Green Version]
- Van Dijk, G. Vallisneria and its interactions with other species. Aquatic 1985, 7, 6–10. [Google Scholar]
- Simberloff, D.; Schmitz, D.C.; Brown, T.C. Strangers in Paradise: Impact and Management of Nonindigenous Species in Florida; Island Press: Washington, DC, USA, 1997. [Google Scholar]
- Labrada, R.; Fornasari, L. Management of the Worst Aquatic Weeds in Africa. FAO Efforts and Achievements during the Period 1991–2001; FAO: Rome, Italy, 2002. [Google Scholar]
- ARSO 2023. ARHIV—Opazovani in Merjeni Meteorološki Podatki po Sloveniji. Available online: https://meteo.arso.gov.si/met/sl/archive/ (accessed on 11 January 2023).
- Records ICAO/OACI: SBCR. Available online: https://en.tutiempo.net/records/sbcr (accessed on 7 November 2018).
- Records ICAO/OACI: HKEL. Available online: https://en.tutiempo.net/records/hkel (accessed on 7 November 2018).
- ESRI. ArcView GIS 3.2.; Environmental Systems Research Institute, Inc.: Redlands, CA, USA, 1999. [Google Scholar]
- Roháček, K.; Soukupová, J.; Barták, M. Chlorophyll fluorescence: A wonderful tool to study plant physiology and plant stress. Res. Signpost. 2008, 37, 41–104. [Google Scholar]
- EPPO A2 List of Pests Recommended for Regulation as Quarantine Pests. Version 2022-09. Available online: https://www.eppo.int/ACTIVITIES/plant_quarantine/A2_list (accessed on 11 January 2023).
- Lukács, B.A.; Mesterházy, A.; Vidéki, R.; Király, G. Alien aquatic vascular plants in Hungary (Pannonian ecoregion): Historical aspects, data set and trends. Plant Biosyst. 2016, 150, 388–395. [Google Scholar] [CrossRef] [Green Version]
- García Murillo, P.; Dana Sanchez, E.D.; Rodrigez Hiraldo, C. Pistia stratiotes L. (Araceae) una planta acuatica en las proximidades del parquet nacional de Doñana (SW Espana). Acta Bot. Malacit. 2005, 30, 235–236. [Google Scholar] [CrossRef]
- Pall, K.; Mayerhofer, V.; Mayerhofer, S. Aquatische Neobiota in Österreich-Stand 2013; BMLFUW: Wien, Austria, 2013; Available online: https://info.bml.gv.at/service/publikationen/wasser/Aquatische-Neobiota-in--sterreich---Stand-2013.html (accessed on 11 January 2023).
- Verloove, F. Catalogue of Neophytes in Belgium (1800–2005); Scripta Botanica Belgica; National Botanic Garden of Belgium: Meise, Belgium, 2006; Volume 39. [Google Scholar]
- Kaplan, Z.; Danihelka, J.; Lepsi, M.; Lepší, P.; Ekrt, L.; Chrtek, J.; Kocián, J.; Prančl, J.; Kobrlová, L.; Hroneš, M.; et al. Distributions of vascular plants in the Czech Republic. Part 3. Preslia 2016, 88, 459–544. [Google Scholar]
- SI Observation Flore. Pistia stratiotes L. Available online: https://siflore.fcbn.fr/?cd_ref=447733&r=metro (accessed on 11 January 2023).
- Diekjobst, H. Pistia stratiotes L. und Lemna aequinoctialis Welwitsch vorübergehend im Gebiet der unteren Erft. Gött. Flor. Rundbr. 1984, 18, 90–95. [Google Scholar]
- D`Auria, G.; Zavagno, F. Indagine sui “Bodri” della Provincia di Cremona. Monografie di «Pianura» N. 3 -1999; Provincia di Cremona: Cremona, Italy, 1999. [Google Scholar]
- Mennema, J. Is water lettuce (Pistia stratiotes L.) becoming a new aquatic weed in the Netherlands? Natura 1977, 74, 187–190. [Google Scholar]
- Elven, R.; Hegre, H.; Solstad, H.; Pedersen, O.; Pedersen, P.A.; Åsen, P.A.; Bjureke, K.; Vandvik, V. Pistia stratiotes, Vurdering av Økologisk Risiko. Artsdatabanken. Available online: https://artsdatabanken.no/Fab2018/N/1794 (accessed on 15 November 2018).
- Krajewski, Ł. Pistia rozetkowa—Efemerofit flory regionu. Przyr. Górnego Śląska 2013, 72, 13. [Google Scholar]
- Almeida, J.D.; Freitas, H. Exotic naturalized flora of continental Portugal—A reassessment. Bot. Complut. 2006, 30, 117–130. [Google Scholar]
- Lansdown, R.V.; Anastasiu, P.; Barina, Z.; Bazos, I.; Çakan, H.; Caković, D.; Delipetrou, P.; Matevski, V.; Mitić, B.; Ruprecht, E.; et al. Review of Alien Freshwater Vascular Plants in South-east Europe. In ESENIAS Scientific Reports 1. State of the Art of Alien Species in South-Eastern Europe; Rat, M., Trichkova, T., Scalera, R., Tomov, R., Uludag, A., Eds.; University of Novi Sad Faculty of Sciences: Novi Sad, Serbia, 2016; pp. 137–154. ISBN 978-86-7031-3316. [Google Scholar]
- Pilipenko, V.N. The tropical species Pistia stratiotes (Araceae) in the delta of Volga river. Bot. Zhurnal 1993, 78, 119–120. [Google Scholar]
- Ranđelović, V.; Zlatković, B.; Jović, D. Tropska biljna vrsta Pistia stratiotes L. (Araceae) u flori Srbije. In II Simpozijum o flori Srbije, IV Simpozijum o flori Jugoistočne Srbije; Zbornik rezimea: Vranje, Srbia, 1995; pp. 33–34. [Google Scholar]
- Hrivnák, R.; Medvecká, J.; Baláži, P.; Bubíková, K.; Otahelová, H.; Svitok, M. Alien aquatic plants in Slovakia over 130 years: Historical overview, current distribution and future perspectives. NeoBiota 2019, 49, 37–56. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.; Coulson, S. Artportalen (Swedish Species Observation System). Version 92.144. ArtDatabanken. Occurrence Dataset. 2019. Available online: https://www.gbif.org/occurrence/1835762190 (accessed on 12 January 2023).
- Newman, J. Rapid Risk Assessment Summary Sheet: Water Lettuce (Pistia stratiotes). Available online: https://www.nonnativespecies.org/assets/Uploads/RSS_RA_Pistia_stratiotes.pdf (accessed on 12 January 2023).
- Dikiy, N.P.; Dovbnya, A.N.; Lyashko Yu, V.; Medvedev, D.V.; Medvedeva, E.P.; Botova, M.A.; Khlapova, N.P.; Fedorets, I.D. Radionuclide Biosorption by the Aquatic Plants of Pistia stratiotes. Probl. Act. Sci. Technol. 2014, 5, 50–53. [Google Scholar]
- Hussner, A.; van de Weyer, K.; Gross, E.M.; Hilt, S. Comments on increasing number and abundance of non indigenous aquatic macrophyte species in Germany. Weed Res. 2010, 50, 519–526. [Google Scholar] [CrossRef]
- Peres, C.K.; Lambrecht, R.W.; Tavares, D.A.; Chiba de Castro, W.A. Alien Express: The threat of aquarium e-commerce introducing invasive aquatic plants in Brazil. Perspect. Ecol. Conserv. 2018, 16, 221–227. [Google Scholar] [CrossRef]
- Zelnik, I.; Haler, M.; Gaberščik, A. Vulnerability of a riparian zone towards invasion by alien plants depends on its structure. Biologia 2015, 70, 869–878. [Google Scholar] [CrossRef]
- Feuchtmayr, H.; Moran, R.; Hatton, K.; Connor, L.; Heyes, T.; Moss, B.; Harvey, I.; Atkinson, D. Global warming and eutrophication: Effects on water chemistry and autotrophic communities in experimental hypertrophic shallow lake mesocosms. J. Appl. Ecol. 2009, 46, 713–723. [Google Scholar] [CrossRef]
- Netten, J.C.; Arts, G.H.P.; Gylstra, R.; Van Nes, E.H.; Scheffer, M.; Roijackers, R.M.M. Effect of temperature and nutrients on the competition between free-floating Salvinia natans and submerged Elodea nuttallii in mesocosms. Fund. Appl. Limnol. 2010, 177, 125–132. [Google Scholar] [CrossRef]
- Adebayo, A.A.; Briski, E.; Briski, E.; Kalaci, O.; Hernandez, M.; Ghabooli, S.; Beric, B.; Chan, F.T.; Zhan, A.; Fifield, E.; et al. Water hyacinth (Eichhornia crassipes) and water lettuce (Pistia stratiotes) in the Great Lakes: Playing with fire? Aquat. Invasions 2011, 6, 91–96. [Google Scholar] [CrossRef] [Green Version]
- Simberloff, D.; Von Holle, B. Positive interactions of nonindigenous species: Invasional meltdown? Biol. Invasions 1999, 1, 21–32. [Google Scholar] [CrossRef]
- Pieterse, A.H.; Lange, L.D.; Verhagen, L. A study on certain aspects of seed germination and growth of Pistia stratiotes L. Acta Bot. Neerl. 1981, 30, 47–57. [Google Scholar] [CrossRef]
- Venema, P. Fast spread of water lettuce (Pistia stratiotes L.) around Meppe. Gorteria 2001, 27, 133–135. [Google Scholar]
- FLORON Verspreidingsatlas Vaatplanten: Pistia stratiotes L. Available online: https://www.verspreidingsatlas.nl/5269# (accessed on 15 January 2023).
- Parker, S. Reports of Meetings. Somerset Rare Plant Group Newsletter 2010, 11. Available online: http://www.somersetrareplantsgroup.org.uk/wp-content/uploads/2014/11/2010-Newsletter-11.pdf (accessed on 12 January 2023).
- Thiébaut, G.; Dutartre, A. Management of invasive aquatic plants in France. In Aquatic Ecosystem Research Trends; Nairne, G.H., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2009. [Google Scholar]
- Ercolini, P. Pistia stratiotes L. (Alismatales: Araceae) in Versilia (Toscana nordoccidantale). Biol. Ambient. 2008, 22, 45–49. [Google Scholar]
- Stinca, A.; D’Auria, G.; Motti, R. Sullo status invasivo di Bidens bipinnata, Phoenix canariensis, Pistia stratiotes e Tradescantia fluminensis in Campania (Sud Italia). Inf. Bot. Ital. 2012, 44, 295–299. [Google Scholar]
- Ružičková, J.; Lehotská, B.; Takáčová, A.; Semerád, M. Morphometry of alien species Pistia stratiotes L. in natural conditions of the Slovak Republic. Biologia 2020, 75, 1–10. [Google Scholar] [CrossRef]
- Shapovalov, M.I.; Saprykin, M.A. Alien Species Pistia Stratiotes L. (Araceae) in Water Bodies of Urbanized Territories of Southern Russia. Russ. J. Biol. Invasions 2016, 7, 195–199. [Google Scholar] [CrossRef]
- Živković, M.M.; Anđelković, A.A.; Cvijanović, D.L.; Novković, M.Z.; Vukov, D.M.; Šipoš, Š.Š.; Ilić, M.M.; Pankov, N.P.; Miljanović, B.M.; Marisavljević, D.P.; et al. The beginnings of Pistia stratiotes L. invasion in the lower Danube delta: The first record for the Province of Vojvodina (Serbia). BioInvasions Rec. 2019, 8, 218–229. [Google Scholar] [CrossRef]
- Hussner, A.; Heiligtag, S. Pistia stratiotes L. (Araceae), die Muschelblume, im Gebiet der unteren Erft (Nordrhein-Westfalen): Ausbreitungstendenz und Problempotenzial. Veröffentlichungen Des. Boch. Bot. Ver. 2013, 5, 1–6. [Google Scholar]
- Hussner, A.; Heidbüchel, P.; Coetzee, J.; Gross, E.M. From introduction to nuisance growth: A review of traits of alien aquatic plants which contribute to their invasiveness. Hydrobiologia 2021, 848, 2119–2151. [Google Scholar] [CrossRef]
- Tamada, K.; Itoh, K.; Uchida, Y.; Higuchi, S.; Sasayama, D.; Azuma, T. Water lettuce overwinter with heat waste. Weed Biol. Manag. 2012, 15, 20–26. [Google Scholar] [CrossRef]
- Hoveka, L.N.; Bezeng, B.S.; Yessoufou, K.; Boatwright, J.S.; Van der Bank, M. Effects of climate change on the future distributions of the top five freshwater invasive plants in South Africa. S. Afr. J. Bot. 2016, 102, 33–38. [Google Scholar] [CrossRef]
- Cancian, L.F.; Camargo, A.F.M.; Silva, G.H.G. Crescimento de Pistia stratiotes em diferentes condiçõesde temperatura e fotoperíodo. Acta Bot. Bras. 2009, 23, 552–557. [Google Scholar] [CrossRef] [Green Version]
- van der Heide, T.; Roijackers, R.M.; van Nes, E.H.; Peeters, E.T.H.M. A simple equation for describing the temperature dependent growth of free-floating macrophytes. Aquat. Bot. 2006, 84, 171–175. [Google Scholar] [CrossRef]
- Whiteman, J.B.; Room, P.M. Temperatures lethal to Salvinia molesta Mitchell. Aquat. Bot. 1991, 40, 27–35. [Google Scholar] [CrossRef]
- Desotgiu, R.; Pollastrini, M.; Cascio, C.; Gerosa, G.; Marzuoli, R.; Bussotti, F. Chlorophyll a fluorescence analysis along a vertical gradient of the crown in a poplar (Oxford clone) subjected to ozone and water stress. Tree Physiol. 2012, 32, 976–986. [Google Scholar] [CrossRef] [Green Version]
- Hussner, A.; Lösch, R. Alien aquatic plants in a thermally abnormal river and their assembly to neophyte-dominated macrophyte stands (River Erft, Northrhine-Westphalia). Limnologica 2005, 35, 18–30. [Google Scholar] [CrossRef] [Green Version]
- Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Available online: http://www.plantsoftheworldonline.org/ (accessed on 9 January 2023).
- Király, G.; Steták, D.; Bányász, A. Spread of invasive macrophytes in Hungary. In Biological Invasions—From Ecology to Conservation; Rabitsch, W., Essl, F., Klingenstein, F., Eds.; NeoBiota: Sofia, Bulgaria, 2008; pp. 123–130. [Google Scholar]
- Shcherbakov, A.V.; Ershkova, E.V.; Khapugin, A.A.; Lyubeznova, N.V. Alien Aquatic Plant Species in European Russia. Bot. Zh. 2022, 107, 70–80. [Google Scholar] [CrossRef]
- Ofenböck, G. Aquatische Neobiota in Österreich; Bundesministerium für Land—Und Forstwirtschaft, Umwelt und Wasserwirtschaft: Wien, Österreich, 2018; ISBN 978-3-89432-287-8. [Google Scholar]
- Bunea, G.; Oroian, I.G.; Todea, A. Law helps, but shows up rather late: The case of Biharean thermal endemic species. Poeciliid Res. 2014, 4, 13–18. [Google Scholar]
- Wasowicz, P.; Przedpelska-Wasowicz, E.M.; Guðmundsdóttir, L.; Tamayo, M. Vallisneria spiralis and Egeria densa (Hydrocharitaceae) in arctic and subarctic Iceland. New J. Botany 2014, 4, 85–89. [Google Scholar] [CrossRef]
- Zlatković, B.K.; Bogosavljević, S.S. Risk analysis of alien plants recorded in thermal waters of Serbia. Weed Res. 2020, 60, 85–95. [Google Scholar] [CrossRef]
- Kempkes, M.; Lukas, J.; Bierbach, D. Tropische Neozoen in heimischen Fließgewässern Guppys und andere Exoten in Gillbach und Erft—Ursachen, Folgen, Perspektiven; Wolf, VerlagsKG: Magdeburg, Germany, 2018; ISBN 978-3-89432-287-8. [Google Scholar]
- Gabka, M.; Owsianny, P. First records of the Hygrophila polysperma Roxb T. Anderson (Acanthaceae) in Poland. Bot. Steciana 2009, 13, 9–14. [Google Scholar]
- Milenković, M.; Žikić, V.; Stanković, S.S.; Marić, S. First study of the guppy fish (Poecilia reticulate Peters,) occurring in natural thermal waters of Serbia. J. Appl. Ichthyol. 2014, 30, 160–163. [Google Scholar] [CrossRef]
- Veler, A. Nymphaea lotus up North, Naturally. Water Gardeners International 2008, 3. Available online: http://www.watergardenersinternational.org/journal/3-4/ana/page1.html (accessed on 4 January 2023).
- Mühlberg, H. Sagittaria subulata im Stadtgebiet von Berlin. Schlechtendalia 2000, 5, 27–30. [Google Scholar]
- Eliáš, P.; Hájek, M.; Hájková, P. A European warm waters neophyte Shinnersia rivularis—New alien species to the Slovak flora. Biologia 2009, 64, 684–686. [Google Scholar] [CrossRef]
- Katsman, E.A.; Kuchkina, M.A. Introduction of Vallisneria spiralis into the Desnogorsk Reservoir. Russ. J. Biol. Invasions 2010, 1, 159–161. [Google Scholar] [CrossRef]
- Babko, R.; Fyda, J.; Kuzmina, T.; Hutorowicz, A. Ciliates on the Macrophytes in Industrially Heated Lakes (Kujawy Lakeland, Poland). Vestn. Zool. 2010, 44, 1–11. [Google Scholar] [CrossRef]
- Zdanowski, B.; Napiorkowska-Krzebietke, A.; Stawecki, K.; Świątecki, A.; Babko, R.; Bogacka-Kapusta, E.; Czarnecki, B.; Kapusta, A. Heated Konin Lakes: Structure, Functioning, and Succession. In Polish River Basins and Lakes—PART I: Hydrology and Hydrochemistry; Korzeniewska, E., Harnisz, M., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 321–349. [Google Scholar]
- Yanygina, L.; Kirillov, V.; Zarubina, E.Y. Invasive species in the biocenosis of the cooling reservoir of Belovskaya power plant (Southwest Siberia). Russ. J. Biol. Invasions 2010, 1, 50–54. [Google Scholar] [CrossRef]
- Lukas, J.; Kalinkat, G.; Kempkes, M.; Rose, U.; Bierbach, D. Feral guppies in Germany-a critical evaluation of a citizen science approach as biomonitoring tool. Bull. Fish Biol. 2017, 17, 13–27. [Google Scholar]







| Country | The First Report (Location) | Reference |
|---|---|---|
| Austria | 1980 | [31] |
| Belgium | 2000 | [32] |
| Croatia | 2017 (River Sava) | [10] |
| Czech Republic | 1991 | [33] |
| France | 1992 (Auvergne) | [34] |
| Germany | 1981 (thermally polluted River Erft) | [35] |
| Hungary | 1966 | [29] |
| Italy | 1998 (Bodrio le Margherite) | [36] |
| Netherlands | 1973 | [37] |
| Norway | 1989 (Lake Stilla, Ak Skedsmo) | [38] |
| Poland | 2012 (Lake Stawiki in Sosnowiecu) | [39] |
| Portugal | 1990 | [40] |
| Romania | 2005 | [41] |
| Russia | 1989 (River Volga) | [42] |
| Serbia | 1994 (thermal spring Banjica) | [43] |
| Slovakia | 2007 | [44] |
| Slovenia | 2001 (thermal stream Topla) | [11] |
| Spain | 2001 (Guipúzcoa) | [30] |
| Sweden | 2006 | [45] |
| United Kingdom | 1983 (pond in London) | [46] |
| Ukraine | 2011 (River Seversky Donets) | [47] |
| Estimate | Std. Error | t-Value | p-Value | |
|---|---|---|---|---|
| Fresh mass | ||||
| a | 14.372 | 1.034 | 13.900 | <0.0001 |
| T0 | 28.779 | 0.324 | 88.944 | <0.0001 |
| sigma | 3.516 | 0.402 | 8.744 | <0.0001 |
| Leaf length | ||||
| a | 7.131 | 0.211 | 33.80 | <0.0001 |
| T0 | 27.231 | 0.133 | 204.16 | <0.0001 |
| sigma | 3.516 | 0.158 | 22.26 | <0.0001 |
| Estimate | Std. Error | t-Value | p-Value | |
|---|---|---|---|---|
| Chl Fluorescence Fv/Fm—actual (GLM, Gaussian) | ||||
| Intercept | 0.464 | 0.020 | 22.873 | <0.0001 |
| Sampling date | 0.019 | 0.001 | 14.213 | <0.0001 |
| Temperature | −0.004 | 0.001 | −3.509 | <0.001 |
| Chl Fluorescence Fv/Fm—dark adapted; Fv/Fm (GLM, Gaussian) | ||||
| Intercept | 0.702 | 0.015 | 45.845 | <0.0001 |
| Sampling date | 0.009 | 0.001 | 8.729 | <0.0001 |
| Temperature | −0.002 | 0.001 | −2.198 | 0.028 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šajna, N.; Urek, T.; Kušar, P.; Šipek, M. The Importance of Thermally Abnormal Waters for Bioinvasions—A Case Study of Pistia stratiotes. Diversity 2023, 15, 421. https://doi.org/10.3390/d15030421
Šajna N, Urek T, Kušar P, Šipek M. The Importance of Thermally Abnormal Waters for Bioinvasions—A Case Study of Pistia stratiotes. Diversity. 2023; 15(3):421. https://doi.org/10.3390/d15030421
Chicago/Turabian StyleŠajna, Nina, Tina Urek, Primož Kušar, and Mirjana Šipek. 2023. "The Importance of Thermally Abnormal Waters for Bioinvasions—A Case Study of Pistia stratiotes" Diversity 15, no. 3: 421. https://doi.org/10.3390/d15030421
APA StyleŠajna, N., Urek, T., Kušar, P., & Šipek, M. (2023). The Importance of Thermally Abnormal Waters for Bioinvasions—A Case Study of Pistia stratiotes. Diversity, 15(3), 421. https://doi.org/10.3390/d15030421