


Natal Philopatry in a Long-Lived Species: The Return of Reproductive River Turtles Marked and Released as Hatchlings

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
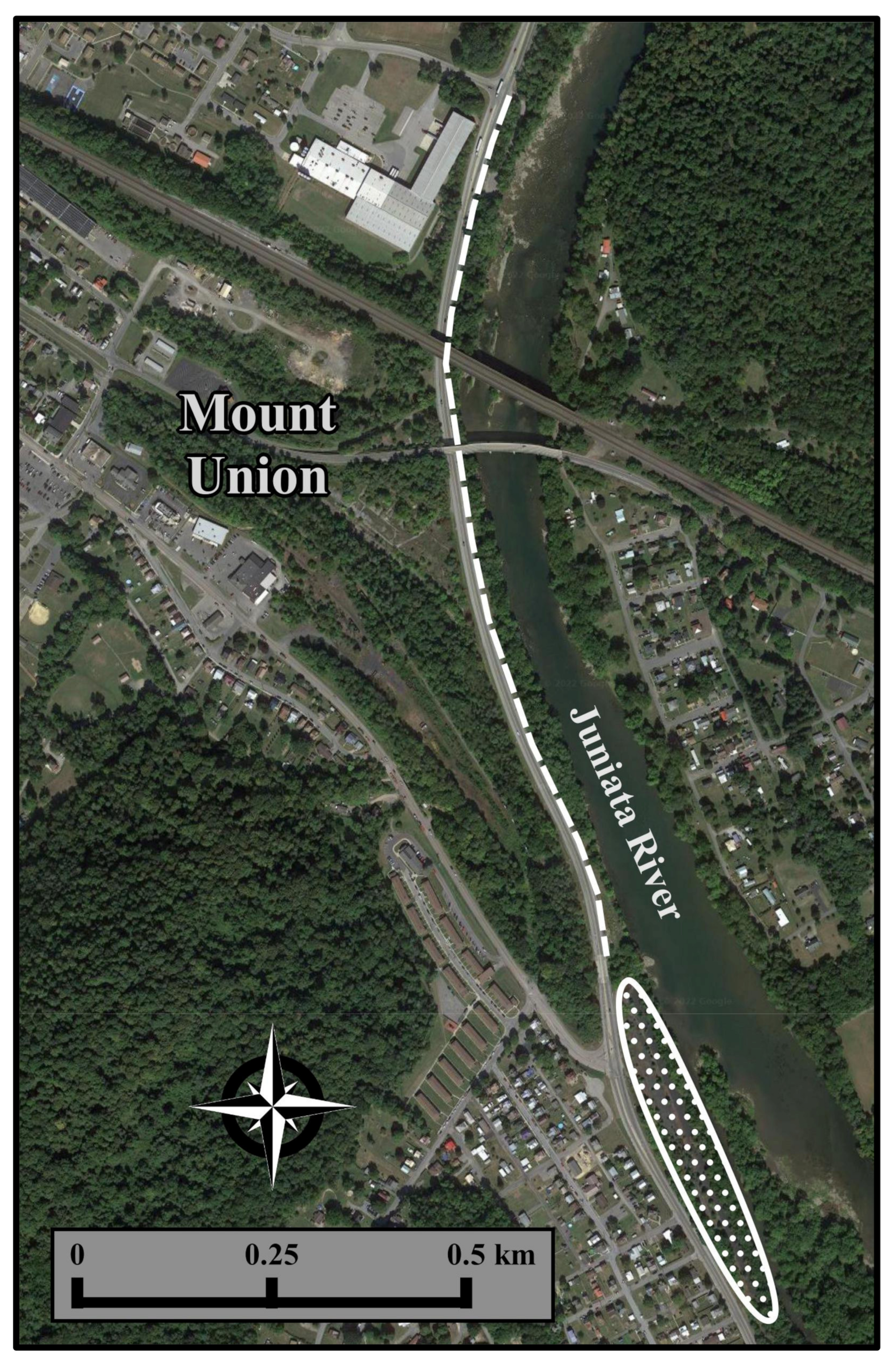
2.1. Study Site
2.2. Adult Females
2.3. Nest Protection
2.4. Hatchling Mark and Release
2.5. Recent Monitoring and Recaptures
2.6. Data Analysis
3. Results
3.1. Nest Success and Hatchlings
3.2. Return of Hatchling-Marked Reproductive Females
3.3. Influence of Maternal Identity and Timing of Release
3.4. Female Longevity and Reproductive Lifetime
3.5. Recapture Locations and Natal Philopatry
3.6. Shell Shape Abnormalities
4. Discussion
4.1. Return of Hatchling-Marked Females
4.2. Super Moms
4.3. Timing of Release
4.4. Recapture Rate
4.5. Natal Philopatry and Recapture Locations
4.6. Shell Shape Abnormalities
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tinkle, D.W. Long-term field studies. BioScience 1979, 29, 717. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Likens, G.E.; Anderson, A.; Bowman, D.; Bull, C.M.; Burns, E.; Dickman, C.R.; Hoffmann, A.A.; Keith, D.A.; Liddell, M.J.; et al. Value of long-term ecological studies. Austral. Ecol. 2012, 37, 7745–7757. [Google Scholar] [CrossRef]
- Sheldon, B.C.; Loeske, E.B.; Albert, K.; Albert, S.C. The expanding value of long-term studies of individuals in the wild. Nat. Ecol. Evol. 2022, 6, 1799–1801. [Google Scholar] [CrossRef] [PubMed]
- Plummer, M.V.; Ferner, J.W. Marking Reptiles. In Reptile Biodiversity: Standard Methods for Inventory and Monitoring; McDiarmid, R.W., Foster, M.S., Guyer, C., Gibbons, J.W., Chernoff, N., Eds.; Univ. of California Press: Berkeley, CA, USA, 2012; pp. 143–150. [Google Scholar]
- Buhlmann, K.A.; Tuberville, T.D. Use of passive integrated transponder (PIT) tags for marking small freshwater turtles. Chelonian Conserv. Biol. 1998, 3, 102–104. [Google Scholar]
- Gibbons, J.W.; Andrews, K.M. Pit tagging: Simple technology at its best. BioScience 2004, 54, 447–454. [Google Scholar] [CrossRef] [Green Version]
- Anderson, K.P.; Byer, N.W.; McGehee, R.J.; Richards–Dimitrie, T. A new system for marking hatchling turtles using visible implant elastomer. Herpetol. Rev. 2015, 46, 25–27. [Google Scholar]
- Davy, C.M.; Coombes, S.M.; Whitear, A.K.; MacKenzie, A.S. Visible implant elastomer: A simple, non-harmful method for marking hatchling turtles. Herpetol. Rev. 2010, 41, 442–445. [Google Scholar]
- Nagle, R.D.; Kinney, O.M.; Gibbons, J.W.; Congdon, J.D. A simple and reliable system for marking hard–shelled turtles: The North American Code. Herpetol. Rev. 2017, 48, 327–330. [Google Scholar]
- Ashmore, G.M.; Janzen, F.J. Phenotypic variation in Smooth Softshell Turtles (Apalone mutica) from eggs incubated in constant versus fluctuating temperatures. Oecologia 2003, 134, 182–188. [Google Scholar] [CrossRef]
- Mullins, M.A.; Janzen, F.J. Phenotypic effects of thermal means and variances on Smooth Softshell Turtle (Apalone mutica) embryos and hatchlings. Herpetologica 2006, 62, 27–36. [Google Scholar] [CrossRef]
- Les, H.L.; Paitz, R.; Bowden, R.M. Experimental test of the effects of fluctuating incubation temperatures on hatchling phenotype. J. Exp. Zool. A Ecol. Genet. Physiol. 2007, 307, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Rowe, C.L.; Liang, D.; Woodland, R.J. Effects of constant and fluctuating incubation temperatures on hatching success and hatchling traits in the Diamondback Terrapin (Malaclemys terrapin) in the context of the warming climate. J. Therm. Biol. 2020, 88, 102528. [Google Scholar] [CrossRef] [PubMed]
- Buhlmann, K.A.; Osborne, C.P. Use of an artificial nesting mound by Wood Turtles (Glyptemys insculpta): A tool for turtle conservation. Northeast. Nat. 2011, 18, 315–334. [Google Scholar] [CrossRef]
- Paterson, J.E.; Steinberg, B.D.; Litzgus, J.D. Revealing a cryptic life-history stage: Differences in habitat selection and survivorship between hatchlings of two turtle species at risk (Glyptemys insculpta and Emydoidea blandingii). Wildl. Res. 2012, 39, 408–418. [Google Scholar] [CrossRef]
- Bougie, T.A.; Byer, N.W.; Lapin, C.N.; Peery, Z.M.; Woodford, J.E.; Pauli, J.N. Wood Turtle (Glyptemys insculpta) nest protection reduces depredation and increases success, but annual variation influences its effectiveness. Can. J. Zool. 2020, 98, 715–724. [Google Scholar] [CrossRef]
- Nagle, R.D. Northern Map Turtle (Graptemys geographica). In Terrestrial Vertebrates of Pennsylvania: A Complete Guide to Species of Conservation Concern; Steele, M.A., Brittingham, M.C., Maret, T.J., Merritt, J.F., Eds.; The Johns Hopkins University Press: Baltimore, MD, USA, 2010; pp. 93–95. [Google Scholar]
- Nagle, R.D.; Congdon, J.D. Reproductive ecology of Graptemys geographica of the Juniata River in central Pennsylvania, with recommendations for conservation. Herpetol. Conserv. Biol. 2016, 11, 232–243. [Google Scholar]
- Nagle, R.D.; Russell, T.J. Nest site fidelity of Northern Map Turtles, Graptemys geographica. Chelonian Conserv. Biol. 2020, 19, 209–216. [Google Scholar] [CrossRef]
- Nagle, R.D.; Lutz, C.L.; Pyle, A.L. Overwintering in the nest by hatchling map turtles (Graptemys geographica). Can. J. Zool. 2004, 82, 1211–1218. [Google Scholar] [CrossRef]
- Bowen, B.W.; Bass, A.L.; Chow, S.M.; Bostrom, M.; Bjorndal, K.A.; Bolten, A.B.; Okuyama, T.; Bolker, B.M.; Epperly, S.; Lacasella, E.; et al. Natal homing in juvenile loggerhead turtles (Caretta caretta). Mol. Ecol. 2004, 13, 3797–3808. [Google Scholar] [CrossRef]
- Weatherhead, P.J.; Forbes, M.R. Natal philopatry in passerine birds: Genetic or ecological influences? Behav. Ecol. 1994, 5, 426–433. [Google Scholar] [CrossRef]
- Nagle, R.D.; Rowe, C.L.; Grant, C.J.; Sebastian, E.R.; Martin, B.E. Abnormal shell shapes in Northern Map Turtles of the Juniata River, Pennsylvania, USA. J. Herpetol. 2018, 52, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Congdon, J.D.; Nagle, R.D.; Kinney, O.M. Front-loading life histories: The enduring influence of juvenile growth on age, size, and reproduction of primiparous female freshwater turtles. Evol. Ecol. Res. 2018, 19, 353–364. [Google Scholar]
- Scribner, K.T.; Congdon, J.D.; Chesser, R.K.; Smith, M.H. Annual differences in female reproductive success affect spatial and cohort-specific genotypic heterogeneity in Painted Turtles. Evolution 1993, 47, 1360–1373. [Google Scholar]
- Roosenburg, W.M. Maternal condition and nest site choice: An alternative for the maintenance of environmental sex determination? Am. Zool. 1996, 36, 157–168. [Google Scholar] [CrossRef] [Green Version]
- Refsnider, J.M.; Janzen, F.J. Putting eggs in one basket: Ecological and evolutionary hypotheses for variation in oviposition-site choice. Ann. Rev. Ecol. Evol. Syst. 2010, 41, 39–57. [Google Scholar] [CrossRef]
- Ferrara, C.R.; Vogt, R.C.; Sousa-Lima, R.S.; Tardio, B.M.; Bernardes, V.C.D. Sound communication and social behavior in an Amazonian River turtle (Podocnemis expansa). Herpetologica 2014, 70, 149–156. [Google Scholar] [CrossRef]
- Bernardo, J. Maternal effects in animal ecology. Am. Zool. 1996, 36, 83–105. [Google Scholar] [CrossRef] [Green Version]
- Moore, M.P.; Whiteman, H.H.; Martin, R.A. A mother’s legacy: The strength of maternal effects in animal populations. Ecol. Lett. 2019, 22, 1620–1628. [Google Scholar] [CrossRef]
- Brooks, R.J.; Bobyn, M.L.; Galbraith, D.A.; Layfield, J.A.; Nancekivell, E.G. Maternal and environmental influences on growth and survival of embryonic and hatchling snapping turtles (Chelydra serpentina). Can. J. Zool. 1991, 69, 2667–2676. [Google Scholar] [CrossRef]
- Páez, V.P.; Echeverri, G.L.; Bock, B.C.; Bowden, R.M.; Hinestroza, L.M. Preovulatory maternal effects on intra- and interpopulation variation in sex ratios and phenotypic characteristics of Magdalena River Turtles (Podocnemis lewyana). Herpetologica 2015, 71, 196–202. [Google Scholar] [CrossRef]
- Lindeman, P.V. The Map Turtle and Sawback Atlas: Ecology, Evolution, Distribution, and Conservation; University of Oklahoma Press: Norman, OK, USA, 2013; pp. 1–460. [Google Scholar]
- Gibbons, J.W. A long-term perspective of delayed emergence (aka overwintering) in hatchling turtles: Some they do and some they don’t, and some you just can’t tell. J. Herpetol. 2013, 47, 203–214. [Google Scholar] [CrossRef]
- Gibbons, J.W.; Nelson, D.H. The evolutionary significance of delayed emergence from the nest by hatchling turtles. Evolution 1978, 32, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Bendick, E. Juniata River at Lewistown Narrows, 2013 to 2015 Continuous Instream Monitoring Report, Pennsylvania Department of Environmental Protection, Bureau of Clean Water. Available online: https://files.dep.state.pa.us/water/Drinking%20Water%20and%20Facility%20Regulation/WaterQualityPortalFiles/CIMR/Juniata_Lewistown_CIMR_2013_2015.pdf. (accessed on 25 January 2023).
- Congdon, J.D. Proximate and evolutionary constraints on energy relations of reptiles. Physiol. Zool. 1989, 62, 356–373. [Google Scholar] [CrossRef] [Green Version]
- Nagle, R.D.; Burke, V.J.; Congdon, J.D. Egg components and hatchling lipid reserves: Parental investment in kinosternid turtles from the southeastern United States. Comp. Biochem. Physiol. B 1998, 120, 145–152. [Google Scholar] [CrossRef]
- Ferrara, C.R.; Vogt, R.C.; Sousa-Lima, R.S. Turtle vocalizations as the first evidence of post-hatching parental care in chelonians. J. Comp. Psychol. 2012, 127, 24–32. [Google Scholar] [CrossRef] [Green Version]
- Iverson, J.B. Patterns of survivorship in turtles (order Testudines). Can. J. Zool. 1991, 69, 385–391. [Google Scholar] [CrossRef]
- Gibbons, J.W. Life History and Ecology of the Slider Turtle; Smithsonian Institution Press: Washington, DC, USA, 1990; pp. 1–368. [Google Scholar]
- Frazer, N.B.; Gibbons, J.W.; Greene, J.L. Life history and demography of the Common Mud Turtle (Kinosternon subrubrum) in South Carolina, USA. Ecology 1991, 72, 2218–2231. [Google Scholar] [CrossRef]
- Congdon, J.D.; Dunham, A.E.; Van Loben Sels, R.C. Demographics of Common Snapping Turtles: Implications for conservation and management of long-lived organisms. Am. Zool. 1994, 34, 397–408. [Google Scholar] [CrossRef] [Green Version]
- Congdon, J.D.; Nagle, R.D.; Kinney, O.M.; Osentoski, M.F.; Avery, H.W.; Van Loben Sels, R.C.; Tinkle, D.W. Nesting ecology and embryo mortality: Implications for hatching success and demography of Blanding’s Turtles (Emydoidea blandingii). Chelonian Conserv. Biol. 2000, 3, 569–579. [Google Scholar]
- Kolbe, J.J.; Janzen, F.J. Experimental analysis of an early life-history stage: Water loss and migrating hatchling turtles. Copeia 2002, 2002, 220–226. [Google Scholar] [CrossRef]
- Janzen, F.J.; Tucker, J.K.; Paukstis, G.L. Experimental analysis of an early life-history stage: Direct or indirect selection on body size of hatchling turtles? Funct. Ecol. 2007, 21, 162–170. [Google Scholar] [CrossRef]
- Delaney, D.M.; Janzen, F.J. Offspring dispersal ability covaries with nest-site choice. Behav. Ecol. 2018, 30, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Congdon, J.D.; Pappas, M.; Brecke, B.; Capps, J. Conservation implications of initial orientation of naive hatchling Snapping Turtles (Chelydra serpentina) and Painted Turtles (Chrysemys picta belli) dispersing from experimental nests. Chelonian Conserv. Biol. 2011, 10, 42–53. [Google Scholar] [CrossRef]
- Pappas, M.J.; Congdon, J.D.; Brecke, B.J. Orientation in five species of hatchling turtles dispersing from artificial nests. Chelonian Conserv. Biol. 2017, 16, 3–11. [Google Scholar] [CrossRef]
- Geller, G.A.; Casper, G.S.; Halstead, B.J. Dispersal of hatchling Ouachita Map Turtles (Graptemys ouachitensis) from natural nests on the Lower Wisconsin River, Wisconsin. Chelonian Conserv. Biol. 2020, 19, 236–245. [Google Scholar] [CrossRef]
- Pluto, T.G.; Bellis, E.D. Seasonal and annual movements of riverine map turtles, Graptemys geographica. J. Herpetol. 1988, 22, 152–158. [Google Scholar] [CrossRef]
- Carrière, M.A.; Bulté, G.; Blouin-Demers, G. Spatial ecology of Northern Map Turtles (Graptemys geographica) in a lotic and a lentic habitat. J. Herpetol. 2009, 43, 597–604. [Google Scholar] [CrossRef]
- Lohmann, K.J.; Lohmann, C.M.; Endres, C.S. The sensory ecology of ocean navigation. J. Exp. Biol. 2008, 211, 1719–1728. [Google Scholar] [CrossRef] [Green Version]
- Lohmann, K.J.; Putman, N.F.; Lohmann, C.M. Geomagnetic imprinting: A unifying hypothesis of long-distance natal homing in salmon and sea turtles. Proc. Natl. Acad. Sci. USA 2008, 105, 19096–19101. [Google Scholar] [CrossRef] [Green Version]
- Brothers, J.R.; Lohmann, K.J. Evidence for geomagnetic imprinting and magnetic navigation in the natal homing of sea turtles. Curr. Biol. 2015, 25, 392–396. [Google Scholar] [CrossRef] [Green Version]
- Lohmann, K.J.; Lohmann, C.M. There and back again: Natal homing by magnetic navigation in sea turtles and salmon. J. Exp. Biol. 2019, 222, jeb184077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindeman, P.V. Nest–site fixity among Painted Turtles (Chrysemys picta) in northern Idaho. Northwest. Nat. 1992, 73, 27–30. [Google Scholar] [CrossRef]
- Doody, J.S.; Dinets, V.; Burghardt, G.M. The Secret Social Lives of Reptiles; Johns Hopkins University Press: Baltimore, MD, USA, 2017; pp. 1–440. [Google Scholar]
- Reid, B.N.; Thiel, R.P.; Palsboll, P.J.; Peery, M.Z. Linking genetic kinship and demographic analyses to characterize dispersal: Methods and application to Blanding’s Turtle. J. Hered. 2016, 107, 603–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freedberg, S.; Ewert, M.A.; Ridenhour, B.J.; Neiman, M.; Nelson, C.E. Nesting fidelity and molecular evidence for natal homing in the freshwater turtle, Graptemys kohnii. Proc. Biol. Sci. 2005, 272, 1345–1350. [Google Scholar] [CrossRef] [Green Version]
- Reinhold, K. Nest–site philopatry and selection for environmental sex determination. Evol. Ecol. 1998, 12, 245–250. [Google Scholar] [CrossRef]
- Byer, N.W.; Reid, B.N. The emergence of imperfect philopatry and fidelity in spatially and temporally heterogeneous environments. Ecol. Modell. 2022, 468, 109968. [Google Scholar] [CrossRef]
- Sheridan, C.M.; Spotila, J.R.; Bien, W.F.; Avery, H.W. Sex-biased dispersal and natal philopatry in the Diamondback Terrapin, Malaclemys terrapin. Mol. Ecol. 2010, 19, 5497–5510. [Google Scholar] [CrossRef]
- Valenzuela, N. Genetic differentiation among nesting beaches in the highly migratory giant river turtle (Podocnemis expansa) from Colombia. Herpetologica 2001, 57, 48–57. [Google Scholar]
- Allard, M.W.; Miyamoto, M.M.; Bjorndal, K.A.; Bolten, A.B.; Bowen, B.W. Support for natal homing in Green Turtles from mitochondrial DNA sequences. Copeia 1994, 1994, 34–41. [Google Scholar] [CrossRef]
- Meylan, A.B.; Bowen, B.W.; Avise, J.C. A genetic test of the natal homing versus social facilitation models for Green Turtle migration. Science 1990, 248, 724–727. [Google Scholar] [CrossRef] [Green Version]
- Bowen, B.; Avise, J.C.; Richardson, J.I.; Meylan, A.B.; Margaritoulis, D.; Hopkins Murphy, S.R. Population structure of Loggerhead Turtles (Caretta caretta) in the northwestern Atlantic Ocean and Mediterranean Sea. Conserv. Biol. 1993, 7, 834–844. [Google Scholar] [CrossRef] [Green Version]
- Bass, A.L.; Good, D.A.; Bjorndal, K.A.; Richardson, J.I.; Hillis, Z.M.; Horrocks, J.A.; Bowen, B.W. Testing models of female reproductive migratory behaviour and population structure in the Caribbean Hawksbill Turtle, Eretmochelys imbricata, with mtDNA sequences. Mol. Ecol. 1996, 5, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Dutton, P.H.; Bowen, B.W.; Owens, D.W.; Barragan, A.; Davis, S.K. Global phylogeography of the Leatherback Turtle (Dermochelys coriacea). J. Zool. 1999, 248, 397–409. [Google Scholar] [CrossRef]
- Freedberg, S.; Lee, C.; Pappas, M. Agricultural practices alter sex ratios in a reptile with environmental sex determination. Biol. Conserv. 2011, 144, 1159–1166. [Google Scholar] [CrossRef]
- Freedberg, S.; Wade, M.J. Cultural inheritance as a mechanism for population sex-ratio bias in reptiles. Evolution 2001, 55, 1049–1055. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | n | Mean | Std Dev | Range |
|---|---|---|---|---|
| Age (years) | 46 | 17.0 | 2.9 | 11–22 |
| Plastron Length (mm) | 46 | 192.8 | 7.7 | 172–210 |
| Carapace Length (mm) | 46 | 226.1 | 9.7 | 207–249 |
| Body Mass (g) | 45 | 1568.4 | 219.3 | 1173–2016 |
| Distance to natal nests on coal pile (m) | 11 | 196.5 | 109.1 | 67.2–446.2 |
| Distance to natal nests in fenced mitigation area (m) | 61 | 665.3 | 381.3 | 39.1–1527.9 |
| Number of Female Siblings Returned Per Nest | Number of Nests | Product (Number of Females) |
|---|---|---|
| 1 | 14 | 14 |
| 2 | 5 | 10 |
| 3 | 2 | 6 |
| 4 | 1 | 4 |
| 5 | 1 | 5 |
| 7 | 1 | 7 |
| Total | 24 | 46 |
| Month Released | Number Released | Number Returned | Percent Returned |
|---|---|---|---|
| March | 66 | 9 | 13.6 |
| April | 477 | 14 | 2.9 |
| May | 77 | 21 | 27.3 |
| August | 69 | 2 | 2.9 |
| September | 3 | 0 | 0 |
| Total | 691 | 46 | 6.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagle, R.D.; Russell, T.J.; Grant, C.J.; Innerst, M.; Strawser, S.J. Natal Philopatry in a Long-Lived Species: The Return of Reproductive River Turtles Marked and Released as Hatchlings. Diversity 2023, 15, 444. https://doi.org/10.3390/d15030444
Nagle RD, Russell TJ, Grant CJ, Innerst M, Strawser SJ. Natal Philopatry in a Long-Lived Species: The Return of Reproductive River Turtles Marked and Released as Hatchlings. Diversity. 2023; 15(3):444. https://doi.org/10.3390/d15030444
Chicago/Turabian StyleNagle, Roy D., Travis J. Russell, Christopher J. Grant, Melissa Innerst, and Seth J. Strawser. 2023. "Natal Philopatry in a Long-Lived Species: The Return of Reproductive River Turtles Marked and Released as Hatchlings" Diversity 15, no. 3: 444. https://doi.org/10.3390/d15030444
APA StyleNagle, R. D., Russell, T. J., Grant, C. J., Innerst, M., & Strawser, S. J. (2023). Natal Philopatry in a Long-Lived Species: The Return of Reproductive River Turtles Marked and Released as Hatchlings. Diversity, 15(3), 444. https://doi.org/10.3390/d15030444