
Artificial Waterholes for European Bison as Biodiversity Hotspots in Forest Ecosystems: Ecological Effects of Species Reintroduction Activities

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
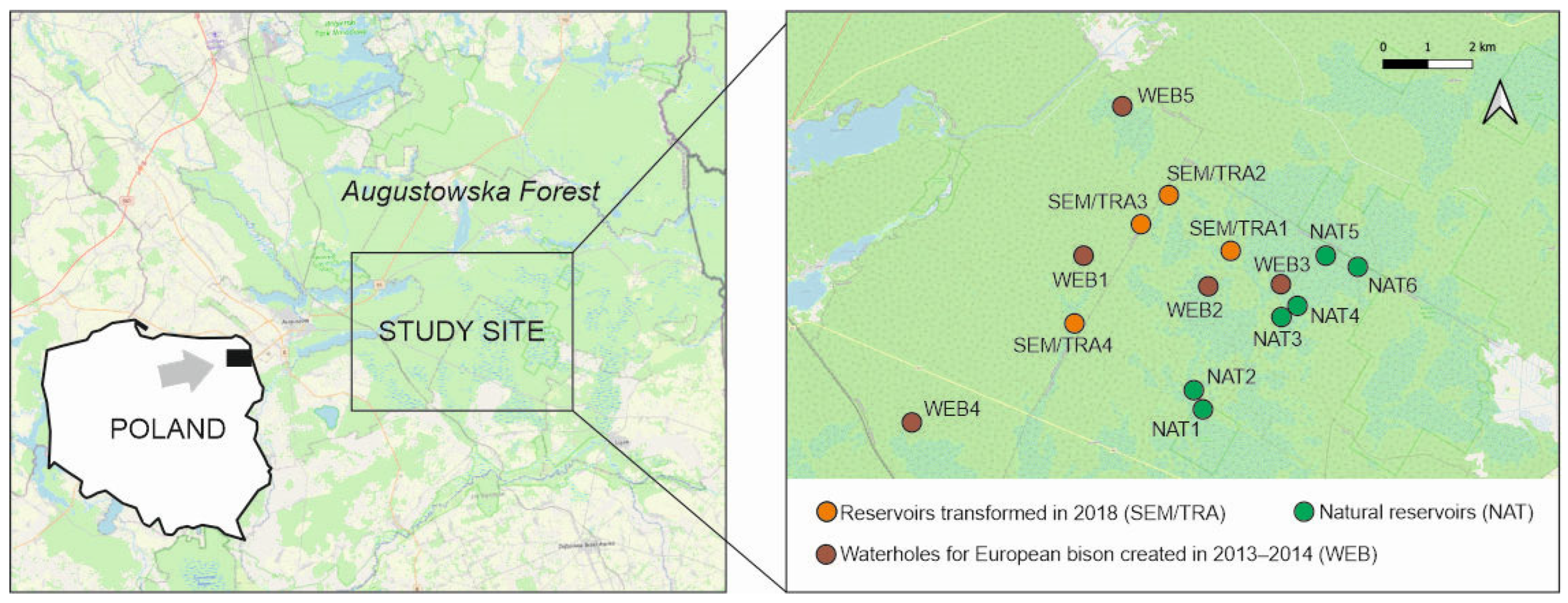
2. Material and Methods
- -
- Five waterholes for the European bison in mid-forest meadows created in 2013–2014, herein called WEB;
- -
- Four seminatural waterholes, which were transformed into waterholes for the E. bison in 2018. Transformation into waterholes consisted in cleaning the reservoir, enlarging its surface, creating shallow banks, and uncovering the surroundings by cutting down the tree vegetation near the reservoir. Before transformation, waterholes were called SEM, and after transformation, they were named TRA. The semi-natural reservoir’s (SEM) numbers correspond to the same reservoir after transformation (TRA), i.e., TRA1 is the same reservoir as SEM1, but after transformation;
- -
- Six natural water reservoirs detected near artificial/transformed waterholes for European bison. Waterholes for European bison are usually created in places where natural water reservoirs [22] are not available; therefore, natural reservoirs located as close as possible to artificial/transformed waterholes for European bison were selected for comparative studies. The reservoirs were named NAT. Natural reservoirs were either ephemeral or heavily overgrown and significantly shaded.
2.1. Dragonflies
- -
- Autochthonous species: when exuviae were collected, and/or teneral individuals were observed, and/or numerous breeding behaviors were detected;
- -
- Probable autochthonous species: when few or isolated breeding behaviors such as tandems, copulation, or oviposition were recorded and/or at least a few individuals displaying territorial behavior and/or numerous adults in an environment suitable for development were detected;
- -
- Recorded species: in other cases.
2.2. Amphibians
- -
- The listening method—to record the presence of anuran amphibians during the visit, lasting from 10 to 30 min, depending on size of the pond;
- -
- Direct observation of water (including water plants) and banks, where individuals in different stages of development (including eggs) were visually sought. On each reservoir, one passage was made along the banks around the entire reservoir, except for in inaccessible places;
- -
- Capturing individuals with a herpetological net.
2.3. Bats
- -
- Capturing bats in chiropterological nets. A net, 6 m long, was placed as close as possible to a given reservoir. Capturing was carried out for three full, consecutive nights (in each reservoir and in each year of the study). The caught individuals were taken out of the net and marked according to species using standard methods [32,34,40].
3. Results
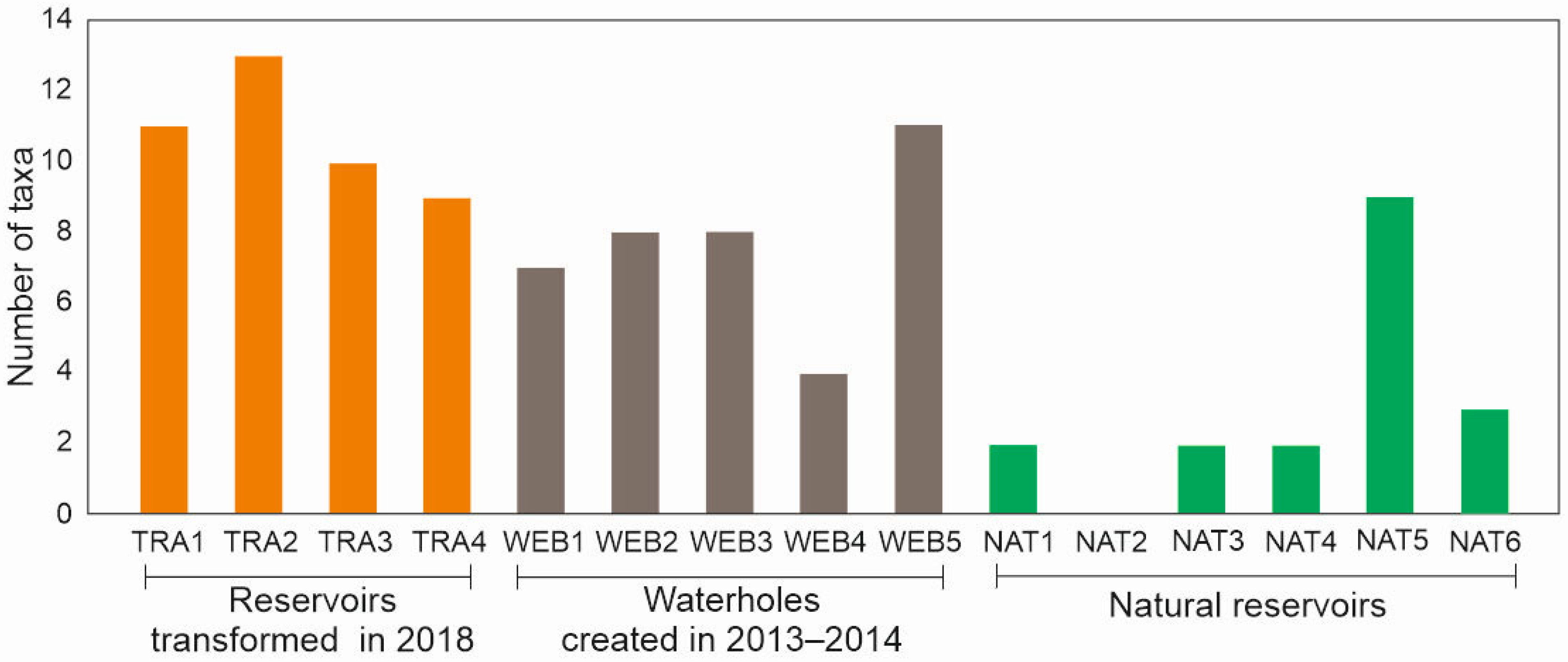
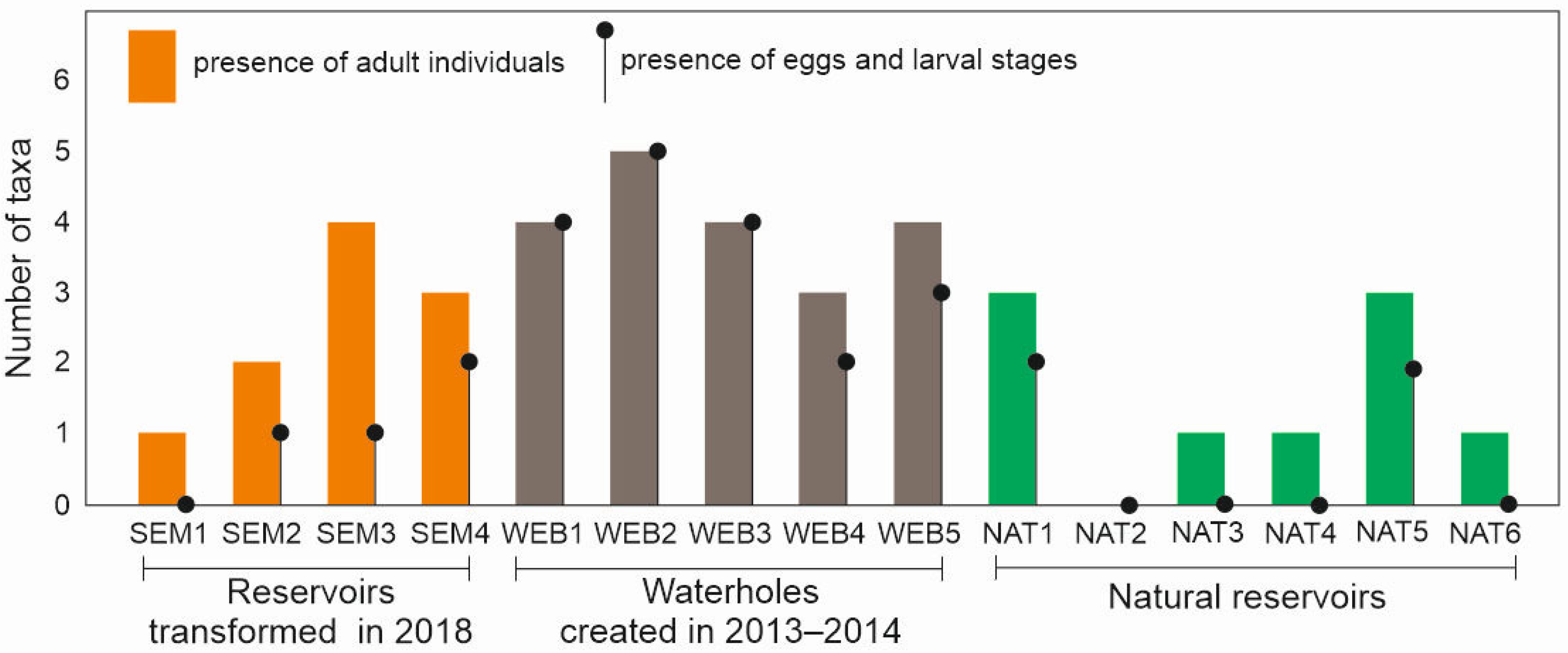
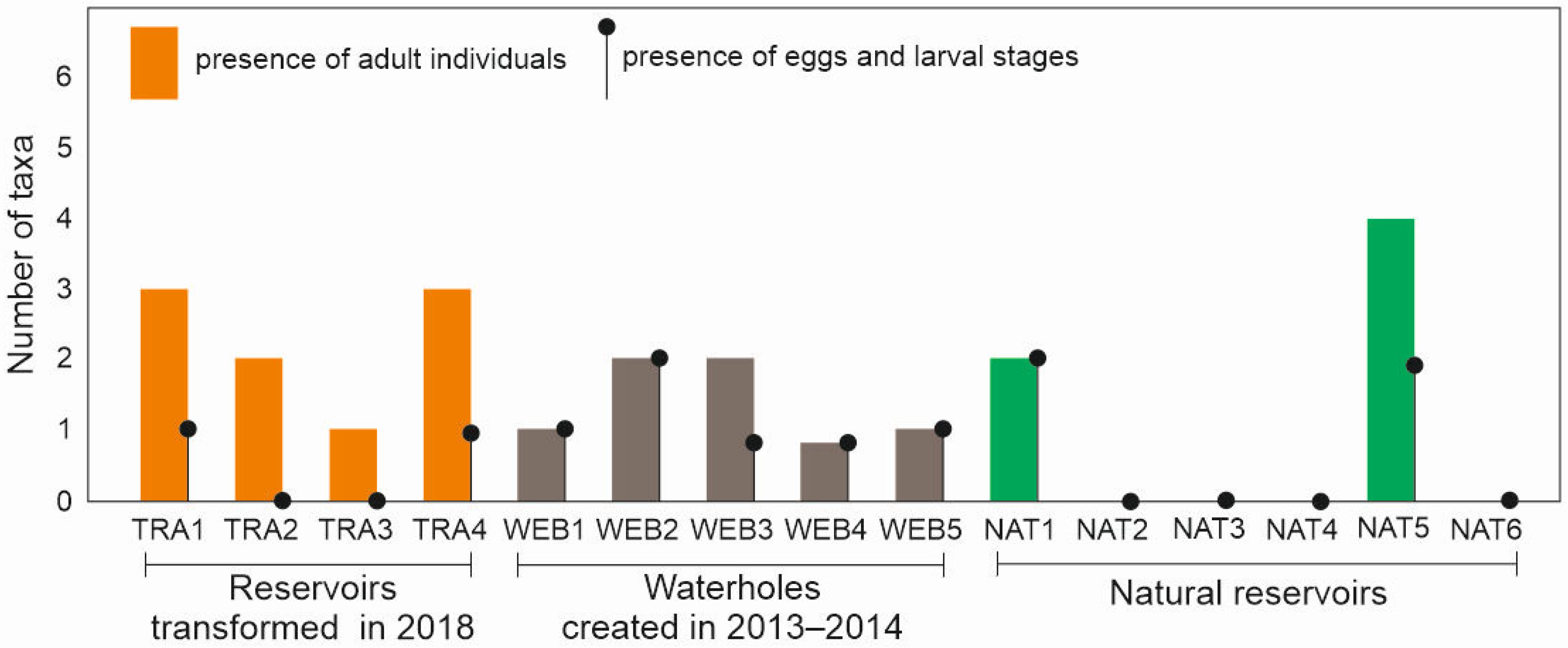
3.1. Dragonflies
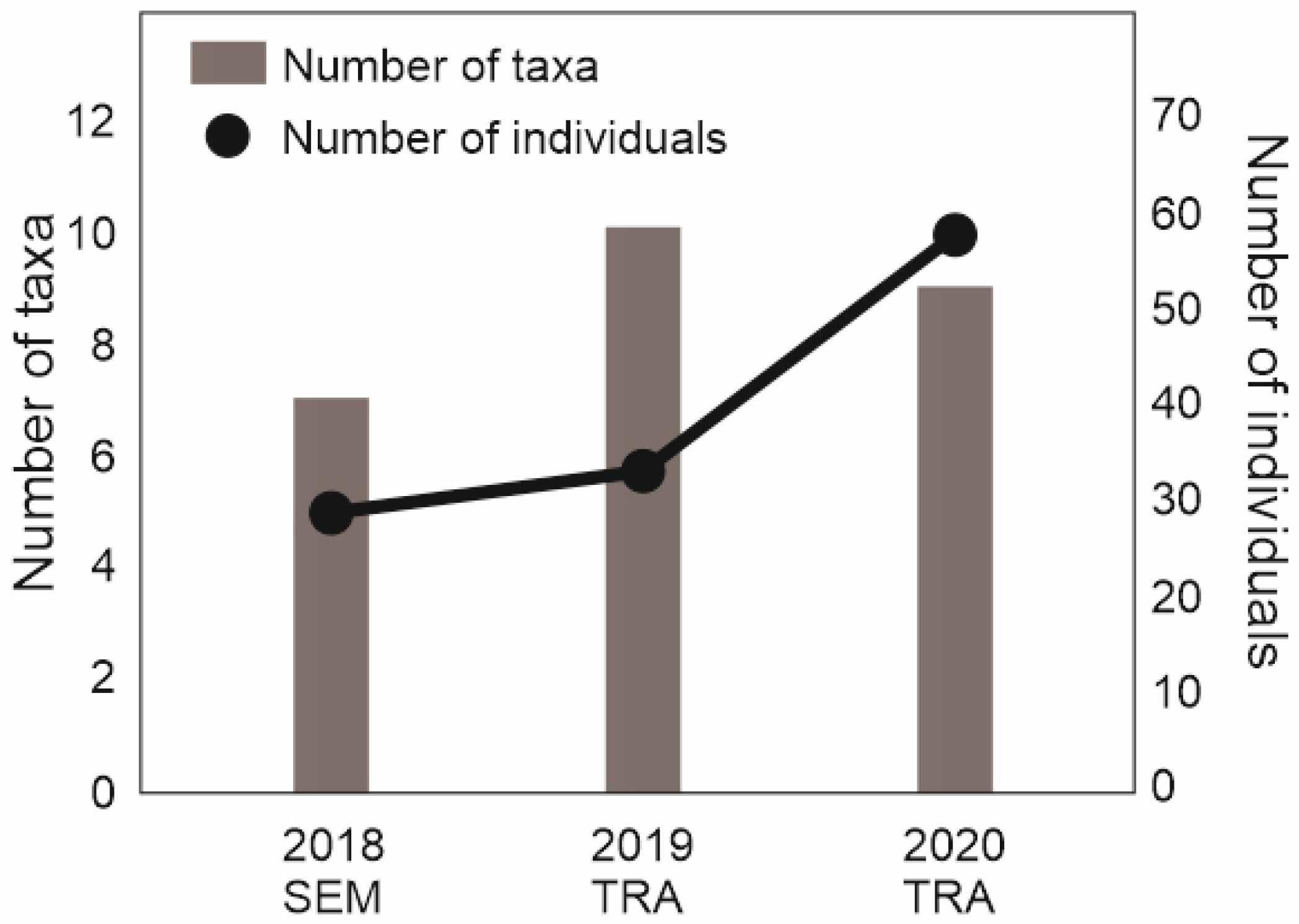
3.2. Amphibians
3.3. Bats
4. Discussion
4.1. Dragonflies
4.2. Amphibians
4.3. Bats
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Olech, W.; Perzanowski, K. (Eds.) European Bison (Bison bonasus) Strategic Species Status Review 2020; IUCN SSC Bison Specialist Group and European Bison Conservation Center: Warsaw, Poland, 2022. [Google Scholar]
- Olech, W.; Klich, D.; Perzanowski, K. Development of a new Action Plan for the European bison. Oryx 2019, 53, 214. [Google Scholar] [CrossRef] [Green Version]
- Perzanowski, K.; Klich, D.; Olech, W. European Union needs urgent strategy for the European bison. Conserv. Lett. 2022, 15, e12923. [Google Scholar] [CrossRef]
- Raczyński, J. The European Bison Pedigree Book 2021; Białowieża National Park: Białowieża, Poland, 2022. [Google Scholar]
- Klich, D.; Olech, W.; Łopucki, R.; Danik, K. Community attitudes to the European bison Bison bonasus in areas where its reintroduction is planned and in areas with existing populations in northeastern Poland. Eur. J. Wildl. Res. 2018, 64, 61. [Google Scholar] [CrossRef] [Green Version]
- Lord, C.M.; Wirebach, K.P.; Tompkins, J.; Bradshaw-Wilson, C.; Shaffer, C.L. Reintroduction of the European bison (Bison bonasus) in central-eastern Europe: A case study. Int. J. Geogr. Inf. Sci. 2020, 34, 1628–1647. [Google Scholar] [CrossRef]
- Kuemmerle, T.; Radeloff, V.C.; Perzanowski, K.; Kozlo, P.; Sipko, T.; Khoyetskyy, P.; Bashta, A.-T.; Chikurova, E.; Parnikoza, I.; Baskin, L.; et al. Predicting potential European bison habitat across its former range. Ecol. Appl. 2011, 21, 830–843. [Google Scholar] [CrossRef] [PubMed]
- Kołodziej-Sobocińska, M.; Demiaszkiewicz, A.W.; Pyziel, A.M.; Kowalczyk, R. Increased Parasitic Load in Captive-Released European Bison (Bison bonasus) has Important Implications for Reintroduction Programs. EcoHealth 2018, 15, 467–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perzanowski, K.; Bleyhl, B.; Olech, W.; Kuemmerle, T. Connectivity or isolation? Identifying reintroduction sites for multiple conservation objectives for wisents in Poland. Anim. Conserv. 2020, 23, 212–221. [Google Scholar] [CrossRef] [Green Version]
- Gałązka, M.; Klich, D.; Anusz, K.; Pyziel-Serafin, A.M. Veterinary monitoring of gastrointestinal parasites in European bison, Bison bonasus designated for translocation: Comparison of two coprological methods. Int. J. Parasitol. Parasites Wildl. 2022, 17, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Borowski, S.; Kossak, S. Bisoniana LI. The natural food preferences of the European bison in seasons free of snow cover. Acta Thériol. 1972, 17, 151–169. [Google Scholar] [CrossRef] [Green Version]
- Kowalczyk, R.; Wójcik, J.M.; Taberlet, P.; Kamiński, T.; Miquel, C.; Valentini, A.; Craine, J.M.; Coissac, E. Foraging plasticity allows a large herbivore to persist in a sheltering forest habitat: DNA metabarcoding diet analysis of the European bison. For. Ecol. Manag. 2019, 449, 117474. [Google Scholar] [CrossRef]
- Kuemmerle, T.; Levers, C.; Bleyhl, B.; Olech, W.; Perzanowski, K.; Reusch, C.; Kramer-Schadt, S. One size does not fit all: European bison habitat selection across herds and spatial scales. Landsc. Ecol. 2018, 33, 1559–1572. [Google Scholar] [CrossRef]
- Jaroszewicz, B.; Pirożnikow, E.; Sagehorn, R. Endozoochory by European bison (Bison bonasus) in Białowieża Primeval Forest across a management gradient. For. Ecol. Manag. 2009, 258, 11–17. [Google Scholar] [CrossRef]
- Jaroszewicz, B. Endozoochory by European bison influences the build-up of the soil seed bank in subcontinental coniferous forest. Eur. J. For. Res. 2013, 132, 445–452. [Google Scholar] [CrossRef] [Green Version]
- Schulze, K.A.; Buchwald, R.; Heinken, T. Epizoochory via the hooves–the European bison (Bison bonasus L.) as a dispersal agent of seeds in an open-forest-mosaic. Tuexenia 2014, 34, 131–144. [Google Scholar]
- Jaroszewicz, B.; Borysowicz, J.; Cholewińska, O. Forest floor plant diversity drives the use of mature spruce forests by European bison. Ecol. Evol. 2021, 11, 636–647. [Google Scholar] [CrossRef]
- Paszkiewicz, R.; Januszczak, M. Szkody powodowane przez żubry w środowisku leśnym Bieszczadów w ocenie leśników. Eur. Bison Conserv. Newsl. 2010, 3, 53–62. [Google Scholar]
- Nieszała, A.; Klich, D.; Perzanowski, K.; Januszczak, M.; Wołoszyn-Gałęza, A.; Olech, W. Debarking intensity of European bison in the Bieszczady Mountains in relation to forest habitat features. For. Ecol. Manag. 2022, 508, 120057. [Google Scholar] [CrossRef]
- Kowalczyk, R.; Kamiński, T.; Borowik, T. Do large herbivores maintain open habitats in temperate forests? For. Ecol. Manag. 2021, 494, 119310. [Google Scholar] [CrossRef]
- Schwerk, A.; Klich, D.; Wójtowicz, E.; Olech, W. Impact of European Bison Grazing (Bison bonasus (L.)) on Species and Functional Traits of Carabid Beetle Assemblages in Selected Habitats in Poland. Biology 2021, 10, 123. [Google Scholar] [CrossRef] [PubMed]
- Olech, W.; Perzanowski, K. Podręcznik Najlepszych Praktyk Ochrony Żubra; Centrum Koordynacji Projektów Środowiskowych: Warszawa, Poland, 2014; pp. 1–96. [Google Scholar]
- Klich, D.; Olech, W.; Cielniak, K. A complex project for the conservation of European bison in Poland by State Forests (2017–2020). Eur. Bison Conserv. Newsl. 2017, 10, 11–20. [Google Scholar]
- Wright, J.P.; Jones, C.G.; Flecker, A.S. An ecosystem engineer, the beaver, increases species richness at the landscape scale. Oecologia 2002, 132, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Orazi, V.; Hagge, J.; Gossner, M.M.; Müller, J.; Heurich, M. A Biodiversity Boost from the Eurasian Beaver (Castor fiber) in Germany’s Oldest National Park. Front. Ecol. Evol. 2022, 10. [Google Scholar] [CrossRef]
- Chovanec, A. Man-made wetlands in urban recreational areas—a habitat for endangered species? Landsc. Urban Plan. 1994, 29, 43–54. [Google Scholar] [CrossRef]
- Rannap, R.; Lõhmus, A.; Briggs, L. Restoring ponds for amphibians: A success story. Hydrobiologia 2009, 634, 87–95. [Google Scholar] [CrossRef]
- Rydell, J.; Bushby, A.; Cosgrove, C.C.; Racey, P.A. Habitat Use by Bats Along Rivers in North East Scotland. Folia Zool. 1994, 43, 417–424. [Google Scholar]
- Zahn, A.; Maier, S. Jagdaktivität von Fledermäusen an Bächen und Teichen. Ztschr. Säugetierk. 1997, 62, 1–11. [Google Scholar]
- Buczyński, P. Dragonflies (Odonata) of Anthropogenic Waters in Middle-Eastern Poland; Wydawnictwo Mantis: Olsztyn, Poland, 2015; pp. 1–272. [Google Scholar]
- Bobrek, R. Post-mining ponds in the Sandomierz Forest (SE Poland) as an important site for the conservation of a species-rich odonate assemblage. Acta Zool. Cracov. 2021, 64, 159–168. [Google Scholar] [CrossRef]
- Dietz, C.; Helversen, O.; Nill, D. Nietoperze Europy i Afryki Północno-Zachodniej; Multico Oficyna Wydawnicza: Warszawa, Poland, 2009; pp. 1–400. [Google Scholar]
- Van De Sijpe, M.; Vandendriessche, B.; Voet, P.; Vandenberghe, J.; Duyck, J.; Naeyaert, E.; Manhaeve, M.; Martens, E. Summer distribution of the Pond bat Myotis dasycneme (Chiroptera, Vespertilionidae) in the west of Flanders (Belgium) with regard to water quality. Mammalia 2004, 68, 377–386. [Google Scholar] [CrossRef]
- Sachanowicz, K.; Ciechanowski, M. Nietoperze Polski; Multico Oficyna Wydawnicza: Warszawa, Poland, 2005. [Google Scholar]
- Buczyński, P.; Tończyk, G.; Daraż, B.; Djatlova, E.; Michalczuk, W.; Miszta, A.; Szpala, B.; Szymański, J.; Tondys, J. Ważki zebrane podczas III Ogólnopolskiego Sympozjum Odonatologicznego PTE (Zwierzyniec, 15-17 IX 2006). Odonatrix 2006, 2 (Suppl. S1), 1–12. [Google Scholar]
- Bernard, R.; Buczyński, P.; Tończyk, G.; Wendzonka, J. Atlas Rozmieszczenia Ważek (Odonata) w Polsce; Bogucki Wydawnictwo Naukowe: Poznań, Poland, 2009; pp. 1–256. [Google Scholar]
- Juszczyk, W. Płazy i Gady Krajowe. Część 2. Płazy–Amphibia; Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1987. [Google Scholar]
- Kierzkowski, P.; Kosiba, P.; Rybacki, M.; Socha, M.; Ogielska, M. Genome dosage effect and colouration features in hybridogenetic water frogs of the Pelophylax esculentus complex. Amphibia-Reptilia 2013, 34, 493–504. [Google Scholar] [CrossRef]
- Rachwald, A.; Fuszara, M. Podręcznik Najlepszych Praktyk Ochrony Nietoperzy w Lasach; CKPŚ: Warszawa, Poland, 2014. [Google Scholar]
- Kowalski, M.; Lesiński, G. (Eds.) Oznaczamy krajowe gatunki nietoperzy. In Poznajemy Nietoperze. ABC Wiedzy o Nietoperzach, ich Badaniu i Ochronie; OTON: Warszawa, Poland, 2000; pp. 38–42. [Google Scholar]
- Rozporządzenie Ministra Środowiska z Dnia 16 Grudnia 2016 r. w Sprawie Ochrony Gatunkowej Zwierząt. Dziennik Ustaw (2016), pozycja 2183. 2016. Available online: https://www.infor.pl/akt-prawny/DZU.2016.253.0002183,rozporzadzenie-ministra-srodowiska-w-sprawie-ochrony-gatunkowej-zwierzat.html (accessed on 17 September 2022).
- Directive, H. Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. Off. J. Eur. Communities 1992, 206, 50. [Google Scholar]
- Mackiewicz, K.; Tończyk, G. Obserwacje ważek (Odonata) w okolicach Sejn (Polska północno-wschodnia) w latach 2016–2017. Odonatrix 2019, 15, 1–28. [Google Scholar]
- Bried, J.T.; Hager, B.J.; Hunt, P.D.; Fox, J.N.; Jensen, H.J.; Vowels, K.M. Bias of reduced-effort community surveys for adult Odonata of lentic waters. Insect Conserv. Divers. 2011, 5, 213–222. [Google Scholar] [CrossRef]
- Oertli, B.; Joye, D.A.; Castella, E.; Juge, R.; Cambin, D.; Lachavanne, J.-B. Does size matter? The relationship between pond area and biodiversity. Biol. Conserv. 2002, 104, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Kadoya, T.; Suda, S.-I.; Washitani, I. Dragonfly species richness on man-made ponds: Effects of pond size and pond age on newly established assemblages. Ecol. Res. 2004, 19, 461–467. [Google Scholar] [CrossRef]
- Remsburg, A.J.; Olson, A.C.; Samways, M.J. Shade Alone Reduces Adult Dragonfly (Odonata: Libellulidae) Abundance. J. Insect Behav. 2008, 21, 460–468. [Google Scholar] [CrossRef]
- Holtmann, L.; Juchem, M.; Brüggeshemke, J.; Möhlmeyer, A.; Fartmann, T. Stormwater ponds promote dragonfly (Odonata) species richness and density in urban areas. Ecol. Eng. 2018, 118, 1–11. [Google Scholar] [CrossRef]
- Bernard, R.; Buczyński, P.; Tończyk, G. Present state, threats, and conservation of dragonflies (Odonata) in Poland. Nat. Conserv. 2002, 59, 53–71. [Google Scholar]
- Reading, C.J. Linking global warming to amphibian declines through its effects on female body condition and survivorship. Oecologia 2006, 151, 125–131. [Google Scholar] [CrossRef]
- Hairston, N.G. Predation and competition in salamander communities. In Long Term Study of Vertebrate Communities; Cody, M.L., Smallwood, J.A., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 161–169. [Google Scholar]
- Pechmann, J.H.K.; Scott, D.E.; Semlitsch, R.D.; Caldwell, T.; Hartel, J.P.; Vitt, L.J.; Gibbons, J.W. Declining Amphibian Populations: The Problem of Separating Human Impacts from Natural Fluctuations. Science 1991, 253, 892–895. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, R.A. Temporary ponds as amphibian habitats. Aquat. Conserv. Mar. Freshw. Ecosyst. 1997, 7, 119–126. [Google Scholar] [CrossRef]
- Juszczyk, W.; Świerad, J. Rocznik Naukowo-Dydaktyczny. Z. 50. Pr. Zool. 1973, 3, 47–66. [Google Scholar]
- Krzysztofiak, L.; Krzysztofiak, A. Czynna Ochrona Płazów; Stowarzyszenie, Człowiek i Przyroda: Krzywe, Poland, 2016; pp. 1–68. [Google Scholar]
- Rafiński, J.; Babik, W. Traszka grzebieniasta Triturus cristatus (Laurenti, 1768). In Atlas płazów i gadów Polski. Status–rozmieszczenie–ochrona; Głowaciński, Z., Rafiński, J., Eds.; Biblioteka Monitoringu Środowiska; Główny Inspektorat Ochrony Środowiska: Warszawa-Kraków, Poland, 2003; pp. 30–32. [Google Scholar]
- Miró, A.; O’Brien, D.; Hall, J.; Jehle, R. Habitat requirements and conservation needs of peripheral populations: The case of the great crested newt (Triturus cristatus) in the Scottish Highlands. Hydrobiologia 2016, 792, 169–181. [Google Scholar] [CrossRef] [Green Version]
- Arntzen, J.W.; Teunis, S.F.M. A six year study on the population dynamics of the crested newt (Triturus cristatus) following the colonization of a newly created pond. Herpetol. J. 1993, 3, 99–110. [Google Scholar]
- Jarvis, L.E. Microhabitat Requirements of the Great Crested Newt (Triturus cristatus) in a Woodland Area. Ph.D. Thesis, The Open University, Milton Keynes, UK, 2012. [Google Scholar]
- Vaughan, N.; Jones, G.; Harris, S. Habitat Use by Bats (Chiroptera) Assessed by Means of a Broad-Band Acoustic Method. J. Appl. Ecol. 1997, 34, 716. [Google Scholar] [CrossRef]
- Hintze, F.; Duro, V.; Carvalho, J.C.; Eira, C.; Rodrigues, P.C.; Vingada, J. Influence ofreservoirs created by small dams on the activity of bats. Acta Chiropt. 2016, 18, 395–408. [Google Scholar] [CrossRef]
- Zeale, M.R.K.; Davidson-Watts, I.; Jones, G. Home range use and habitat selection by barbastelle bats (Barbastella barbastellus): Implications for conservation. J. Mammal. 2012, 93, 1110–1118. [Google Scholar] [CrossRef] [Green Version]
- Rachwald, A.; Apoznański, G.; Thor, K.; Więcek, M.; Zapart, A. Nursery Roosts Used by Barbastelle Bats, Barbastella barbastellus (Schreber, 1774) (Chiroptera: Vespertilionidae) in European Lowland Mixed Forest Transformed by Spruce Bark Beetle, Ips typographus (Linnaeus, 1758) (Coleoptera: Curculionidae). Forests 2022, 13, 1073. [Google Scholar] [CrossRef]
- Greenaway, F. Advice for the management of flightlines and foraging habitats of the barbastelle bat Barbastella barbastellus. In English Nature Research Reports; English Nature: Peterborough, UK, 2004. [Google Scholar]
- Goerlitz, H.R.; ter Hofstede, H.M.; Zeale, M.R.; Jones, G.; Holderied, M.W. An Aerial-Hawking Bat Uses Stealth Echolocation to Counter Moth Hearing. Curr. Biol. 2010, 20, 1568–1572. [Google Scholar] [CrossRef] [Green Version]
- Carr, A.; Zeale, M.R.K.; Weatherall, A.; Froidevaux, J.S.P.; Jones, G. Ground-based and LiDAR-derived measurements reveal scale-dependent selection of roost characteristics by the rare tree-dwelling bat Barbastella barbastellus. For. Ecol. Manag. 2018, 417, 237–246. [Google Scholar] [CrossRef] [Green Version]
- Warren, R.D.; Waters, D.A.; Altringham, J.D.; Bullock, D.J. The distribution of Daubenton’s bats (Myotis daubentonii) and pipistrelle bats (Pipistrellus pipistrellus (Vespertilionidae) in relation to small-scale variation in riverine habitat. Biol. Conserv. 2000, 92, 85–91. [Google Scholar] [CrossRef]
- Russo, D.; Jones, G. Use of foraging habitats by bats in a Mediterranean area determined by acoustic surveys: Conservation implications. Ecography 2003, 26, 197–209. [Google Scholar] [CrossRef]
- Vesterinen, E.J.; Ruokolainen, L.; Wahlberg, N.; Peña, C.; Roslin, T.; Laine, V.N.; Vasko, V.; Sääksjärvi, I.E.; Norrdahl, K.; Lilley, T.M. What you need is what you eat? Prey selection by the bat Myotis daubentonii. Mol. Ecol. 2016, 25, 1581–1594. [Google Scholar] [CrossRef] [PubMed]
- Stewart, D.A.B.; Samways, M.J. 1988. Conserving Dragonfly (Odonata) Assemblages Relative to River Dynamics in an African Savanna Game Reserve. Conserv. Biol. 1988, 12, 683–692. [Google Scholar] [CrossRef]
- Foote, A.L.; Rice Hornung, C.L. Odonates as biological indicators of grazing effects on Canadian prairie wetlands. Ecol. Entomol. 2005, 30, 273–283. [Google Scholar] [CrossRef]
- Burton, E.C.; Gray, M.J.; Schmutzer, A.C.; Miller, D.L. Differential Responses of Postmetamorphic Amphibians to Cattle Grazing in Wetlands. J. Wildl. Manag. 2009, 73, 269–277. [Google Scholar] [CrossRef]
- Arntzen, J.W.; Abrahams, C.; Meilink, W.R.; Iosif, R.; Zuiderwijk, A. Amphibian decline, pond loss and reduced population connectivity under agricultural intensification over a 38 year period. Biodivers. Conserv. 2017, 26, 1411–1430. [Google Scholar] [CrossRef] [Green Version]
- Weather Online. Available online: www.weatheronline.pl (accessed on 1 January 2023).








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thor, K.A.; Ołdak, K.A.; Klich, D.; Gajewska, K.; Popczyk, B.; Klimaszewski, K.; Olech, W. Artificial Waterholes for European Bison as Biodiversity Hotspots in Forest Ecosystems: Ecological Effects of Species Reintroduction Activities. Diversity 2023, 15, 446. https://doi.org/10.3390/d15030446
Thor KA, Ołdak KA, Klich D, Gajewska K, Popczyk B, Klimaszewski K, Olech W. Artificial Waterholes for European Bison as Biodiversity Hotspots in Forest Ecosystems: Ecological Effects of Species Reintroduction Activities. Diversity. 2023; 15(3):446. https://doi.org/10.3390/d15030446
Chicago/Turabian StyleThor, Katarzyna Anna, Krystian Adam Ołdak, Daniel Klich, Karolina Gajewska, Bartłomiej Popczyk, Krzysztof Klimaszewski, and Wanda Olech. 2023. "Artificial Waterholes for European Bison as Biodiversity Hotspots in Forest Ecosystems: Ecological Effects of Species Reintroduction Activities" Diversity 15, no. 3: 446. https://doi.org/10.3390/d15030446
APA StyleThor, K. A., Ołdak, K. A., Klich, D., Gajewska, K., Popczyk, B., Klimaszewski, K., & Olech, W. (2023). Artificial Waterholes for European Bison as Biodiversity Hotspots in Forest Ecosystems: Ecological Effects of Species Reintroduction Activities. Diversity, 15(3), 446. https://doi.org/10.3390/d15030446