
Analysis of the Genetic Diversity of the European Bison (Bison bonasus) Population in Lithuania

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
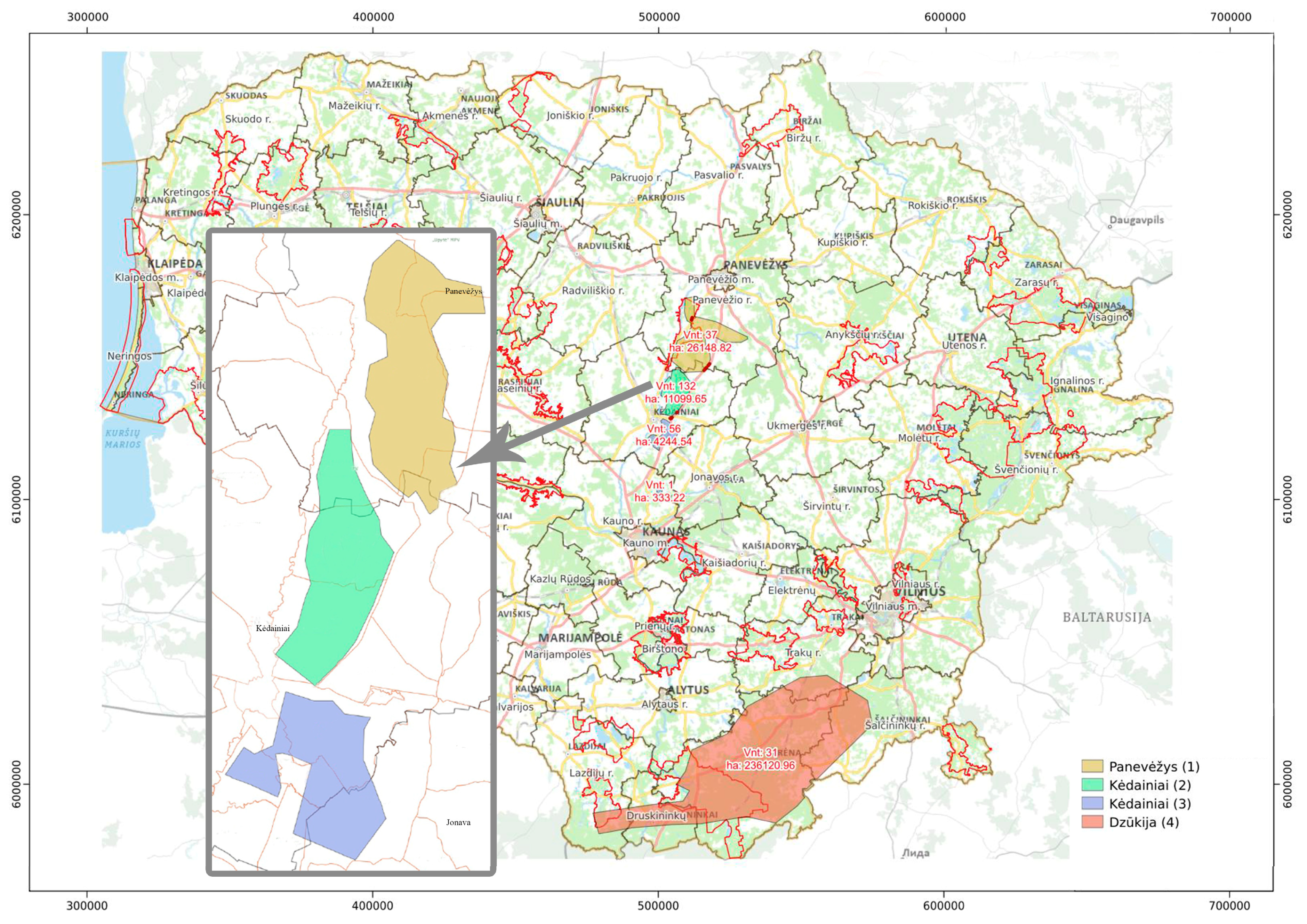
2.1. Sample Collection and DNA Isolation
2.2. Microsatellite Analysis
2.3. mtDNA Analysis
2.4. Data Analysis
3. Results
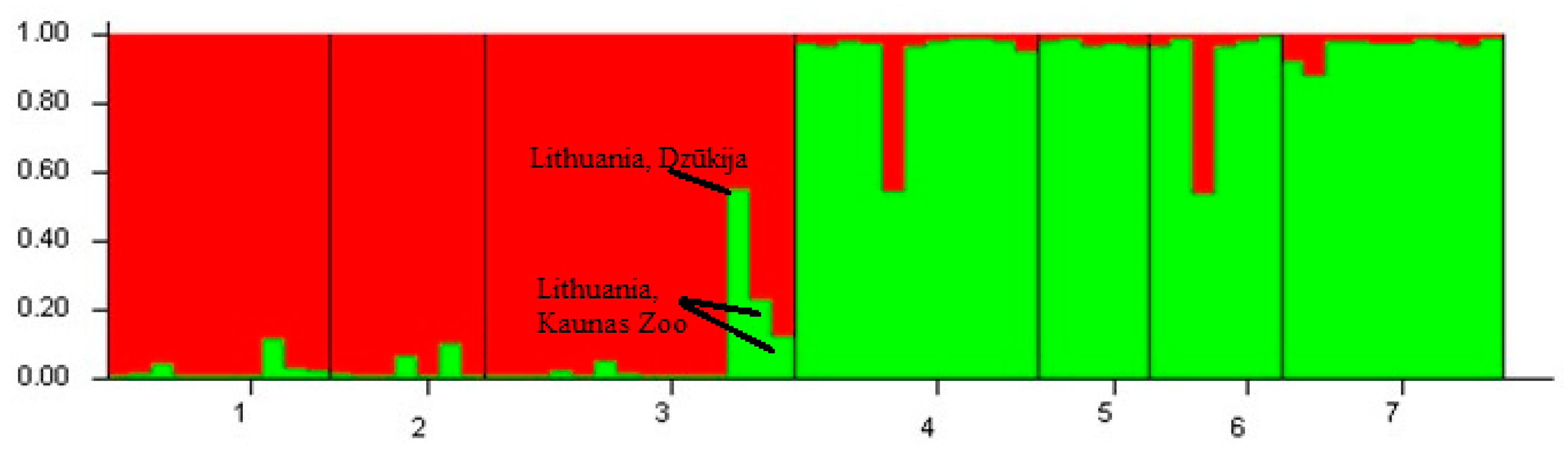
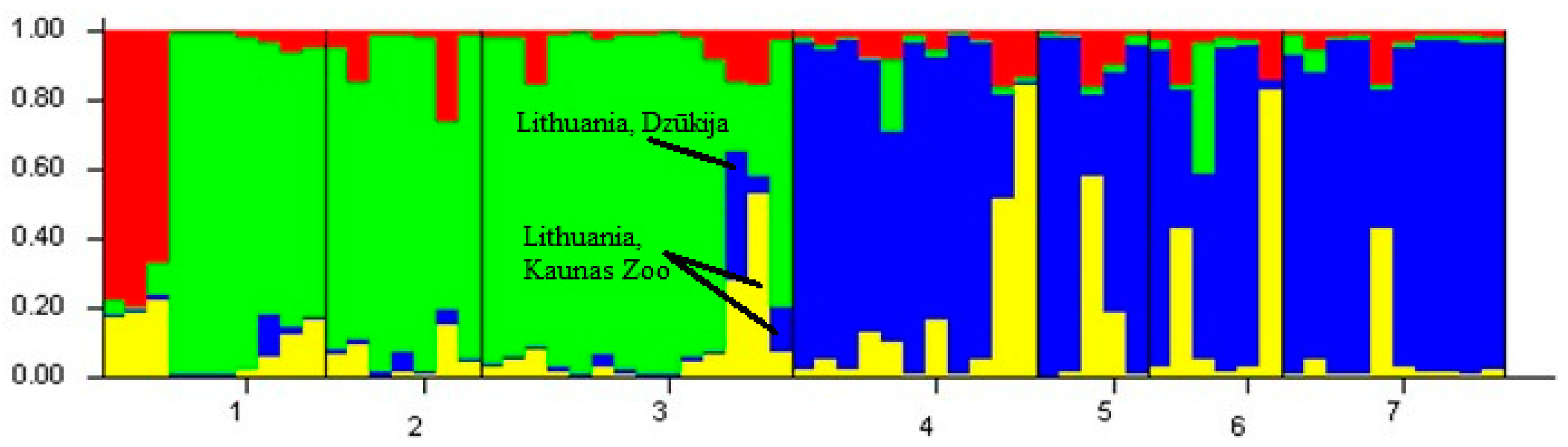
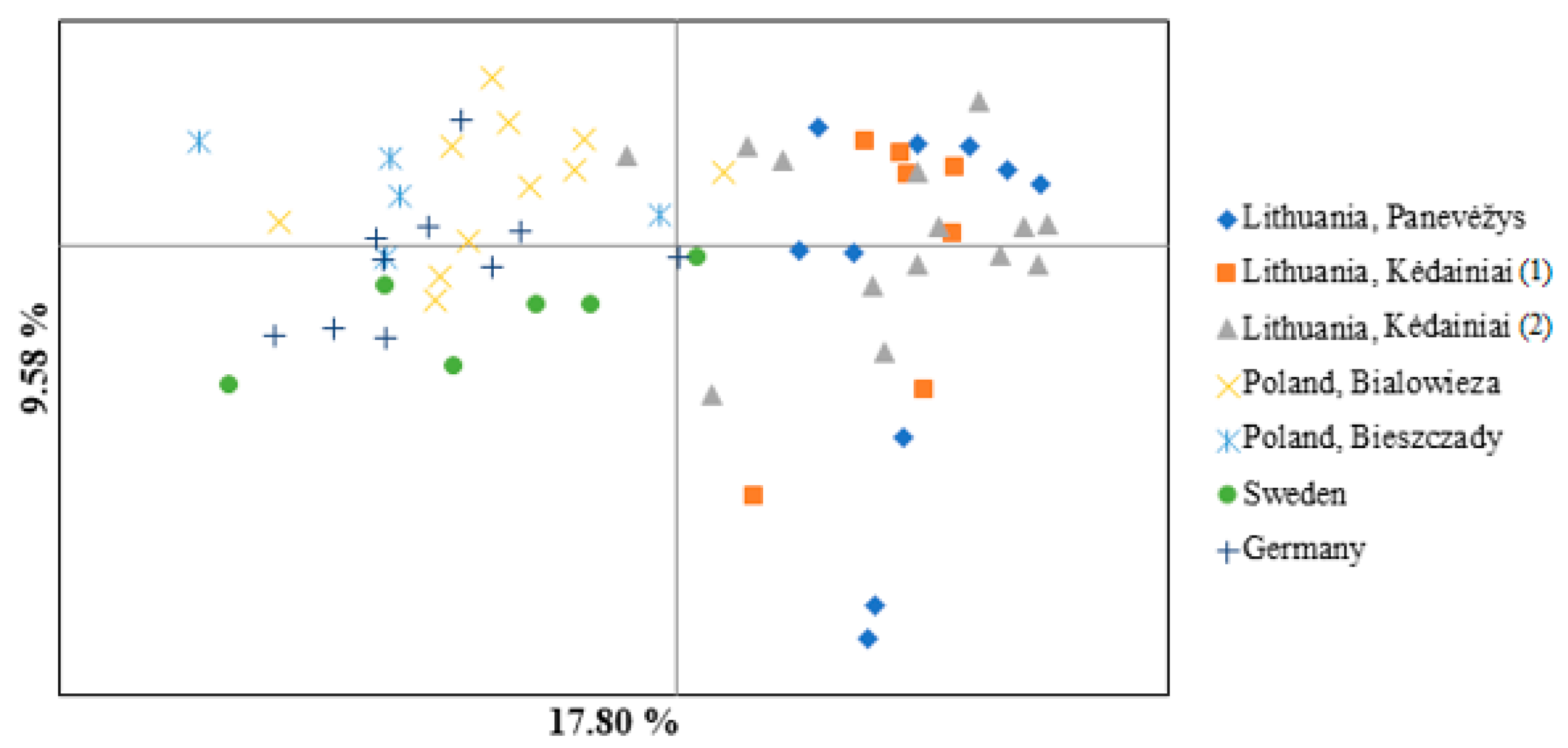
3.1. Microsatellite Genetic Diversity and Genetic Structure
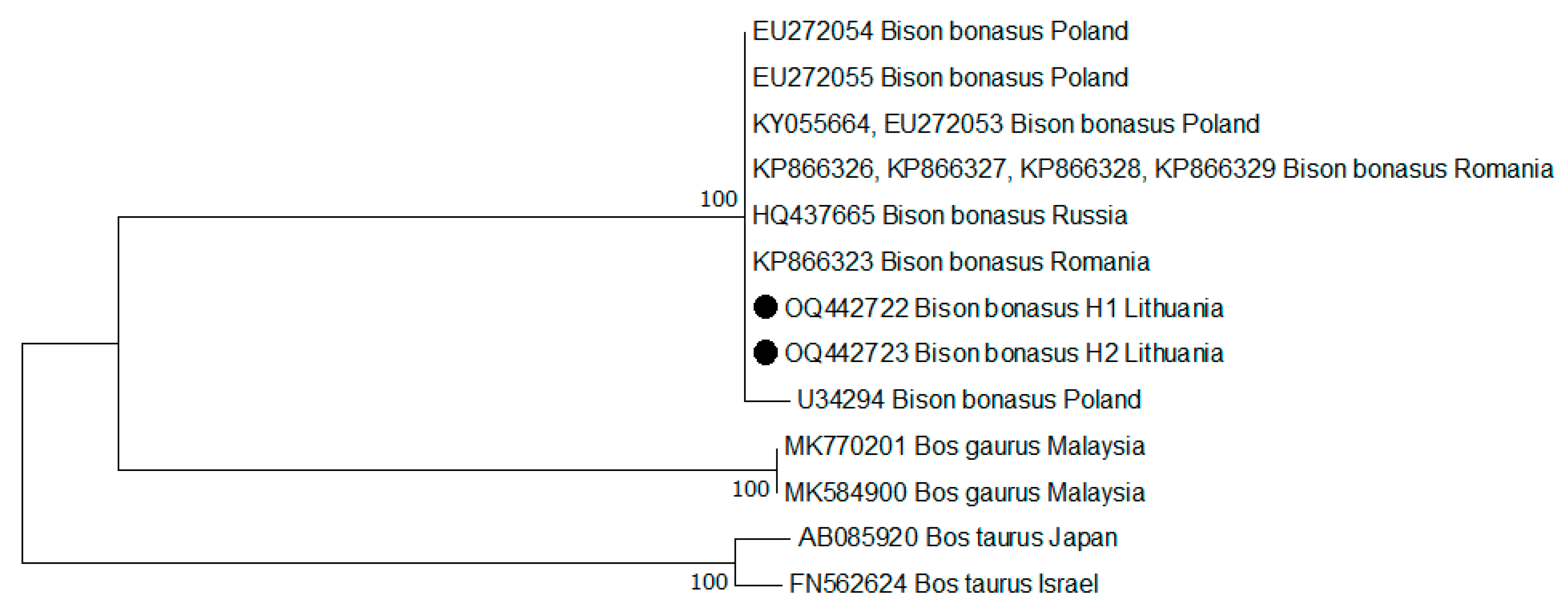
3.2. Genetic Variation Based on mtDNA
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pucek, Z.; Belousove, I.P.; Krasińska, M.; Krasiński, Z.A.; Olech, W. European Bison. Status Survey and Conservation Action Plan; IUCN: Gland, Switzerland, 2004. [Google Scholar]
- Krasińska, M.; Krasiński, Z.A. European Bison: The Nature Monograph; Mammal Research Institute Polish Academy of Sciences: Białowieża, Poland, 2007. [Google Scholar]
- Slatis, H.M. An analysis of inbreeding in the European bison. Genetics 1960, 45, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Olech, W. The number of ancestors and their contribution to European bison (Bison bonasus L.) population. Ann. Wars. Agric. University. Anim. Sci. 1999, 35, 111–117. [Google Scholar]
- Olech, W. The changes of founders’ numbers and their contribution to the European bison population during 80 years of species’ restitution. Eur. Bison Conserv. Newsl. 2009, 2, 54–60. [Google Scholar]
- Plumb, G.; Kowalczyk, R.; Hernandez-Blanco, J.A. Bison bonasus. The IUCN Red List of Threatened Species 2020: e.T2814A45156279. Available online: https://www.iucnredlist.org/species/2814/45156279 (accessed on 7 October 2022).
- Raczyński, J.; Bolbot, M. European Bison Pedigree Book 2020; Białowieski Park Narodowy: Białowieża, Poland, 2020. [Google Scholar]
- Olech, W. The genetic variability within Bison bonasus species 90 years after bottleneck. In Restoration of Endangered and Extinct Animals; Słomski, R., Ed.; Poznan University of Life Sciences: Poznan, Poland, 2010; pp. 48–57. [Google Scholar]
- Wojciechowska, M.; Nowak, Z.; Gurgul, A.; Olech, W.; Drobik, W.; Szmatoła, T. Panel of informative SNP markers for two genetic lines of European bison: Lowland and lowland-caucasian. Anim. Biodivers. Conserv. 2017, 40, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Olech, W.; Klich, D.; Perzanowski, K. An approach towards an improvement of genetic structure of a wisent (Bison bonasus) population in the Carpathians. Eur. Bison Conserv. Newsl. 2021, 13, 11–20. [Google Scholar]
- Tokarska, M.; Pertoldi, C.; Kowalczyk, R.; Perzanowski, K. Genetic status of the European bison Bison bonasus after extinction in the wild and subsequent recovery. Mamm. Rev. 2011, 41, 151–162. [Google Scholar] [CrossRef]
- Tokarska, M.; Bunevich, A.N.; Demontis, D.; Sipko, T.; Perzanowski, K.; Baryshnikov, G.; Kowalczyk, R.; Voitukhovskaya, Y.; Wójcik, J.M.; Marczuk, B.; et al. Genes of the extinct Caucasian bison still roam the Białowieza Forest and are the source of genetic discrepances between Polish and Belarusian populations of the European bison, Bison bonasus. Biol. J. Linn. Soc. 2015, 114, 752–763. [Google Scholar] [CrossRef] [Green Version]
- Balčiauskas, L. European Bison (Bison Bonasus) in Lithuania: Status and possibilities of range extension. Acta Zool. Litu. 1999, 9, 3–18. [Google Scholar] [CrossRef]
- Yanuta, G.; Klich, D.; Balcerak, M. Free living European bison population in Belarus. Eur. Bison Conserv. Newsl. 2021, 13, 57–72. [Google Scholar]
- Kostyunina, O.V.; Mikhailova, M.E.; Dotsev, A.V.; Zemlyanko, I.I.; Volkova, V.V.; Fornara, M.S.; Akopyan, N.A.; Kramarenko, A.S.; Okhlopkov, I.M.; Aksenova, P.V.; et al. Comparative Genetic characteristics of the Russian and Belarusian populations of wisent (Bison bonasus), North American Bison (Bison bison) and Cattle (Bos taurus). Cytol. Genet. 2020, 54, 116–123. [Google Scholar] [CrossRef]
- Daleszczyk, K.; Bunevich, A. Population viability analysis of European bison populations in Polish and Belarusian parts of Bialowieza Forest with and without gene exchange. Biol. Conserv. 2009, 142, 3068–3075. [Google Scholar] [CrossRef]
- Frankham, R.; Ballou, J.D.; Briscoe, D.A. Introduction to Conservation Genetics; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Hartway, C.; Hardy, A.; Jones, L.; Moynahan, B.; Traylor-Holzer, K.; McCann, B.; Aune, K.; Plumb, G. Long-Term Viability of Department of the Interior Bison under Current Management and Potential Metapopulation Management Strategies; Natural Resource Report NPS/NRSS/BRD—2020/2097; National Park Service: Fort Collins, CO, USA, 2020.
- Davies, G.; McCann, B.; Jones, L.; Liccioli, S.; Penedo, M.C.; Ovchinnikov, I.V. Genetic variation of the mitochondrial DNA control region across plains bison herds in USA and Canada. PLoS ONE 2022, 17, e0264823. [Google Scholar] [CrossRef] [PubMed]
- De Barba, M.; Miquel, C.; Lobréaux, S.; Quenette, P.Y.; Swenson, J.E.; Taberlet, P. High-throughput microsatellite genotyping in ecology: Improved accuracy, efficiency, standardization and success with low-quantity and degraded DNA. Mol. Ecol. Resour. 2017, 17, 492–507. [Google Scholar] [CrossRef] [PubMed]
- Demir, E.; Karsli, T.; Balcioğlu, M.S. A comprehensive review on genetic diversity and phylogenetic relationships among native Turkish cattle breeds based on microsatellite markers. Turk. J. Vet. Anim. Sci. 2021, 45, 1–10. [Google Scholar] [CrossRef]
- Wójcik, J.M.; Kawałko, A.; Tokarska, M.; Jaarola, M.; Vallenback, P.; Pertoldi, C. Post-bottleneck mtDNA diversity in a free-living population of European bison: Implications for conservation. J. Zool. 2009, 277, 81–87. [Google Scholar] [CrossRef]
- FAO. Molecular Genetic Characterization of Animal Genetic Resources. FAO Animal Production and Health Guidelines. 2011. Available online: https://www.fao.org/3/i2413e/i2413e00.pdf (accessed on 7 October 2022).
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Earl, D.A.; vonHoldt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large datasets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Kowalczyk, R.; Schmidt, K.; Jedrzejewski, W. Do fences or humans inhibit the movements of large mammals in Białowieża Primeval Forest? In Fencing for Conservation; Somers, M.J., Hayward, M.W., Eds.; Springer: New York, NY, USA, 2012; pp. 235–244. [Google Scholar]
- Homel, K.V.; Śliwińska, K.; Valnisty, A.A.; Nikiforov, M.E. New data on the genetic diversity of European bison Bison bonasus (Linnaeus, 1758) in Belarus. Theriol. Ukr. 2020, 19, 45–53. [Google Scholar] [CrossRef]
- Roth, T.; Pfeiffer, I.; Weising, K.; Brenig, B. Application of bovine microsatellite markers for genetic diversity analysis of European bison (Bison bonasus). J. Anim. Breed. Genet. 2006, 123, 406–409. [Google Scholar] [CrossRef] [PubMed]
- Tokarska, M.; Kawałko, A.; Wójcik, J.M.; Pertoldi, C. Genetic variability in the European bison (Bison bonasus) population from Białowieża forest over 50 years. Biol. J. Linn. Soc. 2009, 97, 801–809. [Google Scholar] [CrossRef] [Green Version]
- Tokarska, M.; Marshall, T.; Kowalczyk, R.; Wójcik, J.M.; Pertoldi, C.; Kristensen, T.N.; Loeschcke, V.; Gregersen, V.R.; Bendixen, C. Effectiveness of microsatellite and SNP markers for parentage and identity analysis in species with low genetic diversity: The case of European bison. Heredity 2009, 103, 326–332. [Google Scholar] [CrossRef] [Green Version]
- Machová, K.; Štruncová, P.; Calta, J.; Tichý, L.; Vostrý, L. Genealogical analysis of European bison population revealed a growing up population despite very low genetic diversity. PLoS ONE 2022, 17, e0277456. [Google Scholar] [CrossRef]
- Larska, M.; Krzysiak, M.K. Infectious disease monitoring of European bison (Bison bonasus). In Wildlife Population Monitoring; Ferretti, M., Ed.; IntechOpen: London, UK, 2019; pp. 248–269. [Google Scholar]
- Gralak, B.; Krasińska, M.; Niemczewski, C.; Krasiński, Z.A.; Żurkowski, M. Polymorphism of bovine microsatellite DNA sequences in the lowland European bison. Acta Theriol. 2004, 49, 449–456. [Google Scholar] [CrossRef]
- Luenser, K.; Fickel, J.; Lehnen, A.; Speck, S.; Ludwig, A. Low level of genetic variability in European bisons (Bison bonasus) from the Bialowieza National Park in Poland. Eur. J. Wildl. Res. 2005, 51, 84–87. [Google Scholar] [CrossRef]
- Wojciechowska, M.; Puchała, K.; Nowak-Życzyńska, Z.; Perlińska-Teresiak, M.; Kloch, M.; Drobik-Czwarno, W.; Olech, W. From wisent to the lab and back again—A complex SNP set for population management as an effective tool in European Bison conservation. Diversity 2023, 15, 116. [Google Scholar] [CrossRef]
- Yudin, N.S.; Kulikov, I.V.; Gunbin, K.V.; Aitnazarov, R.B.; Kushnir, A.V.; Sipko, T.P.; Moshkin, M.P. Detection of mitochondrial DNA from domestic cattle in European bison (Bison bonasus) from the Altai Republic in Russia. Anim. Genet. 2012, 43, 362. [Google Scholar] [CrossRef]
- Cronin, M.A.; MacNeil, M.D.; Vu, N.; Leesburg, V.; Blackburn, H.D.; Derr, J.D. Genetic variation and differentiation of bison (Bison bison) subspecies and cattle (Bos taurus) breeds and subspecies. J. Hered. 2013, 104, 500–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herman, J.A.; Piaggio, A.J.; Halbert, N.D.; Rhyan, J.C.; Salman, M.D. Genetic analysis of a Bison bison herd derived from the Yellowstone National Park population. Wildl. Biol. 2014, 20, 335–343. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | Forward Primer Sequence (5′-3′) | Reverse Primer Sequence (5′-3′) | Fluorescent Dye |
|---|---|---|---|
| BOVIRBP | 5′-GTATGATCACCTTCTATGCTTC-3′ | 5′-GCTTTAGGTAATCATCAGATAGC-3′ | 6FAM |
| BTJAB1 | 5′-CATTAAGGGCTGGGATTCCT-3′ | 5′-AGATTTCTGGAGGAGGCTCACAGCA-3′ | 6FAM |
| BM6438 | 5′-TTGAGCACAGACACAGACTGG-3′ | 5′-ACTGAATGCCTCCTTTGTGC-3′ | 6FAM |
| BM2830 | 5′- ATGGGCGTATAAACACAGATG-3′ | 5′-TGAGTCCTGTCACCATCAGC-3′ | CY3 |
| TGLA122 | 5′-CCCTCCTCCAGGTAAATCAGC-3′ | 5′-AATCACATGGCAAATAAGTACATAC-3′ | 6FAM |
| ETH10 | 5′-GTTCAGGACTGGCCCTGCTAACA-3′ | 5′-CCTCCAGCCCACTTTCTCTTCTC-3′ | 6FAM |
| BM1225 | 5′-TTTCTCAACAGAGGTGTCCAC-3′ | 5′-ACCCCTATCACCATGCTCTG-3′ | 6FAM |
| BM1818 | 5′-AGCTGGGAATATAACCAAAGG-3′ | 5′-AAGTGCTTTCAAGGTCCATGC-3′ | CY3 |
| TGLA53 | 5′-GCTTTCAGAAATAGTTTGCATTCA-3′ | 5′-ATCTTCACATGATATTACAGCAGA-3′ | CY3 |
| BM723 | 5′-ACCCTTGGTTTTCTGCTGG-3′ | 5′-CATCCTGTGTGAGTGTTGTGG-3′ | 6FAM |
| ETH121 | 5′-CCAACTCCTTACAGCAAATGTC-3′ | 5′-ATTTAGAGCTGGCTGGTAAGTG-3′ | 6FAM |
| HEL9 | 5′-CCCATTCAGTCTTCAGAGGT-3′ | 5′-CACATCCATGTTCTCACCAC-3′ | 6FAM |
| TGLA227 | 5′-CGAATTCCAAATCTGTTAATTTGCT-3′ | 5′-ACAGACAGAAACTCAATGAAAGCA′-3′ | CY3 |
| Loci | Allele | Lithuania | Poland (Białowieża) | Poland (Bieszczady) | Sweden | Germany |
|---|---|---|---|---|---|---|
| BM6438 | 262 | 0.190 | 0 | 0 | 0 | 0 |
| 264 | 0.776 | 0.727 | 0.400 | 0.500 | 0.500 | |
| 274 | 0.034 | 0.273 | 0.600 | 0.500 | 0.500 | |
| BM723 | 146 | 0.371 | 0 | 0 | 0 | 0 |
| 148 | 0.629 | 0.864 | 0.600 | 0.667 | 0.650 | |
| 152 | 0 | 0.136 | 0.400 | 0.333 | 0.350 | |
| ETH121 | 186 | 0.050 | 0.091 | 0.100 | 0.500 | 0.350 |
| 192 | 0.633 | 0.909 | 0.900 | 0.417 | 0.650 | |
| 194 | 0 | 0 | 0 | 0.083 | 0 | |
| 204 | 0.100 | 0 | 0 | 0 | 0 | |
| 214 | 0.217 | 0 | 0 | 0 | 0 |
| Population | N | Na | Ho | He | Fis | |
|---|---|---|---|---|---|---|
| Lithuania | 31 | Mean | 2.91 | 0.32 | 0.34 | −0.038 |
| SE | 0.59 | 0.05 | 0.05 | 0.102 | ||
| Poland, Białowieża | 11 | Mean | 2.21 | 0.23 | 0.24 | −0.148 |
| SE | 0.29 | 0.06 | 0.04 | 0.111 | ||
| Poland, Bieszczady | 5 | Mean | 2.10 | 0.25 | 0.27 | −0.122 |
| SE | 0.26 | 0.05 | 0.03 | 0.077 | ||
| Sweden | 6 | Mean | 2.23 | 0.27 | 0.25 | 0.026 |
| SE | 0.20 | 0.05 | 0.03 | 0.100 | ||
| Germany | 10 | Mean | 2.39 | 0.30 | 0.30 | −0.098 |
| SE | 0.26 | 0.05 | 0.05 | 0.095 | ||
| Total | 63 | Weighted mean | 2.58 | 0.29 | 0.30 | −0.076 |
| SE | 0.14 | 0.02 | 0.02 | 0.043 |
| Haplotype | GenBank Accession Numbers | Nucleotide Position | Country | ||||
|---|---|---|---|---|---|---|---|
| 35 | 355 | 381 | 382 | 465 | |||
| H1 | OQ442722 | − | − | C | − | C | Lithuania |
| KP866326, KP866327, KP866328, KP866329 | Romania | ||||||
| KY055664 EU272053 | Poland | ||||||
| HQ437665 | Russia | ||||||
| H2 | OQ442723 | − | − | − | − | ∙ | Lithuania |
| EU272054 | Poland | ||||||
| H3 | EU272055 | − | − | ∙ | C | ∙ | Poland |
| H4 | U34294 | − | G | ∙ | − | T | Poland |
| H5 | KP866323 | A | − | ∙ | − | ∙ | Romania |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lapickis, R.; Griciuvienė, L.; Kibiša, A.; Lipatova, I.; Aleksandravičienė, A.; Ražanskė, I.; Wojciechowska, M.; Kloch, M.; Olech, W.; Paulauskas, A. Analysis of the Genetic Diversity of the European Bison (Bison bonasus) Population in Lithuania. Diversity 2023, 15, 406. https://doi.org/10.3390/d15030406
Lapickis R, Griciuvienė L, Kibiša A, Lipatova I, Aleksandravičienė A, Ražanskė I, Wojciechowska M, Kloch M, Olech W, Paulauskas A. Analysis of the Genetic Diversity of the European Bison (Bison bonasus) Population in Lithuania. Diversity. 2023; 15(3):406. https://doi.org/10.3390/d15030406
Chicago/Turabian StyleLapickis, Romualdas, Loreta Griciuvienė, Artūras Kibiša, Indrė Lipatova, Asta Aleksandravičienė, Irma Ražanskė, Marlena Wojciechowska, Marta Kloch, Wanda Olech, and Algimantas Paulauskas. 2023. "Analysis of the Genetic Diversity of the European Bison (Bison bonasus) Population in Lithuania" Diversity 15, no. 3: 406. https://doi.org/10.3390/d15030406
APA StyleLapickis, R., Griciuvienė, L., Kibiša, A., Lipatova, I., Aleksandravičienė, A., Ražanskė, I., Wojciechowska, M., Kloch, M., Olech, W., & Paulauskas, A. (2023). Analysis of the Genetic Diversity of the European Bison (Bison bonasus) Population in Lithuania. Diversity, 15(3), 406. https://doi.org/10.3390/d15030406