
First Stranding Event of a Common Minke Whale (Balaenoptera acutorostrata Lacépède, 1804) Reported in the Gulf of Thailand


 , and
, and
Abstract
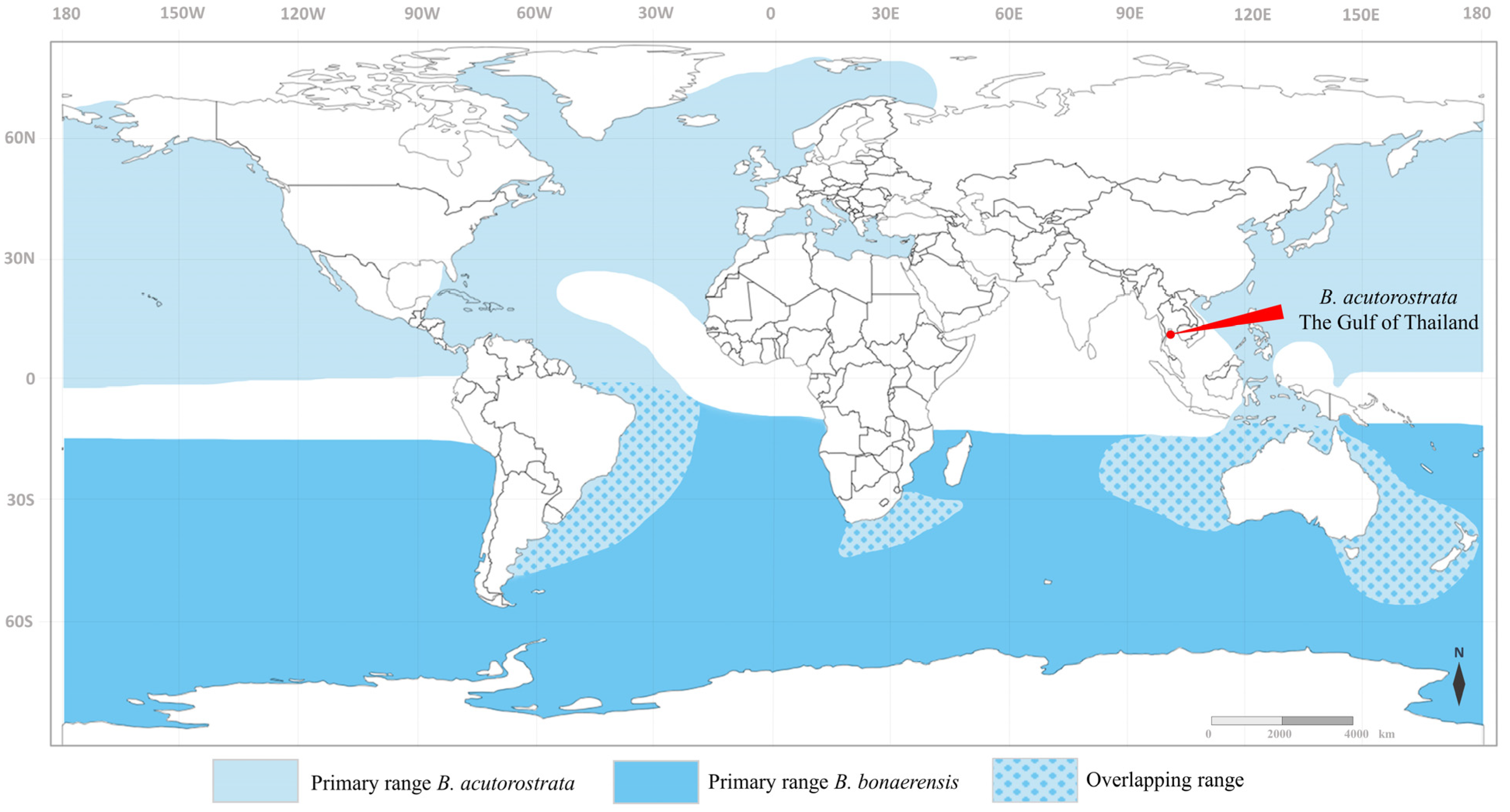
:1. Introduction
2. Materials and Methods
3. Results
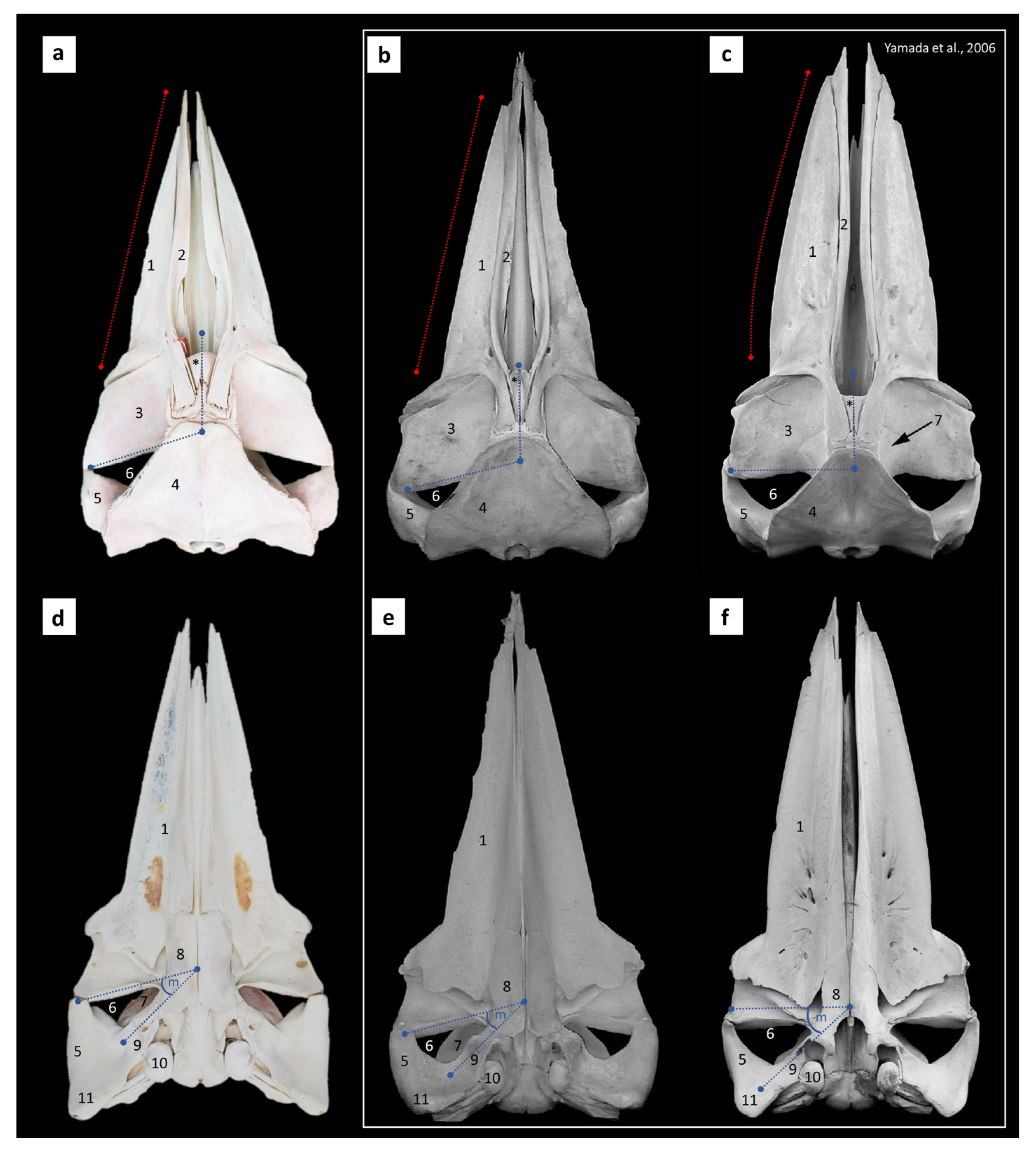
3.1. Cranial Bone Morphology
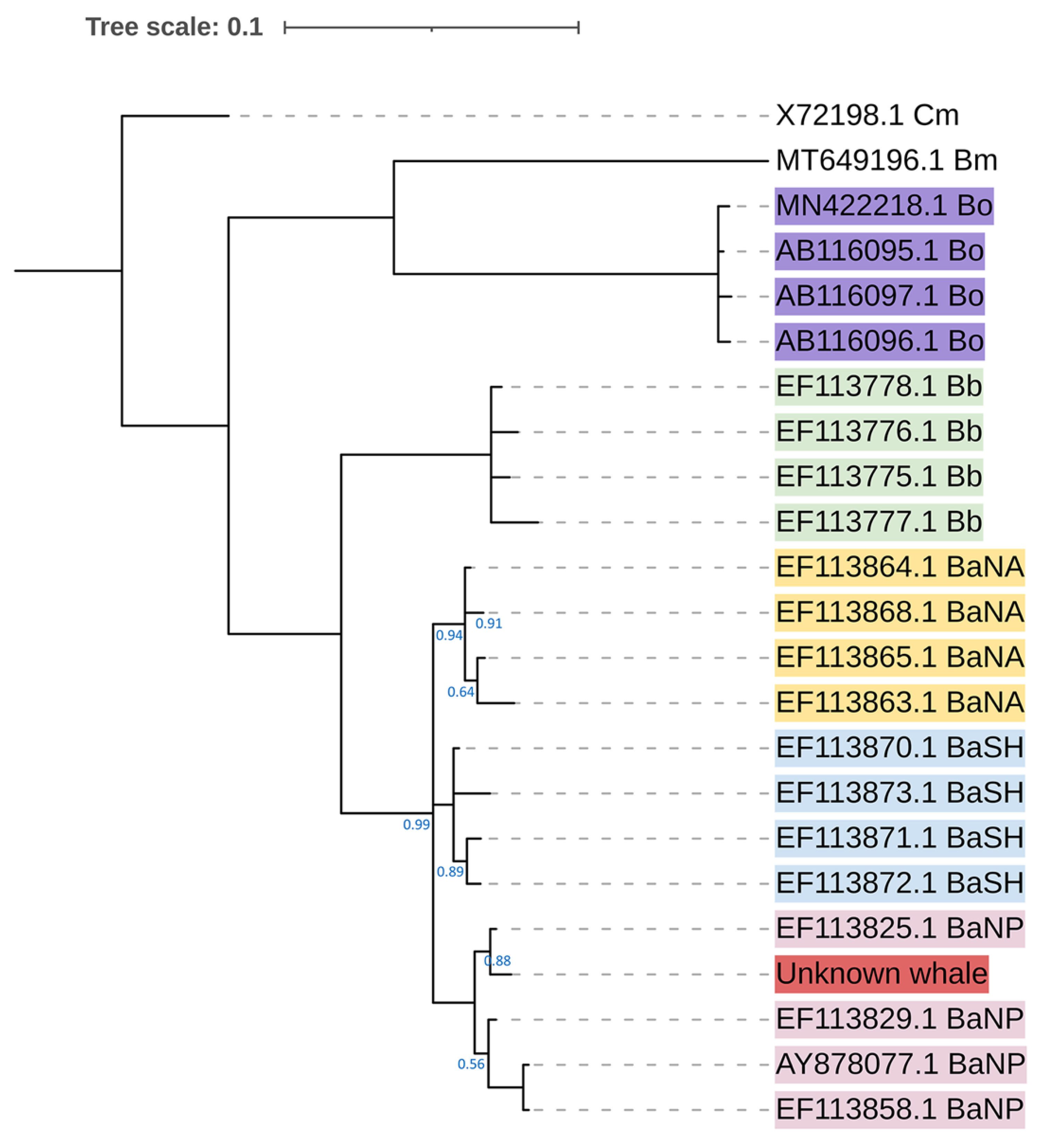
3.2. The Sequence of D-Loop and Phylogenetic Tree
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jefferson, T.; Webber, M.; Pitman, R. Marine Mammals of the World: A Comprehensive Guide to Their Identification; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Dold, C. Cetacea (Whales, Dolphins, Porpoises). Fowler’s Zoo Wild Anim. Med. 2014, 8, 422. [Google Scholar]
- Chantrapornsyl, S.; Adulyanukosol, K.; Kittiwathanawong, K. Records of cetaceans in Thailand. Phuket Mar. Biol. Cent. Res. Bull. 1996, 61, 39–63. [Google Scholar]
- Department of Marine and Coastal Resources. “Whale and Dolphin Species”. Central Database System and Data Standard for Marine and Coastal Resources. 2013. Available online: http://km.dmcr.go.th/th/c_7/d_2448 (accessed on 13 December 2022).
- Hines, E.; Ponnampalam, L.; Jamal Hisne, F.; Tara, S.; Jackson-Ricketts, J.; Kuit, S.; Acebes, J. Report of the Third Southeast Asian Marine Mammal Symposium (SEAMAM III). CMS Tech. Ser. 2015, 32, 105–109. [Google Scholar]
- Kanjana, A.; Surasak, T.; Theerawat, P.; Phaothep, C. Field Guide to Marine Mammals and Sea Turtles of Thailand; Marine and Coastal Resources Research Center, Upper Gulf of Thailand, National Office of Buddhism: Nakhon Pathom, Thailand, 2014.
- Roman, J.; Estes, J.A.; Morissette, L.; Smith, C.; Costa, D.; McCarthy, J.; Nation, J.; Nicol, S.; Pershing, A.; Smetacek, V. Whales as marine ecosystem engineers. Front. Ecol. Environ. 2014, 12, 377–385. [Google Scholar] [CrossRef] [Green Version]
- Department of National Parks Wildlife and Plant Conservation. “Wildlife Conservation and Protection Act, B.E. 2562 (2019)”. Thai Government Gazette. 2019. Available online: http://portal.dnp.go.th/Law (accessed on 28 April 2021).
- Segre, P.S.; Gough, W.T.; Roualdes, E.A.; Cade, D.E.; Czapanskiy, M.F.; Fahlbusch, J.; Kahane-Rapport, S.R.; Oestreich, W.K.; Bejder, L.; Bierlich, K. Scaling of maneuvering performance in baleen whales: Larger whales outperform expectations. J. Exp. Biol. 2022, 225, jeb243224. [Google Scholar] [CrossRef]
- Pastene, L.A.; Acevedo, J.; Goto, M.; Zerbini, A.N.; Acuna, P.; Aguayo-Lobo, A. Population structure and possible migratory links of common minke whales, Balaenoptera acutorostrata, in the Southern Hemisphere. Conserv. Genet. 2010, 11, 1553–1558. [Google Scholar] [CrossRef]
- Glover, K.A.; Kanda, N.; Haug, T.; Pastene, L.A.; Øien, N.; Seliussen, B.B.; Sørvik, A.G.; Skaug, H.J. Hybrids between common and Antarctic minke whales are fertile and can back-cross. Bmc Genet. 2013, 14, 25. [Google Scholar] [CrossRef] [Green Version]
- Pastene, L.A.; Goto, M.; Kanda, N.; Zerbini, A.N.; Kerem, D.; Watanabe, K.; Bessho, Y.; Hasegawa, M.; Nielsen, R.; Larsen, F. Radiation and speciation of pelagic organisms during periods of global warming: The case of the common minke whale, Balaenoptera acutorostrata. Mol. Ecol. 2007, 16, 1481–1495. [Google Scholar] [CrossRef]
- Arnold, P.W.; Birtles, R.A.; Dunstan, A.; Lukoschek, V.; Matthews, M. Colour patterns of the dwarf minke whale Balaenoptera acutorostrata sensu lato: Description, cladistic analysis and taxonomic implications. Mem. Qld. Mus. 2005, 51, 277–307. [Google Scholar]
- Perrin, W.F.; Mallette, S.D.; Brownell, R.L., Jr. Minke whales: Balaenoptera acutorostrata and B. bonaerensis. In Encyclopedia of Marine Mammals; Elsevier: Amsterdam, The Netherlands, 2018; pp. 608–613. [Google Scholar]
- Fraija-Fernández, N.; Crespo-Picazo, J.L.; Domènech, F.; Míguez-Lozano, R.; Palacios-Abella, J.F.; Rodríguez-González, A.; Villar-Torres, M.; Gozalbes, P. First stranding event of a common minke whale calf, Balaenoptera acutorostrata Lacépède, 1804, reported in Spanish Mediterranean waters. Mammal Study 2015, 40, 95–100. [Google Scholar] [CrossRef]
- Öztürk, A.A.; Tonay, A.M.; Dede, A.; Ayhan, D. Strandings of the beaked whales, Risso’s dolphins, and a minke whale on the Turkish coast of the Eastern Mediterranean Sea. J. Black Sea/Mediterr. Environ. 2011, 17, 269–274. [Google Scholar]
- Rudolph, P.; Smeenk, C. Indo-West Pacific marine mammals. In Encyclopedia of Marine Mammals; Elsevier: Amsterdam, The Netherlands, 2009; pp. 608–616. [Google Scholar]
- Zhao, L.; Zhu, Q.; Miao, X.; Xu, M.; Wu, F.; Dai, Y.; Tao, C.; Mou, J.; Wang, X. An overview of cetacean strandings, bycatches and rescues along the western coast of the Taiwan Strait, China: 2010–2015. Acta Oceanol. Sin. 2017, 36, 31–36. [Google Scholar] [CrossRef]
- Lee, D.; An, Y.R.; Park, K.J.; Kim, H.W.; Lee, D.; Joo, H.T.; Oh, Y.G.; Kim, S.M.; Kang, C.K.; Lee, S.H. Spatial distribution of common Minke whale (Balaenoptera acutorostrata) as an indication of a biological hotspot in the East Sea. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2017, 143, 91–99. [Google Scholar] [CrossRef]
- Risch, D.; Norris, T.; Curnock, M.; Friedlaender, A. Common and Antarctic minke whales: Conservation status and future research directions. Front. Mar. Sci. 2019, 6, 247. [Google Scholar] [CrossRef] [Green Version]
- Pastene, L.A.; Goto, M.; Taguchi, M.; Kitakado, T. Temporal and Spatial Distribution of the ‘J’ and ‘O’ Stocks of Common Minke Whale in Waters around Japan Based on Microsatellite DNA; Paper SC/F16/JR38 Presented to the JARPNII Special Permit Expert Panel Review Workshop; Paper SC: Tokyo, Japan, 2016. [Google Scholar]
- Jaaman, S.A.; Najib, R.; Syuhaime, A.; Yuhana, U. Records of Marine Mammals in Peninsular Malaysia: A Review. Tropical Marine Environment: Charting Strategies for the Millennium; Malacca Straits Research and Development Centre (MASDEC), Universiti Putra Malaysia: Serdang, Malaysia, 2002; pp. 499–515. [Google Scholar]
- Mazlan, A.; Zaidi, C.; Wan-Lotfi, W.; Othman, B. On the Current Status of Coastal Marine Biodiversity in Malaysia. Indian J. Mar. Sci. 2005, 34, 76–87. [Google Scholar]
- Smith, B.D.; Jefferson, T.A.; Leatherwood, S.; Ho, D.; Thuoc, C.V.; Quang, L.H. Investigations of marine mammals in Vietnam. Asian Mar. Biol. 1997, 14, 145–172. [Google Scholar]
- Geraci, J.R.; Lounsbury, V.J. Marine Mammals Ashore: A Field Guide for Strandings; National Aquarium in Baltimore: Baltimore, MD, USA, 2005. [Google Scholar]
- Pugliares, K.R.; Bogomolni, A.; Touhey, K.M.; Herzig, S.M.; Harry, C.T. Marine Mammal Necropsy: An Introductory Guide for Stranding Responders and Field Biologists; MBLWHOI Library: Woods Hole, MA, USA, 2007. [Google Scholar]
- Yamada, T.K.; Chou, L.-S.; Chantrapornsyl, S.; Adulyanukosol, K.; Chakravarti, S.K.; Oishi, M.; Wada, S.; Yao, C.-J.; Kakuda, T.; Tajima, Y. Middle-sized balaenopterid whale specimens (Cetacea: Balaenopteridae) preserved at several institutions in Taiwan, Thailand, and India. Mem. Natl. Sci.Mus. 2006, 44, 1–10. [Google Scholar]
- Dalebout, M.; Baker, C.; Mead, J.; Cockcroft, V.; Yamada, T. A comprehensive and validated molecular taxonomy of beaked whales, family Ziphiidae. J. Hered. 2004, 95, 459–473. [Google Scholar] [CrossRef] [Green Version]
- Oremus, M.; Gales, R.; Dalebout, M.L.; Funahashi, N.; Endo, T.; Kage, T.; Steel, D.; Baker, S.C. Worldwide mitochondrial DNA diversity and phylogeography of pilot whales (Globicephala spp.). Biol. J. Linn. Soc. 2009, 98, 729–744. [Google Scholar] [CrossRef] [Green Version]
- Kraft, S.; Pérez-Álvarez, M.; Olavarría, C.; Poulin, E. Global phylogeography and genetic diversity of the long-finned pilot whale Globicephala melas, with new data from the southeastern Pacific. Sci. Rep. 2020, 10, 1769. [Google Scholar] [CrossRef] [Green Version]
- Blair, D.; McMahon, A.; McDonald, B.; Tikel, D.; Waycott, M.; Marsh, H. Pleistocene sea level fluctuations and the phylogeography of the dugong in Australian waters. Mar. Mammal Sci. 2014, 30, 104–121. [Google Scholar] [CrossRef]
- Kriangwanich, W.; Buddhachat, K.; Poommouang, A.; Chomdej, S.; Thitaram, C.; Kaewmong, P.; Kittiwattanawong, K.; Nganvongpanit, K. Feasibility of melting fingerprint obtained from ISSR-HRM curves for marine mammal species identification. PeerJ 2021, 9, e11689. [Google Scholar] [CrossRef]
- Piboon, P.; Kriengsakpichit, N.; Poommouang, A.; Buddhachat, K.; Brown, J.L.; Kampuansai, J.; Chomdej, S.; Kaewmong, P.; Kittiwattanawong, K.; Nganvongpanit, K. Relationship of stranded cetaceans in Thai territorial waters to global populations: Mitochondrial DNA diversity of Cuvier’s beaked whale, Indo Pacific finless porpoise, pygmy sperm whale, and dwarf sperm whale. Sci. Prog. 2022, 105, 00368504221103776. [Google Scholar] [CrossRef]
- Piboon, P.; Poommouang, A.; Buddhachat, K.; Kampuansai, J.; Chomdej, S.; Kaewmong, P.; Kittiwattanawong, K.; Nganvongpanit, K. The Potential Distribution and Maternal Lineage of Two Cetaceans Species (Grampus griseus and Pseudorca crassidens) in the Subfamily Globicephalinae from the Thai Andaman Sea and the Gulf of Thailand. Diversity 2022, 14, 257. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.; Teslenko, M.; Nylander, J. MrBayes Version 3.2 Manual: Tutorials and Model Summaries. 2011. Available online: https://bioweb.pasteur.fr/docs/modules/mrbayes/3.1.2/Manual_MrBayes_v3.2.0_draft.pdf (accessed on 31 December 2022).
- Posada, D. jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Kato, H.; Fujise, Y.; Nakamura, G.; Hakamada, T.; Pastene, L.A.; Best, P.B. Dwarf Minke Whales: Morphology, Growth and Life History Based on Samples Collected from the Higher Latitudes in the Southern Hemisphere. Cetacean Popul. Stud. 2021, 3, 93–128. [Google Scholar]
- Yamato, M.; Ketten, D.R.; Arruda, J.; Cramer, S.; Moore, K. The auditory anatomy of the minke whale (Balaenoptera acutorostrata): A potential fatty sound reception pathway in a baleen whale. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2012, 295, 991–998. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Wu, H.; Wu, C.; Yang, G.; Chen, B. Molecular Identification of Stranded Cetaceans in Coastal China. Aquat. Mamm. 2019, 45, 525–532. [Google Scholar] [CrossRef]
- FalcÃ, L.; Campos, A.; Freitas, J.; Furtado-Neto, M.; Faria, V. Molecular identification of cetaceans from the West Atlantic using the E3-I5 region of COI. Genet. Mol. Res. 2017, 16, 2. [Google Scholar] [CrossRef]
- McGowen, M.R.; Spaulding, M.; Gatesy, J. Divergence date estimation and a comprehensive molecular tree of extant cetaceans. Mol. Phylogenetics Evol. 2009, 53, 891–906. [Google Scholar] [CrossRef]
- Milmann, L.; Taguchi, M.; Siciliano, S.; Baumgarten, J.E.; Oliveira, L.R.; Valiati, V.H.; Goto, M.; Ott, P.H.; Pastene, L.A. New genetic evidences for distinct populations of the common minke whale (Balaenoptera acutorostrata) in the Southern Hemisphere. Polar Biol. 2021, 44, 1575–1589. [Google Scholar] [CrossRef]
- Tulloch, V.J.; Plagányi, É.E.; Brown, C.; Richardson, A.J.; Matear, R. Future recovery of baleen whales is imperiled by climate change. Glob. Change Biol. 2019, 25, 1263–1281. [Google Scholar] [CrossRef] [Green Version]
- MacLeod, C.D. Global climate change, range changes and potential implications for the conservation of marine cetaceans: A review and synthesis. Endanger. Species Res. 2009, 7, 125–136. [Google Scholar] [CrossRef]
- Owen, K.; Jenner, K.C.S.; Jenner, M.-N.M.; McCauley, R.D.; Andrews, R.D. Water temperature correlates with baleen whale foraging behaviour at multiple scales in the Antarctic. Mar. Freshw. Res. 2018, 70, 19–32. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Common Name | Scientific Name |
|---|---|---|
| Balaenopteridae | Blue whale | Balaenoptera musculus |
| Bryde’s whale | Balaenoptera edeni | |
| Fin whale | Balaenoptera physalus | |
| Humpback whale | Megaptera novaeangliae | |
| Omura’s whale | Balaenoptera omurai | |
| Delphinidae | False killer whale | Pseudorca crassidens |
| Fraser’s dolphin | Lagenodelphis hosei | |
| Killer whale | Orcinus orca | |
| Long-beaked common dolphin | Delphinus capensis | |
| Melon-headed whale | Peponocephala electra | |
| Pantropical spotted dolphin | Stenella attenuate | |
| Pygmy killer whale | Feresa attenuata | |
| Risso’s dolphin | Grampus griseus | |
| Rough-toothed dolphin | Steno bredanensis | |
| Short-finned pilot whale | Globicephala macrorhynchus | |
| Spinner dolphin | Stenella longirostris | |
| Striped dolphin | Stenella coeruleoalba | |
| Indo-Pacific bottlenose dolphin | Tursiops aduncus | |
| Indo-Pacific humpback dolphin | Sousa chinensis | |
| Irrawaddy dolphin | Orcaella brevirostris | |
| Kogiidae | Pygmy sperm whale | Kogia breviceps |
| Dwarf sperm whale | Kogia sima | |
| Phocoenidae | Finless porpoise | Neophocaena phocaenoides |
| Physeteridae | Sperm whale | Physeter macrocephalus |
| Ziphiidae | Blainville’s beaked whale | Mesoplodon densirostris |
| Cuvier’s beaked whale | Ziphius cavirostris | |
| Gingko-toothed beaked whale | Mesoplodon ginkgodens |
| Species | Location of Sample | Percent Identity | Accession Number |
|---|---|---|---|
| B. acutorostrata | North Pacific | 98.84 | AJ226105.1 |
| B. acutorostrata | North Pacific | 98.54 | Y17160.1 |
| B. acutorostrata | North Pacific | 98.39 | KT581986.1 |
| B. acutorostrata | North Pacific | 98.34 | AJ226110.1 |
| B. acutorostrata | Unknown | 98.04 | AY878077.1 |
| B. acutorostrata | Unknown | 97.94 | AJ226103.1 |
| B. acutorostrata | Unknown | 97.73 | KY542104.1 |
| B. acutorostrata | Unknown | 97.73 | KY542103.1 |
| B. acutorostrata | Mediterranean | 97.28 | AY230267.1 |
| B. acutorostrata | North Atlantic | 97.07 | KJ586812.1 |
| B. acutorostrata | Unknown | 96.82 | MT410935.1 |
| B. acutorostrata | North Atlantic | 96.71 | X72006.1 |
| B. acutorostrata | North Atlantic | 96.49 | AP006468.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chantra, R.; Nganvongpanit, K.; Yaowasooth, P.; Thongsukdee, S.; Kitiwatthanawong, K.; Thongcharoenchaikit, C.; Brown, J.L.; Piboon, P. First Stranding Event of a Common Minke Whale (Balaenoptera acutorostrata Lacépède, 1804) Reported in the Gulf of Thailand. Diversity 2023, 15, 532. https://doi.org/10.3390/d15040532
Chantra R, Nganvongpanit K, Yaowasooth P, Thongsukdee S, Kitiwatthanawong K, Thongcharoenchaikit C, Brown JL, Piboon P. First Stranding Event of a Common Minke Whale (Balaenoptera acutorostrata Lacépède, 1804) Reported in the Gulf of Thailand. Diversity. 2023; 15(4):532. https://doi.org/10.3390/d15040532
Chicago/Turabian StyleChantra, Rachawadee, Korakot Nganvongpanit, Patcharaporn Yaowasooth, Surasak Thongsukdee, Kongkiat Kitiwatthanawong, Cholawit Thongcharoenchaikit, Janine L. Brown, and Promporn Piboon. 2023. "First Stranding Event of a Common Minke Whale (Balaenoptera acutorostrata Lacépède, 1804) Reported in the Gulf of Thailand" Diversity 15, no. 4: 532. https://doi.org/10.3390/d15040532
APA StyleChantra, R., Nganvongpanit, K., Yaowasooth, P., Thongsukdee, S., Kitiwatthanawong, K., Thongcharoenchaikit, C., Brown, J. L., & Piboon, P. (2023). First Stranding Event of a Common Minke Whale (Balaenoptera acutorostrata Lacépède, 1804) Reported in the Gulf of Thailand. Diversity, 15(4), 532. https://doi.org/10.3390/d15040532