
Effects of Climate on Scorpion Diversity in Arid Ecosystems of the Sahara Desert of Algeria

 ,
, 
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
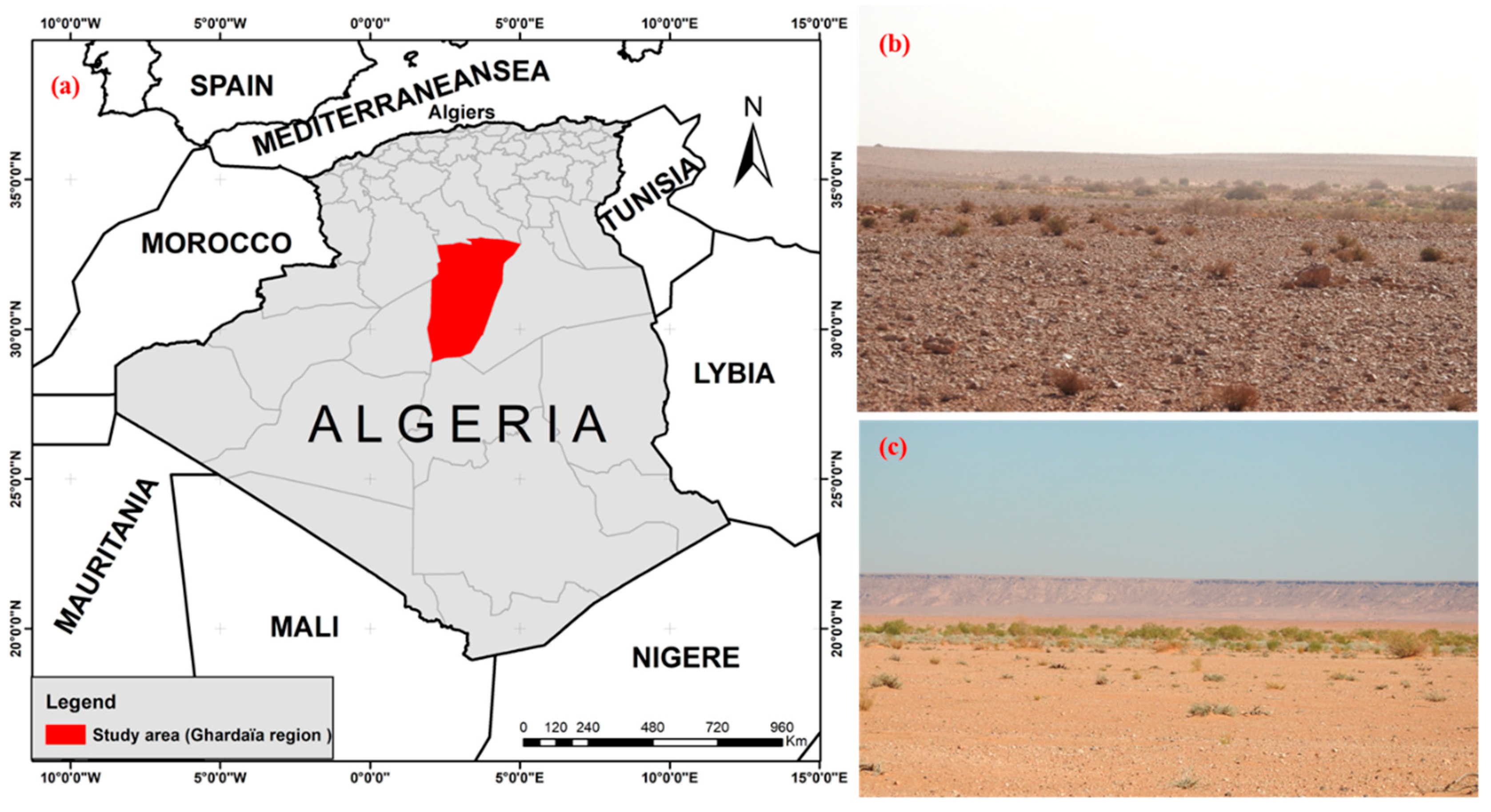
2.1. Study Site
2.2. Scorpion Sampling
2.3. Data Analysis
3. Results
3.1. Taxonomic Composition, Relative Abundance and Occurrence
3.2. Species Phenology
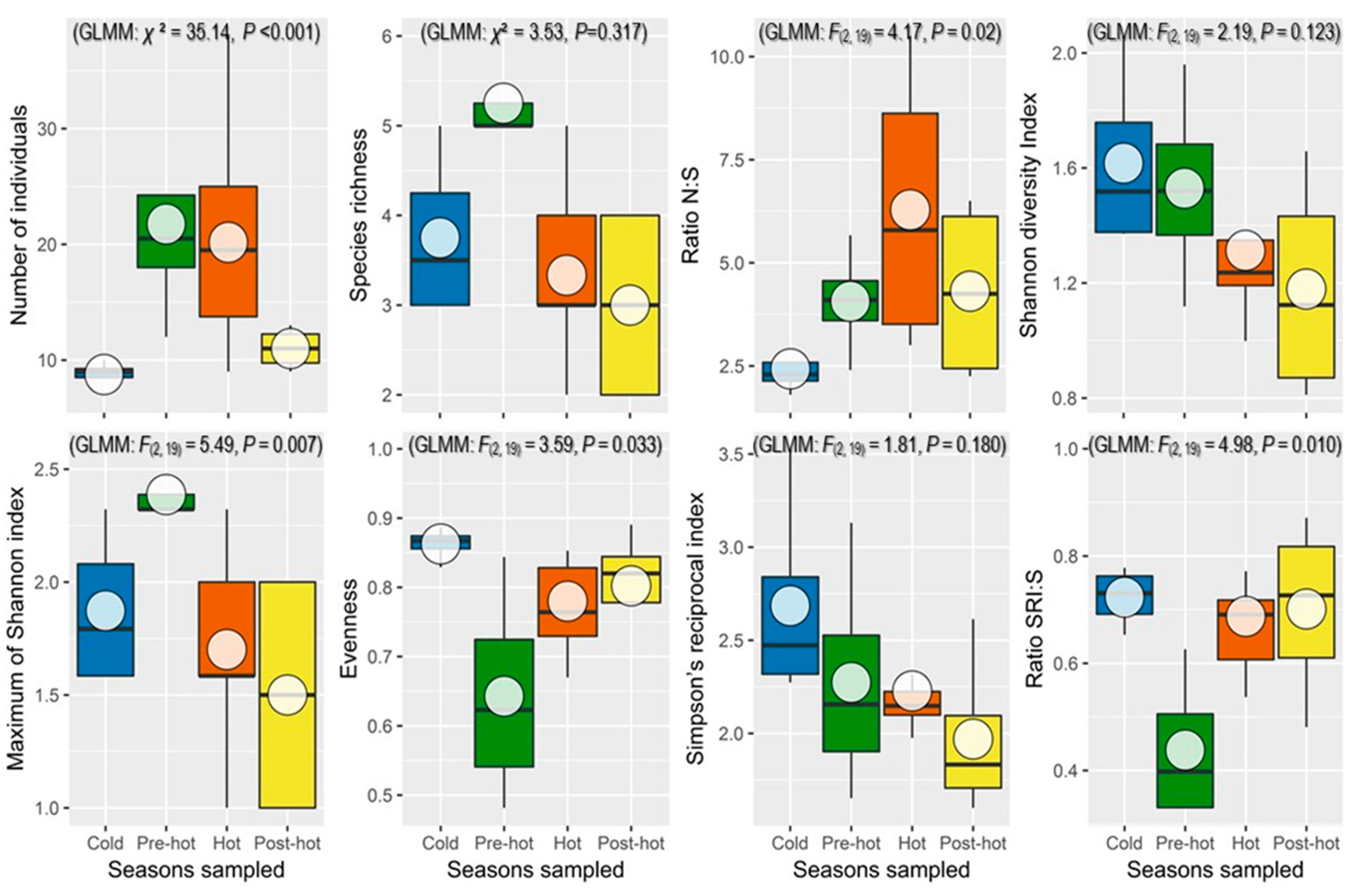
3.3. Variation of Diversity Parameters
3.3.1. Number of Individuals
3.3.2. The Ecological Indices according to Seasons
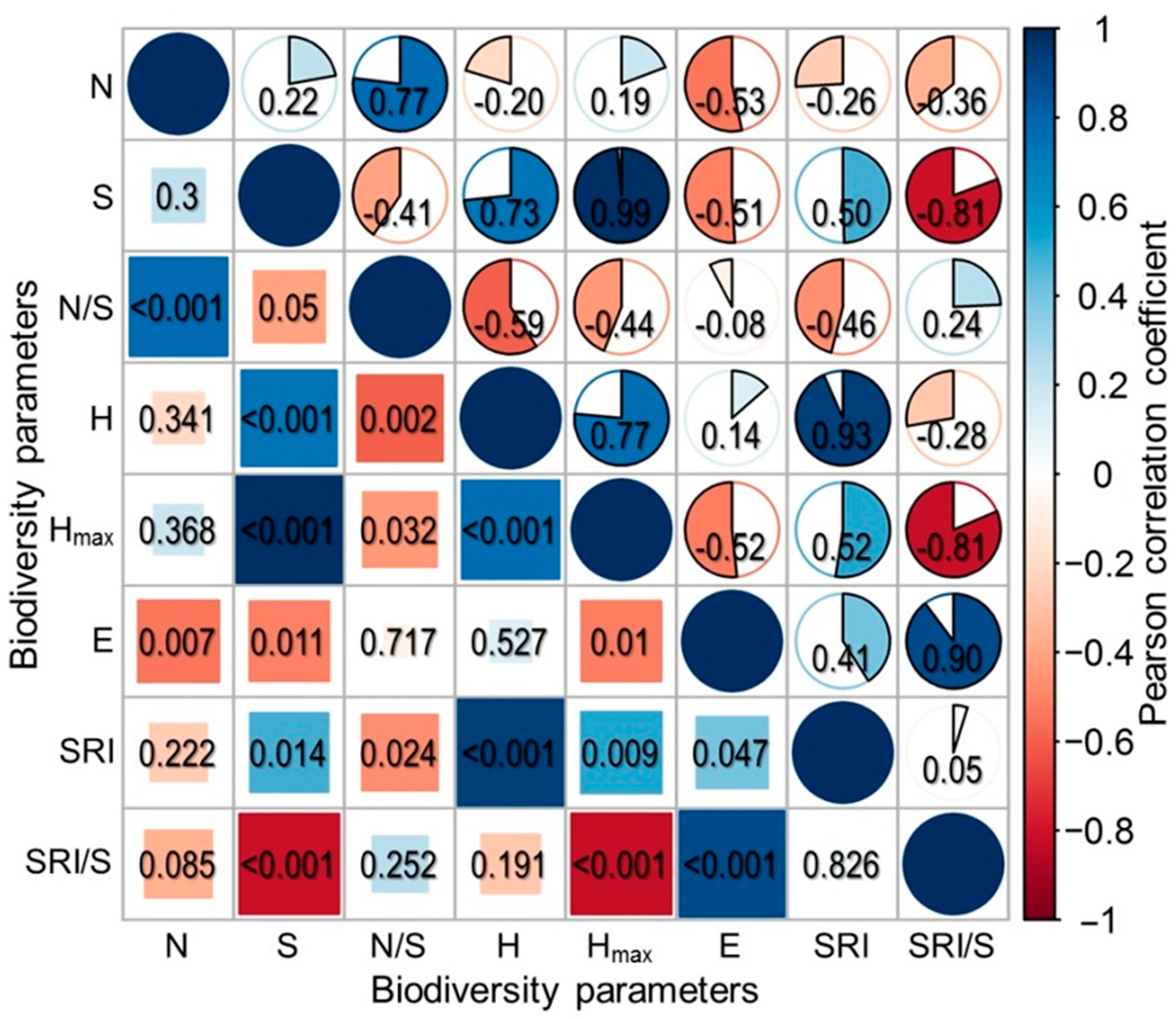
3.4. Relationships between Diversity Parameters
3.5. Effects of Climate on Variations of Biodiversity Parameters
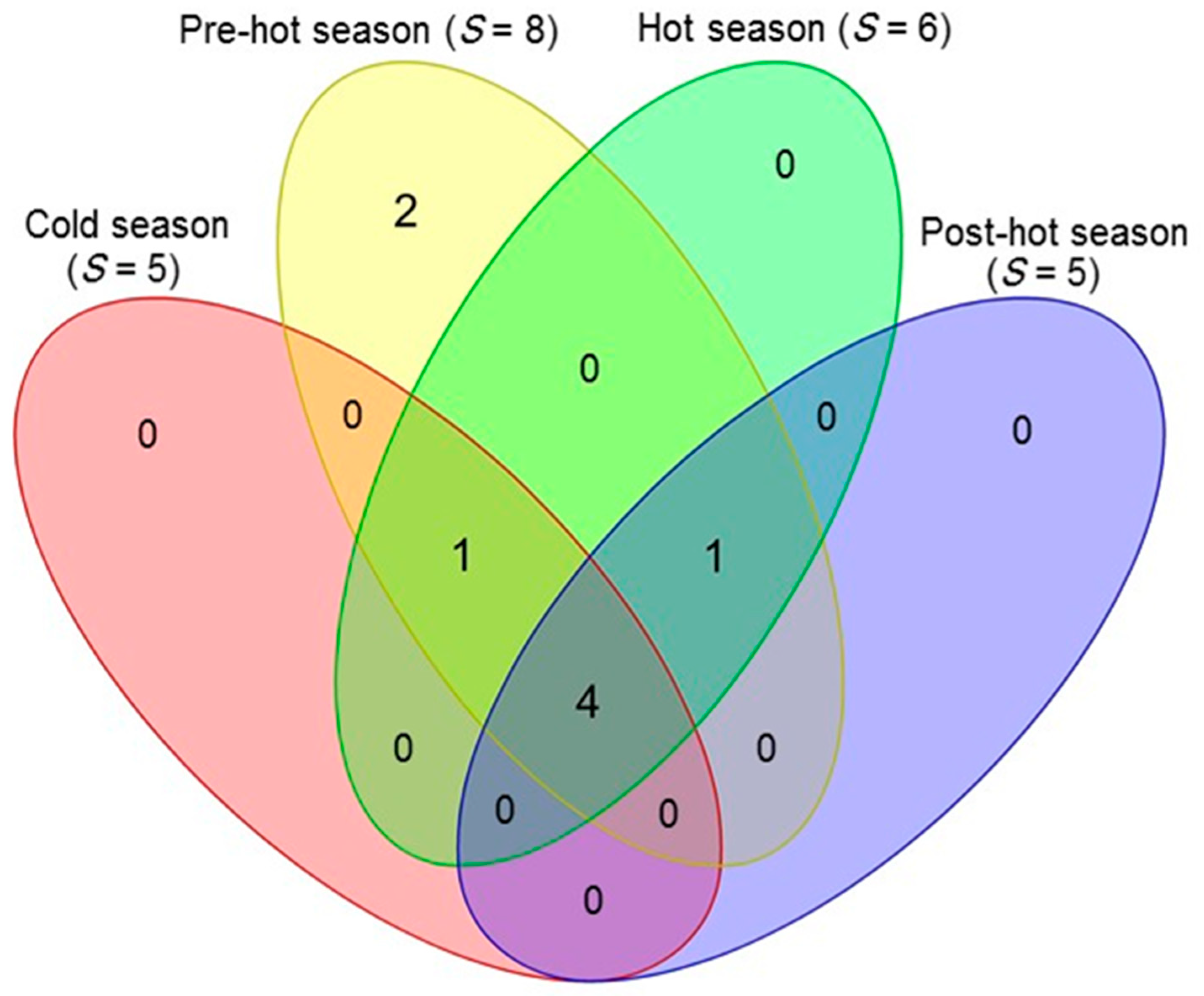
3.6. Distribution and Similarity Analysis of Scorpions According to the Seasons
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilson, E.O.J.B. The current state of biological diversity. Biodiversity 1988, 521, 3–18. [Google Scholar]
- Ceballos, G.; Ehrlich, P.; Barnosky, A.; Garcia, A.; Pringle, R.; Palmer, T. Accelerated Modern Human-Induced Species Losses: Entering the Sixth Mass Extinction. Sci. Adv. 2015, 1, e1400253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habel, J.C.; Segerer, A.; Ulrich, W.; Torchyk, O.; Weisser, W.W.; Schmitt, T. Butterfly community shifts over two centuries. Conserv. Biol. J. Soc. Conserv. Biol. 2016, 30, 754–762. [Google Scholar] [CrossRef] [PubMed]
- Hallmann, C.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Ller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef] [Green Version]
- Seibold, S.; Gossner, M.; Simons, N.; Blüthgen, N.; Müller, J.; Ambarlı, D.; Ammer, C.; Bauhus, J.; Fischer, M.; Habel, J.; et al. Arthropod decline in grasslands and forests is associated with landscape-level drivers. Nature 2019, 574, 671–674. [Google Scholar] [CrossRef]
- Durant, S.; Pettorelli, N.; Bashir, S.; Woodroffe, R.; Wacher, T.; Ornellas, P.; Ransom, C.; Abaigar, T.; Abdelgadir, M.; El Alqamy, H.; et al. Forgotten Biodiversity in Desert Ecosystems. Science 2012, 336, 1379–1380. [Google Scholar] [CrossRef]
- Smith, J.E.; Pettorelli, N. Desert Conservation and Management: Biodiversity Loss. In Reference Module in Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Davies, J.; Poulsen, L.; Schulte-Herbrüggen, B.; MacKinnon, K.; Henwood, W.; Dudley, N.; Smith, J.; Gudka, M. Conserving Drylands Biodiversity; IUCN: Gland, Switzerland, 2012; p. 48. [Google Scholar]
- Lovich, J.; Bainbridge, D. Anthropogenic Degradation of the Southern California Desert Ecosystem and Prospects for Natural Recovery and Restoration. Environ. Manag. 1999, 24, 309–326. [Google Scholar] [CrossRef]
- Brito, D. Overcoming the Linnean shortfall: Data deficiency and biological survey priorities. Basic. Appl. Ecol. 2010, 11, 709–713. [Google Scholar] [CrossRef]
- Cardoso, P.; Erwin, T.L.; Borges, P.A.V.; New, T.R. The seven impediments in invertebrate conservation and how to overcome them. Biol. Conserv. 2011, 144, 2647–2655. [Google Scholar] [CrossRef] [Green Version]
- Schwerdt, L.; Copperi, S.; Pompozzi, G.; Ferretti, N. Diversity and seasonal composition of the scorpion fauna from a mountainous system on pampean grasslands in central Argentina. Stud. Neotrop. Fauna. Environ. 2016, 51, 169–175. [Google Scholar] [CrossRef]
- Polis, G.A. The Biology of Scorpions; Stanford University Press: Redwood City, CA, USA, 1990. [Google Scholar]
- Cloudsley-Thompson, J.J.B.-I.O.B. Scorpions. Biologist 1992, 39, 206–210. [Google Scholar]
- Giribet, G.; Edgecombe, G.D. The Phylogeny and Evolutionary History of Arthropods. Curr. Biol. 2019, 29, R592–R602. [Google Scholar] [CrossRef]
- Dunlop, J.; Tetlie, O.; Prendini, L. Reinterpretation of the Silurian scorpion Proscorpius osborni (Whitfield): Integrating data from Palaeozoic and Recent scorpions. Palaeontology 2008, 51, 303–320. [Google Scholar] [CrossRef]
- Dehghani, R.; Kassiri, H. A Checklist of Scorpions in Iran (By 2017). Asian J. Pharm. 2018, 12, 880. [Google Scholar]
- Rein, J.O. The Scorpion Files; Norwegian University of Science and Technology: Trondheim, Norway, 2016; Available online: https://www.ntnu.no/ub/scorpion-files (accessed on 23 January 2023).
- Porto, T.; Brazil, T.; Lira-da-Silva, R. Scorpions, state of Bahia, northeastern Brazil. Check List 2010, 6, 292–297. [Google Scholar] [CrossRef] [Green Version]
- Mullen, G.R.; Sissom, W.D. Chapter 23—Scorpions (Scorpiones). In Medical and Veterinary Entomology, 3rd eds; Mullen, G.R., Durden, L.A., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 489–504. [Google Scholar]
- Francke, O. Studies of the scorpion subfamilies Superstitioninae and Typhlochactinae, with description of a new genus (Scorpiones, Chactidae). Bull. Assoc. Mex. Cave Stud. 1982, 8, 51–61. [Google Scholar]
- Araújo, V.; Bandeira, A.; Vasconcellos, A. Abundance and stratification of soil macroarthropods in a Caatinga Forest in Northeast Brazil. Braz. J. Biol. 2010, 70, 737–746. [Google Scholar] [CrossRef] [Green Version]
- Chedad, A.; Hammou, M.A.; Chelghoum, H.; Chedad, A.; Amara, O.O.; El Bouhissi, M.; Dahmani, W.; Sadine, S.E. Diversity and distribution pattern of scorpions from the Ouarsenis massif of Tissemsilt, North-West Algeria. Biodiversitas 2022, 23, 2444–2450. [Google Scholar] [CrossRef]
- Dias, S.; Candido, D.; Brescovit, A. Scorpions from Mata do Buraquinho, João Pessoa, Paraíba, Brazil, with ecological notes on a population of Ananteris mauryi Lourenço (Scorpiones, Buthidae). Rev. Bras. Zool. 2006, 23, 707–710. [Google Scholar] [CrossRef] [Green Version]
- Lira, A.F.; Salomão, R.P.; Albuquerque, C.M. Pattern of scorpion diversity across a bioclimatic dry-wet gradient in Neotropical forests. Acta Oecol. 2019, 96, 10–17. [Google Scholar] [CrossRef]
- Nime, M.; Casanoves, F.; Mattoni, C. Scorpion diversity in two different habitats in the Arid Chaco, Argentina. J. Insect. Conserv. 2014, 18, 373–384. [Google Scholar] [CrossRef]
- Nime, M.F.; Casanoves, F.; Vrech, D.E.; Mattoni, C.I. Relationship between environmental variables and surface activity of scorpions in the Arid Chaco ecoregion of Argentina. Invertebr. Biol. 2013, 132, 145–155. [Google Scholar] [CrossRef]
- Pizarro-Araya, J.; Ojanguren, A.; López-Cortés, F.; Agusto, P.; Briones, R.; Cepeda-Pizarro, J. Diversidad y composición estacional de la escorpiofauna (Arachnida: Scorpiones) del archipiélago Los Choros (Región de Coquimbo, Chile). Gayana (Concepción) 2014, 78, 46–56. [Google Scholar] [CrossRef] [Green Version]
- Prendini, L. Scorpion Diversity and Distribution in Southern Africa: Pattern and Process. In African Biodiversity; Springer: Boston, MA, USA, 2005; pp. 25–68. [Google Scholar]
- Yamaguti, H.Y.; Pinto-da-Rocha, R. Ecology of thestylus aurantiurus of the parque estadual da serra da cantareira, São paulo, Brazil (Scorpiones, Bothriuridae). J. Arachnol. 2006, 34, 214–220. [Google Scholar] [CrossRef]
- Jiménez, M.-L.; Navarrete, J. Fauna de arañas del suelo de una comunidad árida-tropical en Baja California Sur, México. Rev. Mex. Biodivers. 2010, 81, 417–426. [Google Scholar] [CrossRef]
- Koch, L. The Scorpions of Australia: Aspects of Their Ecology and Zoogeography; Elsevier: Amsterdam, The Netherlands, 1981; pp. 873–884. [Google Scholar]
- Koch, L.E. The taxonomy, geographic disctribution and evolutionary radiation of austalo-papauan scorpions. Rec. West. Aust. Mus. 1977, 5, 83–367. [Google Scholar]
- Newlands, G. Arachnida (except Acari). In Biogeography and Ecology of Southern Africa; Springer: Berlin/Heidelberg, Germany, 1978; pp. 685–702. [Google Scholar]
- Brownell, P.; Polis, G.A. Scorpion Biology and Research, 1st ed.; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Lira, A.F.A.; Souza, A.M.; Silva Filho, A.A.C.; Albuquerque, C.M.R. Spatio-temporal microhabitat use by two co-occurring species of scorpions in Atlantic rainforest in Brazil. Zoology 2013, 116, 182–185. [Google Scholar] [CrossRef]
- Polis, G.A. The effect of cannibalism on the demography and activity of a natural population of desert scorpions. Behav. Ecol. Sociobiol. 1980, 7, 25–35. [Google Scholar] [CrossRef]
- Schowalter, T.D. Insect responses to major landscape-level disturbance. Annu. Rev. Entomol. 2012, 57, 1–20. [Google Scholar] [CrossRef]
- Uetz, G. The influence of variation in litter habitats on spider communities. Oecologia 1979, 40, 29–42. [Google Scholar] [CrossRef]
- Sadine, S.E.; Bissati, S.; Idder, M. Diversity and structure of scorpion fauna from arid ecosystem in Algerian Septentrional Sahara (2005–2018). Serket 2018, 16, 51–59. [Google Scholar]
- Dupre, G.; El Bouhissi, M.; Sadine, S.E. La Faune des Scorpions d’Algérie. Arachnides 2023, 108. [Google Scholar]
- Mekahlia, M.N.; Abidi, H.; Slimane, F.; Sadine, S.E.; Dekak, A.; Chenchouni, H. Seasonal patterns of scorpion diversity along a gradient of aridity in Algeria. Acta. Oecol. 2021, 113, 103792. [Google Scholar] [CrossRef]
- Sadine, S.E.; Djilani, S.; Kerboua, K.E. Overview on Scorpions of Algeria. Alg. J. Health Sci. 2020, 2, 8–14. [Google Scholar]
- Vachon, M. Etude sur les Scorpions Alger: Institut Pasteur d’Algérie, 482 pp. Vachon, M. 1973 [1974]. Etude des caracteres utilisés pour classer les familles et les genres de scorpions (Arachnides). 1. La trichobothriotaxie en arachnologie: Sigles trichobothriaux et types de trichobothriotaxie chez les scorpions. Bull. Mus. Natl. Hist. Nat. 1952, 3, 857–958. [Google Scholar]
- Qi, J.-X.; Lourenço, W.R. Distribution of endemic relict groups of Saharan scorpions, with the description of new genus and species from Mauritania. C. R. Biol. 2007, 330, 80–85. [Google Scholar] [CrossRef]
- Lourenco, W.R.; Duhem, B. Saharo-Sindian buthid scorpions; description of two new genera and species from Occidental Sahara and Afghanistan. ZooKeys 2009, 14, 37–54. [Google Scholar] [CrossRef]
- Affilastro, A.O. Estudio monográfico de los escorpiones de la República Argentina. Rev. Iber. Aracnol. 2005, 11, 75–241. [Google Scholar]
- Bradley, R. The Influence of Weather and Biotic Factors on the Behaviour of the Scorpion (Paruroctonus utahensis). J. Anim. Ecol. 1988, 57, 533–551. [Google Scholar] [CrossRef]
- Carvalho, L.S.; Sebastian, N.; Araújo, H.F.; Dias, S.C.; Venticinque, E.; Brescovit, A.D.; Vasconcellos, A. Climatic variables do not directly predict spider richness and abundance in semiarid Caatinga vegetation, Brazil. Environ. Entomol. 2015, 44, 54–63. [Google Scholar] [CrossRef]
- Bobka, M.; Jaeger, R.; McNaught, D. Temperature Dependent Assimilation Efficiencies of Two Species of Terrestrial Salamanders. Copeia 1981, 1981, 417–421. [Google Scholar] [CrossRef]
- Zhang, Y.; Ji, X. The thermal dependence of food assimilation and locomotor performance in southern grass lizards, Takydromus sexlineatus (Lacertidae). J. Therm. Biol. 2004, 29, 45–53. [Google Scholar] [CrossRef]
- Angilletta, M.; Steury, T.; Sears, M. Temperature, Growth Rate, and Body Size in Ectotherms: Fitting Pieces of a Life-History Puzzle. Integr. Comp. Biol. 2004, 44, 498–509. [Google Scholar] [CrossRef] [PubMed]
- Forsman, A. Variation in thermal sensitivity of performance among colour morphs of a pygmy grasshopper. J. Evol. Biol. 1999, 12, 869–878. [Google Scholar] [CrossRef]
- Benkenzou, D.; Chegma, S.; Merakchi, F.; Zidane, B. Statistiques au 31 décembre 2006. In Monographie de la Wilaya de Ghardaïa; Direction de la Planification et de l’Aménagement du Territoire (D.P.A.T.): Ghardaïa, Algeria, 2007; p. 122. [Google Scholar]
- Sam, F. Réhabilitation Thermique d’un Local dans une Zone Aride cas de Ghardaia. Ph.D. Thesis, Université Mouloud Mammeri, Tizi Ouzou, Algeria, 2012. [Google Scholar]
- Chehma, A. Séminaire L’effet du Changement Climatique sur l’élevage et la gestion durable des parcours dans les zones arides et semiarides du Maghreb. Du 21 au 24 Novembre 2011. In Le Sahara en Algérie, Situation et Défis; Université Kasdi Merbah: Ouargla, Algeria, 2011; pp. 1–8. [Google Scholar]
- Stahnke, H.L. Scorpion nomenclature and mensuration. Entomol. News 1970, 81, 297–316. [Google Scholar]
- Vachon, M. Étude des caractères utilisés pour classer les familles et les genres de Scorpions (Arachnides). Bull. Mus. Natl. Hist. Nat. 1973, 140, 857–958. [Google Scholar]
- Hjelle, J.T. Anatomy and morphology. In The Biology of Scorpions; Polis, G.A., Ed.; Stanford University Press: Stanford, CA, USA, 1990; pp. 9–63. [Google Scholar]
- Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Magurran, A. Measuring Biological Diversity; Blackwell Publishing: Oxford, UK, 2004. [Google Scholar]
- Bigot, L.; Bodot, P. Contribution à l’étude biocénotique de la garrigue à Quercus coccifera. Composition biotique du peuplement des invertébrés. Vie Milieu 1973, 23, 229–249. Available online: https://hal.sorbonne-universite.fr/hal-02982385 (accessed on 14 October 2022).
- Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples, Version 9 and Earlier; User’s Guide and application; University of Connecticut: Mansfield, Connecticut, USA, 2013. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Sadine, S.E. Scorpion Fauna of the Algerian Septentrional Sahara: Diversity and Ecology. Ph.D. Thesis, Université de Ouargla, Ouargla, Algeria, 2018. [Google Scholar]
- Brown, J.H. On the relationship between abundance and distribution of species. Am. Nat. 1984, 124, 255–279. [Google Scholar] [CrossRef]
- Kaltsas, D.; Stathi, I.; Mylonas, M. Intraspecific differentiation of social behavior and shelter selection in Mesobuthus gibbosus (Brullé, 1832)(Scorpiones: Buthidae). J. Ethol. 2009, 27, 467–473. [Google Scholar] [CrossRef]
- Lankau, R.A. Rapid evolutionary change and the coexistence of species. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 335–354. [Google Scholar] [CrossRef]
- Shivashankar, T. Advanced sub social behaviour in the scorpion Heterometrus fulvipes Brunner (Arachnida). J. Biosci. 1994, 19, 81–90. [Google Scholar] [CrossRef]
- Warburg, M. Intra- and interspecific cohabitation of scorpions in the field and the effect of density, food, and shelter on their interactions. J. Ethol. 2000, 18, 59–63. [Google Scholar] [CrossRef]
- Sadine, S.E. Contribution to the study of scorpion fauna of Northern Sahara, eastern Algeria (El Oued and Ouargla). Master’s Thesis, University of Ouargla, Ouargla, Algeria, 2012. (In French). [Google Scholar]
- Sadine, S.E.; Samia, B.; Ould Elhadj, M. Premieres donnees sur la diversite scorpionique dans la region du Souf (Algerie). Arachnides 2011, 61, 1–10. [Google Scholar]
- Ouici, H.; El Bouhissi, M.; Sadine, S.E.; Abidi, H. Preliminary study and ecological comments on scorpion diversity in Sidi Bel Abbes region, North-west Algeria. Serket 2020, 17, 87–96. [Google Scholar]
- Hasnaoui, C.; Meddour, A.; Lebbal, S. New data on scorpion diversity in the region of Khenchela, Algeria. Serket 2018, 16, 1–6. [Google Scholar]
- Lourenço, W.R.; Sadine, S.E.; Bissati, S.; Houtia, A. The genus Buthacus Birula, 1908 in northern and central Algeria; description of a new species and comments on possible microendemic populations (Scorpiones: Buthidae). Arachnida 2017, 3, 18–30. [Google Scholar]
- Bengaid, Y. Composition et Structure de Peuplement Scorpionique de la Région de Ghardaïa (Algérie). Mémoire de Master Académique en Ecologie, Université de Ghardaïa, Ghardaia, Algeria, 2018. [Google Scholar]
- Lahrech, A.; Souilem, Z. Contribution à L’étude de la Faune Scorpionique de la Région de Ghardaïa (Algérie). Mémoire de Master Académique en Ecologie, Université de Ghardaïa, Ghardaia, Algeria, 2017. [Google Scholar]
- Lourenço, W.R. New taxonomic considerations on the species of the genus Androctonus Ehrenberg, 1828 and description of two new species (Scorpiones, Buthidae). Rev. Suisse. Zool. 2005, 112, 145–171. [Google Scholar] [CrossRef]
- Aboshaala, F.; Badry, A.; Sadine, S.E. Ecological considerations on Buthiscus bicalcaratus Birula, 1905 with a new locality in northern Libya (Scorpiones, Buthidae). Rev. Iber. Aracnol. 2020, 36, 181–183. [Google Scholar]
- Saleh, M.; Younes, M.; Sarhan, M.; Badry, A. Zoogeographical Analysis of the Egyptian Scorpion Fauna. Azhar. Bull. Sci. 2017, 28, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Sadine, S.E. Contribution à L’étude Bioécologique de Quelques Espèces du Scorpion; Androctonus australis, Androctonus amoreuxi, Buthacus arenicola, Buthus tunetanus et Orthochirus innesi dans la Wilaya de Ouargla. Master’s Thesis, University of Ouargla, Ouargla, Algeria, 2005. [Google Scholar]
- Broglio, N.; Goyffon, M. Les accidents d'envenimation scorpionique. Conc. Med. 1980, 38, 5615–5622. [Google Scholar]
- Charnot, A.; Faure, L. Les Scorpions du Maroc, leur venin; leur danger pour l’homme et les animaux. Bull. Inst. Hyg. Maroc. 1934, 4, 1–72. [Google Scholar]
- Sadine, S.E.; Bissati, S. Ecology of Androctonus australis (Linnaeus, 1758) (Scorpiones: Buthidae) from an arid ecosystem of the Algerian Sahara. In Proceedings of the International Conference on Ecology, Ecosystems and Climate Change, Istanbul, Turkey, 15–16 February 2019; pp. 85–92. [Google Scholar]
- Stockmann, M.; Turiel, C.; Althoff, F.; Lowe, G.; Kovařík, F. First report of Lissothus occidentalis Vachon, 1950 (Scorpiones: Buthidae) from Morocco and Western Sahara, with notes on ecology and captive breeding. Euscorpius 2016, 2016, 1–12. [Google Scholar] [CrossRef] [Green Version]
- El Hidan, M.A.; Touloun, O.; Boumezzough, A. Spatial relationship between environmental factors and scorpion distribution in Morocco. J. Entomol. Zool 2017, 5, 674–678. [Google Scholar]
- Badry, A.; Younes, M.; Sarhan, M.M.; Saleh, M. On the scorpion fauna of Egypt, with an identification key (Arachnida: Scorpiones). Zool. Middle East 2018, 64, 75–87. [Google Scholar] [CrossRef]
- Aboshaala, F.; Yağmur, E.; Sadine, S.E.; Ghaliow, M.; Badry, A. On the poorly known species Buthiscus bicalcaratus Birula, 1905 (Scorpiones: Buthidae). Serket 2022, 18, 263–273. [Google Scholar]
- Auber-Thomay, M. Croissance et reproduction d’Androctonus australis (Scorpions, Buthidés). Ann. Sci. Nat. Zool. Biol. Anim. 1974, 12, 45–54. [Google Scholar]
- Fet, V. Ecology of the scorpions (Arachnida, Scorpiones) of the southeastern Kara-Kum. Entomol. Obozr. 1980, 59, 223–228. [Google Scholar]
- Druce, D.; Hamer, M.; Slotow, R. Patterns of millipede (Diplopoda), centipede (Chilopoda) and scorpion (Scorpionida) diversity in savanna habitats within the Greater Makalali Conservancy, South Africa. Afr. Zool. 2007, 42, 204–215. [Google Scholar] [CrossRef]
- Prendini, L. Substratum specialization and speciation in southern African scorpions: The Effect Hypothesis revisited. Scorpions 2001, 2001, 113–138. [Google Scholar]
- Haghani, A.; Khoobdel, M.; Dehghani, R.; Adibzadeh, A.; Sobati, H.; Aliabadian, M. Ecological modeling and distribution analysis of digger scorpions: Odontobuthus doriae, Odonthubutus bidentatus (Scorpiones: Buthidae) and Scorpio maurus (Scorpiones: Scorpionidae) in Iran using the maximum entropy method. Appl. Entomol. Zool. 2020, 55, 17–24. [Google Scholar] [CrossRef]
- Foord, S.H.; Gelebe, V.; Prendini, L. Effects of aspect and altitude on scorpion diversity along an environmental gradient in the Soutpansberg, South Africa. J. Arid Environ. 2015, 113, 114–120. [Google Scholar] [CrossRef]
- Warburg, M.R.; Ben-Horin, A. The response to temperature gradients of scorpions from mesic and xeric habitats. Comp. Biochem. Physiol. Part A Physiol. 1981, 68, 277–279. [Google Scholar] [CrossRef]
- Bryson, R.W., Jr.; Riddle, B.R.; Graham, M.R.; Smith, B.T.; Prendini, L. As old as the hills: Montane scorpions in southwestern North America reveal ancient associations between biotic diversification and landscape history. PLoS ONE 2013, 8, e52822. [Google Scholar] [CrossRef] [Green Version]
- Bryson, R.W., Jr.; Savary, W.E.; Prendini, L. Biogeography of scorpions in the Pseudouroctonus minimus complex (Vaejovidae) from south-western North America: Implications of ecological specialization for pre-Quaternary diversification. J. Biogeogr. 2013, 40, 1850–1860. [Google Scholar] [CrossRef]
- Due, A.D.; Polis, G.A. Trends in scorpion diversity along the Baja California peninsula. Am. Nat. 1986, 128, 460–468. [Google Scholar] [CrossRef]
- Prendini, L.; Bird, T. Scorpions of the Brandberg Massif, Namibia: Species Richness Inversely Correlated with Altitude. Afr. Invertebr. 2008, 49, 77–107. [Google Scholar] [CrossRef]
- Ureta, C.; González, E.; Ramírez-Barrón, M.; Contreras-Félix, G.; Santibáñez-López, C. Climate change will have an important impact on scorpion’s fauna in its most diverse country, Mexico. Perspect. Ecol. Conserv. 2020, 18, 116–123. [Google Scholar] [CrossRef]
- Sadine, S.E. New locality of Orthochirus innesi Simon, 1910 in Algeria (Scorpiones: Buthidae). Serket 2020, 17, 171–175. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus (RA in %) | Species | Traits | Seasons | Overall | |||
|---|---|---|---|---|---|---|---|
| Cold | Pre-Hot | Hot | Post-Hot | ||||
| Androctonus (91.18) | A. aeneas C.L. Koch, 1839 | Ni | — | 5 | 6 | 1 | 12 |
| RA [%] | — | 1.23 | 1.47 | 0.25 | 2.94 | ||
| Occ | — | 75 | 41.7 | 25 | 37.5 | ||
| Scale | — | Omn | Com | Com | Com | ||
| A. amoreuxi (Audouin, 1826) | Ni | 16 | 57 | 125 | 24 | 222 | |
| RA [%] | 3.92 | 13.97 | 30.64 | 5.88 | 54.41 | ||
| Occ | 100 | 100 | 100 | 100 | 100 | ||
| Scale | Omn | Omn | Omn | Omn | Omn | ||
| A. australis (Linnaeus, 1758) | Ni | 10 | 15 | 97 | 16 | 138 | |
| RA [%] | 2.45 | 3.68 | 23.77 | 3.92 | 33.82 | ||
| Occ | 100 | 100 | 100 | 100 | 100 | ||
| Scale | Omn | Omn | Omn | Omn | Omn | ||
| Buthacus (2.45) | B. elmenia Lourenço and Sadine, 2017 | Ni | — | 2 | — | — | 2 |
| RA [%] | — | 0.49 | — | — | 0.49 | ||
| Occ | — | 50 | — | — | 8.3 | ||
| Scale | — | Cst | — | — | Vac | ||
| B. samiae Lourenço and Sadine, 2015 | Ni | 2 | 4 | 2 | — | 8 | |
| RA [%] | 0.49 | 0.98 | 0.49 | — | 1.96 | ||
| Occ | 50 | 75 | 16.7 | — | 29.2 | ||
| Scale | Cst | Omn | Acc | — | Com | ||
| B. spinatus Lourenço, Bissati and Sadine, 2016 | Ni | — | 2 | — | — | 2 | |
| RA [%] | — | 0.49 | — | — | 0.49 | ||
| Occ | — | 50 | — | — | 8.3 | ||
| Scale | — | Cst | — | — | Vac | ||
| Buthus (1.47) | B. saharicus Sadine, Bissati and Lourenço, 2016 | Ni | 2 | 1 | 2 | 1 | 6 |
| RA [%] | 0.49 | 0.25 | 0.49 | 0.25 | 1.47 | ||
| Occ | 50 | 25 | 16.7 | 25 | 25 | ||
| Scale | Cst | Com | Acc | Com | Com | ||
| Lissothus (4.41) | L. chaambi Lourenço and Sadine, 2014 | Ni | 5 | 1 | 10 | 2 | 18 |
| RA [%] | 1.23 | 0.25 | 2.45 | 0.49 | 4.41 | ||
| Occ | 75 | 25 | 58.3 | 50 | 54.2 | ||
| Scale | Cst | Com | Cst | Cst | Cst | ||
| Total | Ni | 35 | 87 | 242 | 44 | 408 | |
| RA [%] | 8.58 | 21.32 | 59.31 | 10.78 | 100 | ||
| Variables | Estimate | S.E. | z-Value | p | Sig. | Estimate | S.E. | z-Value | p | Sig. |
|---|---|---|---|---|---|---|---|---|---|---|
| A. amoreuxi | A. australis | |||||||||
| Intercept | 1.605 | 0.523 | 3.068 | 0.002 | ** | −0.092 | 0.729 | −0.127 | 0.899 | ns |
| T | −0.008 | 0.009 | −0.843 | 0.399 | ns | 0.035 | 0.012 | 2.970 | 0.003 | ** |
| PP | −0.026 | 0.023 | −1.106 | 0.269 | ns | −0.055 | 0.036 | −1.526 | 0.127 | ns |
| W | 0.074 | 0.027 | 2.690 | 0.007 | ** | 0.079 | 0.035 | 2.245 | 0.025 | * |
| RA | −0.125 | 0.054 | −2.298 | 0.022 | * | −0.031 | 0.072 | −0.427 | 0.670 | ns |
| A. aeneas | B. elmenia | |||||||||
| Intercept | −0.481 | 2.180 | −0.221 | 0.825 | ns | −10.691 | 5.408 | −1.977 | 0.048 | * |
| T | −0.052 | 0.040 | −1.294 | 0.196 | ns | −0.143 | 0.096 | −1.486 | 0.137 | ns |
| PP | −0.119 | 0.165 | −0.722 | 0.471 | ns | 0.006 | 0.252 | 0.026 | 0.980 | ns |
| W | 0.096 | 0.120 | 0.802 | 0.423 | ns | 0.769 | 0.339 | 2.267 | 0.023 | * |
| RA | −0.204 | 0.236 | −0.863 | 0.388 | ns | 0.047 | 0.318 | 0.147 | 0.883 | ns |
| B. samiae | B. spinatus | |||||||||
| Intercept | 3.752 | 2.654 | 1.414 | 0.158 | ns | −3.140 | 3.332 | −0.942 | 0.346 | ns |
| T | −0.198 | 0.094 | −2.100 | 0.036 | * | −0.103 | 0.068 | −1.502 | 0.133 | ns |
| PP | 0.048 | 0.144 | 0.337 | 0.736 | ns | −0.294 | 0.371 | −0.791 | 0.429 | ns |
| W | −0.039 | 0.150 | −0.258 | 0.797 | ns | 0.274 | 0.192 | 1.426 | 0.154 | ns |
| RA | −0.469 | 0.284 | −1.649 | 0.099 | ns | 0.085 | 0.261 | 0.328 | 0.743 | ns |
| B. saharicus | L. chaambi | |||||||||
| Intercept | −3.222 | 3.643 | −0.884 | 0.377 | ns | −0.513 | 1.802 | −0.285 | 0.776 | ns |
| T | −0.095 | 0.092 | −1.040 | 0.298 | ns | −0.033 | 0.035 | −0.956 | 0.339 | ns |
| PP | 0.240 | 0.121 | 1.993 | 0.046 | * | 0.068 | 0.062 | 1.110 | 0.267 | ns |
| W | 0.164 | 0.175 | 0.936 | 0.349 | ns | 0.054 | 0.094 | 0.576 | 0.565 | ns |
| RA | 0.325 | 0.340 | 0.956 | 0.339 | ns | 0.024 | 0.182 | 0.133 | 0.894 | ns |
| N | Estimate | S.E. | z-Value | p | Sig. | S | Estimate | S.E. | z-Value | p | Sig. |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Intercept | 2.163 | 0.388 | 5.579 | <0.001 | *** | Intercept | 1.785 | 0.791 | 2.257 | 0.024 | * |
| T | −0.001 | 0.007 | −0.176 | 0.861 | ns | T | −0.029 | 0.015 | −1.925 | 0.054 | ns |
| PP | −0.033 | 0.017 | −1.861 | 0.063 | ns | PP | 0.015 | 0.031 | 0.477 | 0.633 | ns |
| W | 0.066 | 0.020 | 3.251 | 0.001 | ** | W | 0.014 | 0.042 | 0.330 | 0.742 | ns |
| RA | −0.104 | 0.040 | −2.616 | 0.009 | ** | RA | −0.046 | 0.083 | −0.556 | 0.578 | ns |
| N:S | Estimate | S.E. | t-value | p | Sig. | H | Estimate | S.E. | t-value | p | Sig. |
| Intercept | −1.087 | 3.418 | −0.318 | 0.758 | ns | Intercept | 2.087 | 0.417 | 5.001 | <0.001 | *** |
| T | 0.149 | 0.057 | 2.600 | 0.018 | * | T | −0.025 | 0.007 | −3.554 | 0.002 | ** |
| PP | −0.131 | 0.131 | −1.004 | 0.345 | ns | PP | 0.013 | 0.016 | 0.841 | 0.429 | ns |
| W | 0.222 | 0.172 | 1.293 | 0.217 | ns | W | −0.007 | 0.021 | −0.318 | 0.757 | ns |
| RA | −0.073 | 0.340 | −0.214 | 0.839 | ns | RA | −0.045 | 0.042 | −1.068 | 0.318 | ns |
| Hmax | Estimate | S.E. | t-value | p | Sig. | E | Estimate | S.E. | t-value | p | Sig. |
| Intercept | 2.734 | 0.494 | 5.534 | <0.001 | *** | Intercept | 0.763 | 0.180 | 4.236 | <0.001 | ** |
| T | −0.041 | 0.008 | −5.074 | <0.001 | *** | T | 0.004 | 0.003 | 1.150 | 0.271 | ns |
| PP | 0.011 | 0.019 | 0.584 | 0.577 | ns | PP | 0.000 | 0.007 | 0.056 | 0.963 | ns |
| W | 0.013 | 0.024 | 0.511 | 0.618 | ns | W | −0.008 | 0.009 | −0.791 | 0.461 | ns |
| RA | −0.100 | 0.049 | −2.071 | 0.059 | ns | RA | 0.017 | 0.018 | 0.932 | 0.441 | ns |
| SRI | Estimate | S.E. | t-value | p | Sig. | SRI:S | Estimate | S.E. | t-value | p | Sig. |
| Intercept | 3.042 | 0.717 | 4.240 | <0.001 | *** | Intercept | 0.489 | 0.223 | 2.199 | 0.050 | * |
| T | −0.026 | 0.013 | −2.099 | 0.052 | ns | T | 0.010 | 0.004 | 2.640 | 0.017 | * |
| PP | 0.022 | 0.028 | 0.799 | 0.490 | ns | PP | −0.003 | 0.009 | −0.294 | 0.790 | ns |
| W | −0.010 | 0.038 | −0.267 | 0.800 | ns | W | −0.007 | 0.012 | −0.640 | 0.540 | ns |
| RA | −0.048 | 0.073 | −0.653 | 0.573 | ns | RA | 0.022 | 0.023 | 0.992 | 0.375 | ns |
| First Sample | Second Sample | Jaccard Classic | Sorensen Classic | Morisita–Horn | Bray–Curtis |
|---|---|---|---|---|---|
| Cold | Pre-hot | 62.5 | 76.9 | 90.3 | 49.2 |
| Cold | Hot | 83.3 | 90.9 | 95.7 | 25.3 |
| Cold | Post-hot | 50 | 66.7 | 94 | 70.1 |
| Pre-hot | Hot | 75 | 85.7 | 91.5 | 49.2 |
| Pre-hot | Post-hot | 50 | 66.7 | 94.1 | 63.6 |
| Hot | Post-hot | 66.7 | 80 | 99.4 | 29.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sadine, S.E.; Souilem, Z.; Belgaid, Y.; Chedad, A.; Djelloud-Souilem, Z.; Chebihi, B.; Zouaimia, A.; Bensakhri, Z.; Houhamdi, M.; Zebsa, R. Effects of Climate on Scorpion Diversity in Arid Ecosystems of the Sahara Desert of Algeria. Diversity 2023, 15, 541. https://doi.org/10.3390/d15040541
Sadine SE, Souilem Z, Belgaid Y, Chedad A, Djelloud-Souilem Z, Chebihi B, Zouaimia A, Bensakhri Z, Houhamdi M, Zebsa R. Effects of Climate on Scorpion Diversity in Arid Ecosystems of the Sahara Desert of Algeria. Diversity. 2023; 15(4):541. https://doi.org/10.3390/d15040541
Chicago/Turabian StyleSadine, Salah Eddine, Zineb Souilem, Yacine Belgaid, Abdelwahab Chedad, Zineb Djelloud-Souilem, Bahri Chebihi, Abdelheq Zouaimia, Zinette Bensakhri, Moussa Houhamdi, and Rabah Zebsa. 2023. "Effects of Climate on Scorpion Diversity in Arid Ecosystems of the Sahara Desert of Algeria" Diversity 15, no. 4: 541. https://doi.org/10.3390/d15040541
APA StyleSadine, S. E., Souilem, Z., Belgaid, Y., Chedad, A., Djelloud-Souilem, Z., Chebihi, B., Zouaimia, A., Bensakhri, Z., Houhamdi, M., & Zebsa, R. (2023). Effects of Climate on Scorpion Diversity in Arid Ecosystems of the Sahara Desert of Algeria. Diversity, 15(4), 541. https://doi.org/10.3390/d15040541