

The Application of eDNA for Monitoring Aquatic Non-Indigenous Species: Practical and Policy Considerations
 ,
, {kind=link}
{kind=link}
Abstract
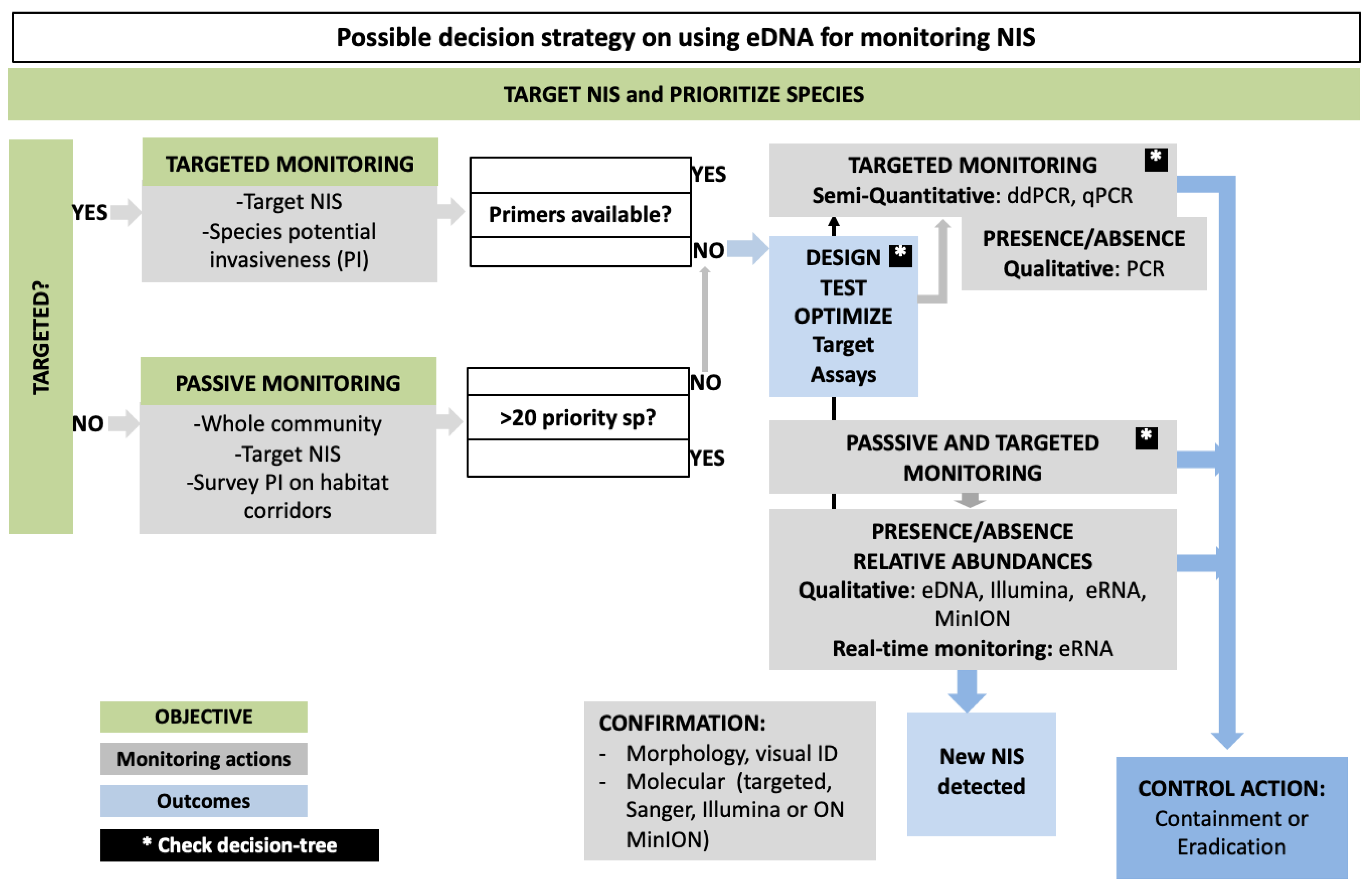
:1. Introduction
2. Detecting Aquatic NIS: Morphology, eDNA and eRNA
3. NIS Detection Using Targeted vs. Passive Approach: New Tools and Estimating Abundances
4. eDNA Fate and Impact on NIS Detection
5. NIS Detection Using PCR-Based Approaches: False Positives, False Negatives, and Sources of Error
6. eDNA as a Biomonitoring Tool: Challenges and Looking Forward
7. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- CBD. Convention on Biological Diversity. Pathways of Introduction of Invasive Species, Their Prioritization and Management. UNEP/CBD/SBSTTA/18/9/Add.1, Subsidiary Body on Scientific, Technical and Technological Advice. 2014. Available online: https://www.cbd.int/doc/meetings/sbstta/sbstta-18/official/sbstta-18-09-add1-en.pdf (accessed on 24 September 2019).
- Tsiamis, K.; Palialexis, A.; Stefanova, K.; Gladan, Ž.N.; Skejić, S.; Despalatović, M.; Cvitković, I.; Dragičević, B.; Dulčić, J.; Vidjak, O.; et al. Non-indigenous species refined national baseline inventories: A synthesis in the context of the European Union’s Marine Strategy Framework Directive. Mar. Pollut. Bull. 2019, 145, 429–435. [Google Scholar] [CrossRef]
- Molnar, J.L.; Gamboa, R.L.; Revenga, C.; Spalding, M.D. Assessing the global threat of invasive species to marine biodiversity. Front. Ecol. Environ. 2008, 6, 485–492. [Google Scholar] [CrossRef]
- Keller, R.P.; Geist, J.; Jeschke, J.M.; Kühn, I. Invasive species in Europe: Ecology, status, and policy. Environ. Sci. Eur. 2011, 23, 1–17. [Google Scholar] [CrossRef]
- Tidbury, H.J.; Taylor, N.G.H.; Copp, G.H.; Garnacho, E.; Stebbing, P.D. Predicting and mapping the risk of introduction of marine non-indigenous species into Great Britain and Ireland. Biol. Invasions 2016, 18, 3277–3292. [Google Scholar] [CrossRef]
- Darling, J.A.; Galil, B.S.; Carvalho, G.R.; Rius, M.; Viard, F.; Piraino, S. Recommendations for developing and applying genetic tools to assess and manage biological invasions in marine ecosystems. Mar. Policy 2018, 85, 56–64. [Google Scholar] [CrossRef]
- Blackburn, T.M.; Pyšek, P.; Bacher, S.; Carlton, J.T.; Duncan, R.P.; Jarošík, V.; Wilson, J.R.; Richardson, D.M. A proposed unified framework for biological invasions. Trends Ecol. Evol. 2011, 26, 333–339. [Google Scholar] [CrossRef]
- Couton, M.; Comtet, T.; Le Cam, S.; Corre, E.; Viard, F. Metabarcoding on planktonic larval stages: An efficient approach for detecting and investigating life cycle dynamics of benthic aliens. Manag. Biol. Invasions 2019, 10, 657–689. [Google Scholar] [CrossRef]
- Crane, L.C.; Goldstein, J.S.; Thomas, D.W.; Rexroth, K.S.; Watts, A.W. Effects of life stage on eDNA detection of the invasive European green crab (Carcinus maenas) in estuarine systems. Ecol. Indic. 2021, 124, 107412. [Google Scholar] [CrossRef]
- Klymus, K.E.; Richter, C.A.; Chapman, D.C.; Paukert, C. Quantification of eDNA shedding rates from invasive bighead carp Hypophthalmichthys nobilis and silver carp Hypophthalmichthys molitrix. Biol. Conserv. 2015, 183, 77–84. [Google Scholar] [CrossRef]
- Venette, R.C.; Gordon, D.R.; Juzwik, J.; Koch, F.H.; Liebhold, A.M.; Peterson, R.K.D.; Sing, S.E.; Yemshanov, D. Early Intervention Strategies for Invasive Species Management: Connections Between Risk Assessment, Prevention Efforts, Eradication, and Other Rapid Responses. In Invasive Species in Forests and Rangelands of the United States: A Comprehensive Science Synthesis for the United States Forest Sector; Poland, T.M., Patel-Weynand, T., Finch, D.M., Miniat, C.F., Hayes, D.C., Lopez, V.M., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 111–131. [Google Scholar]
- Reaser, J.K.; Burgiel, S.W.; Kirkey, J.; Brantley, K.A.; Veatch, S.D.; Burgos-Rodríguez, J. The early detection of and rapid response (EDRR) to invasive species: A conceptual framework and federal capacities assessment. Biol. Invasions 2020, 22, 1–19. [Google Scholar] [CrossRef]
- Bass, D.; Christison, K.W.; Stentiford, G.D.; Cook, L.S.J.; Hartikainen, H. Environmental DNA/RNA for pathogen and parasite detection, surveillance, and ecology. Trends Parasitol. 2023, 39, 285–304. [Google Scholar] [CrossRef] [PubMed]
- Furlan, E.M.; Gleeson, D.; Hardy, C.M.; Duncan, R.P. A framework for estimating the sensitivity of eDNA surveys. Mol. Ecol. Resour. 2016, 16, 641–654. [Google Scholar] [CrossRef] [PubMed]
- Pawlowski, J.; Bruce, K.; Panksep, K.; Aguirre, F.I.; Amalfitano, S.; Apothéloz-Perret-Gentil, L.; Baussant, T.; Bouchez, A.; Carugati, L.; Cermakova, K.; et al. Environmental DNA metabarcoding for benthic monitoring: A review of sediment sampling and DNA extraction methods. Sci. Total Environ. 2022, 818, 151783. [Google Scholar] [CrossRef]
- Blancher, P.; Lefrançois, E.; Rimet, F.; Vasselon, V.; Argillier, C.; Arle, J.; Beja, P.; Boets, P.; Boughaba, J.; Chauvin, C.; et al. A strategy for successful integration of DNA-based methods in aquatic monitoring. Metabarcoding Metagenomics 2022, 6, e85652. [Google Scholar] [CrossRef]
- Bruce, K.; Blackman, R.; Bourlat, S.J.; Hellström, A.M.; Bakker, J.; Bista, I.; Bohmann, K.; Bouchez, A.; Brys, R.; Clark, K.; et al. A Practical Guide to DNA-Based Methods for Biodiversity Assessment; Pensoft Advanced Books: Sofia, Bulgaria, 2021; Volume 1, 90p. [Google Scholar]
- Blackman, R.C.; Ling, K.K.S.; Harper, L.R.; Shum, P.; Hanfling, B.; Lawson-Handley, L. Targeted and passive environmental DNA approaches outperform established methods for detection of quagga mussels, Dreissena rostriformis bugensis in flowing water. Ecol. Evol. 2020, 10, 13248–13259. [Google Scholar] [CrossRef]
- Greco, M.; Lejzerowicz, F.; Reo, E.; Caruso, A.; Maccotta, A.; Coccioni, R.; Pawlowski, J.; Frontalini, F. Environmental RNA outperforms eDNA metabarcoding in assessing impact of marine pollution: A chromium-spiked mesocosm test. Chemosphere 2022, 298, 134239. [Google Scholar] [CrossRef]
- Thomsen, P.F.; Willerslev, E. Environmental DNA—An emerging tool in conservation for monitoring past and present biodiversity. Biol. Conserv. 2015, 183, 4–18. [Google Scholar] [CrossRef]
- Pascher, K.; Švara, V.; Jungmeier, M. Environmental DNA-Based Methods in Biodiversity Monitoring of Protected Areas: Application Range, Limitations, and Needs. Diversity 2022, 14, 463. [Google Scholar] [CrossRef]
- Fediajevaite, J.; Priestley, V.; Arnold, R.; Savolainen, V.A.-O. Meta-analysis shows that environmental DNA outperforms traditional surveys, but warrants better reporting standards. Ecol. Evol. 2021, 11, 4803–4815. [Google Scholar] [CrossRef]
- Hänfling, B.; Lawson-Handley, L.; Lunt, D.H.; Shum, P.; Winfield, I.J.; Read, D.S. A Review of Recent Advances in Genetic Methods to Identify Improvements in CAMERAS Partners Monitoring Activities; University of Hull: Riding, UK, 2017. [Google Scholar]
- Suarez-Bregua, P.; Álvarez-González, M.; Parsons, K.M.; Rotllant, J.; Pierce, G.J.; Saavedra, C. Environmental DNA (eDNA) for monitoring marine mammals: Challenges and opportunities. Front. Mar. Sci. 2022, 9, 987774. [Google Scholar] [CrossRef]
- Sahu, A.; Kumar, N.; Pal Singh, C.; Singh, M. Environmental DNA (eDNA): Powerful technique for biodiversity conservation. J. Nat. Conserv. 2023, 71, 126325. [Google Scholar] [CrossRef]
- Ficetola, G.F.; Miaud, C.; Pompanon, F.; Taberlet, P. Species detection using environmental DNA from water samples. Biol. Lett. 2008, 4, 423–425. [Google Scholar] [CrossRef] [PubMed]
- Jeunen, G.-J.; Lipinskaya, T.; Gajduchenko, H.; Golovenchik, V.; Moroz, M.; Rizevsky, V.; Semenchenko, V.; Gemmell, N.J. Environmental DNA (eDNA) metabarcoding surveys show evidence of non-indigenous freshwater species invasion to new parts of Eastern Europe. Metabarcoding Metagenomics 2022, 6, 171–186. [Google Scholar] [CrossRef]
- Smart, A.S.; Tingley, R.; Weeks, A.R.; van Rooyen, A.R.; McCarthy, M.A. Environmental DNA sampling is more sensitive than a traditional survey technique for detecting an aquatic invader. Ecol. Appl. 2015, 25, 1944–1952. [Google Scholar] [CrossRef]
- Ruppert, K.M.; Kline, R.J.; Rahman, M.S. Past, present, and future perspectives of environmental DNA (eDNA) metabarcoding: A systematic review in methods, monitoring, and applications of global eDNA. Glob. Ecol. Conserv. 2019, 17, e00547. [Google Scholar] [CrossRef]
- Duarte, S.; Vieira, P.E.; Lavrador, A.S.; Costa, F.O. Status and prospects of marine NIS detection and monitoring through (e)DNA metabarcoding. Sci. Total Environ. 2021, 751, 141729. [Google Scholar] [CrossRef]
- Dejean, T.; Valentini, A.; Miquel, C.; Taberlet, P.; Bellemain, E.; Miaud, C. Improved detection of an alien invasive species through environmental DNA barcoding: The example of the American bullfrog Lithobates catesbeianus. J. Appl. Ecol. 2012, 49, 953–959. [Google Scholar] [CrossRef]
- Takahara, T.; Minamoto, T.; Doi, H. Using environmental DNA to estimate the distribution of an invasive fish species in ponds. PLoS ONE 2013, 8, e56584. [Google Scholar] [CrossRef]
- Piggott, M.P.; Banks, S.C.; Broadhurst, B.T.; Fulton, C.J.; Lintermans, M. Comparison of traditional and environmental DNA survey methods for detecting rare and abundant freshwater fish. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 31, 173–184. [Google Scholar] [CrossRef]
- Brantschen, J.; Blackman, R.C.; Walser, J.C.; Altermatt, F. Environmental DNA gives comparable results to morphology-based indices of macroinvertebrates in a large-scale ecological assessment. PLoS ONE 2021, 16, e0257510. [Google Scholar] [CrossRef] [PubMed]
- Meyer, A.; Boyer, F.; Valentini, A.; Bonin, A.; Ficetola, G.F.; Beisel, J.-N.; Bouquerel, J.; Wagner, P.; Gaboriaud, C.; Leese, F.; et al. Morphological vs. DNA metabarcoding approaches for the evaluation of stream ecological status with benthic invertebrates: Testing different combinations of markers and strategies of data filtering. Mol. Ecol. 2021, 30, 3203–3220. [Google Scholar] [CrossRef] [PubMed]
- Lavrador, A.; Fontes, J.T.; Vieira, P.; Costa, F.; Duarte, S. Compilation, Revision, and Annotation of DNA Barcodes of Marine Invertebrate Non-Indigenous Species (NIS) Occurring in European Coastal Regions. Diversity 2023, 15, 174. [Google Scholar] [CrossRef]
- Duarte, S.; Vieira, P.E.; Leite, B.R.; Teixeira, M.A.L.; Neto, J.M.; Costa, F.O. Macrozoobenthos monitoring in Portuguese transitional waters in the scope of the water framework directive using morphology and DNA metabarcoding. Estuar. Coast. Shelf Sci. 2023, 281, 108207. [Google Scholar] [CrossRef]
- Pereira, C.L.; Gilbert, M.T.P.; Araújo, M.B.; Matias, M.G. Fine-tuning biodiversity assessments: A framework to pair eDNA metabarcoding and morphological approaches. Methods Ecol. Evol. 2021, 12, 2397–2409. [Google Scholar] [CrossRef]
- Deiner, K.; Bik, H.M.; Mächler, E.; Seymour, M.; Lacoursière-Roussel, A.; Altermatt, F.; Creer, S.; Bista, I.; Lodge, D.M.; Vere, N.; et al. Environmental DNA metabarcoding: Transforming how we survey animal and plant communities. Mol. Ecol. 2017, 26, 5872–5895. [Google Scholar] [CrossRef]
- Pochon, X.; Zaiko, A.; Fletcher, L.M.; Laroche, O.; Wood, S.A. Wanted dead or alive? Using metabarcoding of environmental DNA and RNA to distinguish living assemblages for biosecurity applications. PLoS ONE 2017, 12, e0187636. [Google Scholar] [CrossRef]
- Giroux, M.S.; Reichman, J.R.; Langknecht, T.; Burgess, R.M.; Ho, K.T. Environmental RNA as a Tool for Marine Community Biodiversity Assessments. Sci. Rep. 2022, 12, 17782. [Google Scholar] [CrossRef]
- Bowers, H.A.; Pochon, X.; von Ammon, U.; Gemmell, N.; Stanton, J.-A.L.; Jeunen, G.-J.; Sherman, C.D.H.; Zaiko, A. Towards the Optimization of eDNA/eRNA Sampling Technologies for Marine Biosecurity Surveillance. Water 2021, 13, 1113. [Google Scholar] [CrossRef]
- Wood, S.A.; Biessy, L.; Latchford, J.L.; Zaiko, A.; von Ammon, U.; Audrezet, F.; Cristescu, M.E.; Pochon, X. Release and degradation of environmental DNA and RNA in a marine system. Sci. Total Environ. 2020, 704, 135314. [Google Scholar] [CrossRef]
- Klymus, K.E.; Merkes, C.M.; Allison, M.J.; Goldberg, C.S.; Helbing, C.C.; Hunter, M.E.; Jackson, C.A.; Lance, R.F.; Mangan, A.M.; Monroe, E.M.; et al. Reporting the limits of detection and quantification for environmental DNA assays. Environ. DNA 2020, 2, 271–282. [Google Scholar] [CrossRef]
- Dougherty, M.M.; Larson, E.R.; Renshaw, M.A.; Gantz, C.A.; Egan, S.P.; Erickson, D.M.; Lodge, D.M. Environmental DNA (eDNA) detects the invasive rusty crayfish Orconectes rusticus at low abundances. J. Appl. Ecol. 2016, 53, 722–732. [Google Scholar] [CrossRef] [PubMed]
- Tamburini, M.; Keppel, E.; Marchini, A.; Repetto, M.F.; Ruiz, G.M.; Ferrario, J.; Occhipinti-Ambrogi, A. Monitoring Non-indigenous Species in Port Habitats: First Application of a Standardized North American Protocol in the Mediterranean Sea. Front. Mar. Sci. 2021, 8, 904. [Google Scholar] [CrossRef]
- Davison, P.I.; Falcou-Préfol, M.; Copp, G.H.; Davies, G.D.; Vilizzi, L.; Créach, V. Is it absent or is it present? Detection of a non-native fish to inform management decisions using a new highly-sensitive eDNA protocol. Biol. Invasions 2019, 21, 2549–2560. [Google Scholar] [CrossRef]
- Wozney, K.M.; Wilson, C.C. Quantitative PCR multiplexes for simultaneous multispecies detection of Asian carp eDNA. J. Great Lakes Res. 2017, 43, 771–776. [Google Scholar] [CrossRef]
- Hernandez, C.; Bougas, B.; Perreault-Payette, A.; Simard, A.; Côté, G.; Bernatchez, L. 60 specific eDNA qPCR assays to detect invasive, threatened, and exploited freshwater vertebrates and invertebrates in Eastern Canada. Environ. DNA 2020, 2, 373–386. [Google Scholar] [CrossRef]
- Hillis, D.M.; Allard, M.W.; Miyamoto, M.M. Analysis of DNA-Sequence Data—Phylogenetic Inference. Mol. Evol. Prod. Biochem. Data 1993, 224, 456–487. [Google Scholar]
- Emerson, J.B.; Adams, R.I.; Román, C.M.B.; Brooks, B.; Coil, D.A.; Dahlhausen, K.; Ganz, H.H.; Hartmann, E.M.; Hsu, T.; Justice, N.B.; et al. Schrödinger’s microbes: Tools for distinguishing the living from the dead in microbial ecosystems. Microbiome 2017, 5, 86. [Google Scholar] [CrossRef]
- Golpayegani, A.; Douraghi, M.; Rezaei, F.; Alimohammadi, M.; Nodehi, R.N. Propidium monoazide-quantitative polymerase chain reaction (PMA-qPCR) assay for rapid detection of viable and viable but non-culturable (VBNC) Pseudomonas aeruginosa in swimming pools. J. Environ. Health Sci. Eng. 2019, 17, 407–416. [Google Scholar] [CrossRef]
- Lee, A.S.; Lamanna, O.K.; Ishida, K.; Hill, E.; Nguyen, A.; Hsieh, M.H. A Novel Propidium Monoazide-Based PCR Assay Can Measure Viable Uropathogenic E. coli In Vitro and In Vivo. Front. Cell. Infect. Microbiol. 2022, 12, 40. [Google Scholar] [CrossRef]
- Hirohara, T.; Tsuri, K.; Miyagawa, K.; Paine, R.T.R.; Yamanaka, H. The Application of PMA (Propidium Monoazide) to Different Target Sequence Lengths of Zebrafish eDNA: A New Approach Aimed Toward Improving Environmental DNA Ecology and Biological Surveillance. Front. Ecol. Evol. 2021, 9, 632973. [Google Scholar] [CrossRef]
- Cristescu, M.E. From barcoding single individuals to metabarcoding biological communities: Towards an integrative approach to the study of global biodiversity. Trends Ecol. Evol. 2014, 29, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Valentini, A.; Taberlet, P.; Miaud, C.; Civade, R.; Herder, J.; Thomsen, P.F.; Bellemain, E.; Besnard, A.; Coissac, E.; Boyer, F.; et al. Next-generation monitoring of aquatic biodiversity using environmental DNA metabarcoding. Mol. Ecol. 2016, 25, 929–942. [Google Scholar] [CrossRef] [PubMed]
- Couton, M.A.-O.; Lévêque, L.; Daguin-Thiébaut, C.A.-O.; Comtet, T.; Viard, F.A.-O. Water eDNA metabarcoding is effective in detecting non-native species in marinas, but detection errors still hinder its use for passive monitoring. Biofouling 2022, 38, 367–383. [Google Scholar] [CrossRef] [PubMed]
- Holman, L.E.; de Bruyn, M.; Creer, S.; Carvalho, G.; Robidart, J.; Rius, M. Detection of introduced and resident marine species using environmental DNA metabarcoding of sediment and water. Sci. Rep. 2019, 9, 11559. [Google Scholar] [CrossRef]
- Pearman, J.; Ammon, U.; Laroche, O.; Zaiko, A.; Wood, S.; Zubia, M.; Planes, S.; Pochon, X. Metabarcoding as a tool to enhance marine surveillance of nonindigenous species in tropical harbors: A case study in Tahiti. Environ. DNA 2021, 3, 173–189. [Google Scholar] [CrossRef]
- King, A.C.; Krieg, R.; Weston, A.; Zenker, A.K. Using eDNA to simultaneously detect the distribution of native and invasive crayfish within an entire country. J. Environ. Manag. 2022, 302, 113929. [Google Scholar] [CrossRef]
- van den Heuvel-Greve, M.J.; van den Brink, A.M.; Glorius, S.T.; de Groot, G.A.; Laros, I.; Renaud, P.E.; Pettersen, R.; Węsławski, J.M.; Kuklinski, P.; Murk, A.J. Early detection of marine non-indigenous species on Svalbard by DNA metabarcoding of sediment. Polar Biol. 2021, 44, 653–665. [Google Scholar] [CrossRef]
- Westfall, K.M.; Therriault, T.W.; Abbott, C.L. Targeted next-generation sequencing of environmental DNA improves detection of invasive European green crab (Carcinus maenas). Environ. DNA 2022, 4, 440–452. [Google Scholar] [CrossRef]
- Taberlet, P.; Coissac, E.; Pompanon, F.; Brochmann, C.; Willerslev, E. Towards next-generation biodiversity assessment using DNA metabarcoding. Mol. Ecol. 2012, 21, 2045–2050. [Google Scholar] [CrossRef]
- Singer, G.A.C.; Fahner, N.A.; Barnes, J.G.; McCarthy, A.; Hajibabaei, M. Comprehensive biodiversity analysis via ultra-deep patterned flow cell technology: A case study of eDNA metabarcoding seawater. Sci. Rep. 2019, 9, 5991. [Google Scholar] [CrossRef] [PubMed]
- McColl-Gausden, E.F.; Weeks, A.R.; Coleman, R.; Song, S.; Tingley, R. Using hierarchical models to compare the sensitivity of metabarcoding and qPCR for eDNA detection. Ecol. Inform. 2023, 75, 102072. [Google Scholar] [CrossRef]
- Bylemans, J.; Gleeson, D.M.; Duncan, R.P.; Hardy, C.M.; Furlan, E.M. A performance evaluation of targeted eDNA and eDNA metabarcoding analyses for freshwater fishes. Environ. DNA 2019, 1, 402–414. [Google Scholar] [CrossRef]
- Knudsen, S.W.; Hesselsøe, M.; Thaulow, J.; Agersnap, S.; Hansen, B.K.; Jacobsen, M.W.; Bekkevold, D.; Jensen, S.K.S.; Møller, P.R.; Andersen, J.H. Monitoring of environmental DNA from nonindigenous species of algae, dinoflagellates and animals in the North East Atlantic. Sci. Total Environ. 2022, 821, 153093. [Google Scholar] [CrossRef]
- Hatfield, R.G.; Batista, F.M.; Bean, T.P.; Fonseca, V.G.; Santos, A.; Turner, A.D.; Lewis, A.; Dean, K.J.; Martinez-Urtaza, J. The Application of Nanopore Sequencing Technology to the Study of Dinoflagellates: A Proof of Concept Study for Rapid Sequence-Based Discrimination of Potentially Harmful Algae. Front. Microbiol. 2020, 11, 844. [Google Scholar] [CrossRef]
- Egeter, B.; Veríssimo, J.; Lopes-Lima, M.; Chaves, C.; Pinto, J.; Riccardi, N.; Beja, P.; Fonseca, N.A. Speeding up the detection of invasive bivalve species using environmental DNA: A Nanopore and Illumina sequencing comparison. Mol. Ecol. Resour. 2022, 22, 2232–2247. [Google Scholar] [CrossRef] [PubMed]
- Hanfling, B.; Lawson Handley, L.; Read, D.S.; Hahn, C.; Li, J.; Nichols, P.; Blackman, R.C.; Oliver, A.; Winfield, I.J. Environmental DNA metabarcoding of lake fish communities reflects long-term data from established survey methods. Mol. Ecol. 2016, 25, 3101–3119. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.A.; Pochon, X.; Laroche, O.; von Ammon, U.; Adamson, J.; Zaiko, A. A comparison of droplet digital polymerase chain reaction (PCR), quantitative PCR and metabarcoding for species-specific detection in environmental DNA. Mol. Ecol. Resour. 2019, 19, 1407–1419. [Google Scholar] [CrossRef]
- Jian, C.; Luukkonen, P.; Yki-Järvinen, H.; Salonen, A.; Korpela, K. Quantitative PCR provides a simple and accessible method for quantitative microbiota profiling. PLoS ONE 2020, 15, e0227285. [Google Scholar] [CrossRef]
- Ushio, M.; Murakami, H.; Masuda, R.; Sado, T.; Miya, M.; Sakurai, S.; Yamanaka, H.; Minamoto, T.; Kondoh, M. Quantitative monitoring of multispecies fish environmental DNA using high-throughput sequencing. Metabarcoding Metagenomics 2018, 2, e23297. [Google Scholar]
- Pont, D.; Meulenbroek, P.; Bammer, V.; Dejean, T.; Erős, T.; Jean, P.; Lenhardt, M.; Nagel, C.; Pekarik, L.; Schabuss, M.; et al. Quantitative monitoring of diverse fish communities on a large scale combining eDNA metabarcoding and qPCR. Mol. Ecol. Resour. 2023, 23, 396–409. [Google Scholar] [CrossRef]
- Yu, Z.; Ito, S.I.; Wong, M.K.; Yoshizawa, S.; Inoue, J.; Itoh, S.; Yukami, R.; Ishikawa, K.; Guo, C.; Ijichi, M.; et al. Comparison of species-specific qPCR and metabarcoding methods to detect small pelagic fish distribution from open ocean environmental DNA. PLoS ONE 2022, 17, e0273670. [Google Scholar] [CrossRef]
- Yates, M.C.; Glaser, D.M.; Post, J.R.; Cristescu, M.E.; Fraser, D.J.; Derry, A.M. The relationship between eDNA particle concentration and organism abundance in nature is strengthened by allometric scaling. Mol. Ecol. 2021, 30, 3068–3082. [Google Scholar] [CrossRef]
- Fonseca, V.G. Pitfalls in relative abundance estimation using eDNA metabarcoding. Mol. Ecol. Resour. 2018, 18, 923–926. [Google Scholar] [CrossRef]
- Krehenwinkel, H.; Wolf, M.; Lim, J.Y.; Rominger, A.J.; Simison, W.B.; Gillespie, R.G. Estimating and mitigating amplification bias in qualitative and quantitative arthropod metabarcoding. Sci. Rep. 2017, 7, 17668. [Google Scholar] [CrossRef]
- DaCosta, J.M.; Sorenson, M.D. Amplification biases and consistent recovery of loci in a double-digest RAD-seq protocol. PLoS ONE 2014, 9, e106713. [Google Scholar] [CrossRef]
- Amarasinghe, S.L.; Su, S.; Dong, X.; Zappia, L.; Ritchie, M.E.; Gouil, Q. Opportunities and challenges in long-read sequencing data analysis. Genome Biol. 2020, 21, 30. [Google Scholar] [CrossRef]
- Wilder, M.L.; Farrell, J.M.; Green, H.C. Estimating eDNA shedding and decay rates for muskellunge in early stages of development. Environ. DNA 2023, 5, 251–263. [Google Scholar] [CrossRef]
- Lamb, P.D.; Fonseca, V.G.; Maxwell, D.L.; Nnanatu, C.C. Systematic review and meta-analysis: Water type and temperature affect environmental DNA decay. Mol. Ecol. Resour. 2022, 22, 2494–2505. [Google Scholar] [CrossRef]
- Walsh, J.R.; Pedersen, E.J.; Vander Zanden, M.J. Detecting species at low densities: A new theoretical framework and an empirical test on an invasive zooplankton. Ecosphere 2018, 9, e02475. [Google Scholar] [CrossRef]
- Urban, P.; Bekkevold, D.; Degel, H.; Hansen, B.K.; Jacobsen, M.W.; Nielsen, A.; Nielsen, E.E. Scaling from eDNA to biomass: Controlling allometric relationships improves precision in bycatch estimation. ICES J. Mar. Sci. 2023, 75, fsad027. [Google Scholar] [CrossRef]
- Holman, L.E.; Chng, Y.; Rius, M. How does eDNA decay affect metabarcoding experiments? Environ. DNA 2022, 4, 108–116. [Google Scholar] [CrossRef]
- Jeunen, G.-J.; Lamare, M.D.; Knapp, M.; Spencer, H.G.; Taylor, H.R.; Stat, M.; Bunce, M.; Gemmell, N.J. Water stratification in the marine biome restricts vertical environmental DNA (eDNA) signal dispersal. Environ. DNA 2020, 2, 99–111. [Google Scholar] [CrossRef]
- Alzate, A.; Onstein, R.E. Understanding the relationship between dispersal and range size. Ecol. Lett. 2022, 25, 2303–2323. [Google Scholar] [CrossRef] [PubMed]
- Gottschling, M.; Czech, L.; Mahe, F.; Adl, S.; Dunthorn, M. The Windblown: Possible Explanations for Dinophyte DNA in Forest Soils. J. Eukaryot Microbiol. 2021, 68, e12833. [Google Scholar] [CrossRef] [PubMed]
- Pilliod, D.S.; Goldberg, C.S.; Arkle, R.S.; Waits, L.P. Factors influencing detection of eDNA from a stream-dwelling amphibian. Mol. Ecol. Resour. 2014, 14, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Pont, D.; Rocle, M.; Valentini, A.; Civade, R.; Jean, P.; Maire, A.; Roset, N.; Schabuss, M.; Zornig, H.; Dejean, T. Environmental DNA reveals quantitative patterns of fish biodiversity in large rivers despite its downstream transportation. Sci. Rep. 2018, 8, 10361. [Google Scholar] [CrossRef]
- Deiner, K.; Altermatt, F. Transport Distance of Invertebrate Environmental DNA in a Natural River. PLoS ONE 2014, 9, e88786. [Google Scholar] [CrossRef]
- Jane, S.F.; Wilcox, T.M.; McKelvey, K.S.; Young, M.K.; Schwartz, M.K.; Lowe, W.H.; Letcher, B.H.; Whiteley, A.R. Distance, flow and PCR inhibition: eDNA dynamics in two headwater streams. Mol. Ecol. Resour. 2015, 15, 216–227. [Google Scholar] [CrossRef]
- Nagarajan, R.P.; Bedwell, M.; Holmes, A.E.; Sanches, T.; Acuña, S.; Baerwald, M.; Barnes, M.A.; Blankenship, S.; Connon, R.E.; Deiner, K.; et al. Environmental DNA Methods for Ecological Monitoring and Biodiversity Assessment in Estuaries. Estuaries Coasts 2022, 45, 2254–2273. [Google Scholar] [CrossRef]
- Andruszkiewicz Allan, E.; Zhang, W.G.; C Lavery, A.; F Govindarajan, A. Environmental DNA shedding and decay rates from diverse animal forms and thermal regimes. Environ. DNA 2021, 3, 492–514. [Google Scholar] [CrossRef]
- Kelly, R.P.; Gallego, R.; Jacobs-Palmer, E. The effect of tides on nearshore environmental DNA. PeerJ 2018, 6, e4521. [Google Scholar] [CrossRef] [PubMed]
- Baker, C.S.; Steel, D.; Nieukirk, S.; Klinck, H. Environmental DNA (eDNA) From the Wake of the Whales: Droplet Digital PCR for Detection and Species Identification. Front. Mar. Sci. 2018, 5, 133. [Google Scholar] [CrossRef]
- Burian, A.; Mauvisseau, Q.; Bulling, M.; Domisch, S.; Qian, S.; Sweet, M. Improving the reliability of eDNA data interpretation. Mol. Ecol. Resour. 2021, 21, 1422–1433. [Google Scholar] [CrossRef]
- Hinz, S.; Coston-Guarini, J.; Marnane, M.; Guarini, J.-M. Evaluating eDNA for Use within Marine Environmental Impact Assessments. J. Mar. Sci. Eng. 2022, 10, 375. [Google Scholar] [CrossRef]
- Port, J.A.; O’Donnell, J.L.; Romero-Maraccini, O.C.; Leary, P.R.; Litvin, S.Y.; Nickols, K.J.; Yamahara, K.M.; Kelly, R.P. Assessing vertebrate biodiversity in a kelp forest ecosystem using environmental DNA. Mol. Ecol. 2016, 25, 527–541. [Google Scholar] [CrossRef]
- Dunker, K.J.; Sepulveda, A.J.; Massengill, R.L.; Olsen, J.B.; Russ, O.L.; Wenburg, J.K.; Antonovich, A. Potential of Environmental DNA to Evaluate Northern Pike (Esox lucius) Eradication Efforts: An Experimental Test and Case Study. PLoS ONE 2016, 11, e0162277. [Google Scholar] [CrossRef]
- Salter, I. Seasonal variability in the persistence of dissolved environmental DNA (eDNA) in a marine system: The role of microbial nutrient limitation. PLoS ONE 2018, 13, e0192409. [Google Scholar] [CrossRef]
- Barnes, M.A.; Turner, C.R. The ecology of environmental DNA and implications for conservation genetics. Conserv. Genet. 2016, 17, 1–17. [Google Scholar] [CrossRef]
- McCartin, L.J.; Vohsen, S.A.; Ambrose, S.W.; Layden, M.; McFadden, C.S.; Cordes, E.E.; McDermott, J.M.; Herrera, S. Temperature Controls eDNA Persistence across Physicochemical Conditions in Seawater. Environ. Sci. Technol. 2022, 56, 8629–8639. [Google Scholar] [CrossRef]
- Piaggio, A.J.; Engeman, R.M.; Hopken, M.W.; Humphrey, J.S.; Keacher, K.L.; Bruce, W.E.; Avery, M.L. Detecting an elusive invasive species: A diagnostic PCR to detect Burmese python in Florida waters and an assessment of persistence of environmental DNA. Mol. Ecol. Resour. 2014, 14, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, K.J.; Soluk, D.A.; Maestas, S.E.M.; Britten, H.B. Persistence and accumulation of environmental DNA from an endangered dragonfly. Sci. Rep. 2021, 11, 18987. [Google Scholar] [CrossRef] [PubMed]
- Collins, R.A.; Wangensteen, O.S.; O’Gorman, E.J.; Mariani, S.; Sims, D.W.; Genner, M.J. Persistence of environmental DNA in marine systems. Commun. Biol. 2018, 1, 185. [Google Scholar] [CrossRef] [PubMed]
- Ogata, M.; Masuda, R.; Harino, H.; Sakata, M.K.; Hatakeyama, M.; Yokoyama, K.; Yamashita, Y.; Minamoto, T. Environmental DNA preserved in marine sediment for detecting jellyfish blooms after a tsunami. Sci. Rep. 2021, 11, 16830. [Google Scholar] [CrossRef]
- Turner, C.R.; Uy, K.L.; Everhart, R.C. Fish environmental DNA is more concentrated in aquatic sediments than surface water. Biol. Conserv. 2015, 183, 93–102. [Google Scholar] [CrossRef]
- Armbrecht, L.; Weber, M.E.; Raymo, M.E.; Peck, V.L.; Williams, T.; Warnock, J.; Kato, Y.; Hernández-Almeida, I.; Hoem, F.; Reilly, B.; et al. Ancient marine sediment DNA reveals diatom transition in Antarctica. Nat. Commun. 2022, 13, 5787. [Google Scholar] [CrossRef]
- Ellegaard, M.; Clokie, M.R.J.; Czypionka, T.; Frisch, D.; Godhe, A.; Kremp, A.; Letarov, A.; McGenity, T.J.; Ribeiro, S.; John Anderson, N. Dead or alive: Sediment DNA archives as tools for tracking aquatic evolution and adaptation. Commun. Biol. 2020, 3, 169. [Google Scholar] [CrossRef]
- Armbrecht, L.H. The potential of sedimentary ancient DNA to reconstruct past ocean ecosystems. Oceanography 2020, 33, 116–123. [Google Scholar] [CrossRef]
- Nevers, M.B.; Przybyla-Kelly, K.; Shively, D.; Morris, C.C.; Dickey, J.; Byappanahalli, M.N. Influence of sediment and stream transport on detecting a source of environmental DNA. PLoS ONE 2020, 15, e0244086. [Google Scholar] [CrossRef]
- Brandt, M.I.; Pradillon, F.; Trouche, B.; Henry, N.; Liautard-Haag, C.; Cambon-Bonavita, M.-A.; Cueff-Gauchard, V.; Wincker, P.; Belser, C.; Poulain, J.; et al. Evaluating sediment and water sampling methods for the estimation of deep-sea biodiversity using environmental DNA. Sci. Rep. 2021, 11, 7856. [Google Scholar] [CrossRef]
- Sakata, M.K.; Yamamoto, S.; Gotoh, R.O.; Miya, M.; Yamanaka, H.; Minamoto, T. Sedimentary eDNA provides different information on timescale and fish species composition compared with aqueous eDNA. Environ. DNA 2020, 2, 505–518. [Google Scholar] [CrossRef]
- Herder, J.; Valentini, A.; Bellemain, E.; Dejean, T.; Delft, J.; Thomsen, P.; Taberlet, P. Environmental DNA—A Review of the Possible Applications for the Detection of (Invasive) Species; Stichting RAVON: Nijmegen, The Netherlands, 2014. [Google Scholar]
- Merkes, C.M.; McCalla, S.G.; Jensen, N.R.; Gaikowski, M.P.; Amberg, J.J. Persistence of DNA in Carcasses, Slime and Avian Feces May Affect Interpretation of Environmental DNA Data. PLoS ONE 2014, 9, e113346. [Google Scholar] [CrossRef] [PubMed]
- Bohmann, K.; Evans, A.; Gilbert, M.T.; Carvalho, G.R.; Creer, S.; Knapp, M.; Yu, D.W.; de Bruyn, M. Environmental DNA for wildlife biology and biodiversity monitoring. Trends Ecol. Evol. 2014, 29, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Brys, R.; Halfmaerten, D.; Neyrinck, S.; Mauvisseau, Q.; Auwerx, J.; Sweet, M.; Mergeay, J. Reliable eDNA detection and quantification of the European weather loach (Misgurnus fossilis). J. Fish Biol. 2021, 98, 399–414. [Google Scholar] [CrossRef]
- Sepulveda, A.J.; Hutchins, P.R.; Jackson, C.; Ostberg, C.; Laramie, M.B.; Amberg, J.; Counihan, T.; Hoegh, A.; Pilliod, D.S. A round-robin evaluation of the repeatability and reproducibility of environmental DNA assays for dreissenid mussels. Environ. DNA 2020, 2, 446–459. [Google Scholar] [CrossRef]
- Doi, H.; Minamoto, T.; Takahara, T.; Tsuji, S.; Uchii, K.; Yamamoto, S.; Katano, I.; Yamanaka, H. Compilation of real-time PCR conditions toward the standardization of environmental DNA methods. Ecol. Res. 2021, 36, 379–388. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Zangaro, F.; Saccomanno, B.; Tzafesta, E.; Bozzeda, F.; Specchia, V.; Pinna, M. Current limitations and future prospects of detection and biomonitoring of NIS in the Mediterranean Sea through environmental DNA. NeoBiota 2021, 70, 151–165. [Google Scholar] [CrossRef]
- Thalinger, B.; Deiner, K.; Harper, L.R.; Rees, H.C.; Blackman, R.C.; Sint, D.; Traugott, M.; Goldberg, C.S.; Bruce, K. A validation scale to determine the readiness of environmental DNA assays for routine species monitoring. Environ. DNA 2021, 3, 823–836. [Google Scholar] [CrossRef]
- Evans, N.T.; Shirey, P.D.; Wieringa, J.G.; Mahon, A.R.; Lamberti, G.A. Comparative Cost and Effort of Fish Distribution Detection via Environmental DNA Analysis and Electrofishing. Fisheries 2017, 42, 90–99. [Google Scholar] [CrossRef]
- Lacoursière-Roussel, A.; Deiner, K. Environmental DNA is not the tool by itself. J. Fish Biol. 2021, 98, 383–386. [Google Scholar] [CrossRef] [PubMed]
- Richardson, M.F.; Sherman, C.D.; Lee, R.S.; Bott, N.J.; Hirst, A.J. Multiple dispersal vectors drive range expansion in an invasive marine species. Mol. Ecol. 2016, 25, 5001–5014. [Google Scholar] [CrossRef] [PubMed]
- Deiner, K.; Walser, J.-C.; Mächler, E.; Altermatt, F. Choice of capture and extraction methods affect detection of freshwater biodiversity from environmental DNA. Biol. Conserv. 2015, 183, 53–63. [Google Scholar] [CrossRef]
- Cristescu, M.E.; Hebert, P.D.N. Uses and Misuses of Environmental DNA in Biodiversity Science and Conservation. Annu. Rev. Ecol. Evol. Syst. 2018, 49, 209–230. [Google Scholar] [CrossRef]
- Sepulveda, A.J.; Nelson, N.M.; Jerde, C.L.; Luikart, G. Are Environmental DNA Methods Ready for Aquatic Invasive Species Management? Trends Ecol. Evol. 2020, 35, 668–678. [Google Scholar] [CrossRef]
- Abbott, C.; Coulson, M.; Gagné, N.; Lacoursière-Roussel, A.; Parent, G.J.; Bajno, R.; Dietrich, C.; May-McNally, S. Guidance on the Use of Targeted Environmental DNA (eDNA) Analysis for the Management of Aquatic Invasive Species and Species at Risk. DFO Can. Sci. Advis. Sec. Res. Doc. 2021, 4, 42. [Google Scholar]
- Mychek-Londer, J.G.; Balasingham, K.D.; Heath, D.D. Using environmental DNA metabarcoding to map invasive and native invertebrates in two Great Lakes tributaries. Environ. DNA 2020, 2, 283–297. [Google Scholar] [CrossRef]
- Haugland, R.A.; Siefring, S.; Varma, M.; Oshima, K.H.; Sivaganesan, M.; Cao, Y.; Raith, M.; Griffith, J.; Weisberg, S.B.; Noble, R.T.; et al. Multi-laboratory survey of qPCR enterococci analysis method performance in U.S. coastal and inland surface waters. J. Microbiol. Methods 2016, 123, 114–125. [Google Scholar] [CrossRef]
- NE. Great Crested Newts: Surveys and Mitigation for Development Projects. Natural England. 2015. Available online: https://www.gov.uk/guidance/great-crested-newts-surveys-and-mitigation-for-development-projects#survey-methods (accessed on 14 January 2022).
- MacDonald, A.J.; Sarre, S.D. A framework for developing and validating taxon-specific primers for specimen identification from environmental DNA. Mol. Ecol. Resour. 2017, 17, 708–720. [Google Scholar] [CrossRef]
- Goldberg, C.S.; Turner, C.R.; Deiner, K.; Klymus, K.E.; Thomsen, P.F.; Murphy, M.A.; Spear, S.F.; McKee, A.; Oyler-McCance, S.J.; Cornman, R.S.; et al. Critical considerations for the application of environmental DNA methods to detect aquatic species. Methods Ecol. Evol. 2016, 7, 1299–1307. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fonseca, V.G.; Davison, P.I.; Creach, V.; Stone, D.; Bass, D.; Tidbury, H.J. The Application of eDNA for Monitoring Aquatic Non-Indigenous Species: Practical and Policy Considerations. Diversity 2023, 15, 631. https://doi.org/10.3390/d15050631
Fonseca VG, Davison PI, Creach V, Stone D, Bass D, Tidbury HJ. The Application of eDNA for Monitoring Aquatic Non-Indigenous Species: Practical and Policy Considerations. Diversity. 2023; 15(5):631. https://doi.org/10.3390/d15050631
Chicago/Turabian StyleFonseca, Vera G., Phil I. Davison, Veronique Creach, David Stone, David Bass, and Hannah J. Tidbury. 2023. "The Application of eDNA for Monitoring Aquatic Non-Indigenous Species: Practical and Policy Considerations" Diversity 15, no. 5: 631. https://doi.org/10.3390/d15050631
APA StyleFonseca, V. G., Davison, P. I., Creach, V., Stone, D., Bass, D., & Tidbury, H. J. (2023). The Application of eDNA for Monitoring Aquatic Non-Indigenous Species: Practical and Policy Considerations. Diversity, 15(5), 631. https://doi.org/10.3390/d15050631