
Genomic Diversity Analyses of Some Indigenous Montenegrin Sheep Populations

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
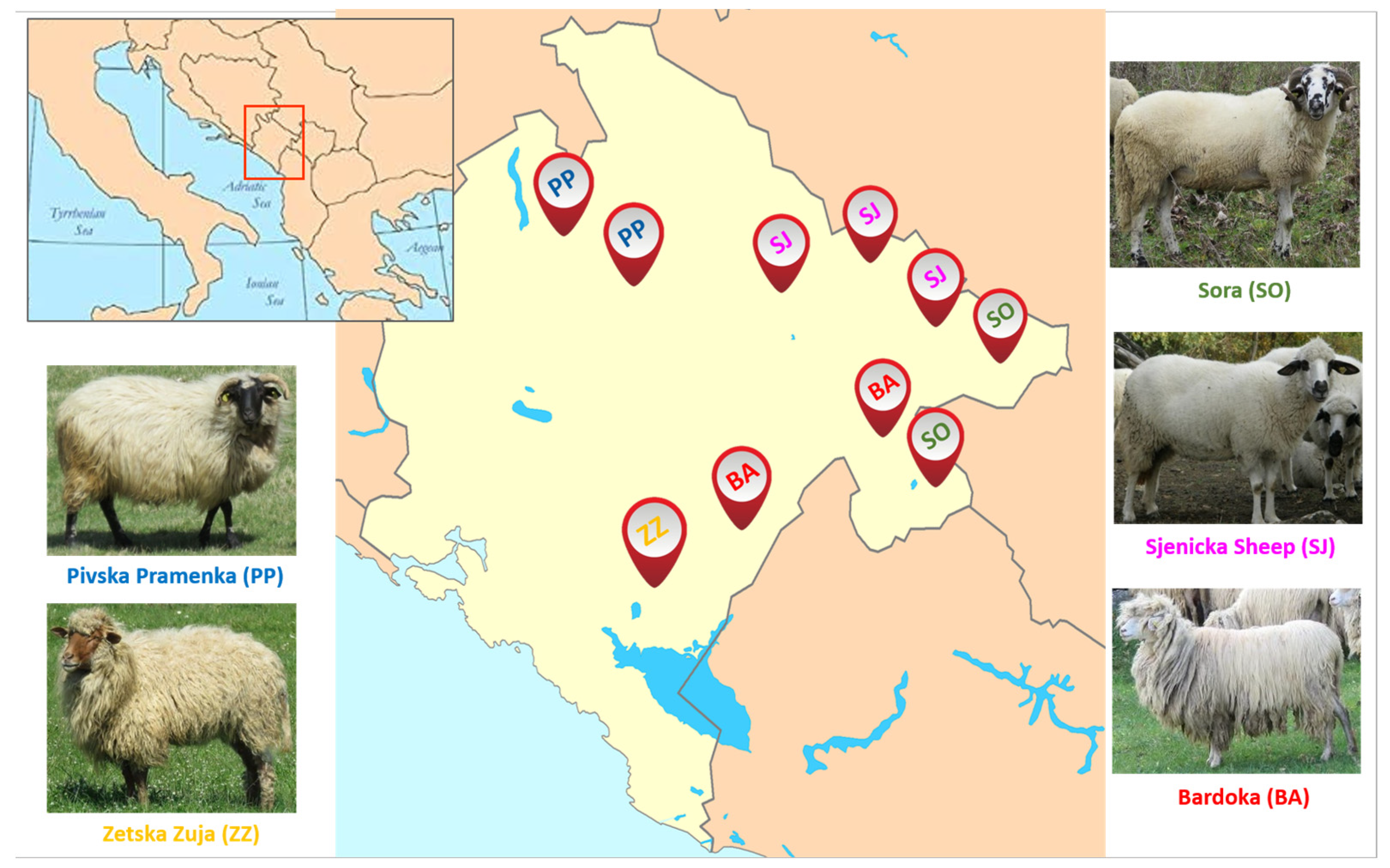
2.1. Animal Resources, SNP Genotyping, and Datasets
2.2. Genetic Diversity Analysis
2.3. Genetic Structure Analysis
3. Results
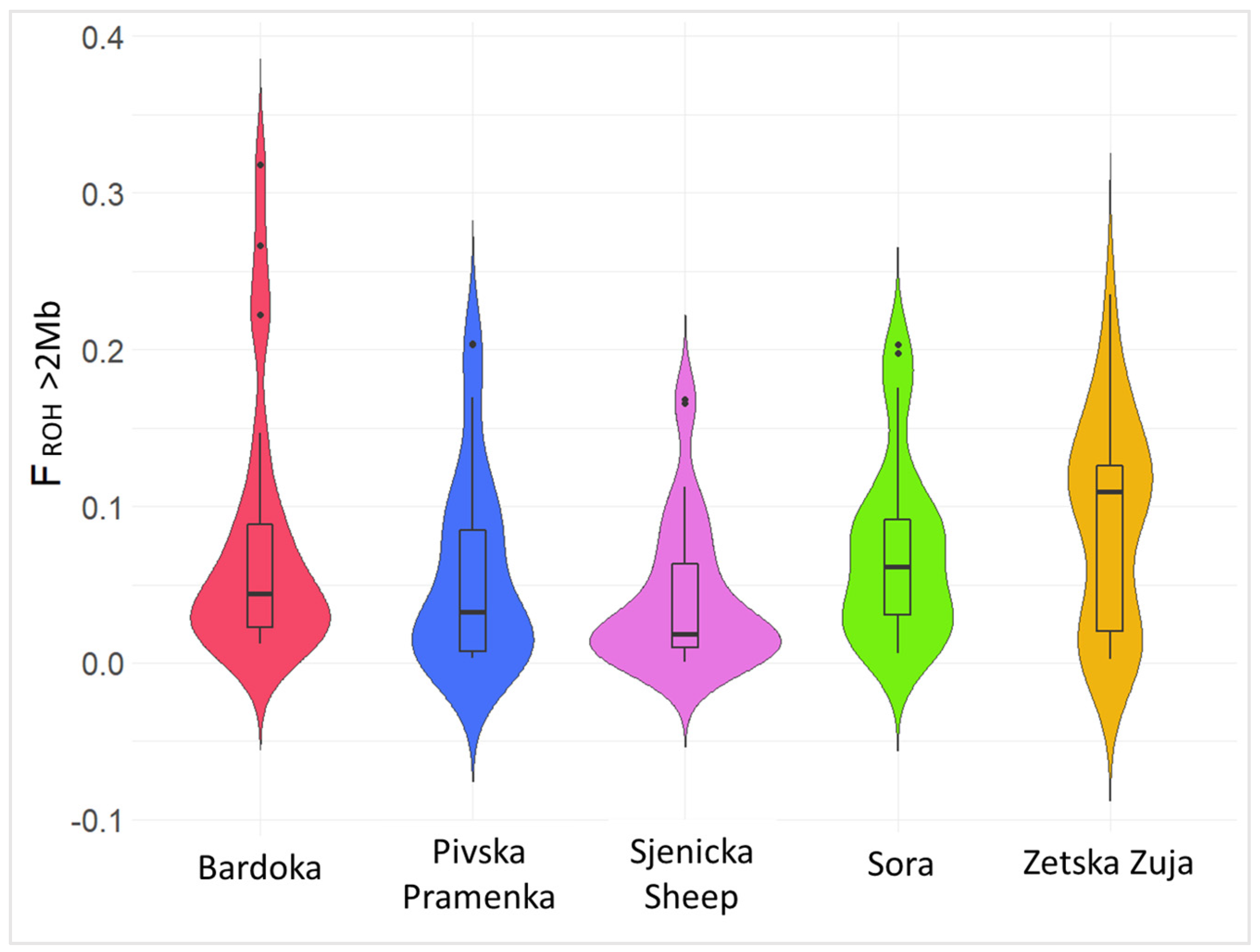
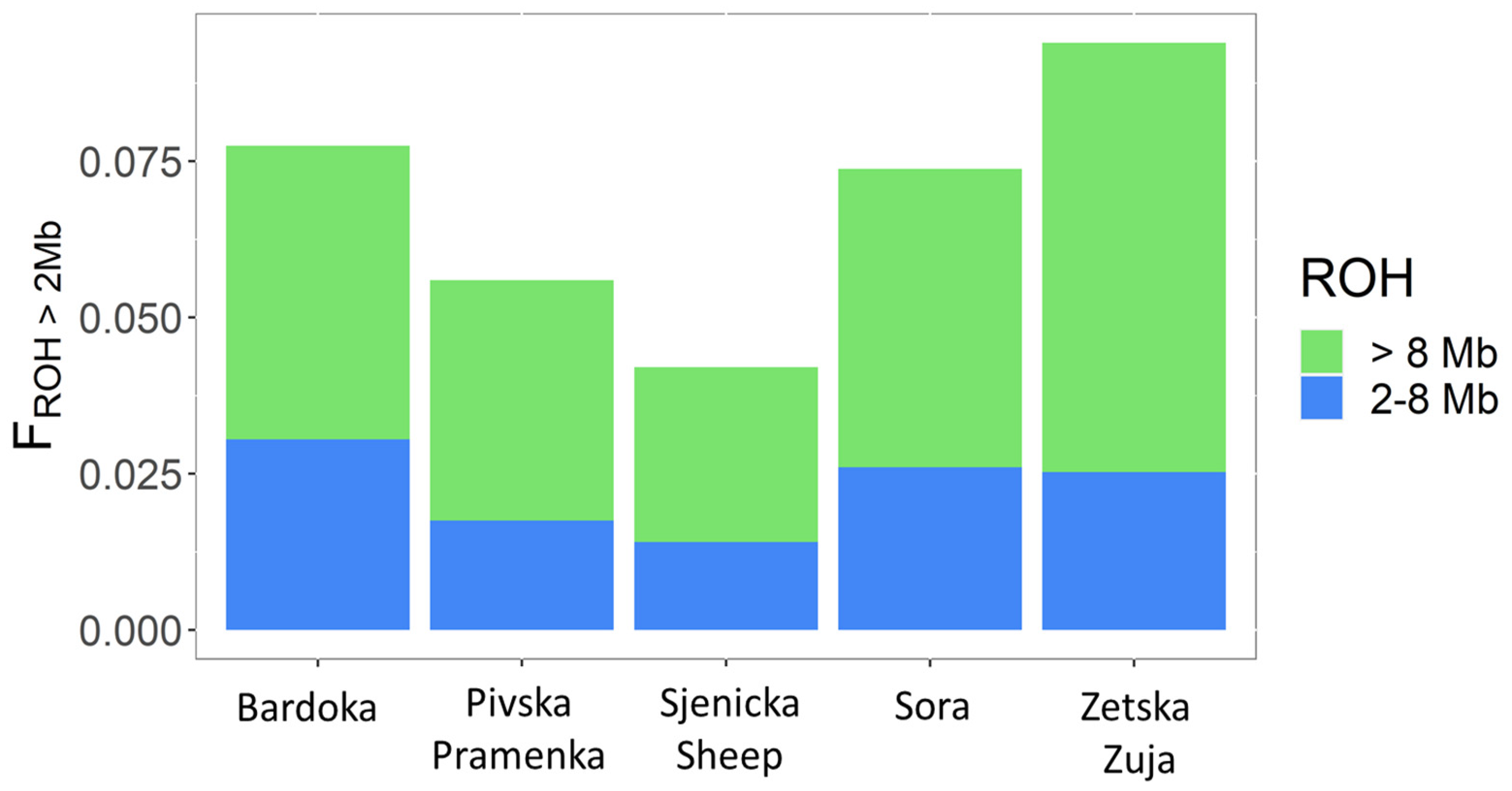
3.1. Analysis of Genetic Diversity
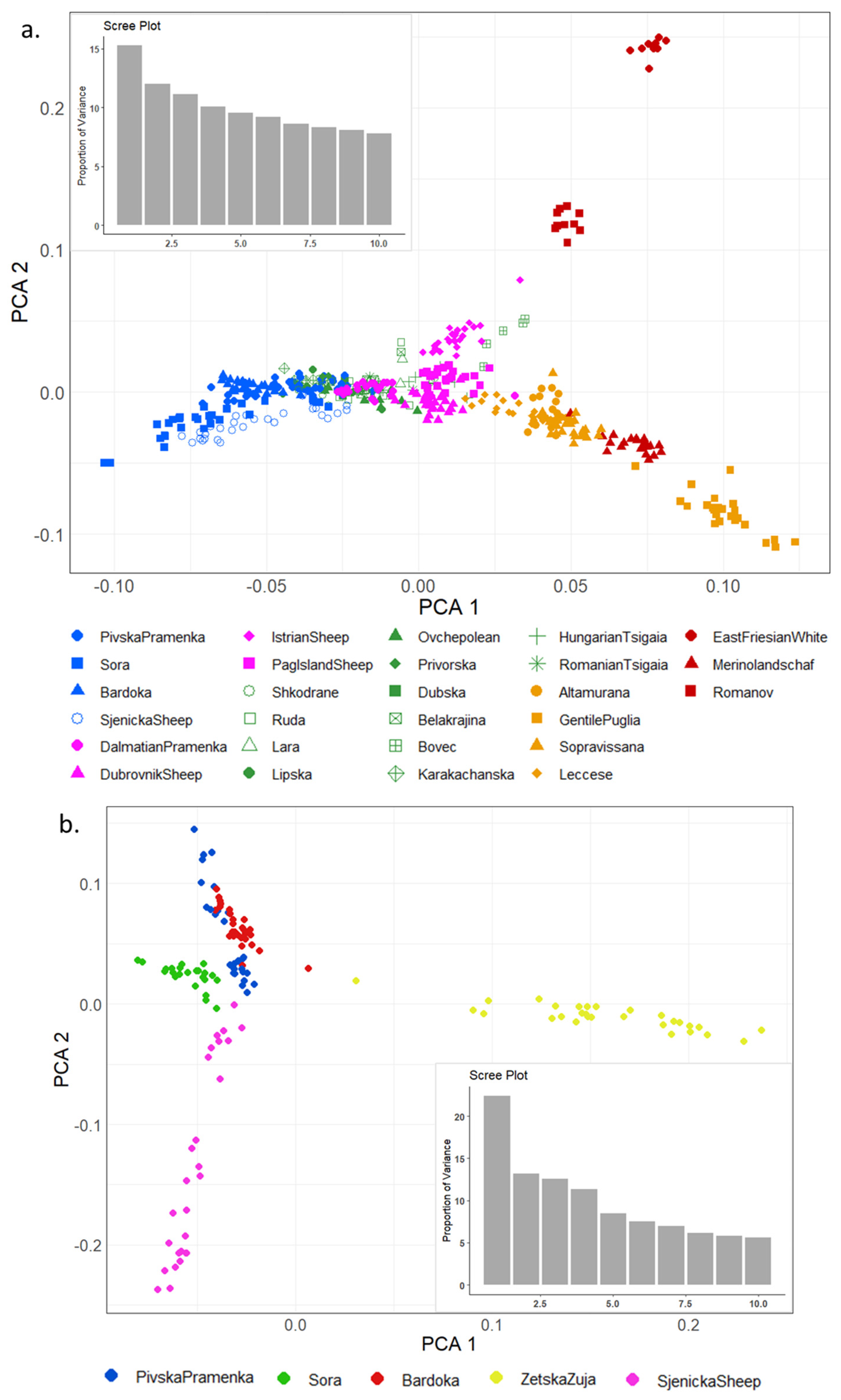
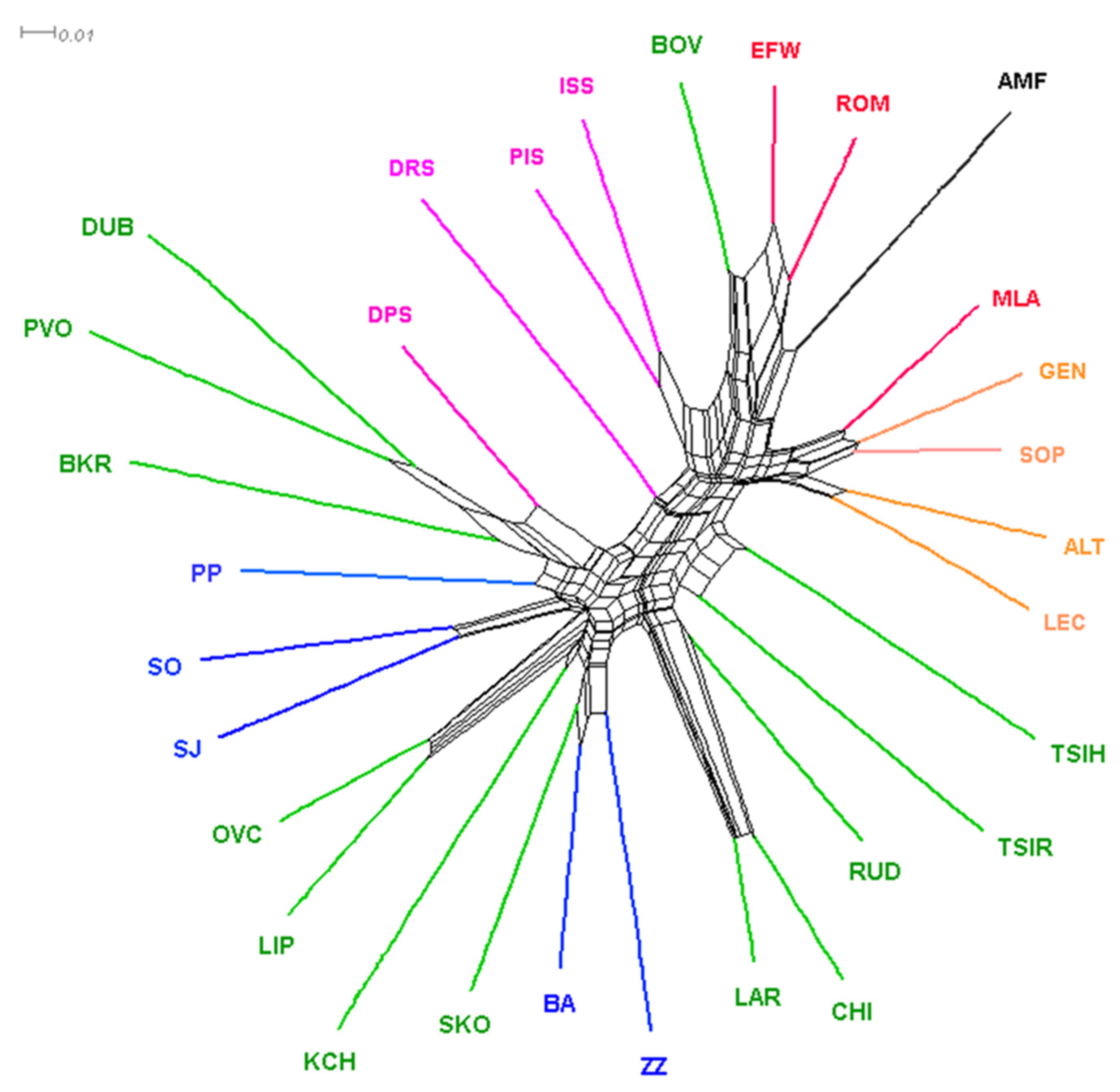
3.2. Population Genetic Differentiation
4. Discussion
4.1. Genetic Diversity, Effective Population Size, and Genome Inbreeding
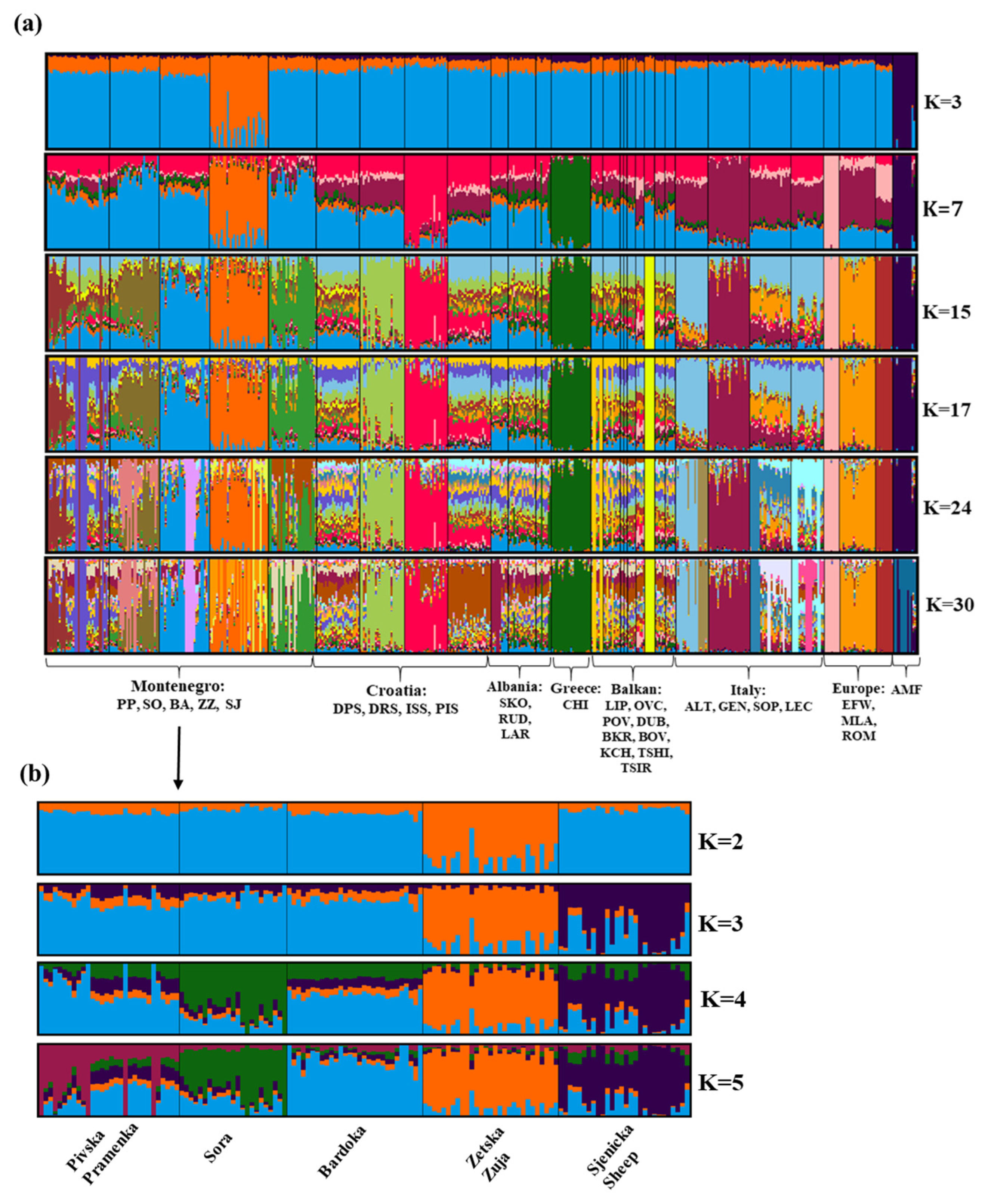
4.2. Population Genetic Structure Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ciani, E.; Cubric-Curik, V.; Simčič, M.; Marković, B.; Esmerov, I.; Ružić-Muslić, D.; Štěpánek, O.; Pieragostini, E.; Sallustio, F.; Arjen Lenstra, J. Genetic structure and admixture in Western Balkans and Central European sheep: Preliminary results from 50K SNP genotypic data. Ital. J. Anim. Sci. 2015, 14, 104–105. [Google Scholar]
- Marković, B.; Knežević, M.; Dubljević, R. Agri-Environmental Policy in Montenegro. In AGRI-Environmental Policy in South-East Europe; Regional Rural Development Standing Working Group in SEE (SWG): Pogradec, Albania, 2018; ISBN 978-608-4760-26-9. [Google Scholar]
- Dubljević, R. Country Pasture/Forage Resource Profiles, Montenegro; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2009; Available online: http://www.fao.org/ag/agp/agpc/doc/counprof/montenegro/montenegro.htm (accessed on 12 October 2022).
- Marković, B.; Marković, M.; Radonjić, D.; Veljić, M. Sustainable sheep and goat production based on local breeds in Montenegro. In Proceedings of the 8th Global Conference on the Conservation of Animal Genetic Resources, Tekirdag, Turkye, 4–8 October 2011; pp. 414–420, ISBN 978-605-87679-4-2. [Google Scholar]
- Marković, B.; Marković, M.; Adzić, N. The farm animal genetic resources of Montenegro. Biotechnol. Anim. Husb. 2007, 23, 1–9. [Google Scholar] [CrossRef]
- Marković, B.; Ivanković, A.; Mirecki, S.; Marković, M. Genetic polymorphism of beta—Lactoglobulin in the three sheep breeds in Montenegro. In Proceedings of the International Symposium on Animal Science, Novi Sad, Serbia, 24–25 October 2015; ISBN 978-86-7520-346-9. [Google Scholar]
- Marković, B.; Marković, M.; Radonjić, D. The morphometric characteristics of indigenous sheep population named Sora. In Proceedings of the 1st International Symposium on Animal Science, Belgrade, Serbia, 8–10 November 2012; pp. 57–64, ISBN 978-86-7834-164-9. [Google Scholar]
- Đokić, M.; Marković, B.; Marković, M.; Mirecki, S.; Veljić, M. Relationship between β-Lactoglobulin Gene Polymorphism and Milk Traits and Milk Composition of Sora Sheep Breed; International Symposium on Animal Science (ISAS): Herceg Novi, Montenegro, 2019; pp. 100–105. [Google Scholar]
- Marković, B.; Dovč, P.; Marković, M.; Radonjić, D.; Adakalić, M.; Simčič, M. Differentiation of some Pramenka sheep breedsbased on morphometric characteristics. Arch. Anim. Breed. 2019, 62, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Bytyqi, H.; Mehmeti, H. Bardhoka Strain. In Catalogue of West Balkan Pramenka Sheep Breed Types: Identification and Conservation of Animal Genetic Resources in South Eastern Europe; Faculty of Agricultural Sciences and Food, University of Skopje: Skopje, Macedonia, 2006. [Google Scholar]
- Đokić, M.; Marković, B.; Gantner, V.; Marković, M. Association of genetic variants of β-lactoglobulin gene with milk traits of Jezeropivska sheep breed. Agric. For. 2020, 66, 15–23. [Google Scholar] [CrossRef]
- Cinkulov, M.; Popovski, Z.; Tanaskovska, B.; Hodžić, A.; Bytyqi, H.; Mehmeti, H.; Margeta, V.; Djedović, R.; Hoda, A.; Trailović, R.; et al. Genetic diversity and structure of the West Balkan Pramenka sheep types as revealed by microsatellite and mitochondrial DNA analysis. J. Anim. Breed. Genet. 2008, 125, 417–426. [Google Scholar] [CrossRef]
- Marković, B.; Marković, M.; Adžić, N. Genetic Resources in Livestock of Montenegro; Montenegrin Academy of Science and Art: Podgorica, Montenegro, 2020; p. 300. ISBN 978-86-7215-470-2. [Google Scholar]
- FAO. Global Plan of Action for Animal Genetic Resources and the Interlaken Declaration; Commission on Genetic Resources for Food and Agriculture; Food and Agriculture Organization of the United Nations: Rome, Italy, 2007; Available online: https://www.fao.org/3/a1404e/a1404e.pdf (accessed on 20 September 2022).
- Ministry of Agriculture and Rural Development. National Program for Conservation and Sustainable Use of Genetic Resources in Agriculture (2008–2013) with Action Plan; Ministry of Agriculture and Rural Development: Podgorica, Montenegro, 2007. [Google Scholar]
- FAO. FAO Animal Production and Health Guidelines; Molecular Genetic Characterization of Animal Genetic Resources. No. 9; FAO: Rome, Italy, 2011; Available online: http://www.fao.org/dad-is (accessed on 9 October 2022).
- Toro, M.A.; Fernández, J.; Caballero, A. Molecular characterization of breeds and its use in conservation. Livestig. Sci. 2009, 120, 174–195. [Google Scholar] [CrossRef]
- Dunisławska, A.; Łachmańska, J.; Sławińska, A.; Siwek, M. Next generation sequencing in animal science—A review. Anim. Sci. Pap. Rep. 2017, 35, 205–224. [Google Scholar]
- Lenstra, J.A.; Groeneveld, L.F.; Eding, H.; Kantanen, J.; Williams, J.L.; Taberlet, P.; Nicolazzi, E.L.; Sölkner, J.; Simianer, H.; Ciani, E.; et al. Molecular tools and analytical approaches for the characterization of farm animal genetic diversity. Anim. Genet. 2012, 43, 483–502. [Google Scholar] [CrossRef]
- Eusebi, P.G.; Martinez, A.; Cortes, O. Genomic Tools for Eective Conservation of Livestock Breed Diversity. Diversity 2020, 12, 2–16. [Google Scholar]
- Kawęcka, A.; Gurgul, A.; Miksza-Cybulska, A. The use of SNP microarrays for biodiversity studies of sheep—A review. Ann. Anim. Sci. 2016, 16, 975–987. [Google Scholar] [CrossRef]
- Davenport, K.M.; Hiemke, C.; McKay, S.D.; Thorne, J.W.; Lewis, R.M.; Taylor, T.M. Genetic structure and admixture in sheep from terminal breeds in the United States. Anim. Genet. 2020, 51, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Mousel, M.R.; Wu, X.; Michal, J.J.; Zhou, X.; Ding, B.; Dodson, M.V.; El-Halawany, N.K.; Lewis, G.S.; Jiang, Z. Genome-Wide Genetic Diversity and Differentially Selected Regions among Suffolk, Rambouillet, Columbia, Polypay, and Targhee Sheep. PLoS ONE 2013, 8, e65942. [Google Scholar] [CrossRef]
- Al-Mamun, H.A.; Clark, S.; Kwan, P.; Gondro, C. Genome-wide linkage disequilibrium and genetic diversity in five populations of Australian domestic sheep. Genet. Sel. Evol. 2015, 47, 2–14. [Google Scholar] [CrossRef]
- Zhao, Y.X.; Yang, J.; Lv, F.H.; Hu, X.J.; Xie, X.L.; Zhang, M.; Li, W.R.; Liu, M.J.; Wang, Y.T.; Li, J.Q.; et al. Genomic reconstruction of the history of native sheep reveals the peopling patterns of nomads and the expansion of early pastoralism in East Asia. Mol. Biol. Evol. 2017, 34, 2380–2395. [Google Scholar] [CrossRef]
- Edea, Z.; Dessie, T.; Dadi, H.; Do, K.-T.; Kim, K.-S. Genetic Diversity and Population Structure of Ethiopian Sheep Populations Revealed by High-Density SNP Markers. Front. Genet. 2017, 8, 218. [Google Scholar] [CrossRef]
- Deniskova, T.E.; Dotsev, A.V.; Selionova, M.I.; Kunz, E.; Medugorac, I.; Reyer, H.; Wimmers, K.; Barbato, M.; Traspov, A.A.; Brem, G.; et al. Population structure and genetic diversity of 25 Russian sheep breeds based on whole-genome genotyping. Genet. Sel. Evol. 2018, 50, 29. [Google Scholar] [CrossRef]
- Deniskova, T.; Dotsev, A.; Lushihina, E.; Shakhin, A.; Kunz, E.; Međugorac, I.; Reyer, H.; Wimmers, K.; Khayatzadeh, N.; Sölkner, J.; et al. Population Structure and Genetic Diversity of Sheep Breeds in the Kyrgyzstan. Front. Genet. 2019, 10, 1311. [Google Scholar] [CrossRef] [PubMed]
- Rochus, C.M.; Tortereau, F.; Plisson-Petit, F.; Restoux, G.; Moreno-Romieux, C.; Tosser-Klopp, G.; Servin, B. Revealing the selection history of adaptive loci using genome-wide scans for selection: An example from domestic sheep. BMC Genom. 2018, 19, 71. [Google Scholar] [CrossRef]
- Rochus, C.M.; Jonas, E.; Johansson, A.M. Population structure of five native sheep breeds of Sweden estimated with high density SNP genotypes. BMC Genet. 2020, 21, 27. [Google Scholar] [CrossRef]
- Grasso, A.N.; Goldberg, V.; Navajas, E.A.; Iriarte, W.; Gimeno, D.; Aguilar, I.; Medrano, J.F.; Rincón, G.; Ciappesoni, G. Genomic variation and population structure detected by single nucleotide polymorphism arrays in Corriedale, Merino and Creole sheep. Genet. Mol. Biol. 2014, 37, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Ciani, E.; Crepaldi, P.; Nicoloso, L.; Lasagna, E.; Sarti, F.M.; Moioli, B.; Napolitano, F.; Carta, A.; Usai, G.; D’Andrea, M.; et al. Genome-wide analysis of Italian sheep diversity reveals a strong geographic pattern and cryptic relationships between breeds. Anim. Genet. 2014, 45, 256–266. [Google Scholar] [CrossRef] [PubMed]
- Beynon, S.E.; Slavov, G.T.; Farré, M.; Sunduimijid, B.; Waddams, K.; Davies, B.; Haresign, W.; Kijas, J.; MacLeod, I.M.; Newbold, C.J.; et al. Population structure and history of the Welsh sheep breeds determined by whole genome genotyping. BMC Genet. 2015, 20, 65. [Google Scholar] [CrossRef] [PubMed]
- Drzaic, I.; Curik, I.; Lukic, B.; Shihabi, M.; Li, M.-H.; Kantanen, J.; Mastrangelo, S.; Ciani, E.; Lenstra, J.A.; Cubric-Curik, V. High-Density Genomic Characterization of Native Croatian Sheep Breeds. Front. Genet. 2022, 13, 940736. [Google Scholar] [CrossRef]
- Adeniyi, O.O.; Simon, R.; Bytyqi, H.; Kugler, W.; Mehmeti, H.; Berisha, K.; Simčić, M.; Magdy, M.; Lühken, G. Capturing Genetic, Diversity and Selection Signatures of the Endangered Kosovar Balusha Sheep Breed. Genes 2022, 13, 866. [Google Scholar] [CrossRef] [PubMed]
- Ahbara, A.; Bahbahani, H.; Almathen, F.; Al Abri, M.; Agoub, M.O.; Abeba, A.; Kebede, A.; Musa, H.H.; Mastrangelo, S.; Pilla, F.; et al. Genome-Wide Variation, Candidate Regions and Genes Associated With Fat Deposition and Tail Morphology in Ethiopian Indigenous Sheep. Front. Genet. 2019, 9, 699. [Google Scholar] [CrossRef] [PubMed]
- Ciani, E.; Lasagna, E.; D’andrea, M.; Alloggio, I.; Marroni, F.; Ceccobelli, S.; Delgado Bermejo, J.V.; Sarti, F.M.; Kijas, J.; Lenstra, J.A.; et al. Merino and Merino-derived sheep breeds: A genome-wide intercontinental study. Genet. Sel. Evol. 2015, 47, 64. [Google Scholar] [CrossRef]
- Kijas, J.W.; Lenstra, J.A.; Hayes, B.; Boitard, S.; Porto Neto, L.R.; San Cristobal, M.; Servin, B.; McCulloch, R.; Whan, V.; Gietzen, K.; et al. Genome-Wide Analysis of the World’s Sheep Breeds Reveals High Levels of Historic Mixture and Strong Recent Selection. PLoS Biol. 2012, 10, e1001258. [Google Scholar] [CrossRef]
- Moradi, M.H.; Nejati-Javaremi, A.; Moradi-Shahrbabak, M.; Dodds, K.G.; McEwan, J.C. Genomic scan of selective sweeps in thin and fat tail sheep breeds for identifying of candidate regions associated with fat deposition. BMC Genet. 2012, 13, 10. [Google Scholar] [CrossRef]
- FAO. The Second Report on the State of the World’s Animal Genetic Resources for Food and Agriculture; Scherf, B.D., Pilling, D., Eds.; FAO Commission on Genetic Resources for Food and Agriculture Assessments: Rome, Italy, 2015; Available online: http://www.fao.org/3/a-i4787e/index.html (accessed on 25 October 2022).
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; De Bakker, P.I.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef]
- Ciani, E.; Mastrangelo, S.; Da Silva, A.; Marroni, F.; Ferenčaković, M.; Ajmone-Marsan, P.; Baird, H.; Barbato, M.; Colli, L.; Delvento, C.; et al. On the Origin of European Sheep as Revealed by the Diversity of the Balkan Breeds and by Optimizing Population-Genetic Analysis Tools. Genet. Sel. Evol. 2020, 52, 25. [Google Scholar] [CrossRef]
- Keenan, K.; McGinnity, P.; Cross, T.F.; Crozier, W.W.; Prodohl, P.A. DiveRsity: An R package for the estimation of population genetics parameters and their associated errors. Methods Ecol. Evol. 2013, 4, 782–788. [Google Scholar] [CrossRef]
- Santiago, E.; Novo, I.; Pardiñas, A.F.; Saura, M.; Wang, J.; Caballero, A. Recent Demographic History Inferred by High-Resolution Analysis of Linkage Disequilibrium. Mol. Biol. Evol. 2020, 37, 3642–3653. [Google Scholar] [CrossRef] [PubMed]
- Ferenčaković, M.; Sölkner, J.; Curik, I. Estimating autozygosity from high-throughput information: Effects of SNP density and genotyping errors. Genet. Sel. Evol. 2013, 45, 42. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016; ISBN 3319242776. [Google Scholar]
- Zheng, X.; Levine, D.; Shen, J.; Gogarten, S.; Laurie, C.; Weir, B. A High-performance Computing Toolset for Relatedness and Principal Component Analysis of SNP Data. Bioinformatics 2012, 28, 3326–3328. [Google Scholar] [CrossRef]
- Nei, M. Genetic distance between populations. Am. Nat. 1972, 106, 283–292. [Google Scholar] [CrossRef]
- Pembleton, L.W.; Cogan, N.O.I.; Forster, J.W. St AMPP: An R Package for Calculation of Genetic Differentiation and Structure of Mixed ploidy Level Populations. Mol. Ecol. Resour. 2013, 13, 946–952. [Google Scholar] [CrossRef]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Alexander David, H.; Novembre, J.; Lange, K. Fast model-based estimation of ancestry in unrelated individuals. Genome Res. 2009, 19, 1655–1664. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Mastrangelo, S.; Portolano, B.; Di Gerlando, R.; Ciampolini, R.; Tolone, M.; Sardina, M.T. Genome-wide analysis in endangered populations: A case study in Barbaresca sheep. Animal 2017, 11, 1107–1116. [Google Scholar] [CrossRef]
- Mitchell, M. An Introduction to Genetic Algorithms; A Bradford Book; The MIT Press: Cambridge, UK; London, UK, 1998. [Google Scholar]
- Novo, I.; Santiago, E.; Caballero, A. The estimates of effective population size based on linkage disequilibrium are virtually unaffected by natural selection. PLoS Genet. 2022, 8, e1009764. [Google Scholar] [CrossRef] [PubMed]
- Lencz, T.; Lambert, C.; DeRosse, P.; Burdick, K.E.; Morgan, T.V.; Kane, J.M.; Kucherlapati, R.; Malhotra, A.K. Runs of homozygosity reveal highly penetrant recessive loci in schizophrenia. Proc. Natl. Acad. Sci. USA 2007, 104, 19942–19947. [Google Scholar] [CrossRef] [PubMed]
- McQuillan, R.; Leutenegger, A.L.; Abdel-Rahman, R.; Franklin, C.S.; Pericic, M.; Barac-Lauc, L.; Smolej-Narancic, N.; Janicijevic, B.; Polasek, O.; Tenesa, A.; et al. Runs of Homozygosity in European Populations. Am. J. Hum. Genet. 2008, 83, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Curik, I.; Ferenčaković, M.; Sölkner, J. Inbreeding and runs of homozygosity: A possible solution to an old problem. Livestig. Sci. 2014, 166, 26–34. [Google Scholar] [CrossRef]
- Marković, M.; Radonjić, D.; Zorc, M.; Đokić, M.; Marković, B. Genetic Diversity of Montenegrin Local Sheep Breeds Based on Microsatellite Markers. Animals 2022, 12, 3029. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breed | N | Ho ± SE | He ± SE | Ar | FIS (CI95%) | Ne0 | Ne5 |
|---|---|---|---|---|---|---|---|
| Bardoka (BA) | 29 | 0.397 ± 0.001 | 0.417 ± 0.001 | 2.074 | 0.049 (0.046 to 0.051) | 81 | 684 |
| Pivska Pramenka (PP) | 30 | 0.411 ± 0.001 | 0.423 ± 0.001 | 2.062 | 0.029 (0.025 to 0.032) | 72 | 72 |
| Sjenicka Sheep (SJ) | 28 | 0.414 ± 0.001 | 0.421 ± 0.001 | 2.145 | 0.016 (0.012 to 0.019) | 47 | 79 |
| Sora (SO) | 23 | 0.392 ± 0.001 | 0.409 ± 0.001 | 2.101 | 0.042 (0.038 to 0.045) | 25 | 5003 |
| Zetska Zuja (ZZ) | 29 | 0.385 ± 0.001 | 0.386 ± 0.001 | 2.136 | 0.001 (−0.002 to 0.0047) | 32 | 92 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Djokic, M.; Drzaic, I.; Shihabi, M.; Markovic, B.; Cubric-Curik, V. Genomic Diversity Analyses of Some Indigenous Montenegrin Sheep Populations. Diversity 2023, 15, 640. https://doi.org/10.3390/d15050640
Djokic M, Drzaic I, Shihabi M, Markovic B, Cubric-Curik V. Genomic Diversity Analyses of Some Indigenous Montenegrin Sheep Populations. Diversity. 2023; 15(5):640. https://doi.org/10.3390/d15050640
Chicago/Turabian StyleDjokic, Milena, Ivana Drzaic, Mario Shihabi, Bozidarka Markovic, and Vlatka Cubric-Curik. 2023. "Genomic Diversity Analyses of Some Indigenous Montenegrin Sheep Populations" Diversity 15, no. 5: 640. https://doi.org/10.3390/d15050640
APA StyleDjokic, M., Drzaic, I., Shihabi, M., Markovic, B., & Cubric-Curik, V. (2023). Genomic Diversity Analyses of Some Indigenous Montenegrin Sheep Populations. Diversity, 15(5), 640. https://doi.org/10.3390/d15050640