
A Theoretical Thermal Tolerance Function for Ectothermic Animals and Its Implications for Identifying Thermal Vulnerability across Large Geographic Scales



Abstract
:1. Introduction
2. Materials and Methods
2.1. Generating a Thermal Death Curve
2.2. Exploring the Thermal Death Curve in Lizards
2.3. Modeling Thermally Deleterious Sites for Urosaurus ornatus
2.4. Geographic Bias in Predictions of Thermally Deleterious Sites for Urosaurus ornatus
2.5. Comparison of Thermal Vulnerability Layers with Known Locations of U. ornatus
3. Results
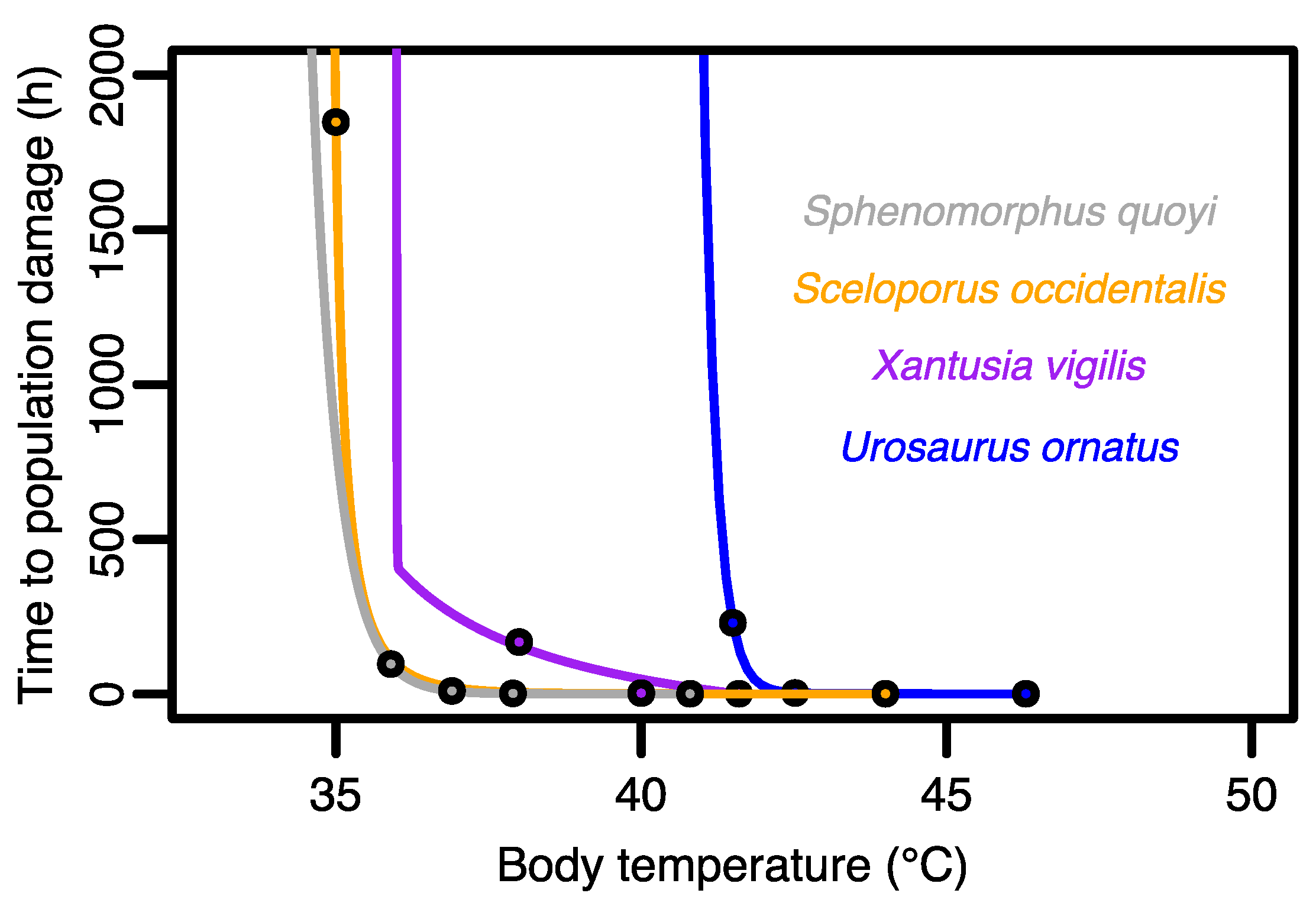
3.1. Exploring the TDC in Four Lizard Species

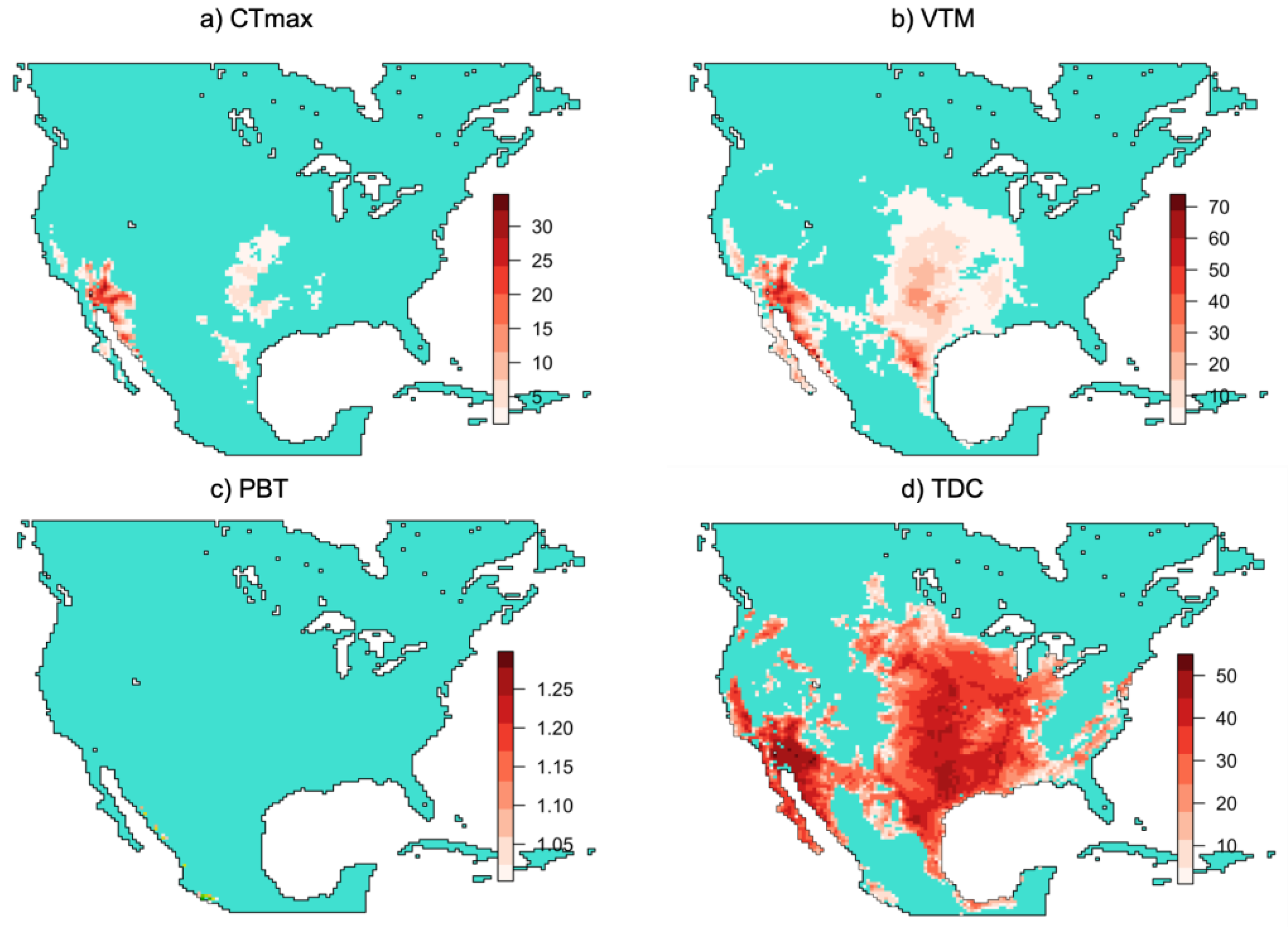
3.2. Geographic Bias in Predictions of Thermally Deleterious Sites for Urosaurus ornatus
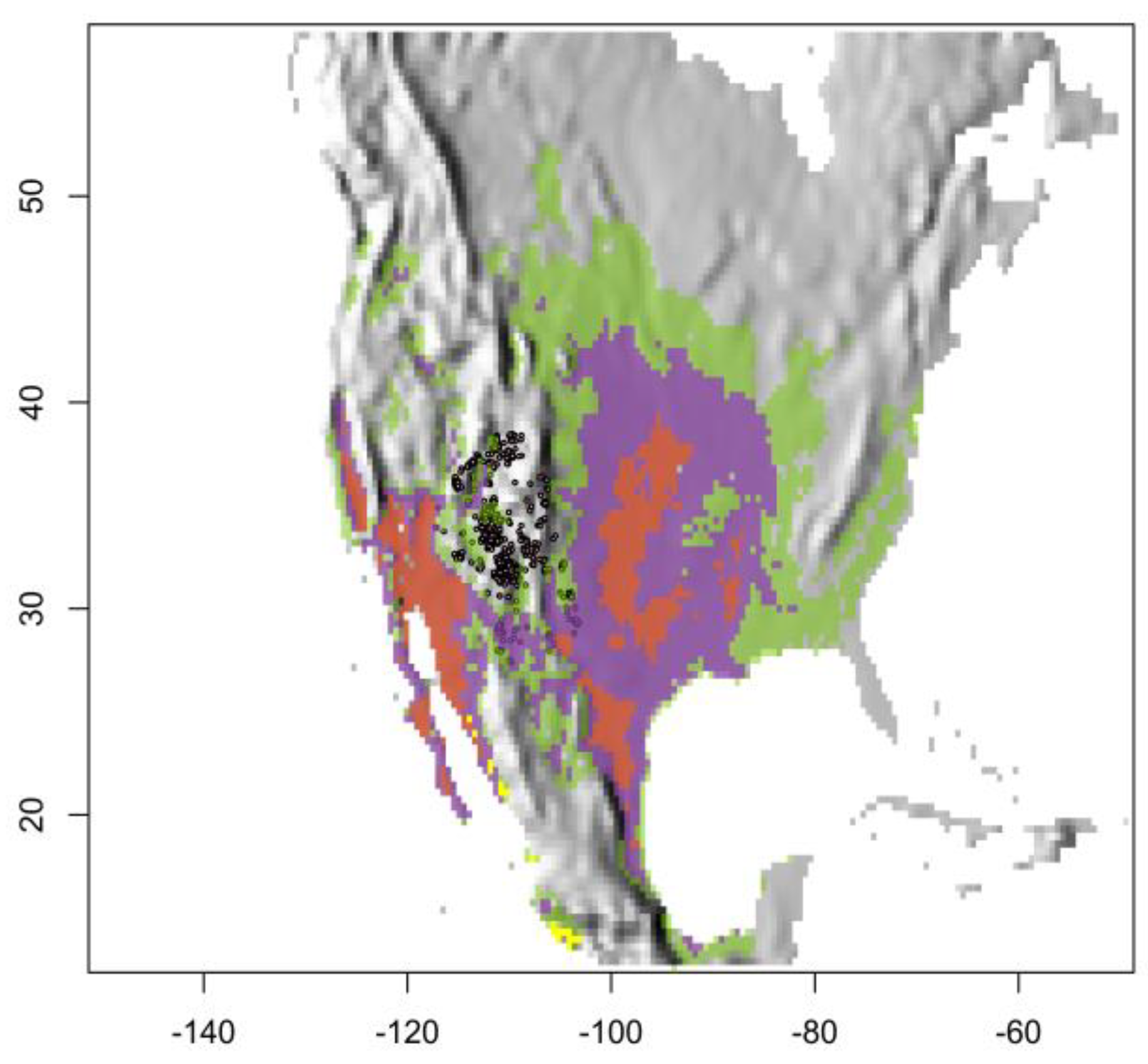
3.3. Relating Predicted Thermally Deleterious Sites with Urosaurus ornatus Locations
4. Discussion

4.1. Predicting Population Vulnerability to Combinations of Temperature and Exposure Time

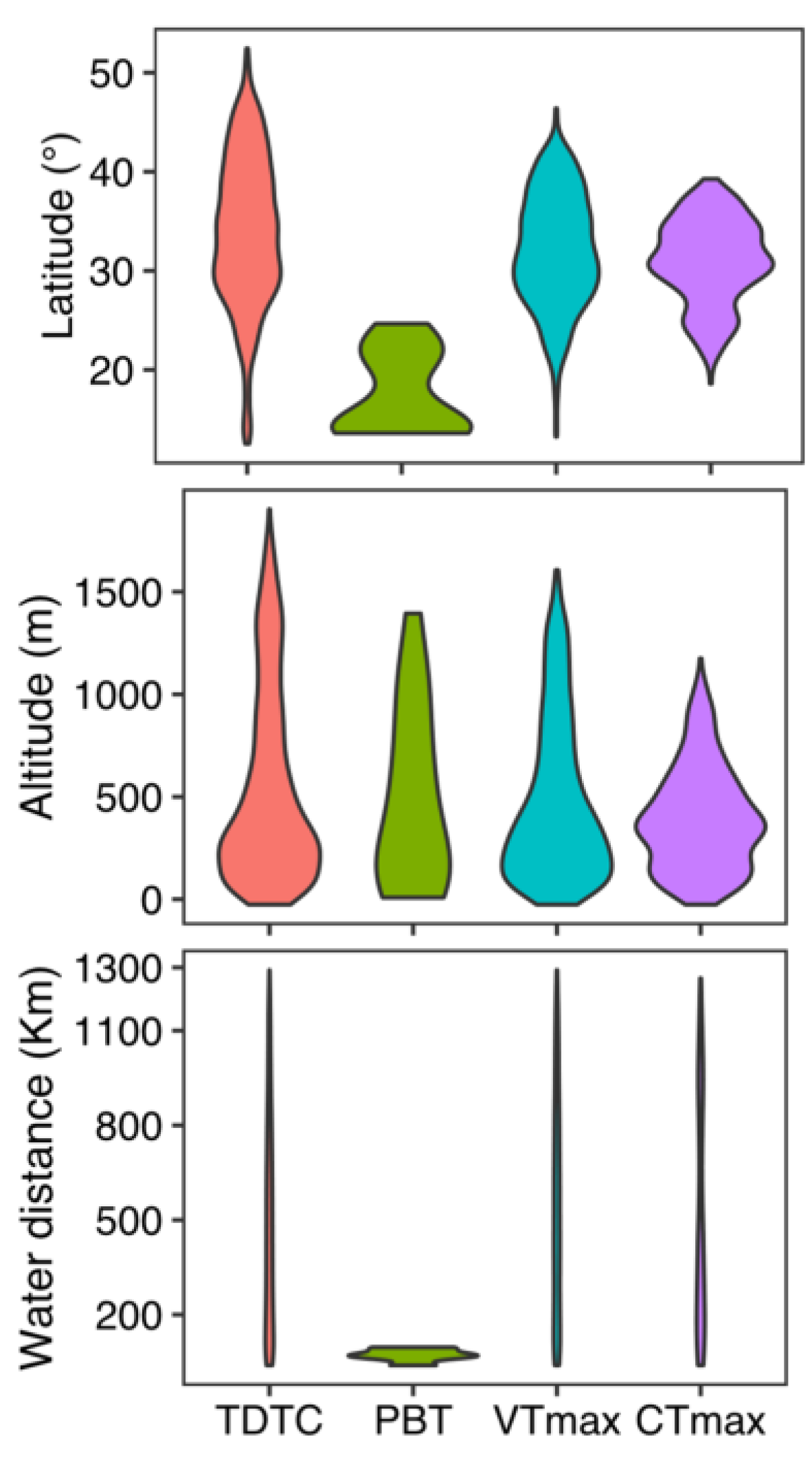
4.2. The Thermal Death Curve and Geographic Biases in Predicted Thermally Deleterious Sites
4.3. Geographic Patterns in the Pulse and Press of Heat Stress
4.4. Relating Predicted Thermally Deleterious Sites with Urosaurus ornatus Locations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Wintle, B.A.; Bekessy, S.A.; Keith, D.A.; Van Wilgen, B.W.; Cabeza, M.; Schröder, B.; Rondinini, C. Ecologi-cal-economic optimization of biodiversity conservation under climate change. Nat. Clim. Chang. 2011, 1, 355–359. [Google Scholar] [CrossRef]
- Williams, S.E.; Shoo, L.P.; Isaac, J.L.; Hoffmann, A.; Langham, G. Towards an Integrated Framework for Assessing the Vulnerability of Species to Climate Change. PLOS Biol. 2008, 6, e325. [Google Scholar] [CrossRef]
- Huey, R.B.; Kearney, M.R.; Krockenberger, A.; Holtum, J.A.; Jess, M.; Williams, S.E. Predicting organismal vul-nerability to climate warming: Roles of behaviour, physiology and adaptation. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 1665–1679. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.M.B.; Beaumont, L.J.; Vance, T.R.; Tozer, C.R.; Remenyi, T.A.; Perkins-Kirkpatrick, S.E.; Mitchell, P.J.; Nicotra, A.B.; McGregor, S.; Andrew, N.R.; et al. Biological responses to the press and pulse of climate trends and extreme events. Nat. Clim. Chang. 2018, 8, 579–587. [Google Scholar] [CrossRef]
- Deutsch, C.A.; Tewksbury, J.J.; Huey, R.B.; Sheldon, K.S.; Ghalambor, C.K.; Haak, D.C.; Martin, P.R. Impacts of climate warming on terrestrial ectotherms across latitude. Proc. Natl. Acad. Sci. USA 2008, 105, 6668–6672. [Google Scholar] [CrossRef]
- Huey, R.B.; Deutsch, C.A.; Tewksbury, J.J.; Vitt, L.J.; Hertz, P.E.; Álvarez Pérez, H.J.; Garland, T., Jr. Why tropical forest lizards are vulnerable to climate warming. Proc. R. Soc. B Biol. Sci. 2009, 276, 1939–1948. [Google Scholar] [CrossRef] [PubMed]
- Kearney, M.R. Activity restriction and the mechanistic basis for extinctions under climate warming. Ecol. Lett. 2013, 16, 1470–1479. [Google Scholar] [CrossRef]
- Khaliq, I.; Hof, C.; Prinzinger, R.; Böhning-Gaese, K.; Pfenninger, M. Global variation in thermal tolerances and vulnerability of endotherms to climate change. Proc. R. Soc. B Boil. Sci. 2014, 281, 20141097. [Google Scholar] [CrossRef] [PubMed]
- Sinervo, B.; Mendez-De-La-Cruz, F.; Miles, D.B.; Heulin, B.; Bastiaans, E.; Villagrán-Santa Cruz, M.; Sites, J.W., Jr. Erosion of lizard diversity by climate change and altered thermal niches. Science 2010, 328, 894–899. [Google Scholar] [CrossRef]
- Kubisch, E.L.; Fernández, J.B.; Ibargüengoytía, N.R. Vulnerability to climate warming of Liolaemus pictus (Squamata, Liolaemidae), a lizard from the cold temperate climate in Patagonia, Argentina. J. Comp. Physiol. B 2016, 186, 243–253. [Google Scholar] [CrossRef]
- Piantoni, C.; Navas, C.A.; Ibargüengoytía, N.R. Vulnerability to climate warming of four genera of New World iguanians based on their thermal ecology. Anim. Conserv. 2016, 19, 391–400. [Google Scholar] [CrossRef]
- Recoder, R.S.; Magalhães-Júnior, A.; Rodrigues, J.; Pinto, H.B.D.A.; Rodrigues, M.T.; Camacho, A. Thermal Constraints Explain the Distribution of the Climate Relict Lizard Colobosauroides carvalhoi (Gymnophthalmidae) in the Semiarid Caatinga. South Am. J. Herpetol. 2018, 13, 248–259. [Google Scholar] [CrossRef]
- Kingsolver, J.G.; Diamond, S.E.; Buckley, L.B. Heat stress and the fitness consequences of climate change for ter-restrial ectotherms. Funct. Ecol. 2013, 27, 1415–1423. [Google Scholar] [CrossRef]
- Pinsky, M.L.; Eikeset, A.M.; McCauley, D.J.; Payne, J.L.; Sunday, J.M. Greater vulnerability to warming of marine versus terrestrial ectotherms. Nature 2019, 569, 108–111. [Google Scholar] [CrossRef]
- Comte, L.; Olden, J.D. Climatic vulnerability of the world’s freshwater and marine fishes. Nat. Clim. Chang. 2017, 7, 718–722. [Google Scholar] [CrossRef]
- Tagliarolo, M.; McQuaid, C.D. Sub-lethal and sub-specific temperature effects are better predictors of mussel distribution than thermal tolerance. Mar. Ecol. Prog. Ser. 2015, 535, 145–159. [Google Scholar] [CrossRef]
- Geiger, R.; Bouyoucos, G.J. The Climate Near the Ground. Am. J. Phys. 1951, 19, 192. [Google Scholar] [CrossRef]
- Sunday, J.M.; Bates, A.E.; Kearney, M.R.; Colwell, R.K.; Dulvy, N.K.; Longino, J.T.; Huey, R.B. Thermal-safety margins and the necessity of thermoregulatory behavior across latitude and elevation. Proc. Natl. Acad. Sci. USA 2014, 111, 5610–5615. [Google Scholar] [CrossRef]
- Rezende, E.L.; Bozinovic, F.; Szilágyi, A.; Santos, M. Predicting temperature mortality and selection in natural Drosophila populations. Science 2020, 369, 1242–1245. [Google Scholar] [CrossRef]
- Rezende, E.L.; Castañeda, L.E.; Santos, M. Tolerance landscapes in thermal ecology. Funct. Ecol. 2014, 28, 799–809. [Google Scholar] [CrossRef]
- Licht, P. The Relation between Preferred Body Temperatures and Testicular Heat Sensitivity in Lizards. Copeia 1965, 1965, 428. [Google Scholar] [CrossRef]
- Cowles, R.B.; Bogert, C.M. A preliminary study of the thermal requirements of desert reptiles. Bull. AMNH 1944, 83, 5. [Google Scholar]
- Camacho, A.; Rusch, T.; Ray, G.; Telemeco, R.S.; Rodrigues, M.T.; Angilletta, M.J. Measuring behavioral thermal tolerance to address hot topics in ecology, evolution, and conservation. J. Therm. Biol. 2018, 73, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Curry-Lindahl, K. Thermal ecology of the tree agama (Agama atricollis) in Zaire with a review of heat tolerance in reptiles. J. Zool. 1979, 188, 185–220. [Google Scholar] [CrossRef]
- Gillooly, J.F.; Brown, J.H.; West, G.B.; Savage, V.M.; Charnov, E.L. Effects of Size and Temperature on Metabolic Rate. Science 2001, 293, 2248–2251. [Google Scholar] [CrossRef] [PubMed]
- Huey, B.R. Temperature, physiology, and the ecology of reptiles. Physiol. Ecol. 1982, 12, 25–95. [Google Scholar]
- Warburg, M.R. The influence of ambient temperature and humidity on the body temperature and water loss from two Australian lizards, Tiliqua rugosa (Gray) (Scincidae) and Amphibolurus barbatus cuvier (Agamidae). Aust. J. Zool. 1965, 13, 331–350. [Google Scholar] [CrossRef]
- Bartholomew, G.A.; Hudson, J.W.; Howell, T.R. Body Temperature, Oxygen Consumption, Evaporative Water Loss, and Heart Rate in the Poor-Will. Condor 1962, 64, 117–125. [Google Scholar] [CrossRef]
- Lowe, C.H., Jr.; Vance, V.J. Acclimation of the critical thermal maximum of the reptile Urosaurus ornatus. Science 1955, 122, 73–74. [Google Scholar] [CrossRef]
- Levy, O.; Buckley, L.B.; Keitt, T.H.; Angilletta, M.J. A Dynamically Downscaled Projection of Past and Future Microclimates. Ecology 2016, in press. [Google Scholar] [CrossRef]
- Arrhenius, S. Quantitative laws in biological chemistry. G. Bell. 1916, 43, 172–173. [Google Scholar]
- Veron, J.; Heatwole, H. Temperature Relations of the Water Skink, Sphenomorphus quoyi. J. Herpetol. 1970, 4, 141–153. [Google Scholar] [CrossRef]
- Wilhoft, D.C.; Anderson, J.D. Effect of Acclimation on the Preferred Body Temperature of the Lizard, Sceloporus occidentalis. Science 1960, 131, 610–611. [Google Scholar] [CrossRef] [PubMed]
- Kour, E.L.; Hutchison, V.H. Critical thermal tolerances and heating and cooling rates of lizards from diverse habitats. Copeia 1970, 1970, 219–229. [Google Scholar] [CrossRef]
- Cowles, R.B.; Burleson, G.L. The Sterilizing Effect of High Temperature on the Male Germ-Plasm of the Yucca Night Lizard, Xantusia vigilis. Am. Nat. 1945, 79, 417–435. [Google Scholar] [CrossRef]
- van Berkum, F.H. Latitudinal patterns of the thermal sensitivity of sprint speed in lizards. Am. Nat. 1988, 132, 327–343. [Google Scholar] [CrossRef]
- Dawson, W.R. On the physiological significance of the preferred body temperatures of reptiles. In Perspectives of Biophysical Ecology; Springer: Berlin/Heidelberg, Germany, 1975; pp. 443–473. [Google Scholar]
- Levy, O.; Buckley, L.B.; Keitt, T.H.; Smith, C.D.; Boateng, K.O.; Kumar, D.S.; Angilletta, M.J., Jr. Resolving the life cycle alters expected impacts of climate change. Proc. R. Soc. B Biol. Sci. 2015, 282, 20150837. [Google Scholar] [CrossRef]
- Levy, O.; Dayan, T.; Porter, W.P.; Kronfeld-Schor, N. Foraging Activity Pattern Is Shaped by Water Loss Rates in a Diurnal Desert Rodent. Am. Nat. 2016, 188, 205–218. [Google Scholar] [CrossRef]
- Bakken, G.S. Measurement and Application of Operative and Standard Operative Temperatures in Ecology. Am. Zool. 1992, 32, 194–216. [Google Scholar] [CrossRef]
- Pierce, D.; Pierce, M.D. Package ‘ncdf4′. 2019. Available online: https://www.vps.fmvz.usp.br/CRAN/web/packages/ncdf4/ncdf4.pdf (accessed on 21 February 2023).
- Fei, L.; Pingfang, H. Energy and Exergy Analysis of a Ground Water Heat Pump System. Phys. Procedia 2012, 24, 169–175. [Google Scholar] [CrossRef]
- Cooper, B.S.; Williams, B.H.; Angilletta, M.J., Jr. Unifying indices of heat tolerance in ectotherms. J. Therm. Biol. 2008, 33, 320–323. [Google Scholar] [CrossRef]
- Jørgensen, L.B.; Malte, H.; Ørsted, M.; Klahn, N.A.; Overgaard, J. A unifying model to estimate thermal tolerance limits in ectotherms across static, dynamic and fluctuating exposures to thermal stress. Sci. Rep. 2021, 11, 12840. [Google Scholar] [CrossRef]
- Willot, Q.; Ørsted, M.; Damsgaard, C.; Overgaard, J. Thermal-death-time model as a tool to analyze heat tolerance, acclimation, and biogeography in ants. Myrmecol. News 2022, 32, 127–138. [Google Scholar]
- Verspagen, N.; Leiva, F.P.; Janssen, I.M.; Verberk, W.C.E.P. Effects of developmental plasticity on heat tolerance may be mediated by changes in cell size in Drosophila melanogaster. Insect Sci. 2019, 27, 1244–1256. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, M.J.; Recoder, R.S.; Camacho, A. A new species of Bachia Gray, 1845 (Squamata: Gymnophthalmidae) from the Eastern Brazilian Cerrado, and data on its ecology, physiology and behavior. Zootaxa 2013, 3616, 173–189. [Google Scholar] [CrossRef]
- Sears, M.W.; Raskin, E.; Angilletta, M.J., Jr. The world is not flat: Defining relevant thermal landscapes in the context of climate change. Integr. Comp. Biol. 2011, 51, 666–675. [Google Scholar] [CrossRef]
- Kearney, M. Hot rocks and much-too-hot rocks: Seasonal patterns of retreat-site selection by a nocturnal ecto-therm. J. Therm. Biol. 2002, 27, 205–218. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Camacho, A.; Rodrigues, M.T.; Jayyusi, R.; Harun, M.; Geraci, M.; Vinagre, C.; Tejedo, M. Does heat tolerance actually predict animals’ geographic thermal limits? bioRxiv 2021. [Google Scholar] [CrossRef]
- Parratt, S.R.; Walsh, B.S.; Metelmann, S.; White, N.; Manser, A.; Bretman, A.J.; Hoffmann, A.A.; Snook, R.R.; Price, T.A.R. Temperatures that sterilize males better match global species distributions than lethal temperatures. Nat. Clim. Chang. 2021, 11, 481–484. [Google Scholar] [CrossRef]
- Morley, S.A.; Peck, L.S.; Sunday, J.M.; Heiser, S.; Bates, A.E. Physiological acclimation and persistence of ectothermic species under extreme heat events. Glob. Ecol. Biogeogr. 2019, 28, 1018–1037. [Google Scholar] [CrossRef]
- Herrando-Pérez, S.; Monasterio, C.; Beukema, W.; Gomes, V.; Ferri-Yáñez, F.; Vieites, D.R.; Araújo, M.B. Heat tolerance is more variable than cold tolerance across species of Iberian lizards after controlling for intraspecific variation. Funct. Ecol. 2020, 34, 631–645. [Google Scholar] [CrossRef]
- Enriquez-Urzelai, U.; Tingley, R.; Kearney, M.R.; Sacco, M.; Palacio, A.S.; Tejedo, M.; Nicieza, A.G. The roles of acclimation and behaviour in buffering climate change impacts along elevational gradients. J. Anim. Ecol. 2020, 89, 1722–1734. [Google Scholar] [CrossRef]
- Strahler, A.N.; Strahler, A.H. Geografía física. Estud. Geogr. 1991, 52, 187. [Google Scholar]
- Wiens, J.J.; Camacho, A.; Goldberg, A.; Jezkova, T.; Kaplan, M.E.; Lambert, S.M.; Miller, E.C.; Streicher, J.W.; Walls, R.L. Climate change, extinction, and Sky Island biogeography in a montane lizard. Mol. Ecol. 2019, 28, 2610–2624. [Google Scholar] [CrossRef]
- Thomas, C.D. Translocation of species, climate change, and the end of trying to recreate past ecological communities. Trends Ecol. Evol. 2011, 26, 216–221. [Google Scholar] [CrossRef]
- Camacho, A.; Rodrigues, M.T.; Navas, C. Extreme operative temperatures are better descriptors of the thermal environment than mean temperatures. J. Therm. Biol. 2015, 49, 106–111. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value | St.Err. | t-Value | p-Value | Geographic Bias |
|---|---|---|---|---|---|
| (Intercept) | 34.059 | 0.104 | 327.383 | 0 | Latitudinal |
| PBT | −16.497 | 1.592 | −10.335 | 0 | Latitudinal |
| VTmax | −2.004 | 0.173 | −11.545 | 0 | Latitudinal |
| CTmax | −3.164 | 0.285 | −11.09 | 0 | Latitudinal |
| (Intercept) | 550.200 | 6.583 | 83.583 | 0 | Altitudinal |
| PBT | −68.824 | 100.995 | −0.681 | 0.495 | Altitudinal |
| VTmax | −67.636 | 10.981 | −6.158 | 0 | Altitudinal |
| CTmax | −154.647 | 18.052 | −8.566 | 0 | Altitudinal |
| (Intercept) | 480,059.4 | 5014.67 | 95.73100 | 0 | Water distance |
| PBT | −407,682.8 | 76,937.02 | −5.29892 | 0 | Water distance |
| VTmax | 60,374.2 | 8365.90 | 7.21670 | 0 | Water distance |
| CTmax | 23,370.1 | 13,752.33 | 1.69936 | 0.089 | Water distance |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camacho, A.; Angilletta, M.J., Jr.; Levy, O. A Theoretical Thermal Tolerance Function for Ectothermic Animals and Its Implications for Identifying Thermal Vulnerability across Large Geographic Scales. Diversity 2023, 15, 680. https://doi.org/10.3390/d15050680
Camacho A, Angilletta MJ Jr., Levy O. A Theoretical Thermal Tolerance Function for Ectothermic Animals and Its Implications for Identifying Thermal Vulnerability across Large Geographic Scales. Diversity. 2023; 15(5):680. https://doi.org/10.3390/d15050680
Chicago/Turabian StyleCamacho, Agustín, Michael J. Angilletta, Jr., and Ofir Levy. 2023. "A Theoretical Thermal Tolerance Function for Ectothermic Animals and Its Implications for Identifying Thermal Vulnerability across Large Geographic Scales" Diversity 15, no. 5: 680. https://doi.org/10.3390/d15050680
APA StyleCamacho, A., Angilletta, M. J., Jr., & Levy, O. (2023). A Theoretical Thermal Tolerance Function for Ectothermic Animals and Its Implications for Identifying Thermal Vulnerability across Large Geographic Scales. Diversity, 15(5), 680. https://doi.org/10.3390/d15050680