
Limobrevibacterium gyesilva gen. nov., sp. nov., Isolated from Forest Soil


Abstract
:1. Introduction
2. Material and Methods
2.1. Isolation and Ecology
2.2. Physiology and Chemotaxonomy
2.3. 16S rRNA Phylogeny
2.4. Genome Features
3. Results and Discussion
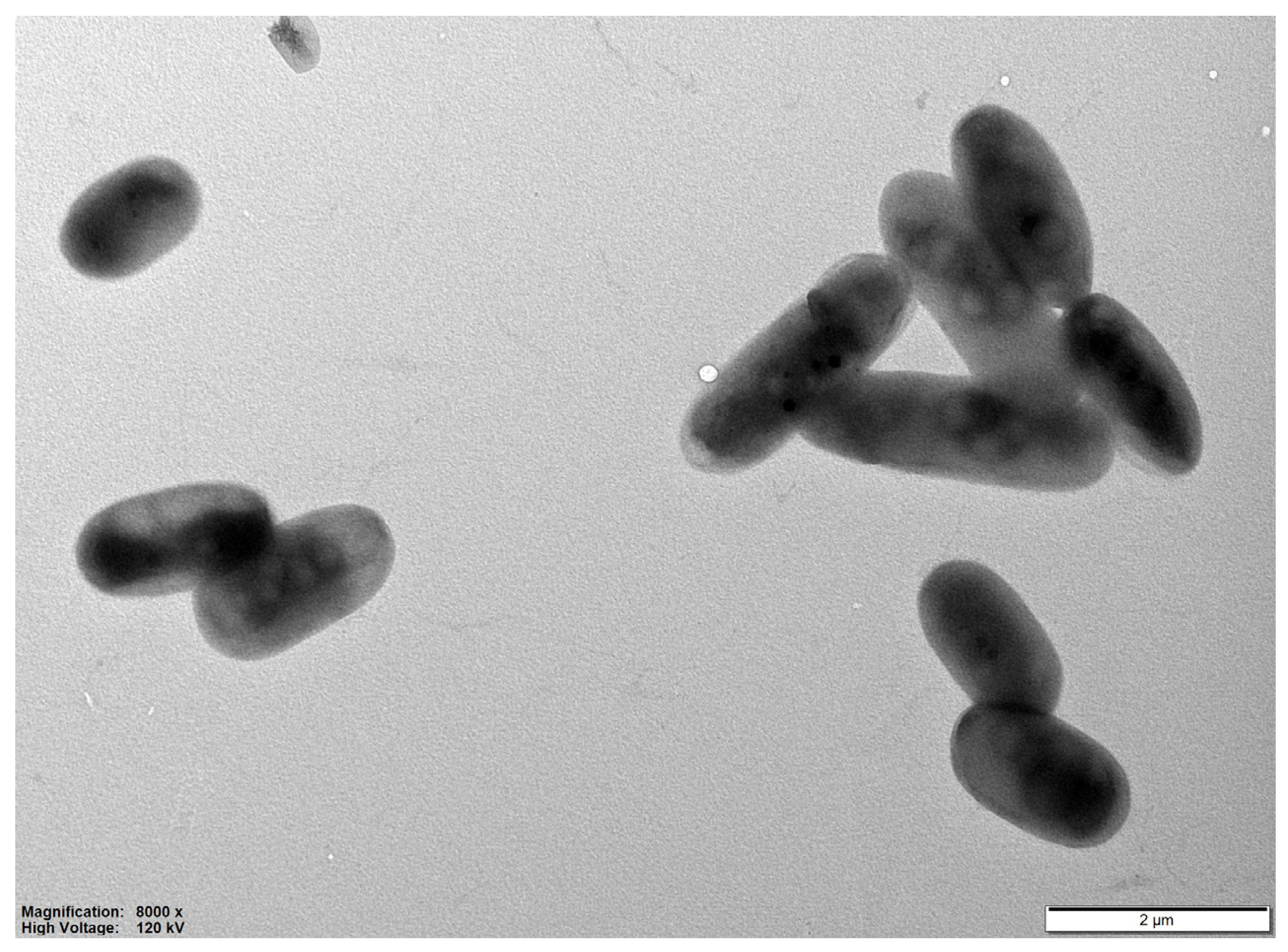
3.1. Isolation of Uncultured Strain
3.2. Physiology and Chemotaxonomy
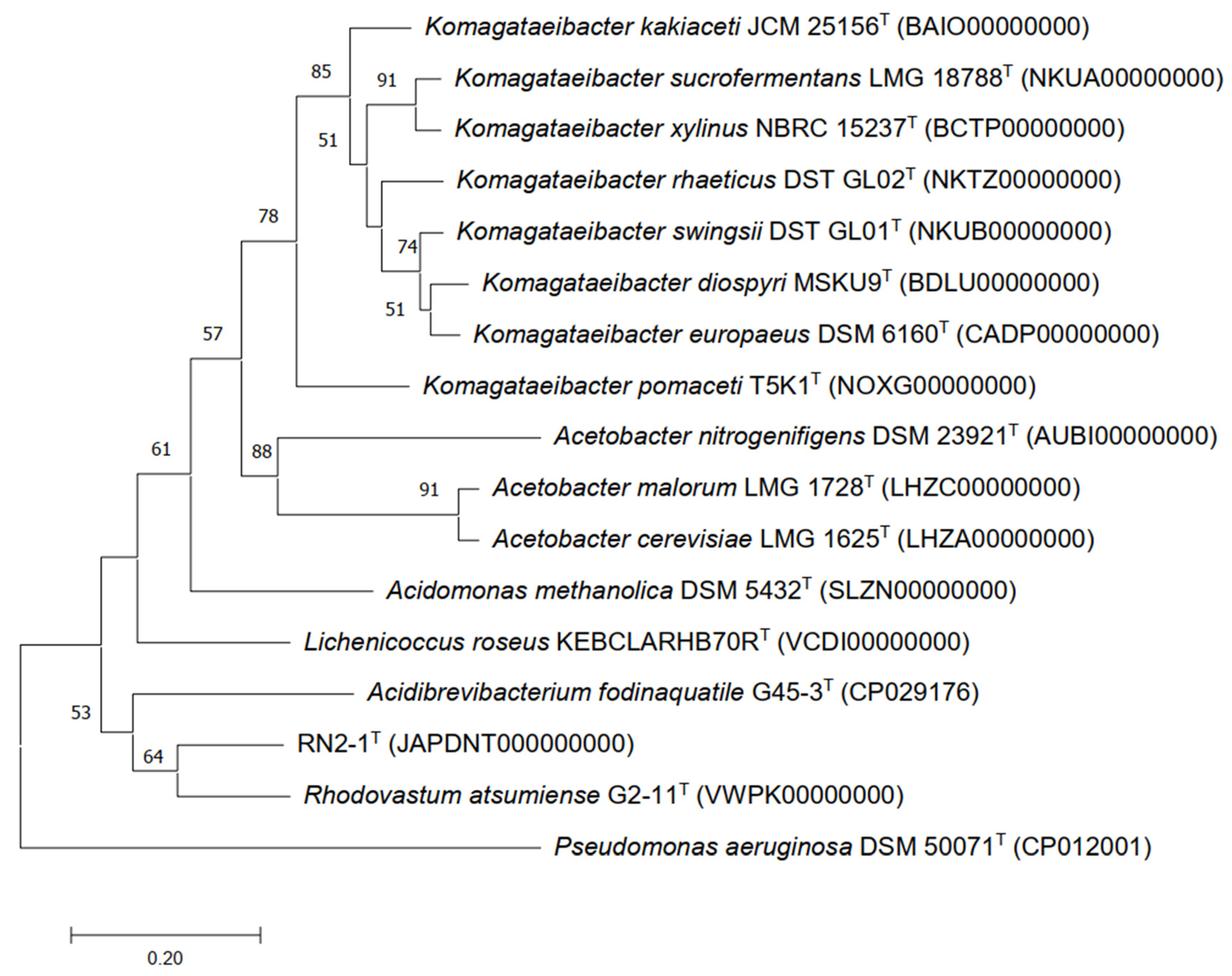
3.3. 16S rRNA Phylogeny
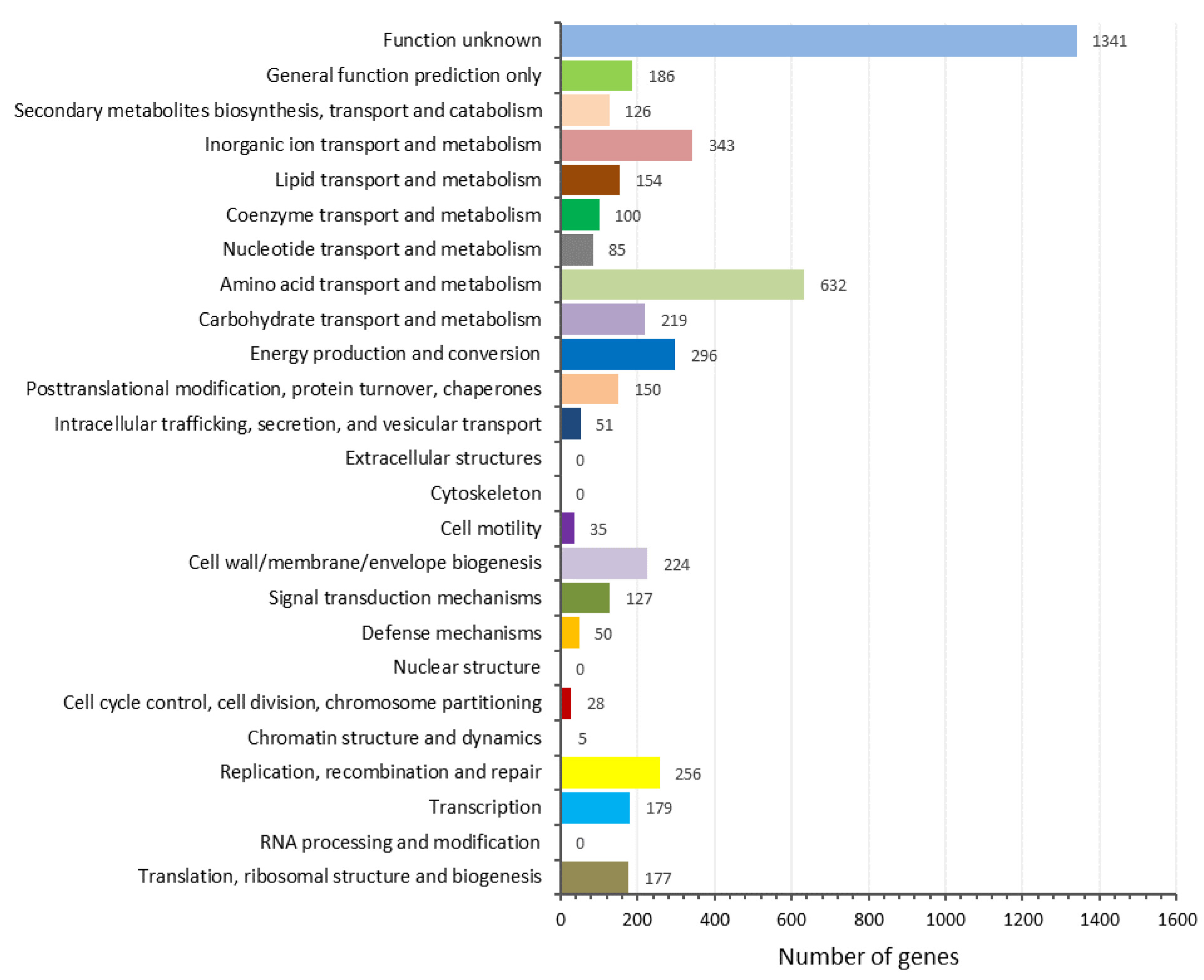
3.4. Genome Features
4. Conclusions
- Description of Limobrevibacterium gen. nov.
- Description of Limobrevibacterium gyesilva sp. nov.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gillis, M.; de Ley, J. Intra- and intergeneric similarities of the ribosomal ribonucleic acid cistrons of Acetobacter and Gluconobacter. Int. J. Syst. Bacteriol. 1980, 30, 7–27. [Google Scholar] [CrossRef]
- Parte, A.C. LPSN—List of prokaryotic names withstanding in nomenclature (bacterio.net), 20 years on. Int. J. Syst. Evol. Microbiol. 2018, 68, 1825–1829. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, M.; de Castro, J.V., Jr.; Müller, H.; Berg, G.; Grube, M. In situ analysis of the bacterial community associated with the reindeer lichen Cladonia arbuscula reveals predominance of Alphaproteobacteria. FEMS Microbiol. Ecol. 2008, 66, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Pankratov, T.A.; Grouzdev, D.S.; Patutina, E.O.; Kolganova, T.V.; Berestovskaya, J.J.; Ashikhmin, A.A. Lichenicoccus roseus gen. nov., sp. nov., the first bacteriochlorophyll a-containing, psychrophilic and acidophilic Acetobacteraceae bacteriobiont of lichen Cladonia species. Int. J. Syst. Evol. Microbiol. 2020, 70, 4591–4601. [Google Scholar] [CrossRef] [PubMed]
- Okamura, K.; Hisada, T.; Kanbe, T.; Hiraishi, A. Rhodovastum atsumiense gen. nov., sp. nov., a phototrophic alphaproteobacterium isolated from paddy soil. J. Gen. Appl. Microbiol. 2009, 55, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Hiraishi, A.; Matsuzawa, Y.; Kanbe, T.; Wakao, N. Acidisphaera rubrifaciens gen. nov., sp. nov., an aerobic bacteriochlorophyll-containing bacterium isolated from acidic environments. Int. J. Syst. Evol. Microbiol. 2000, 50, 1539–1546. [Google Scholar] [CrossRef]
- Muhadesi, J.B.; Huang, Y.; Wang, B.J.; Jiang, C.Y.; Liu, S.J. Acidibrevibacterium fodinaquatile gen. nov., sp. nov., isolated from acidic mine drainage. Int. J. Syst. Evol. Microbiol. 2019, 69, 3243–3250. [Google Scholar] [CrossRef]
- Belova, S.E.; Pankratov, T.A.; Detkova, E.N.; Kaparullina, E.N.; Dedysh, S.N. Acidisoma tundrae gen. nov., sp. nov. and Acidisoma sibiricum sp. nov., two acidophilic, psychrotolerant members of the Alphaproteobacteria from acidic northern wetlands. Int. J. Syst. Evol. Microbiol. 2009, 59, 2283–2290. [Google Scholar] [CrossRef]
- Muthukumarasamy, R.; Revathi, G.; Seshadri, S.; Lakshminarasimhan, C. Gluconacetobacter diazotrophicus (syn. Acetobacter diazotrophicus), a promising diazotrophic endophyte in tropics. Curr. Sci. 2002, 83, 137–145. [Google Scholar]
- Yamada, Y.; Katsura, K.; Kawasaki, H.; Widyastuti, Y.; Saono, S.; Seki, T.; Uchimura, T.; Komagata, K. Asaia bogorensis gen. nov., sp. nov., an unusual acetic acid bacterium in the alpha-Proteobacteria. Int. J. Syst. Evol. Microbiol. 2000, 50, 823–829. [Google Scholar] [CrossRef]
- Yukphan, P.; Malimas, T.; Muramatsu, Y.; Takahashi, M.; Kaneyasu, M.; Potacharoen, W.; Tanasupawat, S.; Nakagawa, Y.; Hamana, K.; Tahara, Y.; et al. Ameyamaea chiangmaiensis gen. nov., sp. nov., an acetic acid bacterium in the alpha-Proteobacteria. Biosci. Biotechnol. Biochem. 2009, 73, 2156–2162. [Google Scholar] [CrossRef] [PubMed]
- Yukphan, P.; Malimas, T.; Muramatsu, Y.; Potacharoen, W.; Tanasupawat, S.; Nakagawa, Y.; Tanticharoen, M.; Yamada, Y. Neokomagataea gen. nov., with descriptions of Neokomagataea thailandica sp. nov. and Neokomagataea tanensis sp. nov., osmotolerant acetic acid bacteria of the alpha-Proteobacteria. Biosci. Biotechnol. Biochem. 2011, 75, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.W.; Salam, N.; Hua, Z.S.; Liu, B.B.; Han, M.X.; Fang, B.Z.; Wang, D.; Xiao, M.; Hozzein, W.N.; Li, W.J. Siccirubricoccus deserti gen. nov., sp. nov., a proteobacterium isolated from a desert sample. Int. J. Syst. Evol. Microbiol. 2017, 67, 4862–4867. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.L.; Li, S.; Wei, Q.C.; Lian, W.H.; Lu, C.Y.; Xu, L.; Huang, L.N.; Dong, L.; Li, W.J. Sabulicella rubraurantiaca gen. nov., sp. nov., a new member of the family Acetobacteraceae, isolated from desert soil. Arch. Microbiol. 2021, 204, 1. [Google Scholar] [CrossRef] [PubMed]
- Ming, H.; Duan, Y.Y.; Yin, Y.R.; Meng, X.L.; Li, S.; Zhou, E.M.; Huang, J.R.; Nie, G.X.; Li, W.J. Crenalkalicoccus roseus gen. nov., sp. nov., a thermophilic bacterium isolated from alkaline hot springs. Int. J. Syst. Evol. Microbiol. 2016, 66, 2319–2326. [Google Scholar] [CrossRef]
- Albuquerque, L.; Rainey, F.A.; Nobre, M.F.; da Costa, M.S. Elioraea tepidiphila gen. nov., sp. nov., a slightly thermophilic member of the Alphaproteobacteria. Int. J. Syst. Evol. Microbiol. 2008, 58, 773–778. [Google Scholar] [CrossRef]
- Khan, S.A.; Jeong, S.E.; Jung, H.S.; Quan, Z.X.; Jeon, C.O. Roseicella frigidaeris gen. nov., sp. nov., isolated from an air-conditioning system. Int. J. Syst. Evol. Microbiol. 2019, 69, 1384–1389. [Google Scholar] [CrossRef]
- Kim, W.H.; Kim, D.H.; Kang, K.; Ahn, T.Y. Dankookia rubra gen. nov., sp. nov., an alphaproteobacterium isolated from sediment of a shallow stream. J. Microbiol. 2016, 54, 420–425. [Google Scholar] [CrossRef]
- Lányi, B. Classical and Rapid Identification Method for Medically Important Bacteria. In Method in Microbiology 19; Cowell, R., Ed.; Academic Press: Oak Ridge, TN, USA, 1987; pp. 1–65. [Google Scholar]
- Trinh, N.H.; Kim, J. Paraburkholderia flava sp. nov., isolated from cool temperate forest soil. Int. J. Syst. Evol. Microbiol. 2020, 70, 2509–2514. [Google Scholar] [CrossRef]
- Okamura, K.; Hisada, T.; Hiraishi, A. Characterization of thermotolerant purple nonsulfur bacteria isolated from hot-spring Chloroflexus mats and the reclassification of “Rhodopseudomonas cryptolactis” Stadtwald-Demchick et al. 1990 as Rhodoplanes cryptolactis nom. rev., comb. nov. J. Gen. Appl. Microbiol. 2007, 53, 357–361. [Google Scholar] [CrossRef]
- Le Olson, T.; Meene, A.M.; Francis, J.N.; Pierson, B.K.; Blankenship, R.E. Pigment analysis of “Candidatus Chlorothrix halophila”, a green filamentous anoxygenic phototrophic bacterium. J. Bacteriol. 2007, 189, 4187–4195. [Google Scholar] [CrossRef] [PubMed]
- Tindall, B.J.; Sikorski, J.; Smibert, R.A.; Krieg, N.R. Phenotypic Characterization and the Principles of Comparative Systematics. In Methods for General and Molecular Microbiology; Reddy, C.A., Ed.; ASM Press: Washington, DC, USA, 2007; pp. 330–393. [Google Scholar]
- Smibert, R.M.; Krieg, N.R. Phenotypic characterization. In Methods for General and Molecular Bacteriology; Gerhardt, P., Murray, R.G.E., Wood, W.A., Krieg, N.R., Eds.; American Society for Microbiology: Washington, DC, USA, 1994; pp. 607–654. [Google Scholar]
- Minnikin, D.E.; O’Donnell, A.G.; Goodfellow, M.; Alderson, G.; Athalye, M.; Schaal, A.; Parlett, J. An integrated procedure for the extraction of bacterial isoprenoid quinones and polar lipids. J. Microbiol. Methods 1984, 2, 233–241. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Frank, J.A.; Reich, C.I.; Sharma, S.; Weisbaum, J.S.; Wilson, B.A.; Olsen, G.J. Critical evaluation of two primers commonly used for amplification of bacterial 16S rRNA genes. Appl. Environ. Microbiol. 2008, 74, 2461–2470. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically United database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Soc. Study Evol. 1985, 39, 783–791. [Google Scholar]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Na, S.I.; Kim, Y.O.; Yoon, S.H.; Ha, S.M.; Baek, I.; Chun, J. UBCG: Up-to-date bacterial core gene set and pipeline for phylogenomic tree reconstruction. J. Microbiol. 2018, 56, 280–285. [Google Scholar] [CrossRef]
- Yoon, S.H.; Ha, S.M.; Lim, J.; Kwon, S.; Chun, J. A large-scale evaluation of algorithms to calculate average nucleotide identity. Antonie Van Leeuwenhoek 2017, 110, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef]
- Sasser, M. Identification of bacteria by gas chromatography of cellular fatty acids. Technol. Note 2001, 101. [Google Scholar]
- Chun, J.; Oren, A.; Ventosa, A.; Christensen, H.; Arahal, D.R.; da Costa, M.S.; Rooney, A.P.; Yi, H.; Xu, X.-W.; De Meyer, S.; et al. Proposed minimal standards for the use of genome data for the taxonomy of prokaryotes. Int. J. Syst. Evol. Microbiol. 2018, 68, 461–466. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| Source of isolation | Forest soil | Paddy soil | Sediment from an acidic hot spring and pyritic mine drainage | The thalli of Cladonia arbuscula and Cladonia stellaris lichens | Acidic mine drainage | Acidic northern wetlands |
| Cell shape | Ovals, Rods | Ovals, rods | Cocci, short rods | Spherical to ovoid | Rods | Coccobacilli |
| Cell size (µm) | 0.8–1.1 × 1.5–3.2 | 2.0–3.0 × 3.0–10 | 0.7–0.9 × 0.9–1.6 | 0.8–1.6 × 0.9–2.26 | 1.3–1.5 × 3.2–3.3 | 0.7–1.5 × 1.4–4.1 |
| Colony color | White, Mucoid | Red brown | Salmon pink | pink or salmon pink | Light yellow | White, cream |
| Motility | − | + | − | − | − | − |
| Anaerobic phototrophy | − | + | − | − | − | − |
| Bacteriochlorophyll a | − | + | + | + | − | − |
| Temperature range for growth (°C) | 20–40 | 20–40 | 20–40 | 5–30 | 20–45 | 2–30 |
| Optimal growth temperature (°C) | 30–35 | 30–35 | 20–35 | 10–15 | 37 | 18–22 |
| pH range for growth | 6–9 | 5.0–8.5 | 3.5–6.0 | 3.0–7.5 | 2.5–5 | 3.7–7.5 |
| pH optimum for growth | 7–8 | 6.0–6.5 | 4.5–5.0 | 4.5–5.5 | 4.0 | 5.0–5.7 |
| Tolerance of >2.5 (%w/v) NaCl | − | − | + | + | − | − |
| Electron donor/carbon source | ||||||
| Acetate | − | ++ | − | − | nd | − |
| Lactate | − | + | + | − | nd | + |
| Mannose | +/− | + | − | + | − | + |
| Sorbitol | +/− | + | + | + | + | + |
| Glycerol | +/− | − | + | − | − | + |
| Asparagine | + | + | − | − | + | + |
| Aspartate | − | + | − | nd | + | − |
| Glutamate | − | ++ | − | − | nd | nd |
| Glutamine | ++ | ++ | − | − | nd | nd |
| Citrate | ++ | − | +/− | − | nd | − |
| Nitrogen fixation | − | + | nd | − | nd | nd |
| Major quinone(s) | Q-10 | Q-10, RQ-10 | Q-10 | Q-10 | Q-10 | Q-10 |
| DNA G+C content (mol%) | 68.7 | 67.8 | 69.1–69.8 | 69.1–69.8 | 65.9 | 60.5–61.9 |
| Fatty Acid | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| Saturated | ||||||
| C14:0 | 1.26 | 2.30 | 1.14 | - | - | - |
| C16:0 | 6.54 | 20.47 | 20.07 | 7.82 | 22.22 | 10.33 |
| C18:0 | 1.02 | 2.17 | 1.31 | 2.71 | - | 4.73 |
| 10-methyl C17:0 | - | 0.68 | - | - | - | - |
| Cyclo-C17:0 | 5.62 | - | - | - | 1.06 | - |
| Unsaturated | ||||||
| C16:1 ω5c | 1.25 | 2.43 | - | - | - | - |
| C18:1 ω5c | - | 0.74 | - | 0.62 | - | - |
| C18:1 ω9c | - | - | - | - | - | 3.33 |
| C20:1 ω7c | - | 0.58 | - | - | - | |
| cyclo C19:0 ω8c | 7.85 | 0.57 | 20.80 | 1.25 | 38.51 | |
| Hydroxy | ||||||
| C16:0 3OH | - | - | 0.68 | 1.27 | - | - |
| C18:0 3OH | 0.68 | 1.56 | 0.81 | 1.64 | - | 9.72 |
| C18:1 2OH | 6.98 | 0.81 | 10.98 | 16.60 | - | 3.62 |
| Summed features * | ||||||
| 2 | - | 3.51 | - | 2.33 | 2.00 | 1.26 |
| 3 | 7.65 | 9.69 | 0.58 | 0.51 | 0.72 | - |
| 8 | 61.14 | 51.84 | 43.64 | 62.50 | 2.97 | 28.50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tuyet, N.L.T.; Kim, J. Limobrevibacterium gyesilva gen. nov., sp. nov., Isolated from Forest Soil. Diversity 2023, 15, 684. https://doi.org/10.3390/d15050684
Tuyet NLT, Kim J. Limobrevibacterium gyesilva gen. nov., sp. nov., Isolated from Forest Soil. Diversity. 2023; 15(5):684. https://doi.org/10.3390/d15050684
Chicago/Turabian StyleTuyet, Nhan Le Thi, and Jaisoo Kim. 2023. "Limobrevibacterium gyesilva gen. nov., sp. nov., Isolated from Forest Soil" Diversity 15, no. 5: 684. https://doi.org/10.3390/d15050684
APA StyleTuyet, N. L. T., & Kim, J. (2023). Limobrevibacterium gyesilva gen. nov., sp. nov., Isolated from Forest Soil. Diversity, 15(5), 684. https://doi.org/10.3390/d15050684