
Expanding Actinomycetota Diversity in the TBRC Culture Collection through Metabarcoding and Simulated In Situ Cultivation of Thailand’s Mekong River Microbiota


Abstract
:1. Introduction
2. Materials and Methods
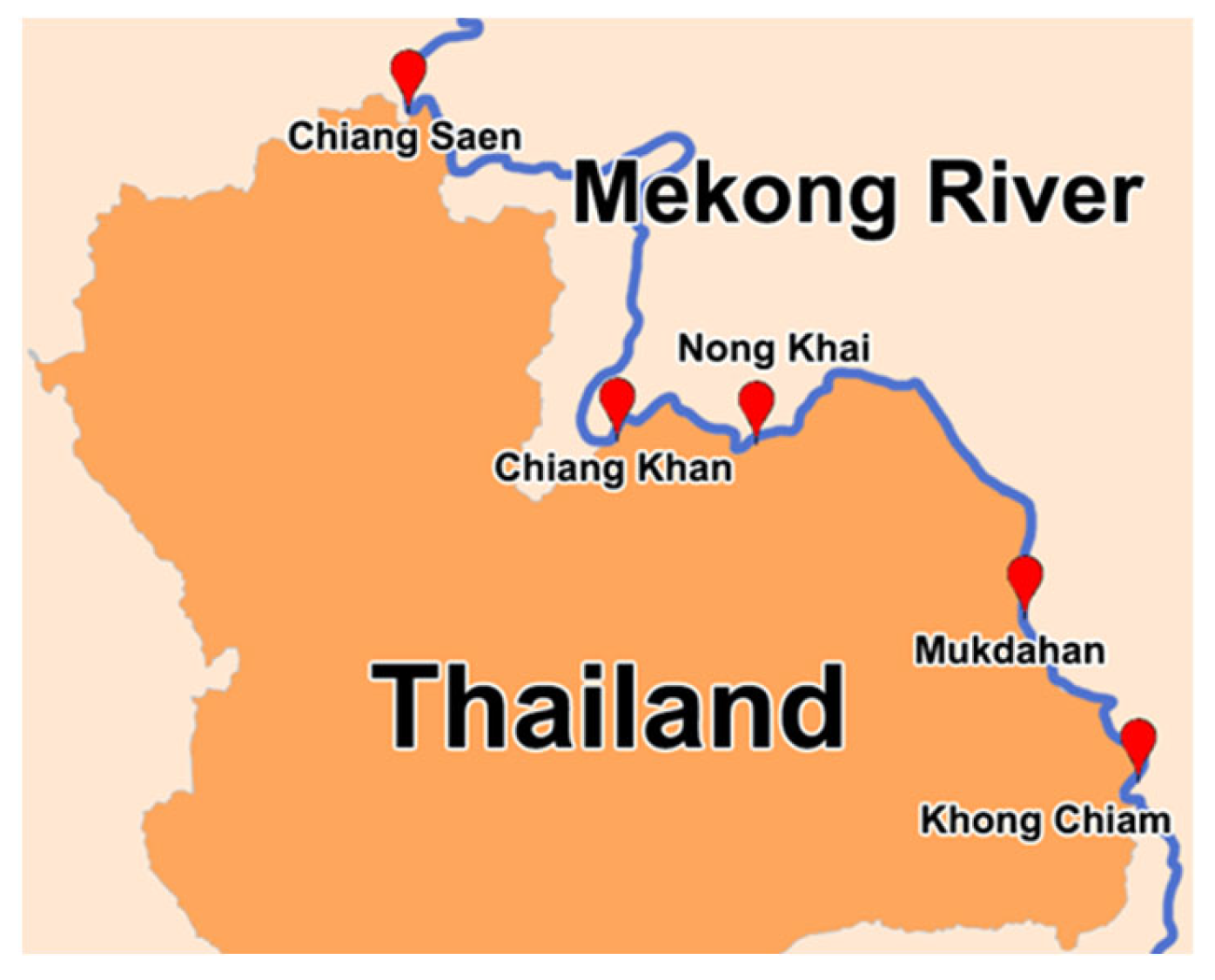
2.1. Sample Collection
2.2. DNA Extraction and 16S-Amplicon Sequencing
2.3. Isolation and Cultivation
2.3.1. Design and Development of the IC Plate
2.3.2. Isolation of Actinomycetota
2.3.3. Identification and Dereplication of Actinomycetota Isolates
2.4. Analysis of Microbial Community Diversity
3. Results
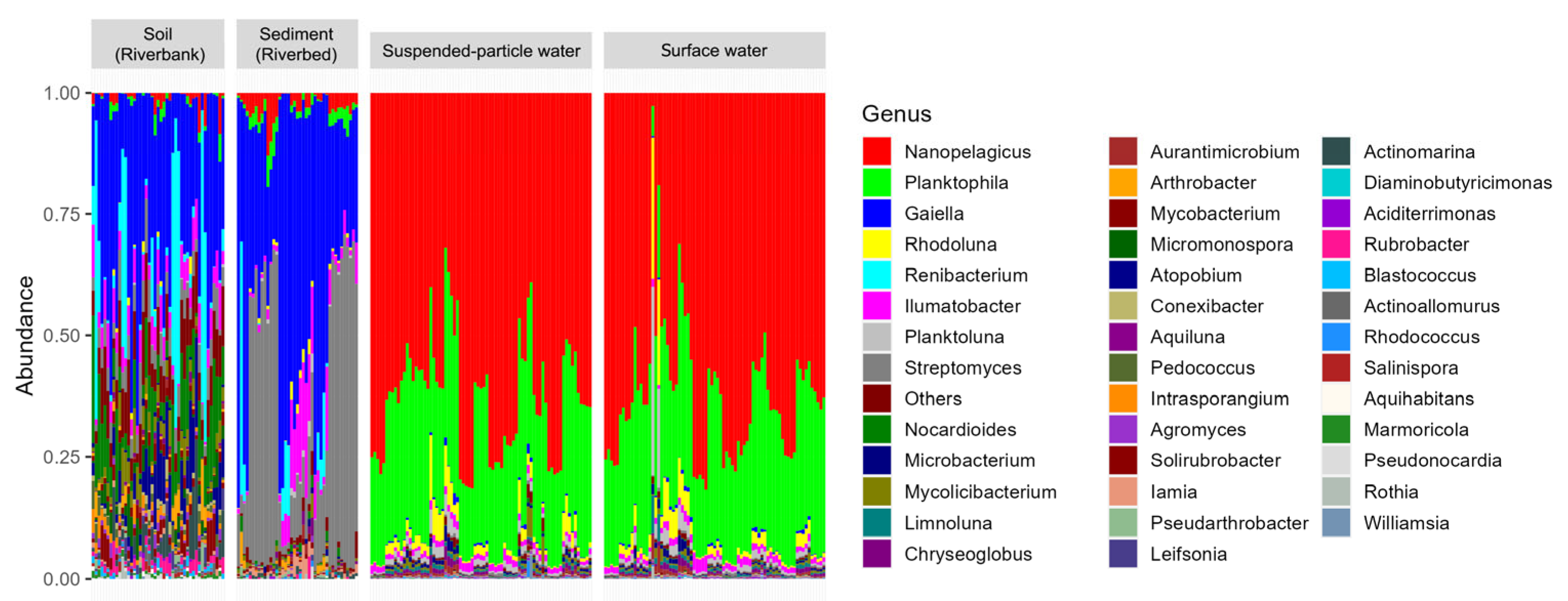
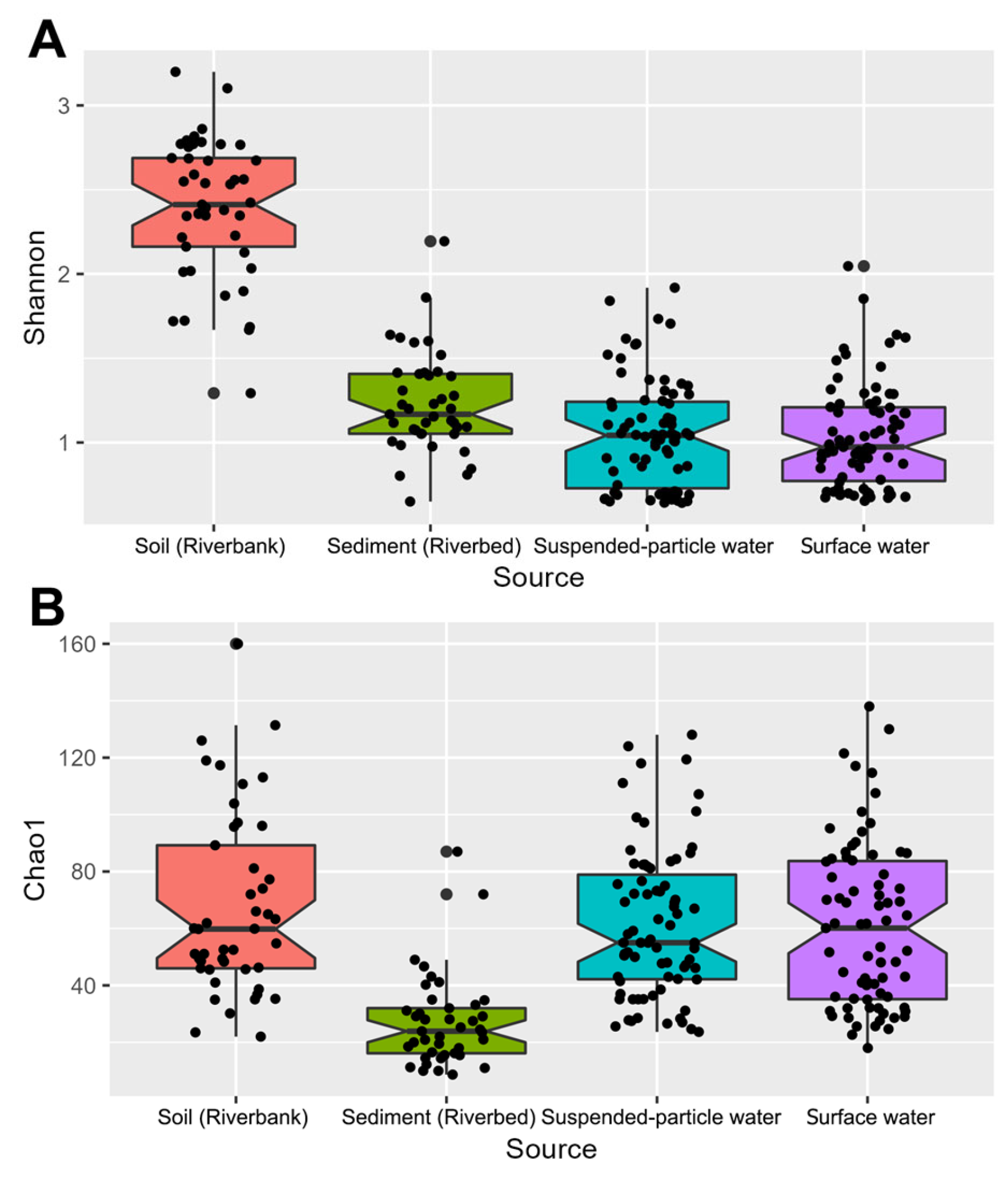
3.1. Culture-Independent, Metabarcoding-Based Diversity, and Composition of Actinomycetota in the Mekong River
3.2. Culture-Dependent Isolation of Actinomycetota in the Mekong River
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sibanda, T.; Mabinya, L.V.; Mazomba, N.; Akinpelu, D.A.; Bernard, K.; Olaniran, A.O.; Okoh, A.I. Antibiotic Producing Potentials of Three Freshwater Actinomycetes Isolated from the Eastern Cape Province of South Africa. Int. J. Mol. Sci. 2010, 11, 2612–2623. [Google Scholar] [CrossRef]
- Allali, K.; Goudjal, Y.; Zamoum, M.; Bouznada, K.; Sabaou, N.; Zitouni, A. Nocardiopsis dassonvillei strain MB22 from the Algerian Sahara promotes wheat seedlings growth and potentially controls the common root rot pathogen Bipolaris sorokiniana. J. Plant Pathol. 2019, 101, 1115–1125. [Google Scholar] [CrossRef]
- Chaiharn, M.; Chunhaleuchanon, S.; Lumyong, S. Screening siderophore producing bacteria as potential biological control agent for fungal rice pathogens in Thailand. World J. Microbiol. Biotechnol. 2009, 25, 1919–1928. [Google Scholar] [CrossRef]
- El-Tarabily, K.A.; Soliman, M.H.; Nassar, A.H.; Al-Hassani, H.A.; Sivasithamparam, K.; McKenna, F.; Hardy, G.E.S.J. Biological control of Sclerotinia minor using a chitinolytic bacterium and actinomycetes. Plant Pathol. 2000, 49, 573–583. [Google Scholar] [CrossRef]
- Errakhi, R.; Bouteau, F.; Lebrihi, A.; Barakate, M. Evidences of biological control capacities of Streptomyces spp. against Sclerotium rolfsii responsible for damping-off disease in sugar beet (Beta vulgaris L.). World J. Microbiol. Biotechnol. 2007, 23, 1503–1509. [Google Scholar] [CrossRef]
- Li, X.; Jing, T.; Zhou, D.; Zhang, M.; Qi, D.; Zang, X.; Zhao, Y.; Li, K.; Tang, W.; Chen, Y.; et al. Biocontrol efficacy and possible mechanism of Streptomyces sp. H4 against postharvest anthracnose caused by Colletotrichum fragariae on strawberry fruit. Postharvest Biol. Technol. 2021, 175, 111401. [Google Scholar] [CrossRef]
- Chukwuneme, C.F.; Babalola, O.O.; Kutu, F.R.; Ojuederie, O.B. Characterization of actinomycetes isolates for plant growth promoting traits and their effects on drought tolerance in maize. J. Plant Interact. 2020, 15, 93–105. [Google Scholar] [CrossRef] [Green Version]
- Franco-Correa, M.; Quintana, A.; Duque, C.; Suarez, C.; Rodríguez, M.X.; Barea, J.-M. Evaluation of actinomycete strains for key traits related with plant growth promotion and mycorrhiza helping activities. Appl. Soil Ecol. 2010, 45, 209–217. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Srinivas, V.; Vidya, M.S.; Rathore, A. Plant growth-promoting activities of Streptomyces spp. in sorghum and rice. SpringerPlus 2013, 2, 574. [Google Scholar] [CrossRef] [Green Version]
- Goudjal, Y.; Toumatia, O.; Sabaou, N.; Barakate, M.; Mathieu, F.; Zitouni, A. Endophytic actinomycetes from spontaneous plants of Algerian Sahara: Indole-3-acetic acid production and tomato plants growth promoting activity. World J. Microbiol. Biotechnol. 2013, 29, 1821–1829. [Google Scholar] [CrossRef] [Green Version]
- Qin, S.; Miao, Q.; Feng, W.-W.; Wang, Y.; Zhu, X.; Xing, K.; Jiang, J.-H. Biodiversity and plant growth promoting traits of culturable endophytic actinobacteria associated with Jatropha curcas L. growing in Panxi dry-hot valley soil. Appl. Soil Ecol. 2015, 93, 47–55. [Google Scholar] [CrossRef]
- Álvarez, A.; Yañez, M.L.; Benimeli, C.S.; Amoroso, M.J. Maize plants (Zea mays) root exudates enhance lindane removal by native Streptomyces strains. Int. Biodeterior. Biodegrad. 2012, 66, 14–18. [Google Scholar] [CrossRef]
- Arya, R.; Mishra, N.K.; Sharma, A.K. Brevibacillus borstelensis and Streptomyces albogriseolus have roles to play in degradation of herbicide, sulfosulfuron. 3 Biotech 2016, 6, 246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Q.S.; Chen, S.H.; Hu, M.Y.; Haq, M.R.U.; Yang, L.; Li, H. Biodegradation of Cypermethrin by a newly isolated actinomycetes HU-S-01 from wastewater sludge. Int. J. Environ. Sci. Technol. 2011, 8, 45–56. [Google Scholar] [CrossRef] [Green Version]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Goodfellow, M.; Williams, S.T. Ecology of Actinomycetes. Annu. Rev. Microbiol. 1983, 37, 189–216. [Google Scholar] [CrossRef]
- Jiang, C.; Xu, L. Diversity of aquatic actinomycetes in lakes of the middle plateau, yunnan, china. Appl. Environ. Microbiol. 1996, 62, 249–253. [Google Scholar] [CrossRef] [Green Version]
- Rifaat, H.M. The biodiversity of Actinomycetes in the River Nile exhibiting antifungal activity. J. Mediterr. Ecol. 2003, 4, 5–8. [Google Scholar]
- Shrestha, B.; Nath, D.K.; Maharjan, A.; Poudel, A.; Pradhan, R.N.; Aryal, S. Isolation and Characterization of Potential Antibiotic-Producing Actinomycetes from Water and Soil Sediments of Different Regions of Nepal. Int. J. Microbiol. 2021, 2021, 5586165. [Google Scholar] [CrossRef]
- Zothanpuia; Passari, A.K.; Leo, V.V.; Chandra, P.; Kumar, B.; Nayak, C.; Hashem, A.; Allah, E.F.A.; Alqarawi, A.A.; Singh, B.P. Bioprospection of actinobacteria derived from freshwater sediments for their potential to produce antimicrobial compounds. Microb. Cell Factories 2018, 17, 68. [Google Scholar] [CrossRef]
- Eccleston, G.P.; Brooks, P.R.; Kurtböke, D.I. The occurrence of bioactive micromonosporae in aquatic habitats of the Sunshine Coast in Australia. Mar. Drugs 2008, 6, 243–261. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; An, R.; Wang, J.; Sun, N.; Zhang, S.; Hu, J.; Kuai, J. Exploring novel bioactive compounds from marine microbes. Curr. Opin. Microbiol. 2005, 8, 276–281. [Google Scholar] [CrossRef] [PubMed]
- Farrell, M.J.; Govender, D.; Hajibabaei, M.; van der Bank, M.; Davies, T.J. Bacterial diversity in the waterholes of the Kruger National Park: An eDNA metabarcoding approach (1). Genome 2019, 62, 229–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, D.; Xia, J.; Song, J.; Sun, H.; Xu, W. Using eDNA to Identify the Dynamic Evolution of Multi-Trophic Communities Under the Eco-Hydrological Changes in River. Front. Environ. Sci. 2022, 10, 853. [Google Scholar] [CrossRef]
- Coates, D.P.O.; Suntornratana, U.; Tung, N.T.; Viravong, S. Biodiversity and fisheries in the Lower Mekong Basin; Phnom Penh: Mekong development series. Mekong River Comm. Phnom Penh Cambodia 2003, 2, 30. [Google Scholar]
- Zhou, J.; Bruns, M.A.; Tiedje, J.M. DNA recovery from soils of diverse composition. Appl. Environ. Microbiol. 1996, 62, 316–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Nichols, D.; Cahoon, N.; Trakhtenberg, E.M.; Pham, L.; Mehta, A.; Belanger, A.; Kanigan, T.; Lewis, K.; Epstein, S.S. Use of ichip for high-throughput in situ cultivation of “uncultivable” microbial species. Appl. Environ. Microbiol. 2010, 76, 2445–2450. [Google Scholar] [CrossRef] [Green Version]
- Suriyachadkun, C.; Chunhametha, S.; Thawai, C.; Tamura, T.; Potacharoen, W.; Kirtikara, K.; Sanglier, J.J. Planotetraspora thailandica sp. nov., isolated from soil in Thailand. Int. J. Syst. Evol. Microbiol. 2009, 59, 992–997. [Google Scholar] [CrossRef] [Green Version]
- Drechsler, C. Morphology of the Genus Actinomyces. II. Bot. Gaz. 1919, 67, 147–168. [Google Scholar] [CrossRef]
- Kanokratana, P.; Chanapan, S.; Pootanakit, K.; Eurwilaichitr, L. Diversity and abundance of Bacteria and Archaea in the Bor Khlueng Hot Spring in Thailand. J. Basic Microbiol. 2004, 44, 430–444. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data; Babraham Bioinformatics, Babraham Institute: Cambridge, UK, 2010. [Google Scholar]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Parks, D.H.; Chuvochina, M.; Rinke, C.; Mussig, A.J.; Chaumeil, P.-A.; Hugenholtz, P. GTDB: An ongoing census of bacterial and archaeal diversity through a phylogenetically consistent, rank normalized and complete genome-based taxonomy. Nucleic Acids Res. 2022, 50, D785–D794. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Oksanen, J.; Simpson, G.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.; Hara, R.; Solymos, P.; Stevens, H.; Szöcs, E.; et al. Vegan Community Ecology Package. Version 2.6. 2 April 2022. [Google Scholar]
- Vavrek, M. Fossil: Palaeoecological and Palaeogeographical Analysis Tools. Palaeontol. Electron. 2011, 14, 16. [Google Scholar]
- Whitman, W.B.; Sutcliffe, I.C.; Rossello-Mora, R. Proposal for changes in the International Code of Nomenclature of Prokaryotes: Granting priority to Candidatus names. Int. J. Syst. Evol. Microbiol. 2019, 69, 2174–2175. [Google Scholar] [CrossRef]
- Ezeobiora, C.E.; Igbokwe, N.H.; Amin, D.H.; Enwuru, N.V.; Okpalanwa, C.F.; Mendie, U.E. Uncovering the biodiversity and biosynthetic potentials of rare actinomycetes. Future J. Pharm. Sci. 2022, 8, 23. [Google Scholar] [CrossRef]
- Zothanpuia; Passari, A.K.; Chandra, P.; Leo, V.V.; Mishra, V.K.; Kumar, B.; Singh, B.P. Production of Potent Antimicrobial Compounds from Streptomyces cyaneofuscatus Associated with Fresh Water Sediment. Front. Microbiol. 2017, 8, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neuenschwander, S.M.; Ghai, R.; Pernthaler, J.; Salcher, M.M. Microdiversification in genome-streamlined ubiquitous freshwater Actinobacteria. ISME J. 2018, 12, 185–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, J.J.; Lenski, E.L.; Zinser, R.E. The Black Queen Hypothesis: Evolution of Dependencies through Adaptive Gene Loss. mBio 2012, 3, e00036-12. [Google Scholar] [CrossRef] [Green Version]
- Bashenkhaeva, M.V.; Galachyants, Y.P.; Khanaev, I.V.; Sakirko, M.V.; Petrova, D.P.; Likhoshway, Y.V.; Zakharova, Y.R. Comparative analysis of free-living and particle-associated bacterial communities of Lake Baikal during the ice-covered period. J. Great Lakes Res. 2020, 46, 508–518. [Google Scholar] [CrossRef]
- Zaitlin, B.; Watson, S.B. Actinomycetes in relation to taste and odour in drinking water: Myths, tenets and truths. Water Res. 2006, 40, 1741–1753. [Google Scholar] [CrossRef]
- Kaeberlein, T.; Lewis, K.; Epstein, S.S. Isolating “Uncultivable” Microorganisms in Pure Culture in a Simulated Natural Environment. Science 2002, 296, 1127–1129. [Google Scholar] [CrossRef] [Green Version]
- dos Santos, J.D.; João, S.A.; Martín, J.; Vicente, F.; Reyes, F.; Lage, O.M. iChip-Inspired Isolation, Bioactivities and Dereplication of Actinomycetota from Portuguese Beach Sediments. Microorganisms 2022, 10, 1471. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kang, I.; Seo, J.H.; Cho, J.C. Culturing the ubiquitous freshwater actinobacterial acI lineage by supplying a biochemical ‘helper’ catalase. ISME J. 2019, 13, 2252–2263. [Google Scholar] [CrossRef]
- Kim, S.; Kang, I.; Lee, J.-W.; Jeon, C.O.; Giovannoni, S.J.; Cho, J.-C. Heme auxotrophy in abundant aquatic microbial lineages. Proc. Natl. Acad. Sci. USA 2021, 118, e2102750118. [Google Scholar] [CrossRef]
- Jiang, C.Y.; Dong, L.; Zhao, J.K.; Hu, X.; Shen, C.; Qiao, Y.; Zhang, X.; Wang, Y.; Ismagilov, R.F.; Liu, S.J.; et al. High-Throughput Single-Cell Cultivation on Microfluidic Streak Plates. Appl. Environ. Microbiol. 2016, 82, 2210–2218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonkou, M.D.M.; Mailhe, M.; Ndongo, S.; Ricaboni, D.; Morand, A.; Cornu, F.; Alou, M.T.; Bilen, M.; Andrieu, C.; Levasseur, A.; et al. Noncontiguous finished genome sequences and descriptions of Actinomyces ihuae, Actinomyces bouchesdurhonensis, Actinomyces urinae, Actinomyces marseillensis, Actinomyces mediterranea and Actinomyces oralis sp. nov. identified by culturomics. New Microbes New Infect. 2018, 25, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Zothanpuia; Passari, A.K.; Gupta, V.K.; Singh, B.P. Detection of antibiotic-resistant bacteria endowed with antimicrobial activity from a freshwater lake and their phylogenetic affiliation. PeerJ 2016, 4, e2103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, K.; Mazumder, A.; Sikdar, S.; Zhao, Y.-M.; Hao, J.; Song, C.; Wang, Y.; Sarkar, R.; Islam, S.; Zhang, Y.; et al. Streptomyces: The biofactory of secondary metabolites. Front. Microbiol. 2022, 13, 968053. [Google Scholar] [CrossRef] [PubMed]
- Kurtböke, D.İ.; Grkovic, T.; Quinn, R.J. Marine Actinomycetes in Biodiscovery. In Springer Handbook of Marine Biotechnology; Kim, S.-K., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 663–676. [Google Scholar]
- Doroghazi, J.R.; Metcalf, W.W. Comparative genomics of actinomycetes with a focus on natural product biosynthetic genes. BMC Genom. 2013, 14, 611. [Google Scholar] [CrossRef] [Green Version]
- Avalon, N.E.; Murray, A.E.; Baker, B.J. Integrated Metabolomic-Genomic Workflows Accelerate Microbial Natural Product Discovery. Anal. Chem. 2022, 94, 11959–11966. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Number of Species | Number of Genomes | Number of Species Containing BGCs with >60% Similarity | Number of >60% Homologous BGCs | Number of Non-Homologous BGCs | Number of BGCs Per Genome * |
|---|---|---|---|---|---|---|
| Acrocarpospora | 4 | 4 | 4 | 9 | 53 | 35.25 |
| Actinoallomurus | 1 | 1 | 1 | 5 | 18 | 34 |
| Actinocatenispora | 1 | 1 | 1 | 2 | 4 | 18 |
| Actinocorallia | 2 | 2 | 2 | 7 | 28 | 32.5 |
| Actinokineospora | 1 | 1 | 1 | 1 | 8 | 32 |
| Actinomadura | 14 | 15 | 14 | 49 | 140 | 34.67 |
| Actinophytocola | 1 | 1 | 1 | 4 | 8 | 44 |
| Actinoplanes | 23 | 24 | 21 | 59 | 158 | 20.08 |
| Allokutzneria | 1 | 1 | 1 | 6 | 16 | 53 |
| Amycolatopsis | 15 | 16 | 15 | 68 | 174 | 35.69 |
| Asanoa | 5 | 5 | 5 | 13 | 40 | 17.4 |
| Catellatospora | 3 | 3 | 3 | 8 | 36 | 27.33 |
| Dactylosporangium | 9 | 10 | 8 | 33 | 106 | 29.9 |
| Frankia | 1 | 1 | 1 | 2 | 16 | 29 |
| Gordonia | 9 | 9 | 8 | 12 | 72 | 16.33 |
| Herbidospora | 3 | 3 | 3 | 9 | 23 | 30 |
| Kibdelosporangium | 2 | 2 | 2 | 12 | 27 | 50 |
| Kineosporia | 1 | 1 | 1 | 2 | 8 | 21 |
| Kitasatospora | 6 | 6 | 6 | 43 | 85 | 41.67 |
| Kutzneria | 5 | 6 | 5 | 34 | 103 | 54.5 |
| Lentzea | 1 | 1 | 1 | 8 | 12 | 40 |
| Marinitenerispora | 1 | 1 | 1 | 3 | 4 | 16 |
| Microbispora | 8 | 8 | 8 | 25 | 80 | 24.75 |
| Micromonospora | 46 | 48 | 46 | 136 | 302 | 20.92 |
| Microtetraspora | 3 | 3 | 3 | 7 | 28 | 25 |
| Mycobacterium | 2 | 2 | 2 | 7 | 13 | 15 |
| Nocardia | 29 | 29 | 29 | 94 | 477 | 37.9 |
| Nocardiopsis | 1 | 1 | 1 | 5 | 6 | 23 |
| Nonomuraea | 14 | 14 | 14 | 40 | 139 | 29 |
| Phytohabitans | 5 | 5 | 5 | 12 | 35 | 27.8 |
| Phytomonospora | 1 | 1 | 1 | 2 | 5 | 17 |
| Planobispora | 4 | 6 | 4 | 18 | 63 | 31.33 |
| Planomonospora | 4 | 4 | 4 | 17 | 38 | 28.5 |
| Planosporangium | 3 | 3 | 3 | 5 | 15 | 15 |
| Plantactinospora | 2 | 2 | 2 | 3 | 10 | 19.5 |
| Polymorphospora | 1 | 1 | 1 | 4 | 11 | 33 |
| Prauserella | 3 | 4 | 3 | 12 | 14 | 20 |
| Pseudonocardia | 5 | 5 | 5 | 14 | 36 | 22 |
| Pseudosporangium | 1 | 1 | 1 | 2 | 4 | 21 |
| Rhodococcus | 5 | 5 | 5 | 5 | 30 | 16.2 |
| Saccharopolyspora | 4 | 4 | 4 | 19 | 23 | 25.5 |
| Sinosporangium | 2 | 3 | 2 | 14 | 29 | 34.33 |
| Sphaerimonospora | 2 | 2 | 2 | 6 | 16 | 18.5 |
| Sphaerisporangium | 8 | 8 | 8 | 22 | 71 | 27.5 |
| Streptacidiphilus | 2 | 2 | 2 | 9 | 18 | 24 |
| Streptomyces | 139 | 140 | 137 | 1401 | 1199 | 36.31 |
| Streptosporangium | 11 | 12 | 11 | 41 | 91 | 29.33 |
| Thermomonospora | 1 | 1 | 1 | 3 | 7 | 20 |
| Tsukamurella | 1 | 1 | 1 | 1 | 8 | 16 |
| Virgisporangium | 3 | 3 | 3 | 9 | 37 | 32.67 |
| Yinghuangia | 1 | 1 | 1 | 6 | 8 | 24 |
| Hydrology Station | Latitude | Longitude |
|---|---|---|
| Chiang Saen | 20.25691 | 100.101 |
| Chiang Khan | 17.90517 | 101.6752 |
| Nong Khai | 17.88286 | 102.7312 |
| Mukdahan | 16.58983 | 104.7398 |
| Khong Chiam | 15.33025 | 105.4863 |
| Genus | Number of Isolates | Order | Family | Species New to TBRC |
|---|---|---|---|---|
| Actinotalea | 2 | Micrococcales | Cellulomonadaceae | Actinotalea fermentans |
| Aeromicrobium | 1 | Propionibacteriales | Nocardioidaceae | Aeromicrobium erythreum |
| Agrococcus | 1 | Micrococcales | Microbacteriaceae | Agrococcus terreus |
| Agromyces | 1 | Micrococcales | Microbacteriaceae | Agromyces indicus |
| Arthrobacter | 1 | Micrococcales | Micrococcaceae | - |
| Asanoa | 1 | Micromonosporales | Micromonosporaceae | - |
| Brevibacterium | 1 | Micrococcales | Brevibacteriaceae | Brevibacterium frigoritolerans |
| Cellulomonas | 2 | Micrococcales | Cellulomonadaceae | Cellulomonas fimi, Cellulomonas oligotrophica |
| Geodermatophilus | 1 | Geodermatophilales | Geodermatophilaceae | Geodermatophilus normandii |
| Kribbella | 1 | Propionibacteriales | Kribbellaceae | Kribbella speibonae |
| Microbacterium | 2 | Micrococcales | Microbacteriaceae | Microbacterium invictum |
| Microbispora | 1 | Streptosporangiales | Streptosporangiaceae | - |
| Micromonospora | 18 | Micromonosporales | Micromonosporaceae | Micromonospora rifamycinica |
| Mycolicibacterium | 4 | Corynebacteriales | Mycobacteriaceae | Mycolicibacterium anyangense, Mycolicibacterium fluoranthenivorans, Mycolicibacterium pallens, Mycolicibacterium tokaiense |
| Nocardia | 3 | Corynebacteriales | Nocardiaceae | Nocardia grenadensis, Nocardia higoensis, Nocardia niwae |
| Nocardioides | 2 | Propionibacteriales | Nocardioidaceae | Nocardioides aquiterrae |
| Nonomuraea | 2 | Streptosporangiales | Streptosporangiaceae | Nonomuraea helvata, Nonomuraea lycopersici |
| Pseudarthrobacter | 2 | Micrococcales | Micrococcaceae | Pseudarthrobacter niigatensis, Pseudarthrobacter oxydans |
| Rhodococcus | 2 | Corynebacteriales | Nocardiaceae | Rhodococcus cerastii, Rhodococcus pedocola |
| Streptomyces | 26 | Streptomycetales | Streptomycetaceae | Streptomyces actinomycinicus, Streptomyces aurantiacus, Streptomyces badius, Streptomyces brasiliensis, Streptomyces durhamensis, Streptomyces echinatus, Streptomyces globisporus, Streptomyces griseoruber, Streptomyces mauvecolor, Streptomyces naganishii, Streptomyces nigra, Streptomyces panaciradicis, Streptomyces pluricolorescens, Streptomyces prasinopilosus, Streptomyces reticuliscabiei, Streptomyces rubiginosohelvolus, Streptomyces sannanensis, Streptomyces sindenensis, Streptomyces turgidiscabies, |
| Williamsia | 1 | Corynebacteriales | Nocardiaceae | Williamsia muralis |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kitikhun, S.; Siriarchawattana, P.; Chunhametha, S.; Suriyachadkun, C.; Rattanawaree, P.; Phithakrotchanakoon, C.; Harnpicharnchai, P.; Eurwilaichitr, L.; Ingsriswang, S. Expanding Actinomycetota Diversity in the TBRC Culture Collection through Metabarcoding and Simulated In Situ Cultivation of Thailand’s Mekong River Microbiota. Diversity 2023, 15, 663. https://doi.org/10.3390/d15050663
Kitikhun S, Siriarchawattana P, Chunhametha S, Suriyachadkun C, Rattanawaree P, Phithakrotchanakoon C, Harnpicharnchai P, Eurwilaichitr L, Ingsriswang S. Expanding Actinomycetota Diversity in the TBRC Culture Collection through Metabarcoding and Simulated In Situ Cultivation of Thailand’s Mekong River Microbiota. Diversity. 2023; 15(5):663. https://doi.org/10.3390/d15050663
Chicago/Turabian StyleKitikhun, Supattra, Paopit Siriarchawattana, Suwanee Chunhametha, Chanwit Suriyachadkun, Pattaraporn Rattanawaree, Chitwadee Phithakrotchanakoon, Piyanun Harnpicharnchai, Lily Eurwilaichitr, and Supawadee Ingsriswang. 2023. "Expanding Actinomycetota Diversity in the TBRC Culture Collection through Metabarcoding and Simulated In Situ Cultivation of Thailand’s Mekong River Microbiota" Diversity 15, no. 5: 663. https://doi.org/10.3390/d15050663
APA StyleKitikhun, S., Siriarchawattana, P., Chunhametha, S., Suriyachadkun, C., Rattanawaree, P., Phithakrotchanakoon, C., Harnpicharnchai, P., Eurwilaichitr, L., & Ingsriswang, S. (2023). Expanding Actinomycetota Diversity in the TBRC Culture Collection through Metabarcoding and Simulated In Situ Cultivation of Thailand’s Mekong River Microbiota. Diversity, 15(5), 663. https://doi.org/10.3390/d15050663