
When Are Cacti Found with Flowers and Fruits? Estimation of the Reproductive Phenology of the Genus Xiquexique Based on Herbarium Data

 , , , , ,
, , , , ,
Abstract
:
1. Introduction
2. Materials and Methods
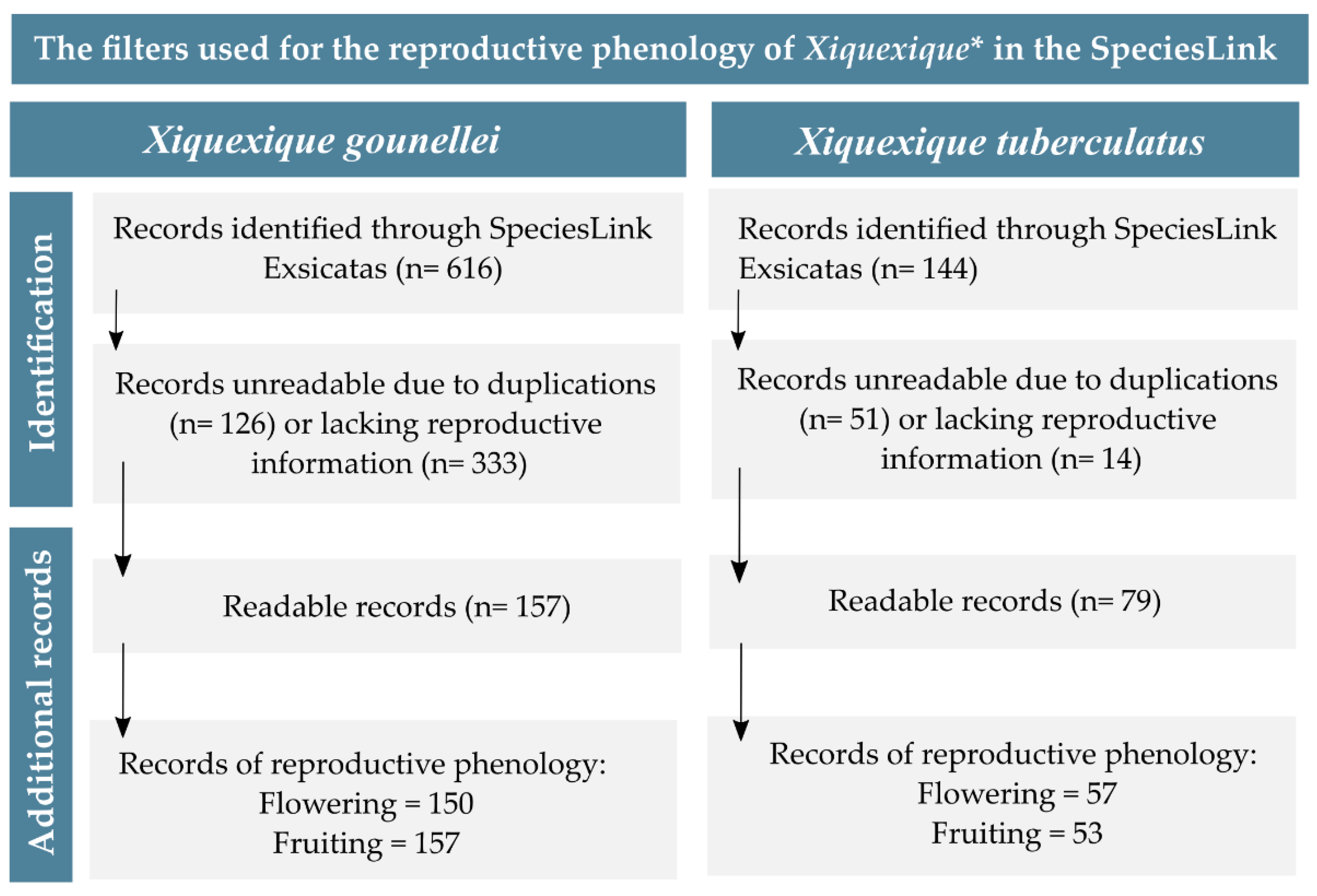
2.1. Species Selection and Phenological Data Collection
2.2. Data Analysis
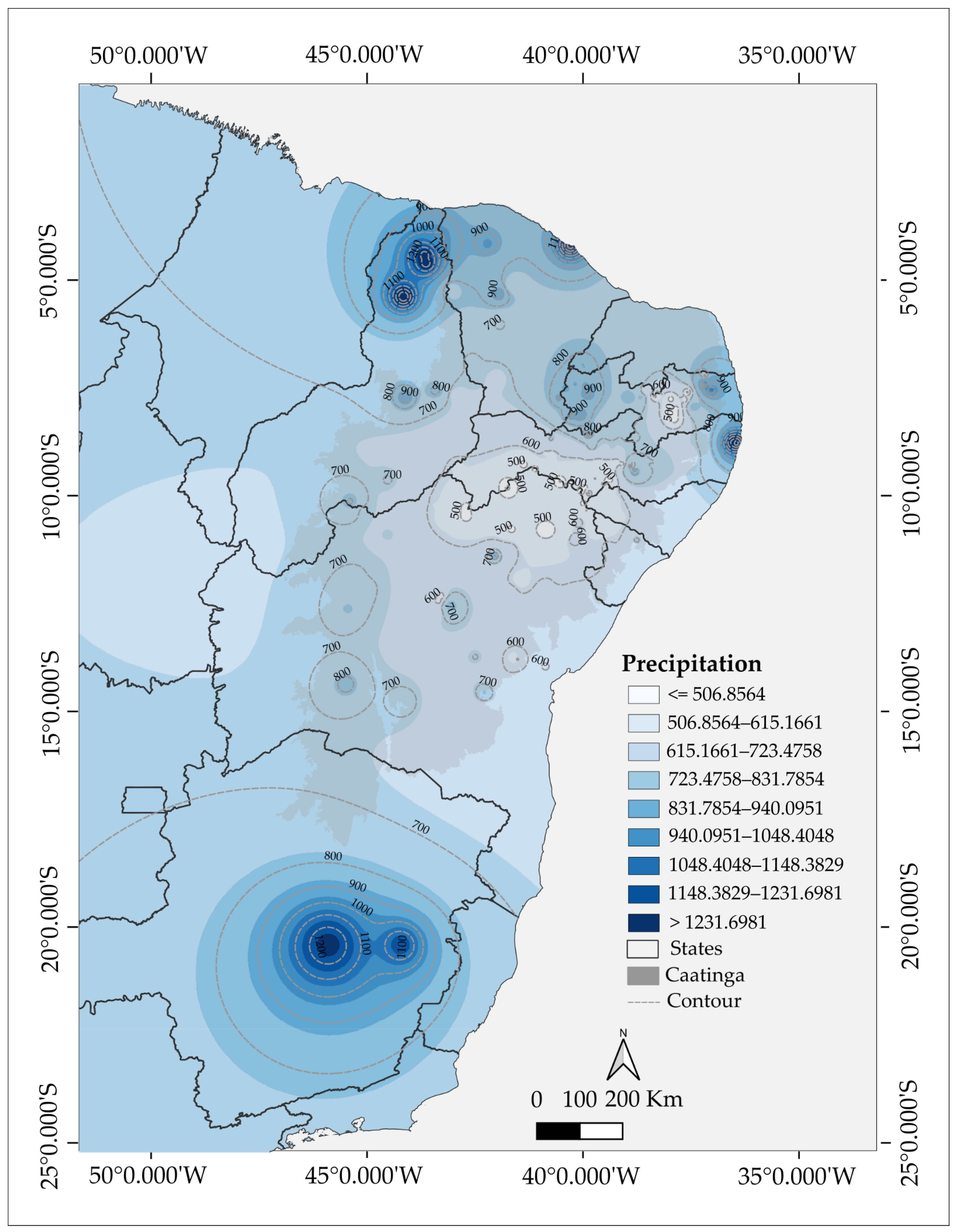
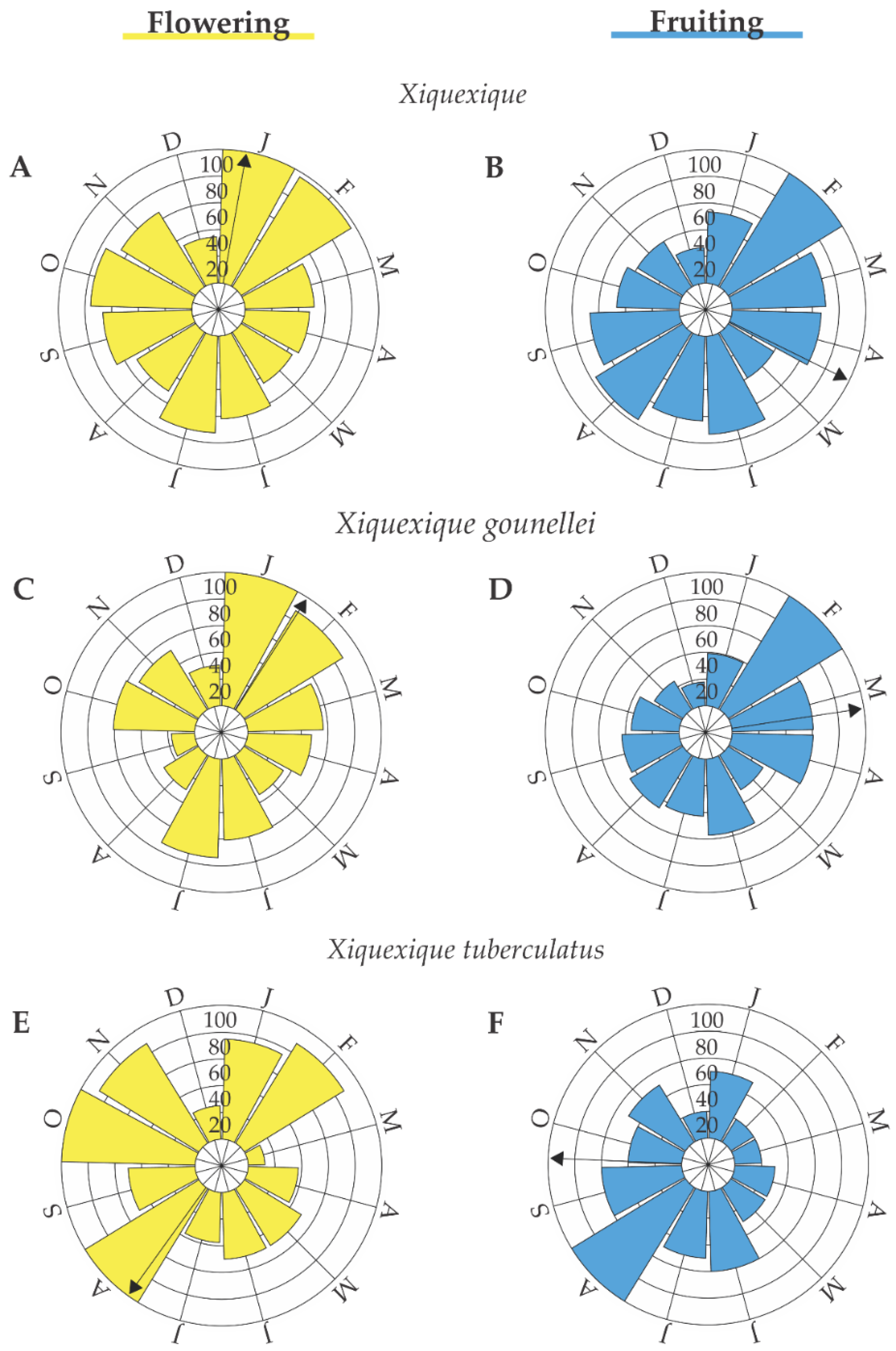
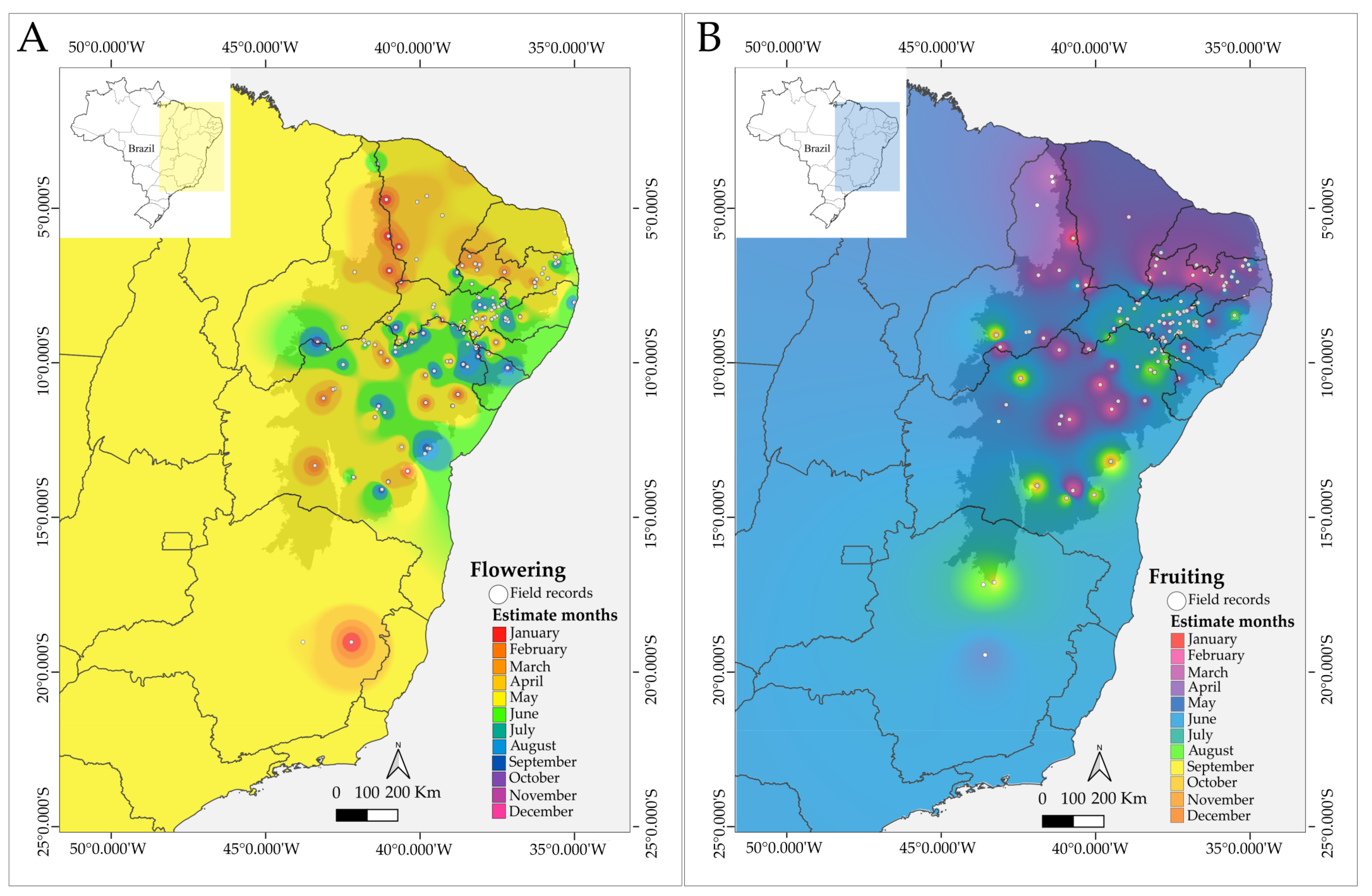
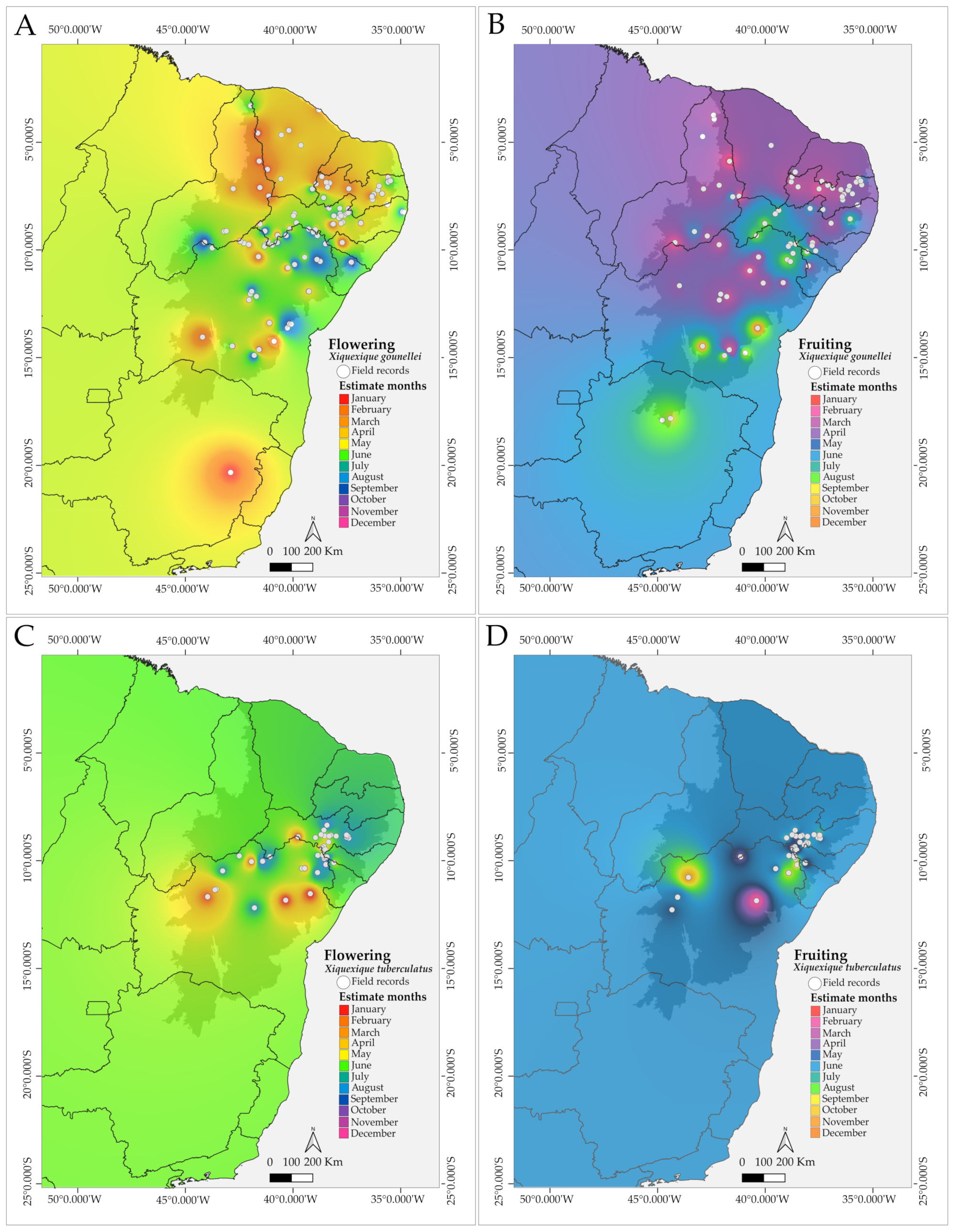
3. Results
4. Discussion
4.1. Phenological Patterns and Resource Availability in Xiquexique
4.2. Challenges and Importance of Herbarium Data for Cactaceae Phenology
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morellato, L.P.C.; Alberton, B.; Alvarado, S.T.; Borges, B.; Buisson, E.; Camargo, M.G.G.; Cancian, L.F.; Carstensen, D.W.; Escobar, D.F.E.; Leite, P.T.P.; et al. Linking plant phenology to conservation biology. Biol. Conser. 2016, 195, 60–72. [Google Scholar] [CrossRef]
- Park, D.S.; Lyra, G.M.; Ellison, A.M.; Maruyama, R.K.B.; Dos Reis Torquato, D.; Asprino, R.C.; Cook, B.I.; Davis, C.C. Herbarium records provide reliable phenology estimates in the understudied tropics. J. Ecol. 2023, 111, 327–337. [Google Scholar] [CrossRef]
- Davis, C.C.; Willis, C.G.; Connolly, B.; Kelly, C.; Ellison, A.M. Herbarium records are reliable sources of phenological change driven by climate and provide novel insights into species’ phenological cueing mechanisms. Am. J. Bot. 2015, 102, 1599–1609. [Google Scholar] [CrossRef]
- Davis, C.C.; Champ, J.; Park, D.S.; Breckheimer, I.; Lyra, G.M.; Xie, J.; Joly, A.; Tarapore, D.; Ellison, A.M.; Bonnet, P. A new method for counting reproductive structures in digitized herbarium specimens using Mask R-CNN. Front. Plant Sci. 2020, 11, 1129. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.C.; Lyra, G.M.; Park, D.S.; Asprino, R.; Maruyama, R.; Torquato, D.; Cook, B.I.; Ellison, A.M. New directions in tropical phenology. Trends Ecol. Evol. 2022, 37, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Fava, W.S.; Da Cunha, N.L.; Lorenz, A.P. Reproductive Phenology of Leptolobium dasycarpum and L. elegans across the brazilian savanna based on herbarium records. Flora 2019, 255, 34–41. [Google Scholar] [CrossRef]
- Dantas, L.V.D.A.; Pereira, V.A.D.M.; Menezes, I.S.; Miranda, L.D.A.P.D.; Funch, L.S. Reproductive phenology of Passiflora cincinnata Mast. in the semi-arid region of Bahia state, Brazil, based on herbarium data and IDW interpolation. Hoehnea 2023, 50, e162023. [Google Scholar] [CrossRef]
- Abernethy, K.; Bush, E.R.; Forget, P.; Mendoza, I.; Morellato, L.P.C. Current issues in tropical phenology: A synthesis. Biotropica 2018, 50, 477–482. [Google Scholar] [CrossRef]
- Burrough, P.A. Principals of Geographical Information Systems for Land Resources Assessment; Clarendon Press: Oxford, UK, 1986. [Google Scholar]
- Taylor, N.; Zappi, D.C. Cacti of Eastern Brazil; Royal Botanic Gardens: Kew, VIC, Australia, 2004; ISBN 1842460560. [Google Scholar]
- Tabarelli, M.; Filgueiras, B.K.C.; Ribeiro, E.M.S.; Lopes, A.V.F.; Leal, I.R. Tropical dry forests. Encycl. Biodivers. 2023, 3, 294–312. [Google Scholar]
- Rocha, E.A.; Domingos-Melo, A.; Zappi, D.C.; Machado, I.C. Reproductive biology of columnar cacti: Are bats the only protagonists in the pollination of Pilosocereus, a typical chiropterophilous genus? Folia Geobot. 2019, 54, 239–256. [Google Scholar] [CrossRef]
- Rocha, E.A.; Machado, I.C.; Zappi, D.C. Floral biology of Pilosocereus tuberculatus (Werderm.) Byles & Rowley: A bat pollinated cactus endemic from the “Caatinga” in Northeastern Brazil. Bradleya 2007, 25, 129–144. [Google Scholar] [CrossRef]
- Albuquerque-Lima, S.; Taylor, N.P.; Zappi, D.C.; Machado, I.C. Floral specialization and bat pollination in subtribe Cereinae (Cactaceae): A morphological approach. Diversity 2023, 15, 207. [Google Scholar] [CrossRef]
- Queiroz, J.A.; Diniz, U.M.; Vázquez, D.P.; Quirino, Z.M.; Santos, F.A.; Mello, M.A.; Machado, I.C. Bats and hawkmoths form mixed modules with flowering plants in a nocturnal interaction network. Biotropica 2021, 53, 596–607. [Google Scholar] [CrossRef]
- Lima-Nascimento, A.M.; Bento Da Silva, J.S.; Casas, A.; Marques de Lucena, C.; Paiva de Lucena, R.F. Traditional management of cactaceae: Cereus jamacaru DC. as the native cactus most managed by rural communities in areas of Caatinga in Brazil. Ethnobot. Res. App. 2021, 21, 1–12. [Google Scholar] [CrossRef]
- Lima-Nascimento, A.M.D.; Bento-Silva, J.S.; Lucena, C.M.D.; Lucena, R.F.P.D. Ethnobotany of native cacti in the Northeast region of Brazil: Can traditional use influence availability? Acta Bot. Bras. 2019, 33, 350–359. [Google Scholar] [CrossRef]
- Simões, S.D.S.; Zappi, D.; Costa, G.M.D.; De Oliveira, G.; Aona, L.Y.S. Spatial niche modelling of five endemic cacti from the brazilian Caatinga: Past, present and future. Austral Ecol. 2020, 45, 35–47. [Google Scholar] [CrossRef]
- Carvalho1, C.E.; Menezes, M.O.T.; Araújo, F.S.; Sfair, J.C. High endemism of cacti remains unprotected in the Caatinga. Biodivers. Conserv. 2022, 31, 1217–1228. [Google Scholar] [CrossRef]
- Goettsch, B.; Hilton-Taylor, C.; Cruz-Piñón, G.; Duffy, J.P.; Frances, A.; Hernández, H.M.; Inger, R.; Pollock, C.; Schipper, J.; Superina, M.; et al. High proportion of cactus species threatened with extinction. Nat. Plants 2015, 1, 15142. [Google Scholar] [CrossRef] [PubMed]
- Gomes, V.G.N.; Meiado, M.V.; Quirino, Z.G.M.; de Araujo, H.F.P.; Machado, I.C. Synchronous fruiting and common seed dispersers of two endemic columnar cacti in the Caatinga, a dry forest in Brazil. Plant Ecol. 2017, 218, 1325–1338. [Google Scholar] [CrossRef]
- Lavor, P.; Versieux, L.M.; Calvente, A. Phylogenetic relationships of Pilosocereus (Cactaceae) and taxonomic implications. PlantNow 2020, 1, 52–70. [Google Scholar] [CrossRef]
- Albuquerque-Lima, S.; Milet-Pinheiro, P.; Navarro, D.F.; Taylor, N.; Zappi, D.; Machado, I.C. Intermediary floral traits between natural hybrid and its parents in the Xiquexique (Cactaceae). Org. Divers. Evol. 2024, 23, 1–18. [Google Scholar] [CrossRef]
- Zappi, D.; Taylor, N. A New species of Pilosocereus subgenus Gounellea, P. frewenii, from SE Brazil. Bradleya 2011, 29, 131–136. [Google Scholar] [CrossRef]
- Morellato, L.P.C.; Alberti, L.F.; Hudson, I.L. Applications of Circular Statistics in Plant Phenology: A Case Studies Approach. In Phenological Research: Methods for Environmental and Climate Change Analysis; Hudson, I.L., Keatley, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Zar, J.H. Biostatistical Analysis; Pearson Prentice-Hall: Upper Saddle River, NJ, USA, 2010. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 8 December 2023).
- Li, J.; Heap, A.D. Spatial interpolation methods applied in the environmental sciences: A review. Environ. Modell. Softw. 2014, 53, 173–189. [Google Scholar] [CrossRef]
- Borchert, R. Phenology and flowering periodicity of neotropical dry forest species: Evidence from herbarium collections. J. Trop. Ecol. 1996, 12, 65–80. [Google Scholar] [CrossRef]
- Borchert, R.; Calle, Z.; Strahler, A.H.; Baertschi, A.; Magill, R.E.; Broadhead, J.S.; Kamau, J.; Njoroge, J.; Muthuri, C. Insolation and photoperiodic control of tree development near the Equator. New Phytol. 2015, 205, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Borchert, R.; Robertson, K.; Schwartz, M.D.; Williams-Linera, G. Phenology of temperate trees in tropical climates. Int. J. Biometeorol. 2005, 50, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Tochigi, K.; Shuri, K.; Kikuchi, S.; Naoe, S.; Koike, S.; Nagamitsu, T. Phenological shift along an elevational gradient and dispersal of pollen and seeds maintain a hybrid zone between two cherry tree species. Plant Spec. Biol. 2021, 36, 230–245. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Phenophase (n) | Mean Date | Mean Angle | Angular Standard Deviation | Mean Vector (r) | Rayleigh Test (p) |
|---|---|---|---|---|---|---|
| Xiquexique | Flowering | 11/January | 10.13° | 120.66° | 0.17 | 0.17 |
| Fruiting | 27/April | 116.24° | 120.96° | 0.08 | 0.17 | |
| X. gounellei | Flowering | 02/February | 32.62° | 113.59° | 0.14 | 0.05 |
| Fruiting | 23/March | 81.35° | 108.23° | 0.26 | 0.01 | |
| X. tuberculatus | Flowering | 02/October | 272.37° | 107.81° | 0.17 | 0.19 |
| Fruiting | 05/August | 215.73° | 93.40° | 0.26 | 0.02 |
| Taxon | Flowering | Fruiting | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Temperature | Precipitation | Temperature | Precipitation | |||||||||
| β | z or F Value | p | β | z or F Value | p | β | z or F Value | p | β | z or F Value | p | |
| Xiquexique | 0.24 | 2.33 | 0.01 | 0.004 | 2.43 | 0.01 | - | - | - | 0.004 | 2.55 | 0.01 |
| X. gounellei | 0.35 | 0.14 | 0.01 | - | - | - | 0.57 | 4.18 | <0.001 | - | - | - |
| X. tuberculatus | - | - | - | - | - | - | - | - | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bezerra-Silva, A.; Albuquerque-Lima, S.; Gomes, V.G.N.; Fagundes, A.C.d.A.; Gomes, M.T.D.; Silva, M.T.d.; Machado, I.C.; Funch, L.S. When Are Cacti Found with Flowers and Fruits? Estimation of the Reproductive Phenology of the Genus Xiquexique Based on Herbarium Data. Diversity 2024, 16, 79. https://doi.org/10.3390/d16020079
Bezerra-Silva A, Albuquerque-Lima S, Gomes VGN, Fagundes ACdA, Gomes MTD, Silva MTd, Machado IC, Funch LS. When Are Cacti Found with Flowers and Fruits? Estimation of the Reproductive Phenology of the Genus Xiquexique Based on Herbarium Data. Diversity. 2024; 16(2):79. https://doi.org/10.3390/d16020079
Chicago/Turabian StyleBezerra-Silva, Alexsandro, Sinzinando Albuquerque-Lima, Vanessa Gabrielle Nóbrega Gomes, Adelly Cardoso de Araujo Fagundes, Maria Thereza Dantas Gomes, Márjori Thays da Silva, Isabel Cristina Machado, and Ligia Silveira Funch. 2024. "When Are Cacti Found with Flowers and Fruits? Estimation of the Reproductive Phenology of the Genus Xiquexique Based on Herbarium Data" Diversity 16, no. 2: 79. https://doi.org/10.3390/d16020079
APA StyleBezerra-Silva, A., Albuquerque-Lima, S., Gomes, V. G. N., Fagundes, A. C. d. A., Gomes, M. T. D., Silva, M. T. d., Machado, I. C., & Funch, L. S. (2024). When Are Cacti Found with Flowers and Fruits? Estimation of the Reproductive Phenology of the Genus Xiquexique Based on Herbarium Data. Diversity, 16(2), 79. https://doi.org/10.3390/d16020079