
Reintroduction of the Extinct-in-the-Wild Spix’s Macaw (Cyanopsitta spixii) in the Caatinga Forest Domain of Brazil

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
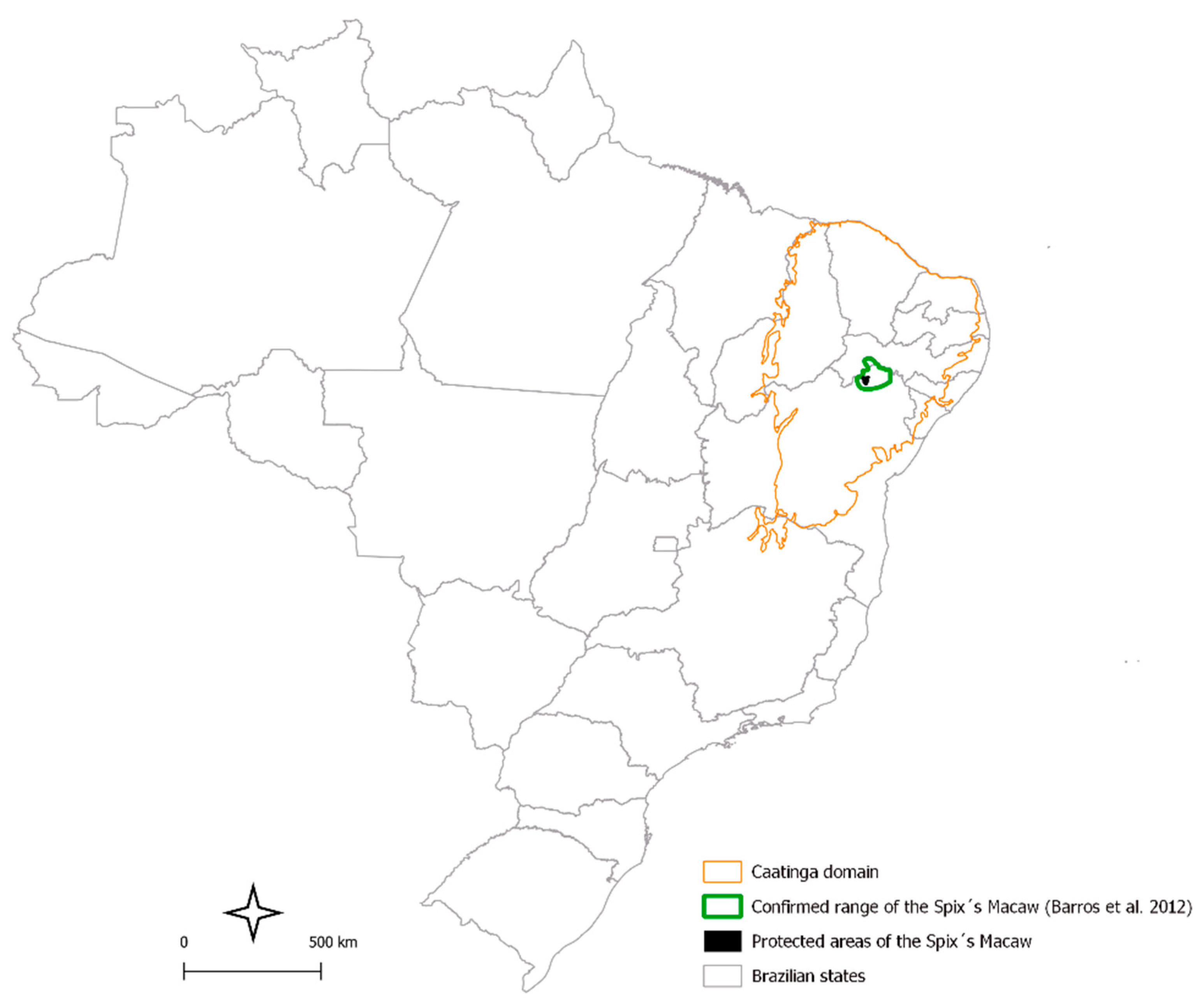
2.1. Reintroduction Site
2.2. Release Candidate Observation and Training
2.3. Release Groups
2.4. Health Protocol
2.5. Telemetry Instrumentation
2.6. Releases
2.7. Post-Release Monitoring
2.8. Habitat Management
2.9. Data Analyses
2.10. Release Success Indicators
- (1)
- Survival, assessed as the percentage of released individuals that survived at 3 months and 1 year following release.
- (2)
- Fidelity to release site, defined as the percentage of macaws that established a stable area of activity within 5 km of the release site, excluding any temporary long-distance forays.
- (3)
- Flock cohesion, defined as the percentage of surviving individuals that directly interacted (e.g., flying, foraging, perching, or roosting together). This includes any direct interactions or integration into wild heterospecific groups at or near the release site.
- (4)
- Prompt reproduction, defined here as reproduction (including egg laying) by released macaws during the initial 18 months post-release [43].
2.11. Social Engagement
3. Results
3.1. Releases
3.1.1. First Release
3.1.2. Second Release
3.2. Success Indicators
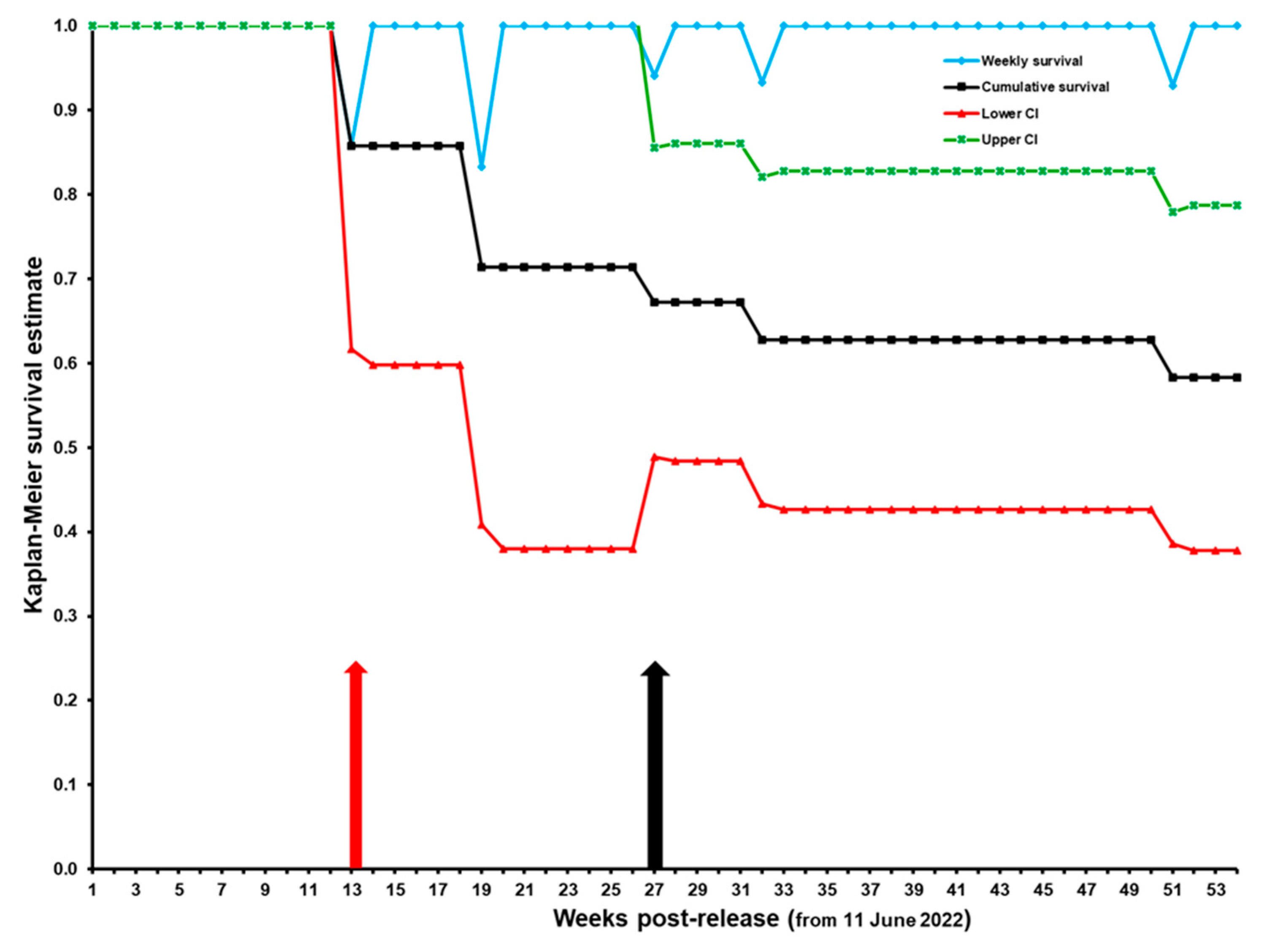
3.2.1. Survival
3.2.2. Site Fidelity and Flock Cohesion
3.2.3. Reproduction Attempts
3.2.4. Local Community Engagement
4. Discussion
4.1. Success of the Release
4.2. Tracking
4.3. The Surrogate Model Species
4.4. Protected Areas
4.5. Human Interaction
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ceballos, G.; Ehrlich, P.R.; Barnosky, A.D.; García, A.; Pringle, R.M.; Palmer, T.M. Accelerated modern human–induced species losses: Entering the sixth mass extinction. Sci. Adv. 2015, 1, 5. [Google Scholar] [CrossRef] [PubMed]
- Ceballos, G.; Ehrlich, P.R. The misunderstood sixth mass extinction. Science 2018, 360, 1080–1081. [Google Scholar] [CrossRef]
- Vergara-Tabares, D.L.; Cordier, J.M.; Landi, M.A.; Olah, G.; Nori, J. Global trends of habitat destruction and consequences for parrot conservation. Global. Chang. Biol. 2020, 26, 4251–4262. [Google Scholar] [CrossRef] [PubMed]
- Berkunsky, I.; Quillfeldt, P.; Brightsmith, D.J.; Abbud, M.C.; Aguilar, J.; Alemán-Zelaya, U.; Aramburú, R.M.; Arce Arias, A.; Balas McNab, R.; Balsby, T.J.S.; et al. Current threats faced by Neotropical parrot populations. Biol. Conserv. 2017, 214, 278–287. [Google Scholar] [CrossRef]
- Olah, G.; Butchart, S.H.M.; Symes, A.; Guzmán, I.M.; Cunningham, R.; Brightsmith, D.J.; Heinsohn, R. Ecological and socio-economic factors affecting extinction risk in parrots. Biod. Conserv. 2016, 25, 205–223. [Google Scholar] [CrossRef]
- Butchart, S.H.M.; Lowe, S.; Martin, R.W.; Symes, A.; Westrip, J.R.S.; Wheatley, H. Which bird species have gone extinct? A novel quantitative classification approach. Biol. Conserv. 2018, 227, 9–18. [Google Scholar] [CrossRef]
- BirdLife International. Species Factsheet: Cyanopsitta spixii. 2019. Available online: https://datazone.birdlife.org/species/factsheet/spixs-macaw-cyanopsitta-spixii (accessed on 4 December 2023).
- Sick, H. Ornitologia Brasileira; Nova Fronteira: Rio de Janeiro, Brazil, 1997. [Google Scholar]
- Kupferschmidt, K. A wild hope. Science 2022, 376, 1148–1153. [Google Scholar] [CrossRef] [PubMed]
- Juniper, A.T. Spix´s Macaw: The Race to Save the World´s Rarest Bird; Washington Square Press: New York, NY, USA, 2002; p. 293. [Google Scholar]
- Roth, P. Bericht fiber ein 1985–1988 durchgefuhrtes Projekt-1. Papageien 1990, 90, 86–88. [Google Scholar]
- Roth, P. Bericht fiber ein 1985–1988 durchgefuhrtes Projekt-2. Papageien 1990, 90, 121–125. [Google Scholar]
- Juniper, A.T.; Yamashita, C. The conservation of Spix´s macaw. Oryx 1990, 24, 224–228. [Google Scholar] [CrossRef]
- Juniper, A.T.; Yamashita, C. The habitat and status of Spix’s Macaw Cyanopsitta spixii. Bird Conserv. Int. 1991, 1, 1–9. [Google Scholar] [CrossRef]
- Barros, Y.M.; Soye, Y.; Miyaki, C.Y.; Watson, R.; Crosta, R.L.; Lugarini, C. Plano de Ação Nacional Para a Conservação da Ararinha-azul: Cyanopsitta spixii; Instituto Chico Mendes de Conservação da Biodiversidade: Brasilia, Brazil, 2012; p. 145. [Google Scholar]
- Barnett, J.M.; Silva, C.L.G.; Araujo, H.F.P.; Roos, A.L.; Machado, C.G.; Uejima, A.M.K.; Naka, L.N. The avifauna of Curaçá (Bahia): The last stronghold of Spix’s Macaw. Rev. Bras. Ornit. 2014, 22, 121–137. [Google Scholar] [CrossRef]
- Barros, Y.M. Biologia Comportamental de Propyrrhura maracana (Aves, Psittacidae): Fundamentos Para Conservação In Situ de Cyanopsitta spixii (Aves, Psittacidae) na Caatinga; Universidade Estadual de São Paulo: São Carlos, Brazil, 2001; p. 97. [Google Scholar]
- Miyaki, C.Y.; Faria, P.J.; Griffiths, R.; Araujo, J.C.C.; Barros, Y.M. The last Spix’s Macaw and an Illiger’s Macaw produced a hybrid. Conserv. Gen. 2001, 2, 53–55. [Google Scholar] [CrossRef]
- White, T.H., Jr.; Barros, Y.M.; Develey, P.F.; Llerandi-Román, I.C.; Monsegur-Rivera, A.O.; Trujillo-Pinto, A.M. Improving reintroduction planning and implementation through quantitative SWOT analysis. J. Nat. Conserv. 2015, 28, 149–159. [Google Scholar] [CrossRef]
- Lugarini, C.; Vercillo, E.U.; Purchase, C.; Watson, R.; Schischakin, N.A. Conservação da Ararinha-azul, Cyanopsitta spixii (Wagler, 1832): Desafios e Conquistas. Biodiversidade Bras. 2021, 11, 1–16. [Google Scholar] [CrossRef]
- Vercillo, U.; Oliveira-Santos, L.G.; Novaes, M.; Purchase, C.; Purchase, C.; Lugarini, C.; Ferreira, A.; Marco, P.; Marcuk, V.; Franco, J.L.A. Spix’s Macaw (Cyanopsitta spixii (Wagler, 1832)) population viability analysis. Bird Conserv. Int. 2023, 33, e67. [Google Scholar] [CrossRef]
- White, T.H., Jr.; Collar, N.J.; Moorhouse, R.J.; Sans, V.; Stolen, E.D.; Brightsmith, D.J. Psittacine reintroductions: Common denominators of success. Biol. Conserv. 2012, 148, 106–115. [Google Scholar] [CrossRef]
- IUCN/SSC. Guidelines for Reintroductions and Other Conservation Translocations, Version 1.0; IUCN Species Survival Commission: Gland, Suisse, 2013; p. 57. ISBN 978-2-8317-1609. [Google Scholar]
- Gobierno do Brasil. Decreto n° 9.402, de 5 de junho de 2018. Cria o Refúgio de Vida Silvestre da Ararinha Azul e a Área de Proteção Ambiental da Ararinha Azul. Available online: https://www.planalto.gov.br/ccivil_03_ato2015-2018/2018/decreto/d9402.htm (accessed on 14 February 2023).
- Marcuk, V.; Purchase, C.; de Boer, D.; Bürkle, M.; Scholtyssek, K. Qualitative description of the submission and agonistic behavior of the Spix’s Macaw (Cyanopsitta spixii, Spix 1824), with special reference to the displacement displays. J. Ethol. 2020, 38, 253–270. [Google Scholar] [CrossRef]
- Lugarini, C.; Vercillo, U. Como realizar a gestão de um projeto de alto risco? O relato da repatriação das Ararinhas-azuis ao Brasil. Biodiversidade Bras. 2021, 11, 1–10. [Google Scholar] [CrossRef]
- Ford, M.J. Selection in captivity during supportive breeding may reduce fitness in the wild. Conserv. Biol. 2002, 16, 815–825. [Google Scholar] [CrossRef]
- Frankham, R. Genetic adaptation to captivity in species conservation programs. Mol. Ecol. 2008, 17, 344–350. [Google Scholar] [CrossRef]
- White, T.H., Jr.; Collazo, J.A.; Dinsmore, S.J.; Llerandi-Román, I. Niche restriction and conservatism in a neotropical psittacine: The case of the Puerto Rican parrot. In Habitat Loss: Causes, Effects on Biodiversity and Reduction Strategies; Devore, B., Ed.; Nova Science Publishers: New York, NY, USA, 2014; pp. 1–83. [Google Scholar]
- Lenormand, T. Gene flow and the limits to natural selection. Trends Ecol. Evol. 2002, 17, 183–189. [Google Scholar] [CrossRef]
- Holt, R.D. Bringing the Hutchinsonian niche into the 21st century: Ecological and evolutionary perspectives. Proc. Nat. Acad. Sci. USA 2009, 106, 19659–19665. [Google Scholar] [CrossRef] [PubMed]
- Morse, D.H. Feeding behavior and predator avoidance in heterospecific groups. Bioscience 1977, 27, 332–339. [Google Scholar] [CrossRef]
- Herrera, C.M. Ecological aspects of heterospecific flocks formation in a Mediterranean passerine bird community. Oikos 1979, 33, 85–96. [Google Scholar] [CrossRef]
- Magrath, R.D.; Pitcher, B.J.; Gardner, J.L. A mutual understanding? Interspecific responses by birds to each other’s aerial alarm calls. Behav. Ecol. 2007, 18, 944–951. [Google Scholar] [CrossRef]
- Thiollay, J.M. Frequency of mixed species flocking in tropical forest birds and correlates of predation risk: An intertropical comparison. J. Avian Biol. 1999, 30, 282–294. [Google Scholar] [CrossRef]
- Goodale, E.; Kotagama, S.W. Response to conspecific and heterospecific alarm calls in mixed-species bird flocks of a Sri Lankan rainforest. Behav. Ecol. 2008, 19, 887–894. [Google Scholar] [CrossRef]
- Sridhar, H.; Beauchamp, G.; Shanker, K. Why do birds participate in mixed-species foraging flocks? A large-scale synthesis. Anim. Behav. 2009, 78, 337–347. [Google Scholar] [CrossRef]
- Martinez, A.E.; Zenil, R.T. Foraging guild influences dependence on heterospecific alarm calls in Amazonian bird flocks. Behav. Ecol. 2012, 23, 544–550. [Google Scholar] [CrossRef]
- Martinez, A.E.; Parra, E.; Gomez, J.P.; Vredenburg, V.T. Shared predators between primate groups and mixed species bird flocks: The potential for forest-wide eavesdropping networks. Oikos 2022, 2022, e0974. [Google Scholar] [CrossRef]
- Stephens, P.A.; Sutherland, W.J. Consequences of the Allee effect for behaviour, ecology and conservation. Trends Ecol. Evol. 1999, 14, 401–405. [Google Scholar] [CrossRef]
- Courchamp, F.; Clutton-Brock, T.; Grenfell, B. Inverse density dependence and the Allee effect. Trends Ecol. Evol. 1999, 14, 405–410. [Google Scholar] [CrossRef]
- Armstrong, D.P.; Wittmer, H.U. Incorporating Allee effects into reintroduction strategies. Ecol. Res. 2011, 26, 687–695. [Google Scholar] [CrossRef]
- White, T.H., Jr.; Abreu, W.; Benitez, G.; Jhonson, A.; Lopez, M.; Ramirez, L.; Rodriguez, I.; Toledo, M.; Torres, P.; Velez, J. Minimizing potential Allee effects in psittacine reintroductions: An example from Puerto Rico. Diversity 2021, 13, 13. [Google Scholar] [CrossRef]
- Brooks, T. Are unsuccessful avian invaders rarer in the native range than successful invaders? In Biotic Homogenization; Lockwood, J.L., McKinney, M.L., Eds.; Kluwer Academic: New York, NY, USA, 2001. [Google Scholar]
- Cassey, P. Life history and ecology influences establishment success of introduced land birds. Biol. J. Linnean Soc. 2002, 76, 465–480. [Google Scholar] [CrossRef]
- Prinzing, A.; Durka, W.; Klotz, S.; Brandl, R. Which species become aliens? Evol. Ecol. Res. 2002, 4, 385–405. [Google Scholar]
- Cassey, P.; Blackburn, T.M.; Russell, G.J.; Jones, K.E.; Lockwood, J.L. Influences on the transport and establishment of exotic bird species: An analysis of the parrots (Psittaciformes) of the world. Global Chang. Biol. 2004, 10, 417–426. [Google Scholar] [CrossRef]
- Blackburn, T.M.; Cassey, P.; Lockwood, J.L. The role of species traits in the establishment success of exotic birds. Global Chang. Biol. 2009, 15, 2852–2860. [Google Scholar] [CrossRef]
- White, T.H., Jr.; Collazo, J.A.; Vilella, F.J. Survival of captive-reared Puerto Rican Parrots released in the Caribbean National Forest. Condor 2005, 107, 426–434. [Google Scholar] [CrossRef]
- Vaz, F.F.; Serafini, P.P.; Locatelli-Dittrich, R.; Meurer, R.; Durigon, E.L.; Araújo, J.; Thomazelli, L.M.; Ometto, T.; Sipinski, E.A.B.; Sezerban, R.M.; et al. Survey of pathogens in threatened wild Red-tailed Amazon Parrot (Amazona brasiliensis) nestlings in Rasa Island, Brazil. Braz. J. Microbiol. 2017, 48, 747–753. [Google Scholar] [CrossRef]
- White, T.H., Jr.; Abreu-González, W. Dummy transmitters for pre-release acclimation of captive-reared birds. Re-introduction News 2007, 26, 28–30. [Google Scholar]
- Kenward, R.E. A Manual for Wildlife Tagging; Academic Press: London, UK, 2001; p. 350. [Google Scholar]
- Godfrey, J.D.; Bryant, D.M.; Williams, M.J. Radio-telemetry increases free-living energy costs in the endangered Takahe Porphyrio mantelli. Biol. Conserv. 2003, 114, 35–38. [Google Scholar] [CrossRef]
- Davenport, L.C.; Boorsman, T.; Carrara, L.; Antas, P.T.Z.; Faria, L.; Brightsmith, D.J.; Herzog, S.K.; Soria-Auza, R.W.; Hennessey, A.B. Satellite telemetry of Blue-throated Macaws in Barba Azul Nature Reserve (Beni, Bolivia) reveals likely breeding areas. Diversity 2021, 13, 564. [Google Scholar] [CrossRef]
- Vilarta, M.R.; Wittkoff, W.; Lobato, C.; Oliveira, R.d.A.; Pereira, N.G.P.; Silveira, L.F. Reintroduction of the Golden Conure (Guaruba guarouba) in northern Brazil: Establishing a population in a protected area. Diversity 2021, 13, 198. [Google Scholar] [CrossRef]
- Koenig, S.E.; Wunderle, J.M., Jr.; Enkerlin-Hoeflich, E.C. Vines and canopy contact: A route for snake predation on parrot nests. Bird Conserv. Inter. 2007, 17, 1–15. [Google Scholar] [CrossRef]
- Pollock, K.H.; Moore, C.T.; Davidson, W.R.; Kellogg, F.E.; Doster, G.L. Survival rates of Bobwhite quail based on band recovery analyses. J. Wildl. Manage. 1989, 3, 1–6. [Google Scholar] [CrossRef]
- Pollock, K.H.; Winterstein, S.R.; Conroy, M.J. Estimation and analysis of survival distributions for radio-tagged animals. Biometrics 1989, 45, 99–109. [Google Scholar] [CrossRef]
- Tsai, K.; Pollock, K.H.; Brownie, C. Effects of violation of assumptions for survival analysis methods in radiotelemetry studies. J. Wildl. Manag. 1999, 63, 1369–1375. [Google Scholar] [CrossRef]
- Collazo, J.A.; White, T.H., Jr.; Vilella, F.J.; Guerrero, S.A. Survival of captive-reared Hispaniolan Parrots released in Parque Nacional del Este, Dominican Republic. Condor 2003, 105, 198–207. [Google Scholar] [CrossRef]
- Harrington, D.P.; Fleming, T.R. A class of rank test procedures for censored survival data. Biometrika 1982, 69, 553–566. [Google Scholar] [CrossRef]
- Byers, O.; Copsey, J.; Lees, C.; Miller, P.; Traylor-Holzer, K. Reversing the decline in threatened species through effective conservation planning. Diversity 2022, 14, 754. [Google Scholar] [CrossRef]
- Brightsmith, D.J.; Hilburn, J.; del Campo, A.; Boyd, J.; Frisius, M.; Frisius, R.; Janik, D.; Guillén, F. The use of hand-raised psittacines for reintroduction: A case study of scarlet macaws (Ara macao) in Peru and Costa Rica. Biol. Conserv. 2005, 121, 465–472. [Google Scholar] [CrossRef]
- Seddon, P.J.; Armstrong, D.P.; Maloney, R.F. Developing the science of reintroduction biology. Conserv. Biol. 2007, 21, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Berger-Tal, O.; Blumstein, D.T.; Swaisgood, R.R. Conservation translocations: A review of common difficulties and promising directions. Animal Conserv. 2019, 23, 121–131. [Google Scholar] [CrossRef]
- Harrington, L.A.; Moehrenschlager, A.; Gelling, M.; Atkinson, R.P.; Hughes, J.; Macdonald, D.W. Conflicting and complementary ethics of animal welfare considerations in reintroductions. Conserv. Biol. 2013, 27, 486–500. [Google Scholar] [CrossRef]
- Williams, S.; Haines, J. Scarlet macaw reintroduction on the Nicoya Peninsula of Costa Rica. In Global Conservation Translocation Perspectives: 2021; Soorae, P.S., Ed.; Case studies from around the globe; IUCN SSC Conservation Translocation Specialist Group, Environment Agency—Abu Dhabi and Calgary Zoo: Calgary, AB, Canada, 2021; pp. 133–136. [Google Scholar]
- Meyers, J.M. Evaluation of three radio transmitters and collar designs for Amazona. Wildl. Soc. Bull. 1996, 24, 15–20. [Google Scholar]
- Evans, B.E.I.; Ashley, J.; Marsden, S.J. Abundance, habitat use, and movements of Blue-winged Macaws (Primolius maracana) and other parrots in and around an Atlantic forest reserve. Wilson Bull. 2005, 117, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Nunes, M.F.C.; Galetti, M. Use of forest fragments by Blue-winged Macaws (Primolius maracana) within a fragmented landscape. Biodiv. Conserv. 2007, 16, 953–967. [Google Scholar] [CrossRef]
- Senar, J.C.; Carrillo-Ortiz, J.; Arroyo, L. Numbered neck collars for long-distance identification of parakeets. J. Field Ornithol. 2012, 83, 180–185. [Google Scholar] [CrossRef]
- Barve, V. Discovering and developing primary biodiversity data from social networking sites: A novel approach. Ecol. Inf. 2014, 24, 194–199. [Google Scholar] [CrossRef]
- MacPhail, V.J.; Colla, S.R. Power of the people: A review of citizen science programs for conservation. Biol. Conserv. 2020, 249, 1–15. [Google Scholar] [CrossRef]
- Burr, D.; Kline, J.; Perryman, A. A smartphone application for monitoring gopher tortoises in Florida. Fla. Sci. 2014, 77, 198–203. [Google Scholar]
- Pennington, C.; Freeborough, K.; Dashwood, C.; Dijkstra, T.; Lawrie, K. The national landslide database of Great Britain: Acquisition, communication and the role of social media. Geomorphology 2015, 249, 44–51. [Google Scholar] [CrossRef]
- Liebenberg, L.; Steventon, J.; Brahman, N.; Benadie, K.; Minye, J.; Langwane, H.; Xhukwe, Q. Smartphone Icon User Interface design for non-literate trackers and its implications for an inclusive citizen science. Biol. Conserv. 2016, 208, 155–162. [Google Scholar] [CrossRef]
- Deredec, A.; Courchamp, F. Importance of the Allee effect for reintroductions. Écoscience 2007, 14, 440–451. [Google Scholar] [CrossRef]
- Somers, M.J.; Graf, J.A.; Szkman, M.; Slotow, R.; Gusset, M. Dynamics of a small re-introduced population of wild dogs over 25 years: Allee effects and the implications of sociality for endangered species’ recovery. Oecologia 2008, 158, 239–247. [Google Scholar] [CrossRef]
- Silva, C.L.G. Densidade e Uso de Habitat de Psitacídeos na Área de Ocorrência Histórica da Extinta Ararinha-Azul; Centro de Ciências Agrárias (CCA)—Programa de Pós-Graduação em Biodiversidade: Areia, Brazil, 2016; p. 60. [Google Scholar]
- Nunes, M.F.C.; Galetti, M.; Marsden, S.; Pereira, R.S.; Peterson, A.T. Are large-scale distributional shifts of the Blue-winged Macaw (Primolius maracana) related to climate change? J. Biogeo. 2007, 34, 816–827. [Google Scholar] [CrossRef]
- Cavalcanti, L.C.S.; Rafael, L.M.; Barbosa, L.C.S.; Braz, A.M.; Ribeiro, J.R. Can landscape unit maps help the conservation of Spix’s Macaw (Cyanopsitta spixii)? Revista Ra’e Ga Espaço Geográfico Análise 2020, 8, 181. [Google Scholar] [CrossRef]
- Wilson, R.J.; Ellis, S.; Baker, J.S.; Lineham, M.E.; Whitehead, R.W.; Thomas, C.D. Large-scale patterns of distribution and persistence at the range margins of a butterfly. Ecology 2002, 83, 3357–3368. [Google Scholar] [CrossRef]
- Pichorim, M.; França Câmara, T.P.; Marques de Oliveira-Júnior, T.; Valdenor de Oliveira, D.; Galvão do Nascimento, E.P.; Mobley, J.A. A population of Blue-winged Macaw Primolius maracana in northeastern Brazil: Recommendations for a local Conservation Action Plan. Trop. Conserv. Sci. 2014, 7, 488–507. [Google Scholar] [CrossRef]
- Dias, D.M.; Massara, R.L.; Bocchiglieri, A. Use of habitats by donkeys and cattle within a protected area of the Caatinga dry forest biome in northeastern Brazil. Persp. in Ecol. Conserv. 2019, 17, 64–70. [Google Scholar] [CrossRef]
- Gomides, S.C.; Machado, T.M.; Evangelista-Vale, J.C.; Martins-Oliveira, A.T.; Pires-Oliveira, J.C.; Muller, A.; da Rosa, L.B.; Santos-Silva, D.L.; Eisenlohr, P.V. Assessing species reintroduction sites based on future climate suitability for food resources. Conserv. Biol. 2021, 35, 1821–1832. [Google Scholar] [CrossRef]
- Kanaan, V. Re-introduction of the Vinaceous-breasted Amazon at the Araucárias National Park, Santa Catarina, Brazil. In Global Re-Introduction Perspectives: 2016; Soorae, P.S., Ed.; Case-studies from around the globe; IUCN.SSC Reintroduction Specialist Group: Abu Dhabi, UAE, 2016; pp. 106–110. [Google Scholar]
- Martins, F.C.; Engel, M.T.; Schulz, F.; Martins, C.S.G. Human dimensions of the reintroduction of Brazilian birds. Front. Conserv. Sci. 2022, 3, 791103. [Google Scholar] [CrossRef]
- Barbosa, A.; Aécio, J.; Nóbrega, A.; Alves, N.; Romeu, R. Aspectos da caça e comércio ilegal da avifauna silvestre por populações tradicionais do semiárido paraibano. Rev. Biol. Ciên. Terra 2010, 10, 39–49. [Google Scholar]
- Ndenecho, E.N. Ecological planning and ecotourism development in Kimbi Game Reserve, Cameroon. J. Human Ecol. 2009, 27, 105–113. [Google Scholar]
- Estrada, A. Reintroduction of the Scarlet Macaw (Ara macao cyanoptera) in the Tropical Rainforests of Palenque, Mexico: Project Design and First Year Progress. Trop. Conserv. Sci. 2014, 7, 342–364. [Google Scholar] [CrossRef]
- Dayer, A.A.; Silva-Rodríguez, E.A.; Albert, S.; Chapman, M.; Zukowski, B.; Ibarra, J.T.; Gifford, G.; Echeverri, A.; Martínez-Salinas, A.; Sepúlveda-Luque, C. Applying conservation social science to study the human dimensions of Neotropical bird conservation. Condor 2020, 122, 1–15. [Google Scholar] [CrossRef]
- McKinley, D.C.; Miller-Rushing, A.J.; Ballard, H.L.; Bonney, R.; Brown, P.; Cook-Patton, S.C.; Evans, D.M.; French, R.A.; Parrish, J.K.; Phillips, T.B.; et al. Citizen science can improve conservation science, natural resource management, and environmental protection. Biol. Conserv. 2017, 208, 15–28. [Google Scholar] [CrossRef]
- Monroe, M.C. Two avenues for encouraging conservation behaviors. Human Ecol. 2003, 10, 113–125. [Google Scholar]
- Toomey, A.H.; Domrose, M.C. Can citizen science lead to positive conservation attitudes and behaviors? Res. Human Ecol. 2013, 20, 50–62. [Google Scholar]
- Carneiro, A.P.B.; Jimenez, J.E.; White, T.H., Jr.; Soto-Gamboa, M.R. Distribution of Slender-billed parakeets (Enicognathus leptorthynchus) in a fragmented agricultural landscape of southern Chile. Ornitol. Neotrop. 2012, 23, 201–213. [Google Scholar]
- White, T.H., Jr.; Jimenez, J.E. Lophozonia tree cavities used for nesting by Slender-billed Parakeets (Enicognathus leptorhynchus) in the central valley of southern Chile: A potentially vanishing keystone resource. Avian Res. 2017, 8, 3. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Purchase, C.; Lugarini, C.; Purchase, C.; Ferreira, A.; Vercillo, U.E.; Stafford, M.L.; White, T.H., Jr. Reintroduction of the Extinct-in-the-Wild Spix’s Macaw (Cyanopsitta spixii) in the Caatinga Forest Domain of Brazil. Diversity 2024, 16, 80. https://doi.org/10.3390/d16020080
Purchase C, Lugarini C, Purchase C, Ferreira A, Vercillo UE, Stafford ML, White TH Jr. Reintroduction of the Extinct-in-the-Wild Spix’s Macaw (Cyanopsitta spixii) in the Caatinga Forest Domain of Brazil. Diversity. 2024; 16(2):80. https://doi.org/10.3390/d16020080
Chicago/Turabian StylePurchase, Cromwell, Camile Lugarini, Candice Purchase, Ariane Ferreira, Ugo Eichler Vercillo, Mark L. Stafford, and Thomas H. White, Jr. 2024. "Reintroduction of the Extinct-in-the-Wild Spix’s Macaw (Cyanopsitta spixii) in the Caatinga Forest Domain of Brazil" Diversity 16, no. 2: 80. https://doi.org/10.3390/d16020080
APA StylePurchase, C., Lugarini, C., Purchase, C., Ferreira, A., Vercillo, U. E., Stafford, M. L., & White, T. H., Jr. (2024). Reintroduction of the Extinct-in-the-Wild Spix’s Macaw (Cyanopsitta spixii) in the Caatinga Forest Domain of Brazil. Diversity, 16(2), 80. https://doi.org/10.3390/d16020080