Spacio-Temporal Distribution and Tourist Impact on Airborne Bacteria in a Cave (Škocjan Caves, Slovenia)


Abstract
:1. Introduction
2. Materials and Methods
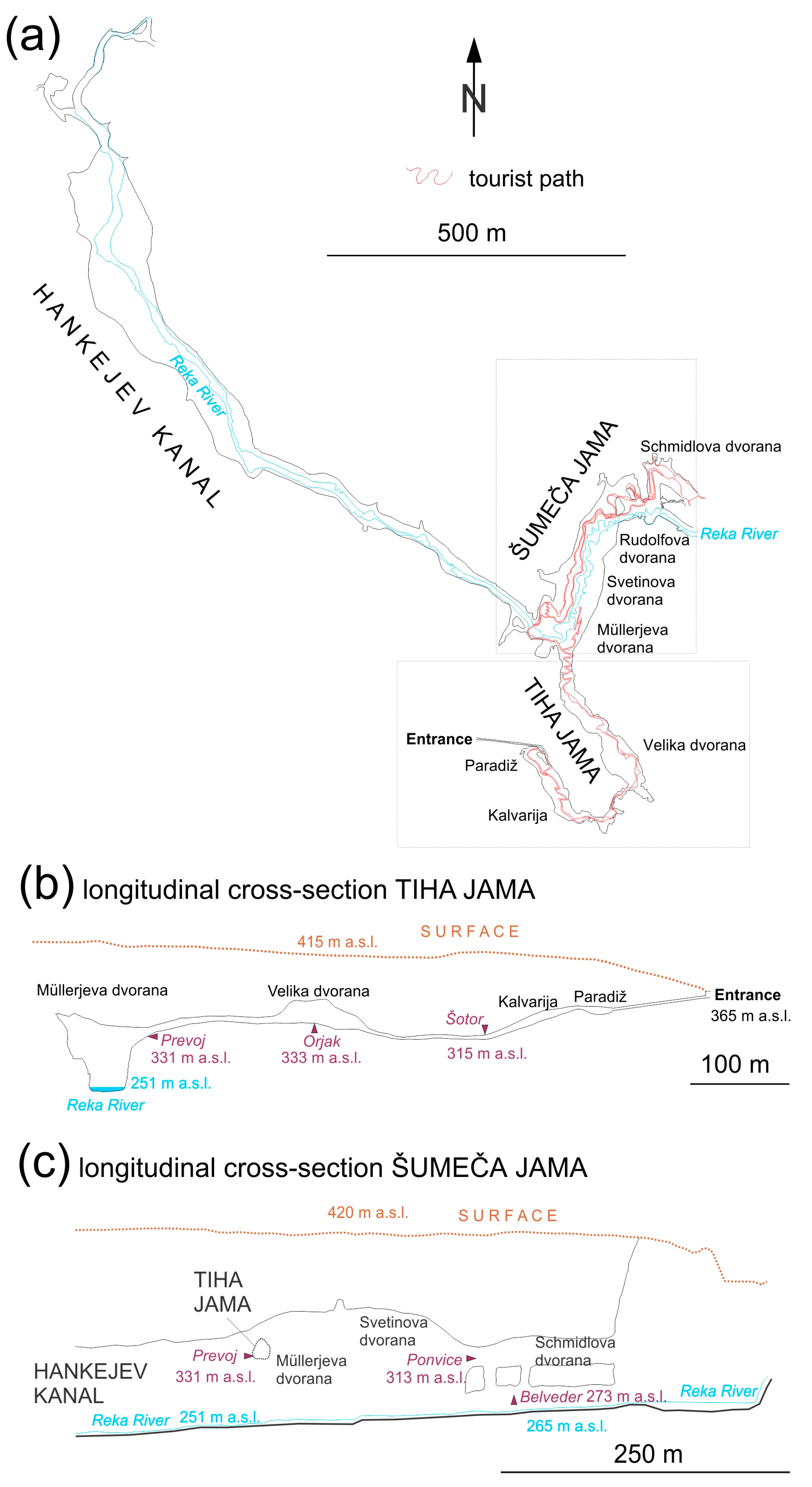
2.1. Cave Description and Sites of Bioaerosol Impacting
2.2. Atmospheric Conditions and Cultivation of Airborne Bacteria
2.3. Characterisation of Isolates
2.4. Statistical Evaluation
3. Results and Discussion
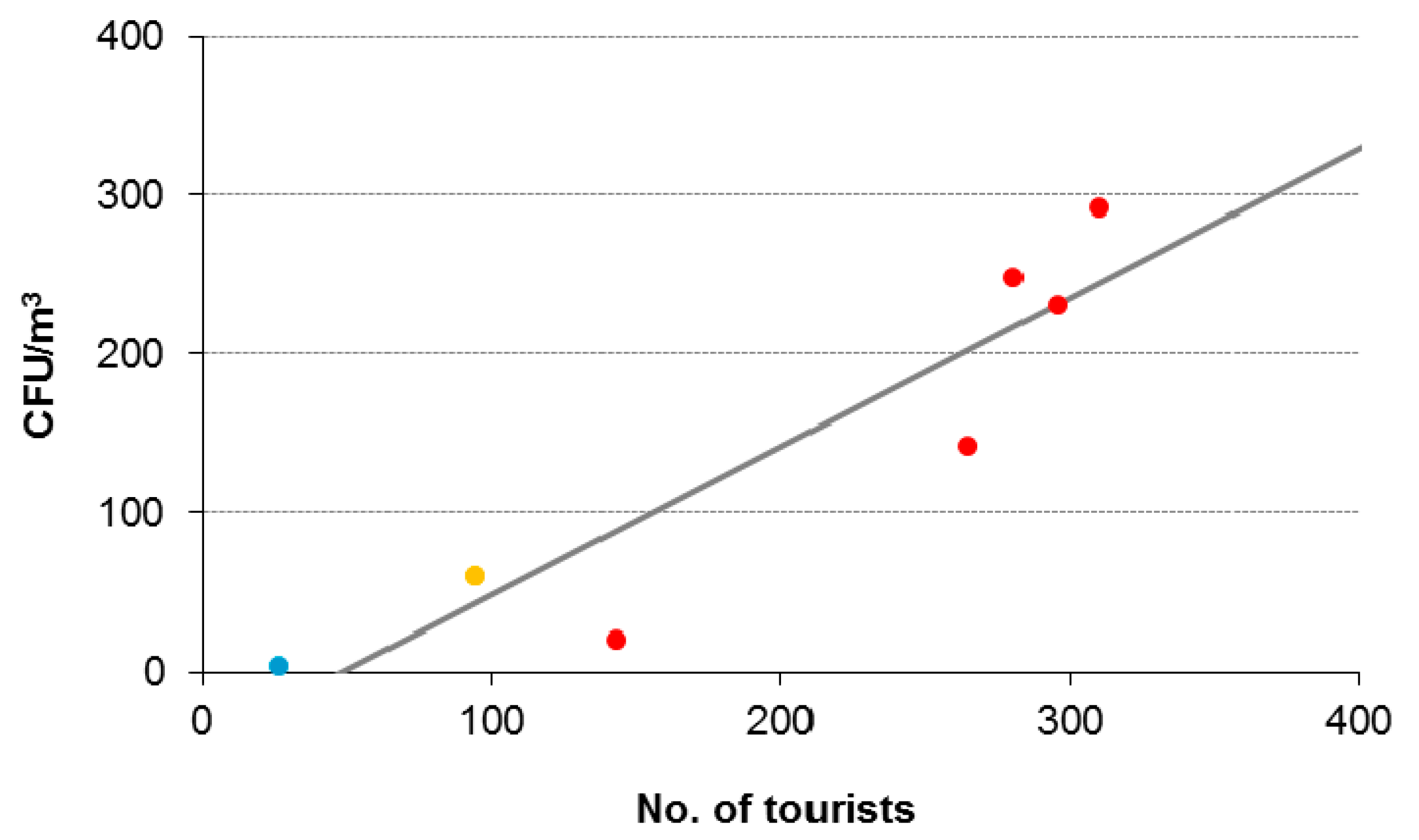
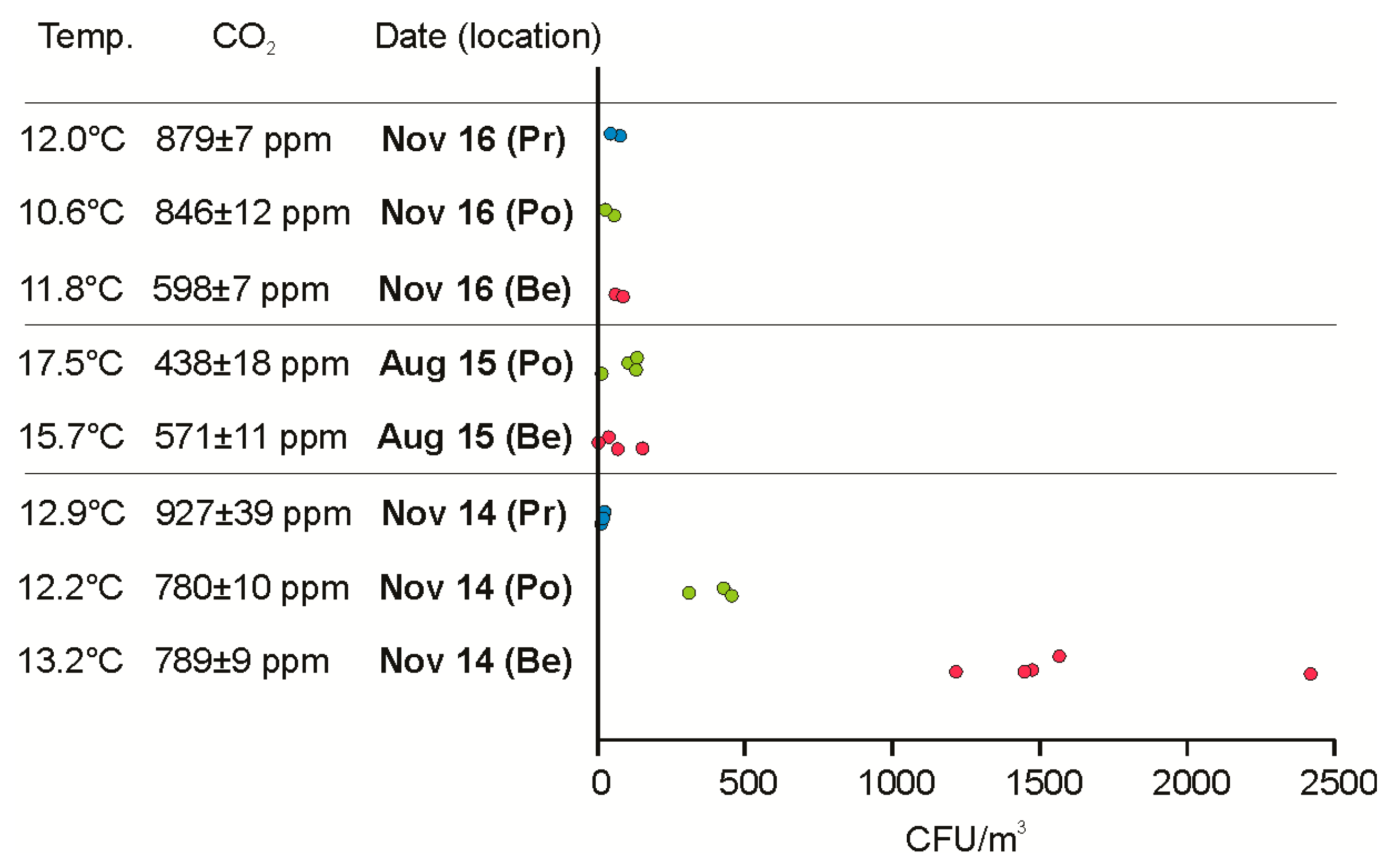
3.1. Tourist-Derived Cave Air Biomass
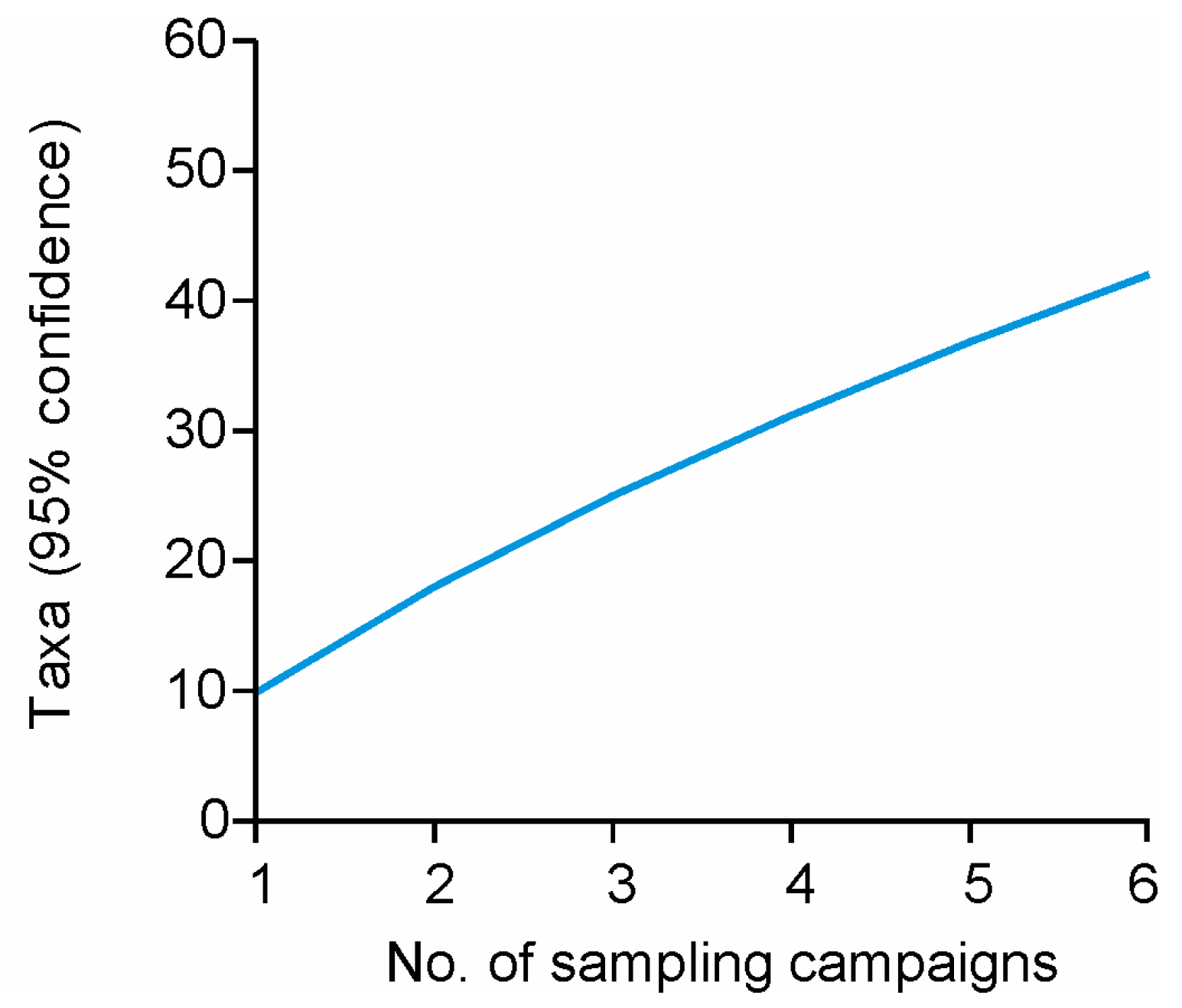
3.2. Diversity of Human-Related Airborne Bacteria
3.3. Attribution of River Aerosolization to Airborne Biomass
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hibbing, M.; Fuqua, C.; Parsek, M.; Peterson, S. Bacterial competition: Surviving and thriving in the microbial jungle. Nat. Rev. Microbiol. 2010, 8, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Adams, R.; Bhangar, S.; Pasut, W.; Arens, E.; Taylor, J.; Lindow, S.; Nazaroff, W.; Bruns, T. Chamber bioaerosol study: Outdoor air and human occupants as sources of indoor airborne microbes. PLoS ONE 2015, 10, e0128022. [Google Scholar] [CrossRef] [PubMed]
- Kellogg, C.; Griffin, D. Aerobiology and the global transport of desert dust. Trends Ecol. Evol. 2006, 21, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Mulec, J.; Vaupotič, J.; Walochnik, J. Prokaryotic and eukaryotic airborne microorganisms as tracers of microclimatic changes in the underground (Postojna Cave, Slovenia). Microb. Ecol. 2012, 64, 654–667. [Google Scholar] [CrossRef] [PubMed]
- Mulec, J. Human impact on underground cultural and natural heritage sites, biological parameters of monitoring and remediation actions for insensitive surfaces: Case of Slovenian show caves. J. Nat. Conserv. 2014, 22, 132–141. [Google Scholar] [CrossRef]
- Wang, W; Ma, X.; Ma, Y.; Mao, L.; Wu, F.; Ma, X.; An, L.; Feng, H. Seasonal dynamics of airborne fungi in different caves of the Mogao Grottoes, Dunhuang, China. Int. Biodeter. Biodegr. 2010, 64, 461–466. [Google Scholar] [CrossRef]
- Wang, W.; Ma, Y.; Ma, X.; Wu, F.; Ma, X.; An, L.; Feng, H. Seasonal variations of airborne bacteria in the Mogao Grottoes, Dunhuang, China. Int. Biodeter. Biodegr. 2010, 64, 309–315. [Google Scholar] [CrossRef]
- Fernandez-Cortes, A.; Cuezva, S.; Sanchez-Moral, S.; Canaveras, J.; Porca, E.; Jurado, V.; Martin-Sanchez, P.M.; Saiz-Jimenez, C. Detection of human-induced environmental disturbances in a show cave. Environ. Sci. Pollut. R. 2011, 18, 1037–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hospodsky, D.; Yamamoto, N.; Nazaroff, W.; Miller, D.; Gorthala, S.; Peccia, J. Characterizing airborne fungal and bacterial concentrations and emission rates in six occupied children’s classrooms. Indoor Air 2015, 25, 641–652. [Google Scholar] [CrossRef] [PubMed]
- Setlhare, G.; Malebo, N.; Shale, K.; Lues, R. Identification of airborne microbiota in selected areas in a health-care setting in South Africa. BMC Microbiol. 2014, 14, 100. [Google Scholar] [CrossRef] [PubMed]
- Bowers, R.M.; Clements, N.; Emerson, J.B.; Wiedinmyer, C.; Hannigan, M.P.; Fierer, N. Seasonal variability in bacterial and fungal diversity of the near-surface atmosphere. Environ. Sci. Technol. 2013, 47, 12097–12106. [Google Scholar] [CrossRef] [PubMed]
- Kramer, A.; Schwebke, I.; Kampf, G. How long do nosocomial pathogens persist on inanimate surfaces? A systematic review. BMC Infect. Dis. 2006, 6, 130. [Google Scholar] [CrossRef] [PubMed]
- Jurado, V.; Laiz, L.; Sanchez-Moral, S.; Saiz-Jimenez, C. Pathogenic microorganisms related to human visits in Altamira Cave, Spain. In The Conservation of Subterranean Cultural Heritage; Saiz-Jimenez, C., Ed.; CRC Press/Balkema: Boca Raton, FL, USA; London, UK; New York, NY, USA; Leiden, The Netherlands, 2014; pp. 229–239. [Google Scholar]
- Porca, E.; Jurado, V.; Martin-Sanchez, P.; Hermosin, B.; Bastian, F.; Alabouvette, C.; Saiz-Jimenez, C. Aerobiology: An ecological indicator for early detection and control of fungal outbreaks in caves. Ecol. Indic. 2011, 11, 1594–1598. [Google Scholar] [CrossRef]
- Sanchez-Moral, S.; Cuezva, S.; Garcia-Anton, E.; Fernandez-Cortes, A.; Elez, J.; Benavente, D.; Canaveras, J.C.; Jurado, V.; Rogerio Candelera, M.A.; Saiz-Jimenez, C. Microclimatic monitoring in Altamira Cave: Two decades of scientific projects for its conservation. In The Conservation of Subterranean Cultural Heritage; Saiz-Jimenez, C., Ed.; CRC Press/Balkema: Boca Raton, FL, USA; London, UK; New York, NY, USA; Leiden, The Netherlands, 2014; pp. 139–144. [Google Scholar]
- Geneste, J.M.; Mauriac, M. The Conservation of Lascaux Cave, France. In The Conservation of Subterranean Cultural Heritage; Saiz-Jimenez, C., Ed.; CRC Press/Balkema: Boca Raton, FL, USA; London, UK; New York, NY, USA; Leiden, The Netherlands, 2014; pp. 165–172. [Google Scholar]
- Gams, I.; Nagel, J. Kras: Zgodovinski, Naravoslovni in Geografski Oris (Kras/Karst: Historical, Natural and Geographical Description); Slovenska matica: Ljubljana, Yugoslavia, 1974; p. 358. [Google Scholar]
- Habič, P.; Knez, M.; Kogovšek, J.; Kranjc, A.; Mihevc, A.; Slabe, T.; Šebela, S.; Zupan, N. Škocjanske Jame speleological revue. Int. J. Speleol. 1989, 18, 1–42. [Google Scholar] [CrossRef]
- Mihevc, A. Škocjanske jame—Prispevek k speleogenezi (Škocjan Caves—A contribution to speleogenesis). Naše Jame 1999, 40, 11–27. [Google Scholar]
- Mihevc, A. Speleogeneza Divaškega Krasa; Založba ZRC: Ljubljana, Slovenia, 2001; p. 180. [Google Scholar]
- Gospodarič, R. O Geologiji in Speleogenezi Škocjanskih Jam. Geološki Zbornik 1983, 4, 163–172. [Google Scholar]
- Knez, M. The influence of bedding-planes on the development of Karst caves (a study of Velika Dolina at Škocjanske Jame caves, Slovenia). Carbonate Evaporite 1998, 13, 121–131. [Google Scholar] [CrossRef]
- Presetnik, P.; Koselj, K.; Zagmajster, M.; Zupančič, N.; Jazbec, K.; Žibrat, U.; Petrinjak, A.; Hudoklin, A. Atlas Netopirjev (Chiroptera) Slovenije (Atlas of Bats (Chiroptera) of Slovenia). Atlas Faunae et Florae Sloveniae 2; Center za kartografijo favne in flore: Miklavž na Dravskem polju, Slovenia, 2009; p. 152. [Google Scholar]
- Vode, Podatki Samodejnih Hidroloških Postaj. Available online: http://www.arso.gov.si/vode/podatki/amp/ (accessed on 24 November 2016).
- Feller, W. An Introduction to the Probability Theory and Its Application; John Wiley and Sons: New York, NY, USA, 1950; 175p. [Google Scholar]
- Singhal, N.; Kumar, M.; Kanaujia, P.K.; Viridi, J.S. MALDI-TOF mass spectrometry: An emerging technology for microbial identification and diagnosis. Front. Microbial. 2015, 6, 791. [Google Scholar] [CrossRef] [PubMed]
- Santos, I.C.; Hildenbrand, Z.L.; Schung, K.A. Applications of MALDI-TOF MS in environmental microbiology. Analyst 2016, 141, 2827–2837. [Google Scholar] [CrossRef] [PubMed]
- Cassagne, C.; Normand, A.C.; L’Ollivier, C.; Ranque, S.; Piarroux, R. Performance of MALDI-TOF MS platforms for fungal identification. Mycoses 2016, 59, 678–690. [Google Scholar] [CrossRef] [PubMed]
- Risk Group Database. Available online: https://my.absa.org/Riskgroups (accessed on 11 May 2017).
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Grgić, I.; Iskra, I.; Podkrajšek, B.; Debevec Gerjevič, V. Measurements of aerosol particles in the Škocjan Caves, Slovenia. Environ. Sci. Pollut. R. 2014, 21, 1915–1923. [Google Scholar] [CrossRef] [PubMed]
- Šebela, S.; Prelovšek, M.; Turk, J. Impact of peak period visits on the Postojna Cave (Slovenia) microclimate. Theor. Appl. Climatol. 2013, 111, 51–64. [Google Scholar] [CrossRef]
- Robertson, L.D. Monitoring viable fungal and bacterial bioaerosol concentrations to identify acceptable levels for common indoor environments. Indoor Built Environ. 1997, 6, 295–300. [Google Scholar] [CrossRef]
- Adams, R.; Miletto, M.; Taylor, J.; Bruns, T. Dispersal in microbes: Fungi in indoor air are dominated by outdoor air and show dispersal limitation at short distances. ISME J. 2013, 7, 1262–1273. [Google Scholar] [CrossRef] [PubMed]
- Mulec, J.; Dietersdorfer, E.; Ustunturk-Onan, M.; Walochnik, J. Acanthamoeba and other free-living amoebae in bat guano, an extreme habitat. Parasitol. Res. 2016, 115, 1375–1383. [Google Scholar] [CrossRef] [PubMed]
- Garrity, G. (Ed.) Bergey’s Manual® of Systematic Bacteriology: Volume 2: The Proteobacteria, Part A Introductory Essays; Springer Science & Business Media: New Delhi, India, 2005; p. 304. [Google Scholar]
- Goodfellow, M.; Kämpfer, P.; Busse, H.J.; Trujillo, M.E.; Suzuki, K.I.; Ludwig, W.; Whitman, W.B. (Eds.) Bergey’s Manual® of Systematic Bacteriology: Volume 3: The Actinobacteria, Part A and B; Springer: New York, NY, USA, 2012; p. 2083. [Google Scholar]
- Reddy, G.S.; Prakash, J.S.; Prabahar, V.; Matsumoto, G.I.; Stackebrandt, E.; Shivaji, S. Kocuria polaris sp. nov., an orange-pigmented psychrophilic bacterium isolated from an Antarctic cyanobacterial mat sample. Int. J. Syst. Evol. Microbiol. 2003, 53, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Sedláček, I. Taxonomie Prokaryot; Masarykova Univerzita: Brno, Czech Republic, 2007; p. 270. [Google Scholar]
- Stackebrandt, E.; Koch, C.; Gvozdiak, O.; Schumann, P. Taxonomic dissection of the genus micrococcus: Kocuria gen. nov., Nesterenkonia gen. nov., Kytococcus gen. nov., Dermacoccus gen. nov., and Micrococcus Cohn 1872 gen. emend. Int. J. Syst. Evol. Microbiol. 1995, 45, 682–692. [Google Scholar] [CrossRef] [PubMed]
- Tvrzová, L.; Schumann, P.; Sedláček, I.; Páčová, Z.; Spröer, C.; Verbarg, S.; Kroppenstedt, R.M. Reclassification of strain CCM 132, previously classified as Kocuria varians, as Kocuria carniphila sp. nov. Int. J. Syst. Evol. Microbiol. 2005, 55, 139–142. [Google Scholar] [CrossRef] [PubMed]
- Vos, P.; Garrity, G.; Jones, D.; Krieg, N.R.; Ludwig, W.; Rainey, F.A.; Schleifer, K.H.; Whitman, W. (Eds.) Bergey’s Manual of Systematic Bacteriology: Volume 3: The Frmicutes; Springer Science & Business Media: New York, NY, USA, 2009; p. 1450. [Google Scholar]
- Mulec, J.; Oarga-Mulec, A.; Tomazin, R.; Matos, T. Characterization and fluorescence of yellow biofilms in karst caves, southwest Slovenia. Int. J. Speleol. 2015, 44, 107–114. [Google Scholar] [CrossRef]
- Kranjc, A.; Opara, B. Temperature monitoring in Škocjanske jame caves. Acta Carsol. 2002, 31, 85–96. [Google Scholar]
- Kogovšek, J.; Kranjc, A. Pollution by surface water: The Reka River. In Karst Hydrogeology and Human Activities: Impacts, Consequences and Implications, International Contributions to Hydrogeology 20; Drew, D.P., Hötzl, H., Eds.; Balkema: Rotterdam, The Netherlands; Brookfield, VT, USA, 1999; pp. 148–153. [Google Scholar]
- Di Giorgio, C.; Krempff, A.; Guiraud, H.; Binder, P.; Tiret, C.; Dumenil, G. Atmospheric pollution by airborne microorganisms in the city of Marseilles. Atmos. Environ. 1996, 30, 155–160. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling | Climate Condition | Before Tourists | After Tourists | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Date | Temp. (°C) | CO2 (ppm) | Bacteria (CFU/m3) | CO2 (ppm) | Bacteria (CFU/m3) | Tourist Count | ||||
| 22 January 2012 | 12.5 | 437 ± 11 | 11 ± 2 | 0 | ||||||
| min/max | 410 | 460 | 8 | 15 | ||||||
| 1 April 2012 | 12.7 | 593 ± 12 | 11 ± 19 | 625 ± 13 | 71 ± 32 | 95 | ||||
| min/max | 560 | 620 | 1 | 54 | 580 | 560 | 25 | 114 | ||
| 3 June 2012 | 13.0 | 1258 ± 17 | 8 ± 4 | 1284 ± 24 | 256 ± 138 | 281 | ||||
| min/max | 1210 | 1310 | 2 | 13 | 1230 | 1340 | 78 | 558 | ||
| 22 July 2012 | 13.0 | 2007 ± 38 | 91 ± 34 | 1957 ± 50 | 382 ± 253 | 310 | ||||
| min/max | 1900 | 2100 | 41 | 138 | 1820 | 2040 | 74 | 1008 | ||
| 22 August 2012 | 13.4 | 1062 ± 43 | 19 ± 7 | 1040 ± 49 | 250 ± 87 | 296 | ||||
| min/max | 960 | 1170 | 10 | 30 | 920 | 1160 | 80 | 438 | ||
| 26 September 2013 | 12.9 | 1367 ± 21 | 10 ± 4 | 1290 ± 49 | 152 ± 75 | 265 | ||||
| min/max | 1310 | 1420 | 4 | 16 | 1150 | 1310 | 46 | 394 | ||
| 3 December 2013 | 12.6 | 453 ± 10 | 2 ± 2 | † | † | † | ||||
| min/max | 430 | 480 | 0 | 6 | ||||||
| 1 July 2014 | 12.9 | 2402 ± 25 | 5 ± 3 * | 2281 ± 74 | 24 ± 19 * | 143 | ||||
| min/max | 2340 | 2480 | 1 | 11 | 2140 | 2340 | 8 | 72 | ||
| 11 November 2014 | 12.7 | 2075 ± 21 | 19 ± 13 | 2069 ± 23 | 22 ± 11 * | 27 | ||||
| min/max | 1950 | 2120 | 9 | 40 | 1950 | 2150 | 9 | 44 | ||
| 12 November 2014 | nd | nd | nd | 2256 ± 24 | 37 ± 28 | 50 | ||||
| min/max | 12.5 | 2200 | 2310 | 9 | 63 | |||||
| Species | 1 July 2014 (0) | 1 July 2014 (143) | 11 November 2014 (0) | 11 November 2014 (27) | 12 November 2014 (0) | 13 November 2014 * (0) | 12 November 2014 ∆ |
|---|---|---|---|---|---|---|---|
| Acidovorax facilis | . | . | . | + | . | . | . |
| Acinetobacter haemolyticus | . | . | . | + | . | . | . |
| Acinetobacter johnsonii | . | + | . | . | . | . | . |
| Acinetobacter junii | . | . | . | + | . | . | . |
| Acinetobacter parvus | . | . | . | + | . | . | . |
| Acinetobacter tjenbergiae | . | . | . | + | . | . | . |
| Aerococcus viridans | + | + | . | . | . | + | . |
| Aeromonas encheleia | . | . | . | . | . | . | + |
| Arthrobacter arilaitensis | + | . | + | . | . | . | . |
| Bacillus cereus | . | . | . | . | . | . | + |
| Bacillus subtilis ssp. subtilis | . | + | . | . | . | . | . |
| Bacillus marisflavi | . | . | . | . | . | . | + |
| Bacillus thuringensis | . | . | . | + | . | . | . |
| Brachybacterium faecium | . | . | . | + | . | . | . |
| Brevundimonas diminuta | . | . | . | + | . | . | + |
| Dietzia maris | . | + | . | . | + | . | . |
| Kocuria carniphila | . | . | . | + | . | . | . |
| Kocuria polaris | . | . | . | . | + | . | . |
| Kocuria rosea | . | . | . | + | . | + | . |
| Kytococcus sedentarius | . | . | . | + | . | . | . |
| Microbacterium hydrocarbonoxydans | . | . | . | + | . | . | . |
| Microbacterium sp. | . | + | . | . | . | . | . |
| Micrococcus luteus | . | + | . | + | + | + | . |
| Micrococcus terreus | . | . | . | + | . | . | . |
| Paenibacillus amyolyticus | + | . | + | . | . | . | . |
| Paenibacillus polymixa | + | . | . | . | . | . | . |
| Pseudochrobactrum asaccharolyticum | . | . | . | + | . | . | . |
| Pseudoclavibacter helvolus | . | + | . | . | . | . | . |
| Pseudomonas antarctica | . | . | + | . | . | . | . |
| Pseudomonas cedrina ssp. cedrina | . | . | + | . | . | . | . |
| Pseudomonas extremorientalis | . | . | . | . | . | . | + |
| Pseudomonas jessenii | . | . | + | . | . | . | . |
| Pseudomonas marginalis | . | . | + | . | . | . | . |
| Pseudomonas synxantha | . | . | . | . | . | . | + |
| Psyhcrobacillus psychrotolerans | + | + | + | . | . | + | . |
| Rhodococcus erythropolis | . | . | . | + | . | . | . |
| Staphylococcus capitis | . | . | . | + | . | . | . |
| Staphylococcus capitis ssp. capitis | . | . | . | + | . | . | . |
| Staphylococcus epidermidis | + | . | + | + | . | . | . |
| Staphylococcus equorum | . | . | . | . | + | + | . |
| Staphylococcus equorum ssp. equorum | . | . | . | + | . | . | . |
| Staphylococcus haemolyticus | . | . | . | . | + | . | . |
| Staphylococcus pasteuri | . | . | . | . | . | + | . |
| Staphylococcus warneri | . | . | . | . | + | . | . |
| Staphylococcus xylosus | . | . | . | + | . | + | . |
| Streptomyces badius | + | . | + | . | . | . | . |
| Total identified species | 7 | 8 | 9 | 21 | 6 | 7 | 6 |
| Species | 1 July 2014 (0) | 1 July 2014 (143) | 11 November 2014 (0) | 11 November 2014 (27) | 12. November 2014 (0) | 13 November 2014 * (0) | 12 November 2014 ∆ |
|---|---|---|---|---|---|---|---|
| Acinetobacter calcoaceticus | . | . | . | . | + | . | . |
| Acinetobacter haemolyticus | . | + | . | . | + | . | . |
| Acinetobacter haemolytius | . | . | . | + | . | . | . |
| Acinetobacter junii | . | + | . | + | . | . | . |
| Acinetobacter lwoffii | . | . | . | . | . | . | + |
| Acinetobacter parvus | . | . | . | + | . | . | . |
| Acinetobacter tjernbergiae | . | . | . | + | . | . | . |
| Aerococcus viridans | + | + | + | + | . | + | . |
| Arthrobacter arilaitensis | + | . | + | . | . | + | . |
| Bacillus arsenicus | . | . | . | + | . | . | . |
| Bacillus cereus | . | . | . | . | + | . | + |
| Bacillus flexus | . | + | . | . | . | . | . |
| Bacillus jeotgali | + | . | . | . | . | . | . |
| Bacillus licheniformis | . | + | . | . | . | . | + |
| Bacillus muralis | . | + | . | . | . | + | . |
| Bacillus pumilus | + | + | + | . | . | . | . |
| Bacillus simplex | . | + | . | . | + | + | . |
| Bacillus subtilis ssp. subtilis | . | + | . | . | . | . | . |
| Bacillus thuringiensis | . | . | . | . | . | + | + |
| Brachybacterium faecium | . | . | . | . | . | + | . |
| Brevundimonas diminuta | . | . | . | + | . | . | + |
| Carnobacterium maltaromaticum | . | . | . | . | . | . | + |
| Dietzia cinnamea | . | . | . | + | . | . | . |
| Dietzia natronolimnaea | . | + | . | . | . | . | . |
| Jeotgalicoccus halotolerans | . | . | . | + | . | . | . |
| Kocuria carniphila | . | . | . | + | . | + | . |
| Kocuria rhizophila | . | . | . | + | . | . | . |
| Kocuria rosea | . | . | . | . | . | + | . |
| Kyotococcus sedentarius | . | . | . | + | . | . | . |
| Micrococcus luteus | . | . | . | + | . | + | . |
| Micrococcus terreus | . | . | . | + | . | . | . |
| Paenibacillus polymixa | + | . | . | . | . | . | . |
| Pseudochrobactrum asaccharolyticum | . | . | . | + | . | . | . |
| Pseudomonas anguilliseptica | . | . | . | . | + | . | . |
| Pseudomonas azotoformans | . | . | + | . | . | . | . |
| Pseudomonas chlororaphis | . | . | . | . | + | . | . |
| Pseudomonas frederiksbergensis | . | . | . | . | . | . | + |
| Pseudomonas libanensis | . | . | . | . | . | . | + |
| Pseudomonas marginalis | . | . | + | . | . | . | . |
| Pseudomonas rhodesiae | . | . | + | . | . | . | . |
| Pseudomonas synxantha | . | . | . | . | . | . | + |
| Pseudomonas syringae ssp. syringae | . | . | + | . | . | . | . |
| Pseudomonas tolaasii | . | . | + | . | . | . | . |
| Psyhcrobacillus psychrotolerans | . | + | . | . | . | + | . |
| Rhodococcus erythropolis | . | . | . | . | . | + | . |
| Rhodococcus epidermidis | . | . | . | . | . | + | . |
| Staphylococcus equorum | . | . | . | + | + | + | . |
| Staphylococcus equorum ssp. equorum | . | . | . | + | . | . | . |
| Staphylococcus hominis ssp. hominis | . | . | . | + | . | . | . |
| Staphylococcus warneri | . | . | . | + | . | . | . |
| Staphylococcus xylosus | . | . | . | + | . | . | . |
| Stenotrophomonas rhizophila | . | . | . | . | . | . | + |
| Total identified genera | 5 | 11 | 8 | 20 | 7 | 13 | 10 |
| Species | Phylum | Optimum Growth Temperature (°C) | Typical Habitat (Pathogenicity) * | Risk Group (Country) |
|---|---|---|---|---|
| Acidovorax facilis | Proteobacteria | 30 | Soil | |
| Acinetobacter haemolyticus | Proteobacteria | 28 | human skin (rarely pathogen) | |
| Acinetobacter johnsonii | Proteobacteria | 26 | humans, animals, activated sludge, food (pathogen) | 2 (AU, CH, NZ) |
| Acinetobacter junii | Proteobacteria | 28 | ubiquitous, humans (pathogen) | 2 (AU, CH, NZ) |
| Acinetobacter parvus | Proteobacteria | 35 | Humans | |
| Acinetobacter tjernbergiae | Proteobacteria | 30 | aquatic, wastewater | |
| Aerococcus viridans | Firmicutes | 30 | ubiquitous, air, humans (pathogen) | 2 (CH, DE) |
| Aeromonas encheleia | Proteobacteria | 28 | aquatic, humans, animals | |
| Arthrobacter arilaitensis | Actinobacteria | 10–30 | food, cheeses | |
| Bacillus cereus | Firmicutes | 20–40 | ubiquitous (pathogen) | 2 (AU, CH, DE, NZ, SG, UK) |
| Bacillus subtilis | Firmicutes | 25–35 | Ubiquitous | |
| Bacillus marisflavi | Firmicutes | 30–37 | marine tide zone | |
| Bacillus thuringiensis | Firmicutes | 30–37 | ubiquitous, (pathogen) | |
| Brachybacterium faecium | Actinobacteria | 25–30 | soil, feaces | |
| Brevundimonas diminuta | Proteobacteria | 30–35 | ubiquitous (pathogen) | 2 (CH, DE) |
| Dietzia maris | Actinobacteria | 26 | humans, animals (pathogen) | |
| Kocuria carniphila | Actinobacteria | 28–37 | food, meat | |
| Kocuria polaris | Actinobacteria | 20 | Aquatic | |
| Kocuria rosea | Actinobacteria | 25–37 | soil, aquatic (opportunistic pathogen) | |
| Kytococcus sedentarius | Actinobacteria | 25–37 | aquatic, human skin (pathogen) | |
| Microbacterium hydrocarbonoxydans | Actinobacteria | 30–37 | crude oil | |
| Micrococcus luteus | Actinobacteria | 25–37 | soil, aquatic, air, human and animal skin | |
| Micrococcus terreus | Actinobacteria | 33 | Soil | |
| Paenibacillus amylolyticus | Firmicutes | 28–37 | Soil | |
| Paenibacillus polymyxa | Firmicutes | 30 | soil, marine sediment, plant roots | |
| Pseudochrobactrum asaccharolyticum | Proteobacteria | 20–30 | Humans | |
| Pseudoclavibacter helvolus | Actinobacteria | 28–30 | food, butter | |
| Pseudomonas antarctica | Proteobacteria | 22 | aquatic, cyanobacterial mats | |
| Pseudomonas cedrina | Proteobacteria | 21–28 | Aquatic | |
| Pseudomonas extremorientalis | Proteobacteria | 30 | Aquatic | |
| Pseudomonas jessenii | Proteobacteria | 25–30 | aquatic (opportunistic pathogen) | |
| Pseudomonas marginalis | Proteobacteria | 28–30 | plants (plant pathogen) | |
| Pseudomonas synxantha | Proteobacteria | 25–30 | Plants | |
| Psychrobacillus psychrotolerans | Firmicutes | 25 | Soil | |
| Rhodococcus erythropolis | Actinobacteria | 20 | Soil | |
| Staphylococcus capitis | Firmicutes | 30–40 | human skin (opportunistic pathogen) | |
| Staphylococcus epidermidis | Firmicutes | 26–37 | human and animal skin (pathogen) | 2 (BE, CH, DE) |
| Staphylococcus equorum | Firmicutes | 30 | human and animal skin, food, sausages, cheeses | |
| Staphylococcus haemolyticus | Firmicutes | 34–35 | human and animal skin (pathogen) | 2 (BE, CH, DE) |
| Staphylococcus pasteuri | Firmicutes | 35–37 | humans, animals, food (pathogen) | 2 (BE, CH) |
| Staphylococcus warneri | Firmicutes | 30–40 | human and animal skin | |
| Staphylococcus xylosus | Firmicutes | 25–35 | ubiquitous, human and animal skin | |
| Streptomyces badius | Actinobacteria | 28 | Soil |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mulec, J.; Oarga-Mulec, A.; Šturm, S.; Tomazin, R.; Matos, T. Spacio-Temporal Distribution and Tourist Impact on Airborne Bacteria in a Cave (Škocjan Caves, Slovenia). Diversity 2017, 9, 28. https://doi.org/10.3390/d9030028
Mulec J, Oarga-Mulec A, Šturm S, Tomazin R, Matos T. Spacio-Temporal Distribution and Tourist Impact on Airborne Bacteria in a Cave (Škocjan Caves, Slovenia). Diversity. 2017; 9(3):28. https://doi.org/10.3390/d9030028
Chicago/Turabian StyleMulec, Janez, Andreea Oarga-Mulec, Samo Šturm, Rok Tomazin, and Tadeja Matos. 2017. "Spacio-Temporal Distribution and Tourist Impact on Airborne Bacteria in a Cave (Škocjan Caves, Slovenia)" Diversity 9, no. 3: 28. https://doi.org/10.3390/d9030028
APA StyleMulec, J., Oarga-Mulec, A., Šturm, S., Tomazin, R., & Matos, T. (2017). Spacio-Temporal Distribution and Tourist Impact on Airborne Bacteria in a Cave (Škocjan Caves, Slovenia). Diversity, 9(3), 28. https://doi.org/10.3390/d9030028