Molecular Dissection of dH3w, A Fluorescent Peptidyl Sensor for Zinc and Mercury

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Steady-State Fluorescence Spectroscopy
2.3. Monte Carlo Modeling of Zn(dH3w)2
3. Results and Discussion
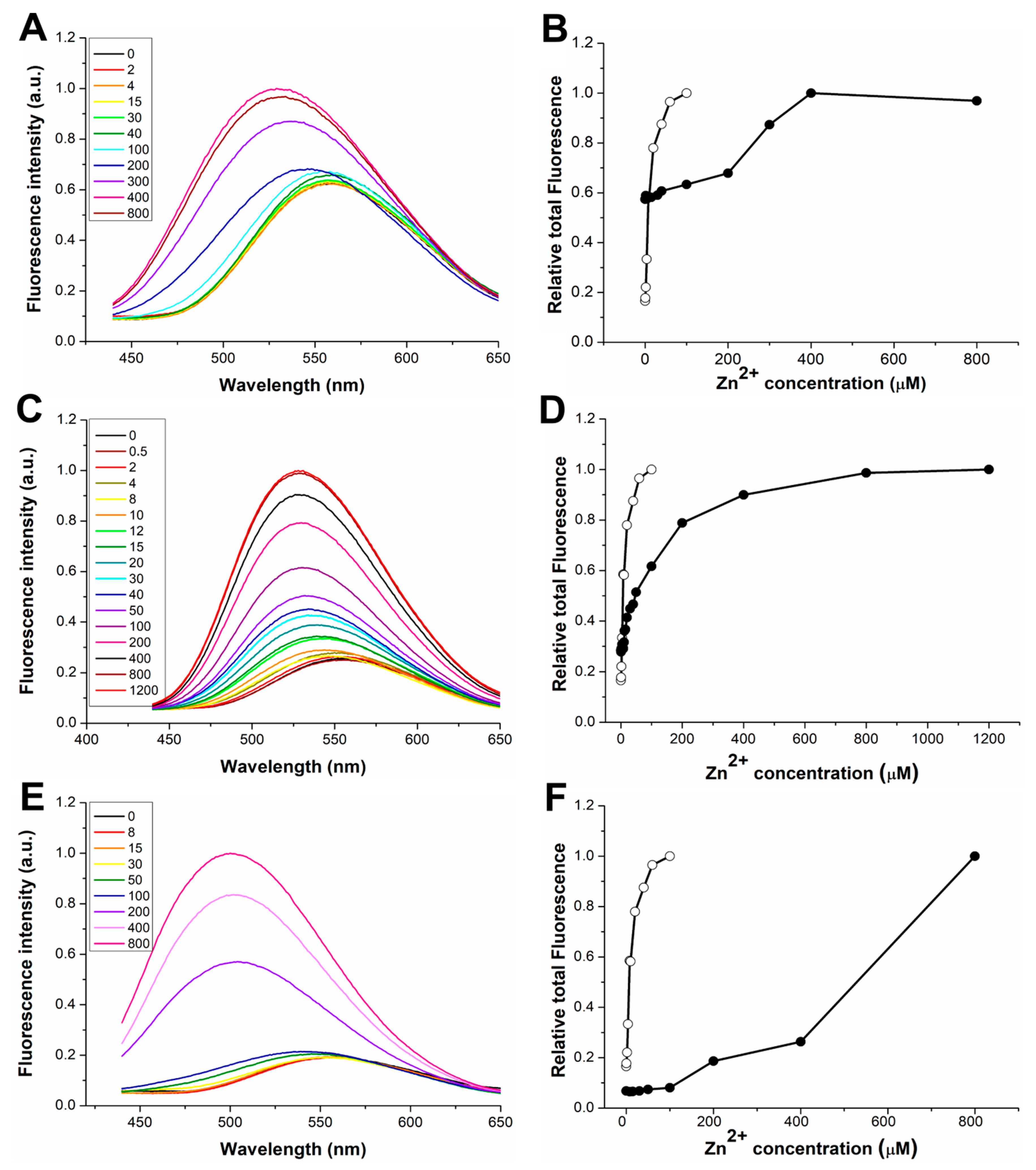
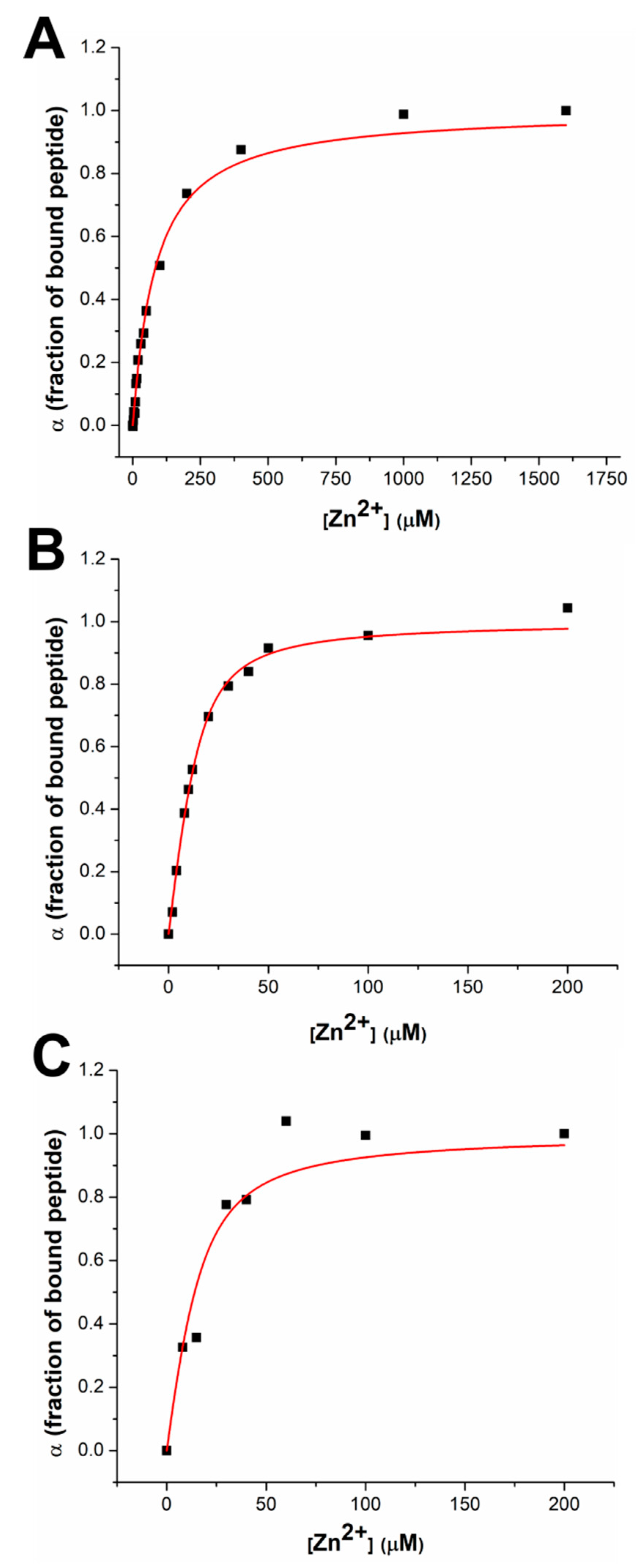
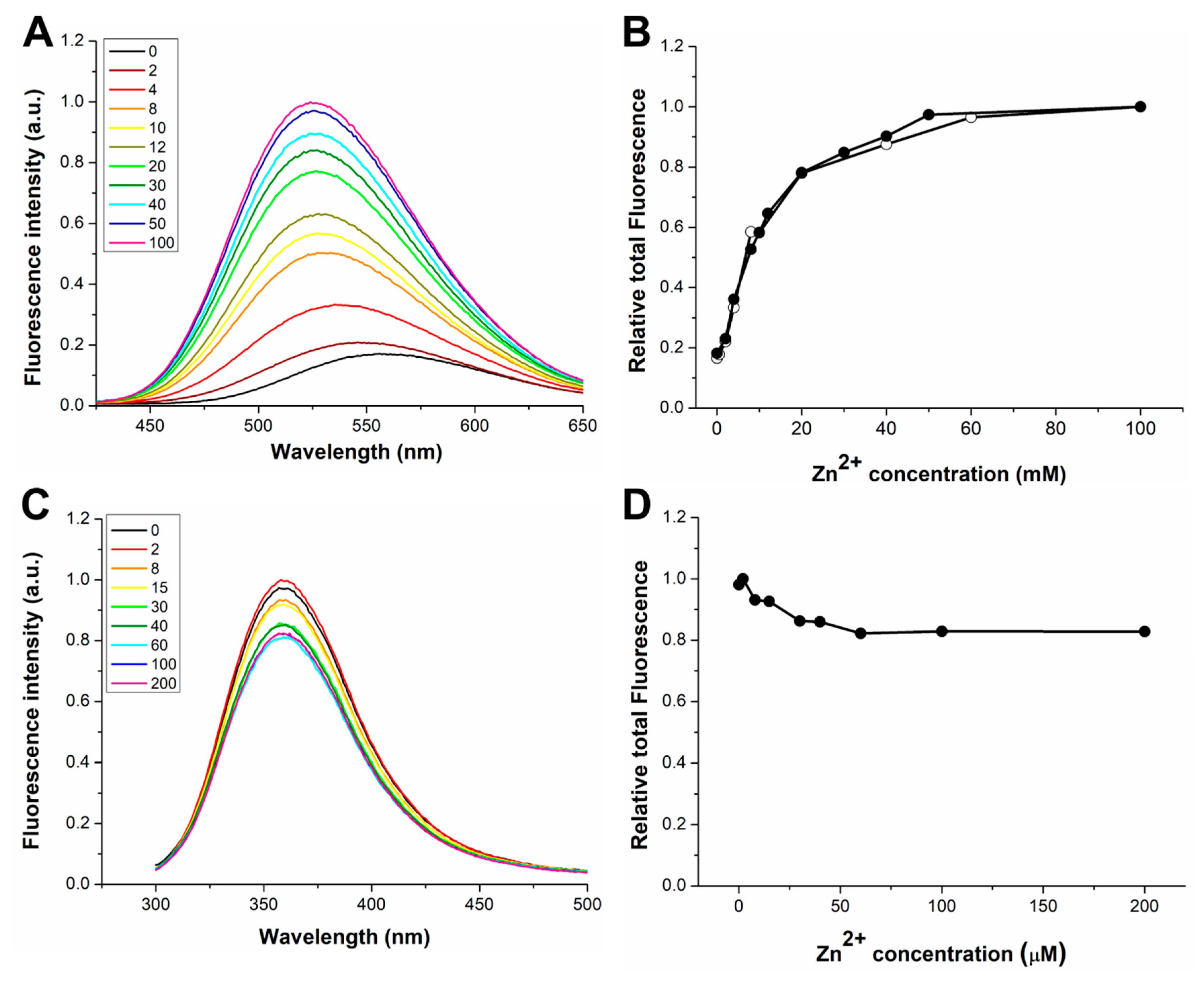
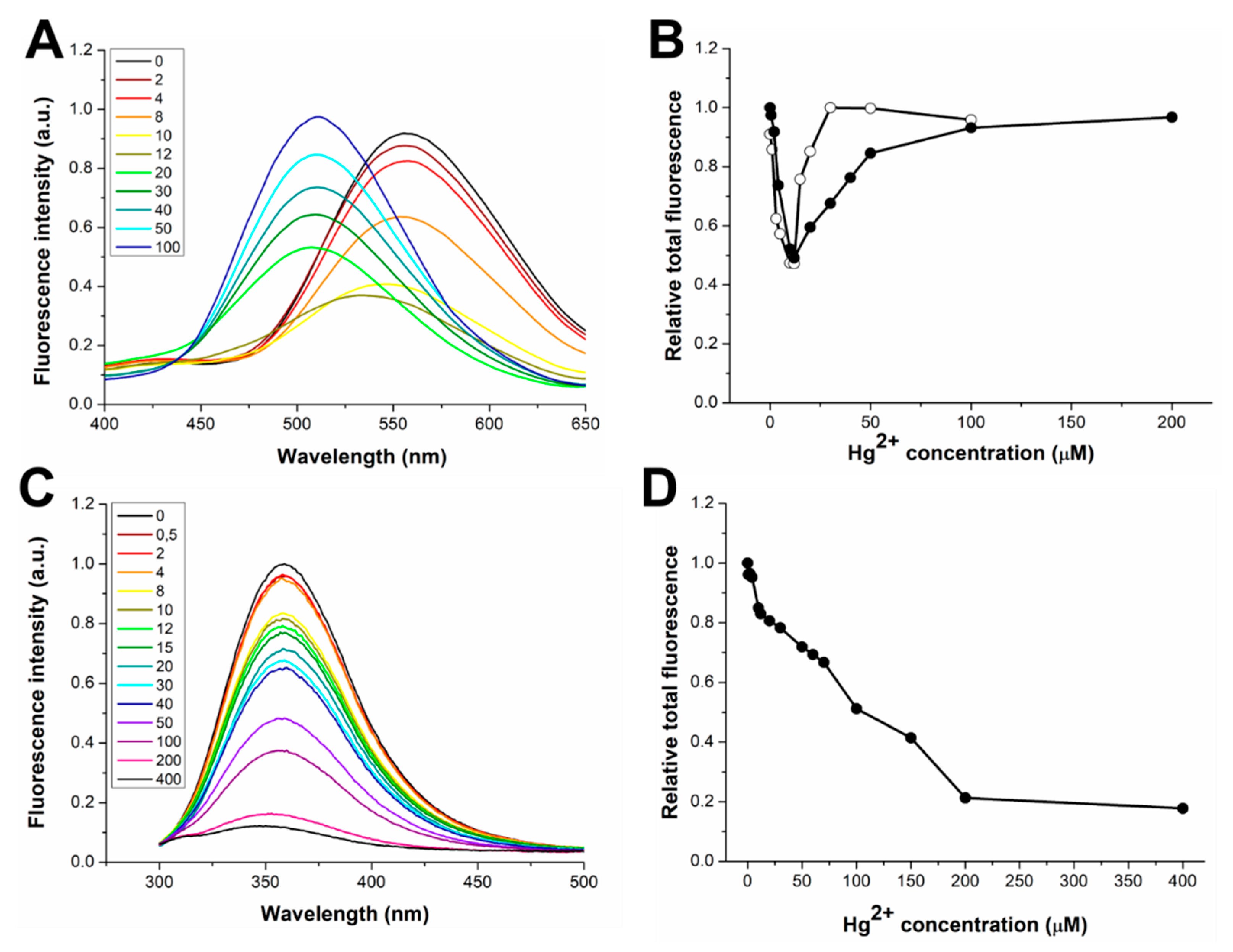
3.1. Response to Zn(II) of the dH3w Variants
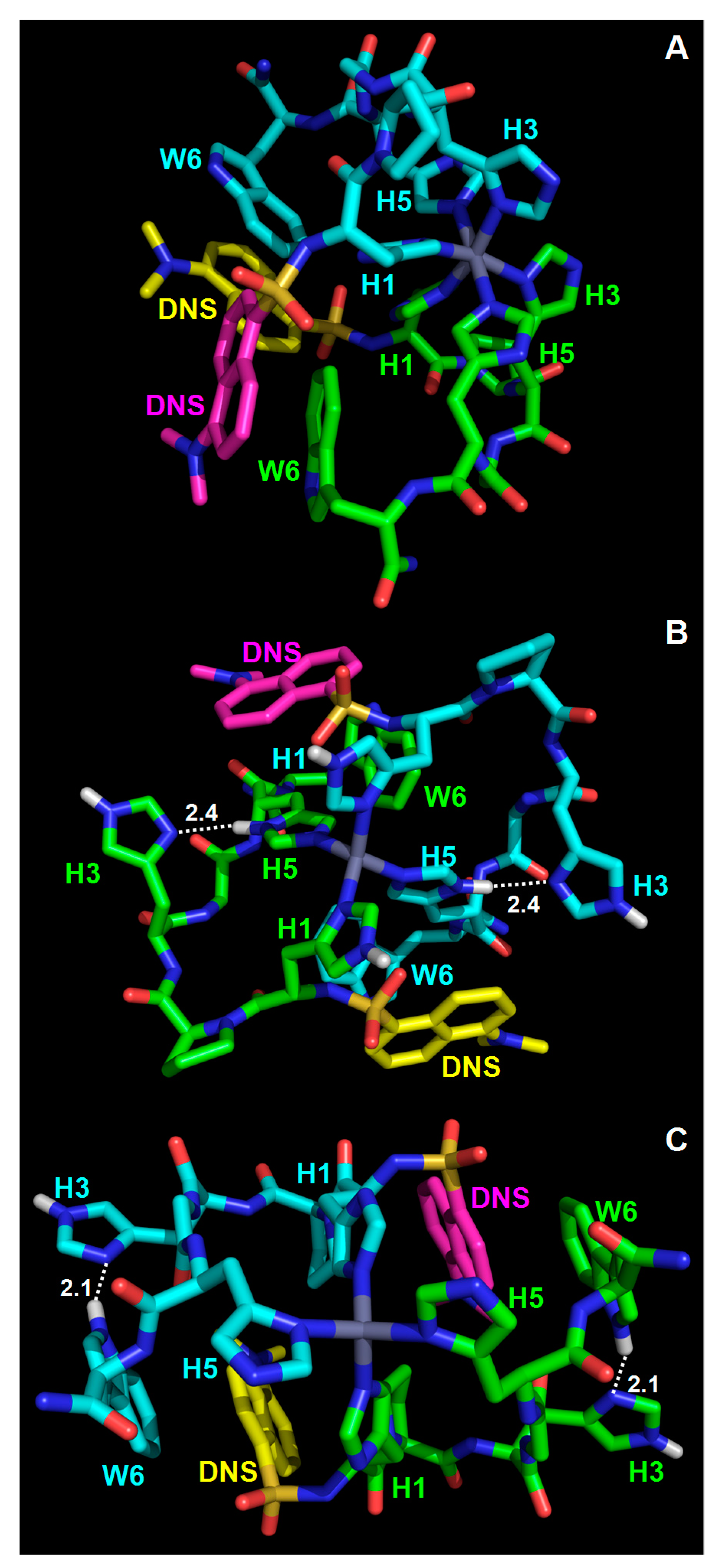
3.2. Modeling of the dH3W/Zn(II) Complex
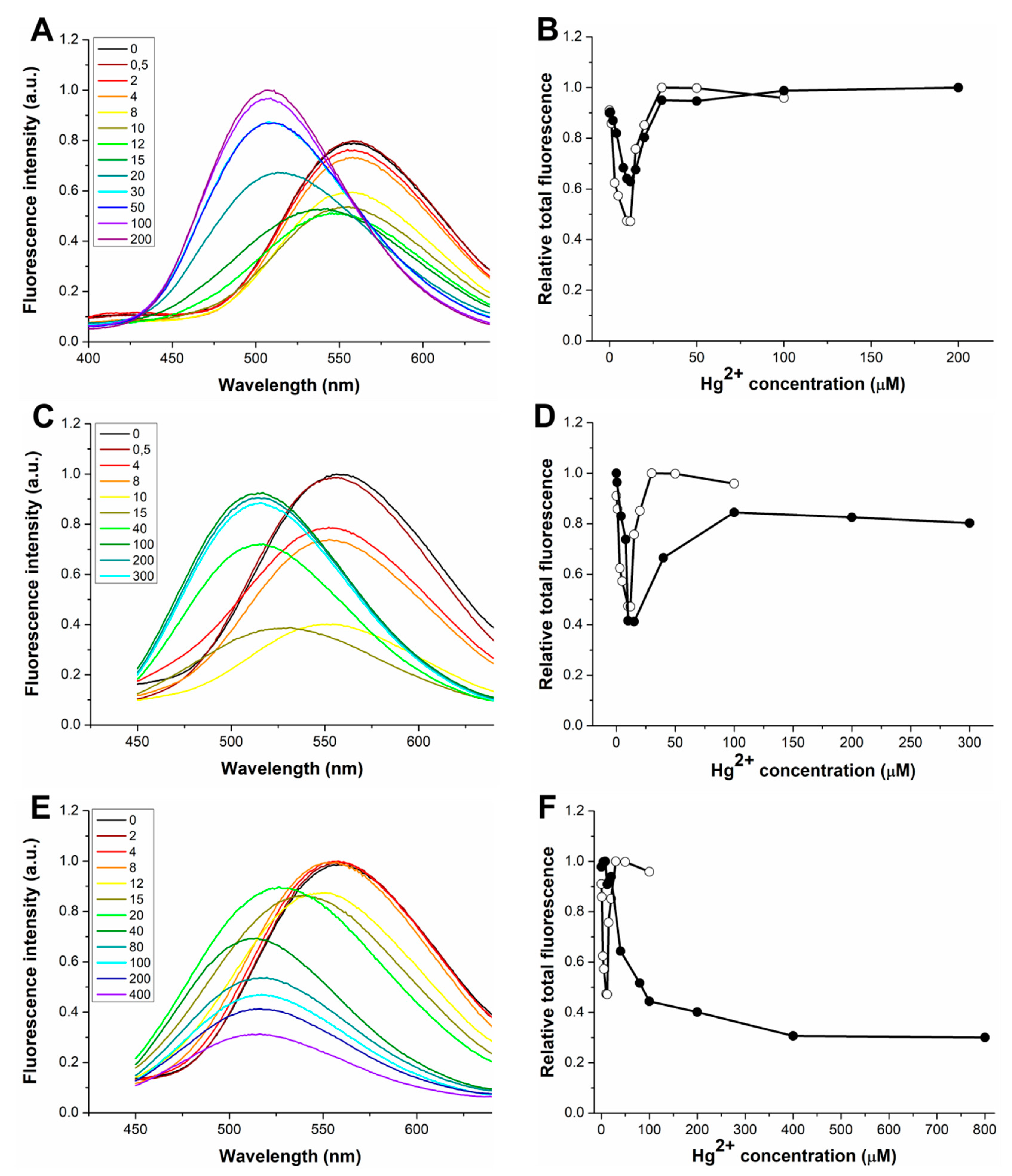
3.3. Response to Hg(II) of the dH3w Variants
3.4. Modeling of the Hg/dH3w Complexes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- DeSilva, T.M.; Veglia, G.; Porcelli, F.; Prantner, A.M.; Opella, S.J. Selectivity in heavy metal-binding to peptides and proteins. Biopolymers 2002, 64, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Jaishankar, M.; Tseten, T.; Anbalagan, N.; Mathew, B.B.; Beeregowda, K.N. Toxicity, mechanism and health effects of some heavy metals. Interdiscip. Toxicol. 2014, 7, 60–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egan, J.G.; Hynes, A.J.; Fruehwald, H.M.; Ebralidze, I.I.; King, S.D.; Alipour Moghadam Esfahani, R.; Naumkin, F.Y.; Easton, E.B.; Zenkina, O.V. A novel material for the detection and removal of mercury(ii) based on a 2,6-bis(2-thienyl)pyridine receptor. J. Mater. Chem. C 2019, 7, 10187–10195. [Google Scholar] [CrossRef]
- Pirzada, M.; Altintas, Z. Nanomaterials for Healthcare Biosensing Applications. Sensors 2019, 19, 5311. [Google Scholar] [CrossRef] [Green Version]
- Shellaiah, M.; Simon, T.; Venkatesan, P.; Sun, K.W.; Ko, F.H.; Wu, S.P. Cysteamine-modified diamond nanoparticles applied in cellular imaging and Hg2+ ions detection. Appl. Surf. Sci. 2019, 465, 340–350. [Google Scholar] [CrossRef]
- Singh, A.K.; Bhattacharjee, G.; Singh, R. Mercury(II)-selective membrane electrode using tetrathia- diazacyclotetradeca-2,9-diene as neutral carrier. Sensors Actuators B Chem. 2004, 99, 36–41. [Google Scholar] [CrossRef]
- Donadio, G.; Di Martino, R.; Oliva, R.; Petraccone, L.; Del Vecchio, P.; Di Luccia, B.; Ricca, E.; Isticato, R.; Di Donato, A.; Notomista, E. A new peptide-based fluorescent probe selective for zinc(II) and copper(II). J. Mater. Chem. B 2016, 4, 6979–6988. [Google Scholar] [CrossRef]
- Siepi, M.; Oliva, R.; Petraccone, L.; Del Vecchio, P.; Ricca, E.; Isticato, R.; Lanzilli, M.; Maglio, O.; Lombardi, A.; Leone, L.; et al. Fluorescent peptide dH3w: A sensor for environmental monitoring of mercury (II). PLoS ONE 2018, 13, e0204164. [Google Scholar] [CrossRef]
- Graciani, F.S.; Ximenes, V.F. Investigation of human albumin-induced circular dichroism in dansylglycine. PLoS ONE 2013, 8, e76849. [Google Scholar] [CrossRef]
- Joshi, B.P.; Lohani, C.R.; Lee, K.H. A highly sensitive and selective detection of Hg(II) in 100% aqueous solution with fluorescent labeled dimerized Cys residues. Org. Biomol. Chem. 2010, 8, 3220–3226. [Google Scholar] [CrossRef]
- Siepi, M.; Donadio, G.; Dardano, P.; De Stefano, L.; Monti, D.M.; Notomista, E. Denatured lysozyme-coated carbon nanotubes: A versatile biohybrid material. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Siepi, M.; Morales-Narváez, E.; Domingo, N.; Monti, D.M.; Notomista, E.; Merkoçi, A. Production of biofunctionalized MoS2 flakes with rationally modified lysozyme: A biocompatible 2D hybrid material. 2D Mater. 2017, 4, 035007. [Google Scholar] [CrossRef] [Green Version]
- Donadio, G.; Sarcinelli, C.; Pizzo, E.; Notomista, E.; Pezzella, A.; Di Cristo, C.; De Lise, F.; Di Donato, A.; Izzo, V. The toluene o-xylene monooxygenase enzymatic activity for the biosynthesis of aromatic antioxidants. PLoS ONE 2015, 10, e0124427. [Google Scholar] [CrossRef] [PubMed]
- De Rosa, M.; Zanfardino, A.; Notomista, E.; Wichelhaus, T.A.; Saturnino, C.; Varcamonti, M.; Soriente, A. Novel promising linezolid analogues: Rational design, synthesis and biological evaluation. Eur. J. Med. Chem. 2013, 69, 779–785. [Google Scholar] [CrossRef]
- Notomista, E.; Cafaro, V.; Bozza, G.; Donato, A. Di Molecular determinants of the regioselectivity of toluene/o-xylene monooxygenase from pseudomonas sp. strain OX1. Appl. Environ. Microbiol. 2009, 75, 823–836. [Google Scholar] [CrossRef] [Green Version]
- Notomista, E.; Scognamiglio, R.; Troncone, L.; Donadio, G.; Pezzella, A.; Di Donato, A.; Izzo, V. Tuning the specificity of the recombinant multicomponent toluene o-xylene monooxygenase from Pseudomonas sp. Strain OX1 for the biosynthesis of tyrosol from 2-phenylethanol. Appl. Environ. Microbiol. 2011, 77, 5428–5437. [Google Scholar] [CrossRef] [Green Version]
- Zanfardino, A.; Restaino, O.F.; Notomista, E.; Cimini, D.; Schiraldi, C.; De Rosa, M.; De Felice, M.; Varcamonti, M. Isolation of an Escherichia coli K4 kfoC mutant over-producing capsular chondroitin. Microb. Cell Fact. 2010, 9, 34. [Google Scholar] [CrossRef] [Green Version]
- Zhorov, B.S.; Bregestovski, P.D. Chloride channels of glycine and GABA receptors with blockers: Monte Carlo minimization and structure-activity relationships. Biophys. J. 2000, 78, 1786–1803. [Google Scholar] [CrossRef] [Green Version]
- Weiner, S.J.; Kollman, P.A.; Case, D.A.; Singh, U.C.; Ghio, C.; Alagona, G.; Profeta, S.; Weiner, P. A new force field for molecular mechanical simulation of nucleic acids and proteins. J. Am. Chem. Soc. 1984, 106, 765–784. [Google Scholar] [CrossRef]
- Ghisaidoobe, A.; Chung, S. Intrinsic Tryptophan Fluorescence in the Detection and Analysis of Proteins: A Focus on Förster Resonance Energy Transfer Techniques. Int. J. Mol. Sci. 2014, 15, 22518–22538. [Google Scholar] [CrossRef]
- Chen, Y.; Barkley, M.D. Toward Understanding Tryptophan Fluorescence in Proteins. Biochemistry 1998, 37, 9976–9982. [Google Scholar] [CrossRef] [PubMed]
- Garrett, T.P.J.; Guss, J.M.; Freeman, H.C. Hexakis(imidazole)manganese(II) dichloride tetrahydrate, [Mn(C3H4N2)6]Cl2.4H2O, and hexakis(imidazole)zinc(II) dichloride tetrahydrate, [Zn(C3H4N2)6]Cl2.4H2O. Acta Crystallogr. Sect. C Cryst. Struct. Commun. 1983, 39, 1027–1031. [Google Scholar] [CrossRef]
- Klitzke, J.S.; Roisnel, T.; Carpentier, J.F.; Casagrande, O.L. Zinc(II) complexes based on sterically hindered hydrotris(pyrazolyl)borate ligands: Synthesis, reactivity and solid-state structures. Inorganica Chim. Acta 2009, 362, 4585–4592. [Google Scholar] [CrossRef]
- Zvargulis, E.S.; Buys, I.E.; Hambley, T.W. Models of the active sites of zinc containing enzymes: The crystal structures of two bis(tripod)zinc(II) complexes. Polyhedron 1995, 14, 2267–2273. [Google Scholar] [CrossRef]
- Lazaridis, T.; Karplus, M. Effective energy function for proteins in solution. Proteins Struct. Funct. Genet. 1999, 35, 133–152. [Google Scholar] [CrossRef]
- Brooks, P.; Davidson, N. Mercury(II) Complexes of Imidazole and Histidine. J. Am. Chem. Soc. 1960, 82, 2118–2123. [Google Scholar] [CrossRef]
- Sénèque, O.; Rousselot-Pailley, P.; Pujol, A.; Boturyn, D.; Crouzy, S.; Proux, O.; Manceau, A.; Lebrun, C.; Delangle, P. Mercury Trithiolate Binding (HgS3) to a de Novo Designed Cyclic Decapeptide with Three Preoriented Cysteine Side Chains. Inorg. Chem. 2018, 57, 2705–2713. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| λmax (nm) | |||
|---|---|---|---|
| Peptide | Unbound | Zn(II) Complex | Hg(II) Complex |
| dH3w | 560 | 515 | 507 |
| dH3w(H1A) | 560 | 534 | 507 |
| dH3w(H3A) | 560 | 528 | 515 |
| dH3w(H5A) | 560 | 500 | 515 |
| dH3w(W6A) | 560 | 524 | 510 |
| AcH3w | 359 | 359 | 350 |
| Peptide | Kb (M−1) | n (peptide/zinc) |
|---|---|---|
| dH3w | (5.9 ± 2.2) × 105 | 0.5 a |
| dH3w(H3A) | (2.6 ± 0.2) × 104 | 0.5 b |
| dH3w(W6A) | (4.6 ± 0.2) × 105 | 0.5 a |
| AcH3w | (2.9 ± 0.1) × 105 | 0.5 b |
| Protonation State of His1, His3, and His5 a | ||||||||
|---|---|---|---|---|---|---|---|---|
| Octahedral complexes | Nδ/Nδ/Nδ | Nδ/Nε/Nδ | Nδ/Nδ/Nε | Nδ/Nε/Nε | Nε/Nδ/Nδ | Nε/Nε/Nδ | Nε/Nδ/Nε | Nε/Nε/Nε |
| Energy (kJ/mol) | −1205.8 | −1295.4 | −1315.4 | −1338.0 | −1297.5 | −1324.2 | −1399.1 | −1377.4 |
| Tetrahedral complexes | Nδ/Nδ/Nδ | Nδ/Nε/Nδ | Nδ/Nδ/Nε | Nδ/Nε/Nε | Nε/Nδ/Nδ | Nε/Nε/Nδ | Nε/Nδ/Nε | Nε/Nε/Nε |
| Energy (kJ/mol) | −1486.6 | −1478.2 | −1511.7 | −1510.0 | −1528.0 | −1526.7 | −1513.4 | −1512.9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siepi, M.; Oliva, R.; Battista, F.; Petraccone, L.; Del Vecchio, P.; Izzo, V.; Dal Piaz, F.; Isticato, R.; Notomista, E.; Donadio, G. Molecular Dissection of dH3w, A Fluorescent Peptidyl Sensor for Zinc and Mercury. Sensors 2020, 20, 598. https://doi.org/10.3390/s20030598
Siepi M, Oliva R, Battista F, Petraccone L, Del Vecchio P, Izzo V, Dal Piaz F, Isticato R, Notomista E, Donadio G. Molecular Dissection of dH3w, A Fluorescent Peptidyl Sensor for Zinc and Mercury. Sensors. 2020; 20(3):598. https://doi.org/10.3390/s20030598
Chicago/Turabian StyleSiepi, Marialuisa, Rosario Oliva, Filomena Battista, Luigi Petraccone, Pompea Del Vecchio, Viviana Izzo, Fabrizio Dal Piaz, Rachele Isticato, Eugenio Notomista, and Giuliana Donadio. 2020. "Molecular Dissection of dH3w, A Fluorescent Peptidyl Sensor for Zinc and Mercury" Sensors 20, no. 3: 598. https://doi.org/10.3390/s20030598
APA StyleSiepi, M., Oliva, R., Battista, F., Petraccone, L., Del Vecchio, P., Izzo, V., Dal Piaz, F., Isticato, R., Notomista, E., & Donadio, G. (2020). Molecular Dissection of dH3w, A Fluorescent Peptidyl Sensor for Zinc and Mercury. Sensors, 20(3), 598. https://doi.org/10.3390/s20030598