
Anti-Inflammatory Potential of Daturaolone from Datura innoxia Mill.: In Silico, In Vitro and In Vivo Studies

 , ,
, ,  ,
, 
Abstract
:1. Introduction
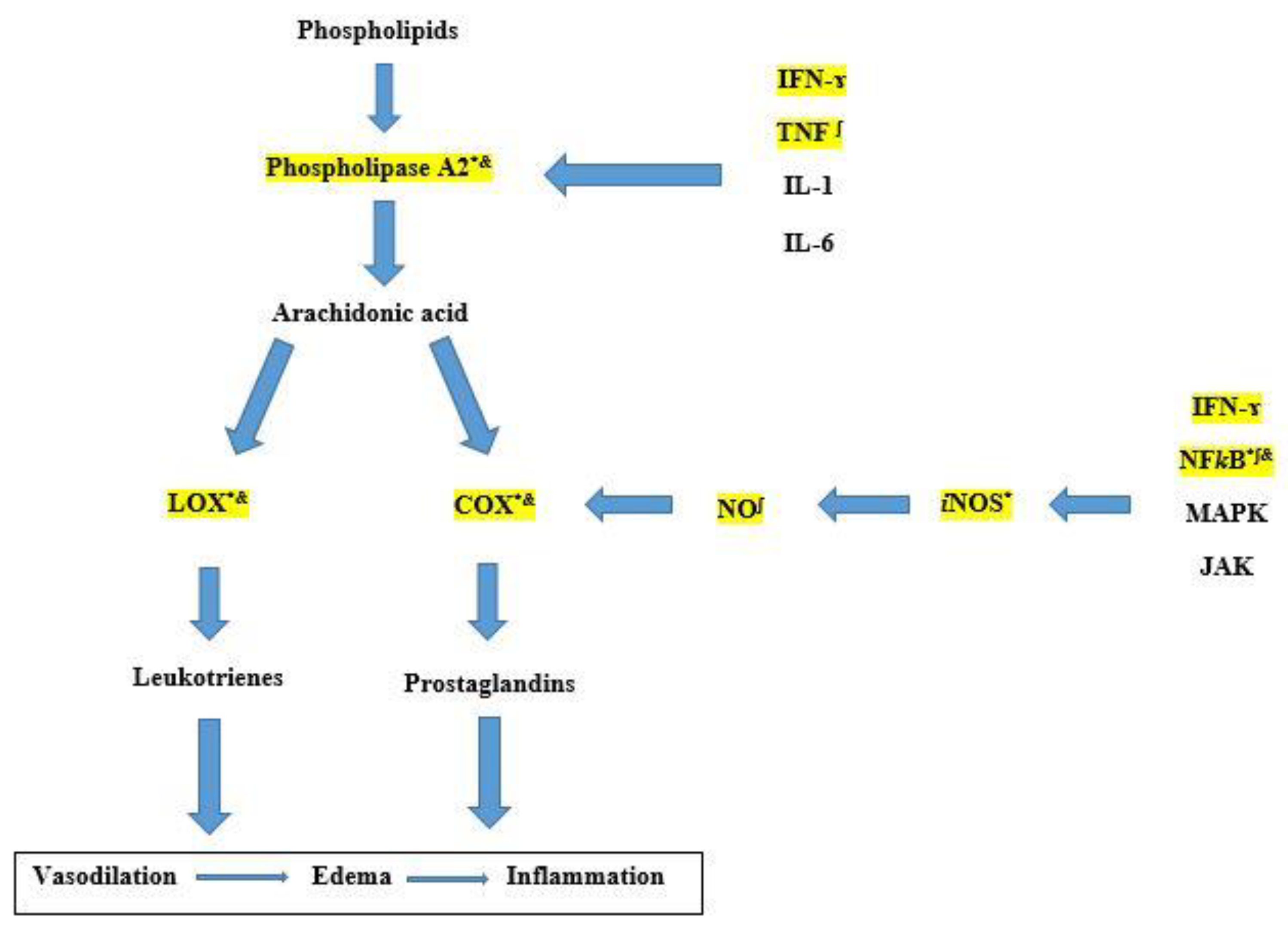
2. Results and Discussion
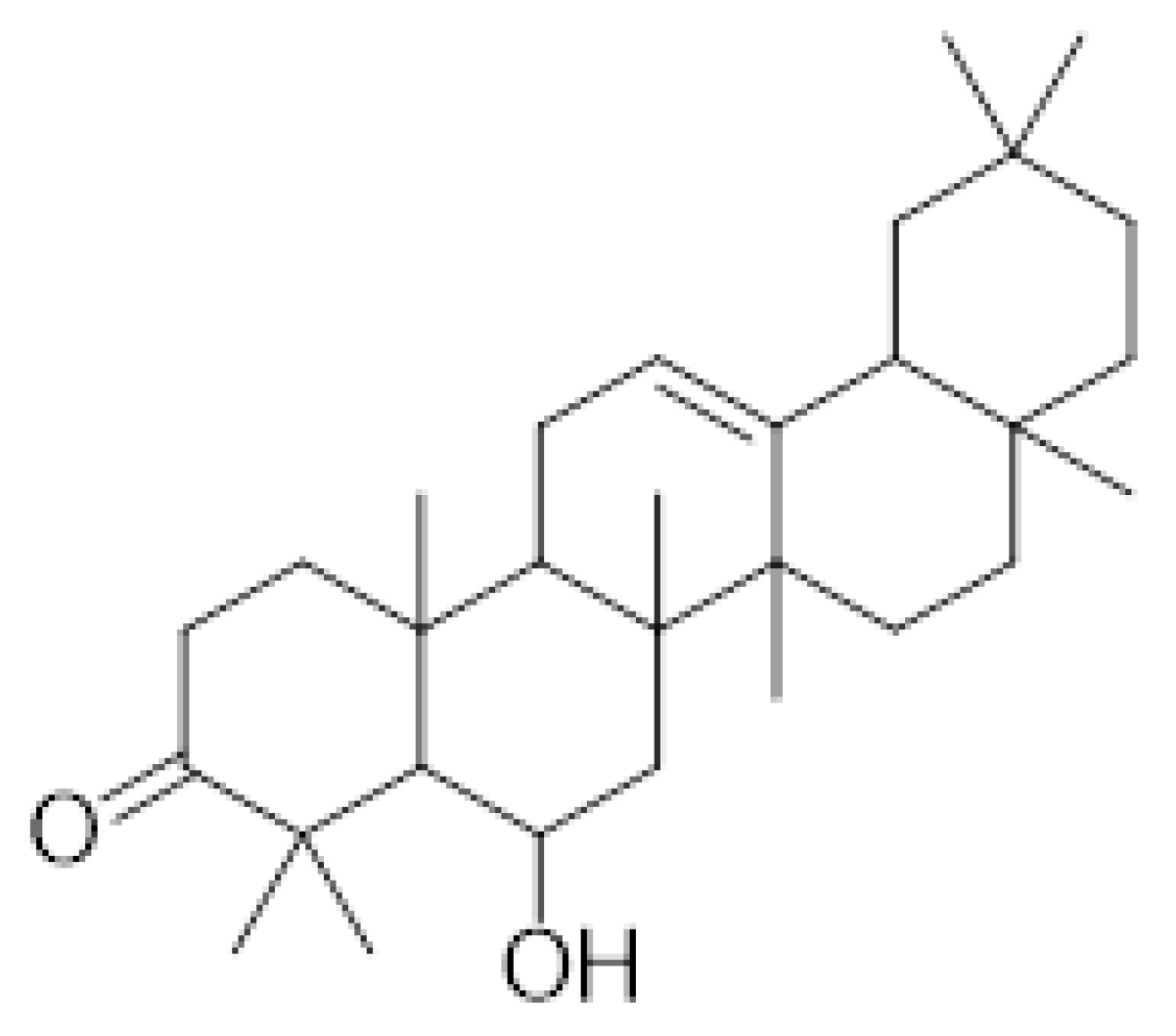
2.1. Daturaolone Was Predicted as a Drug-like Compound
2.2. Absorption, Distribution, Metabolism, Excretion and Toxicity (ADMET) of Daturaolone
2.3. Daturaolone Can Interact with Multiple Molecular Targets
2.4. Predicted Cytotoxicity in Cancer Cell Lines
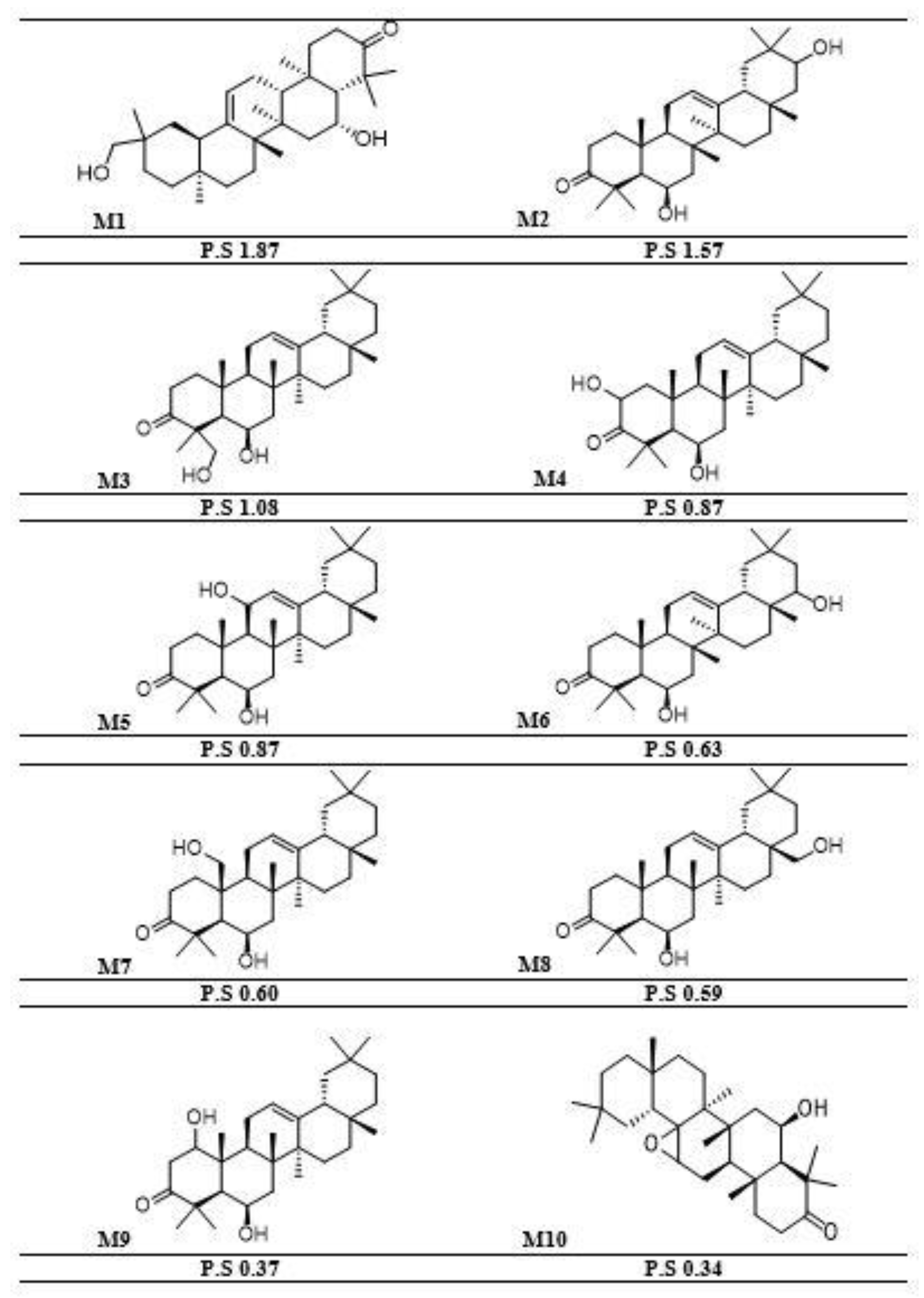
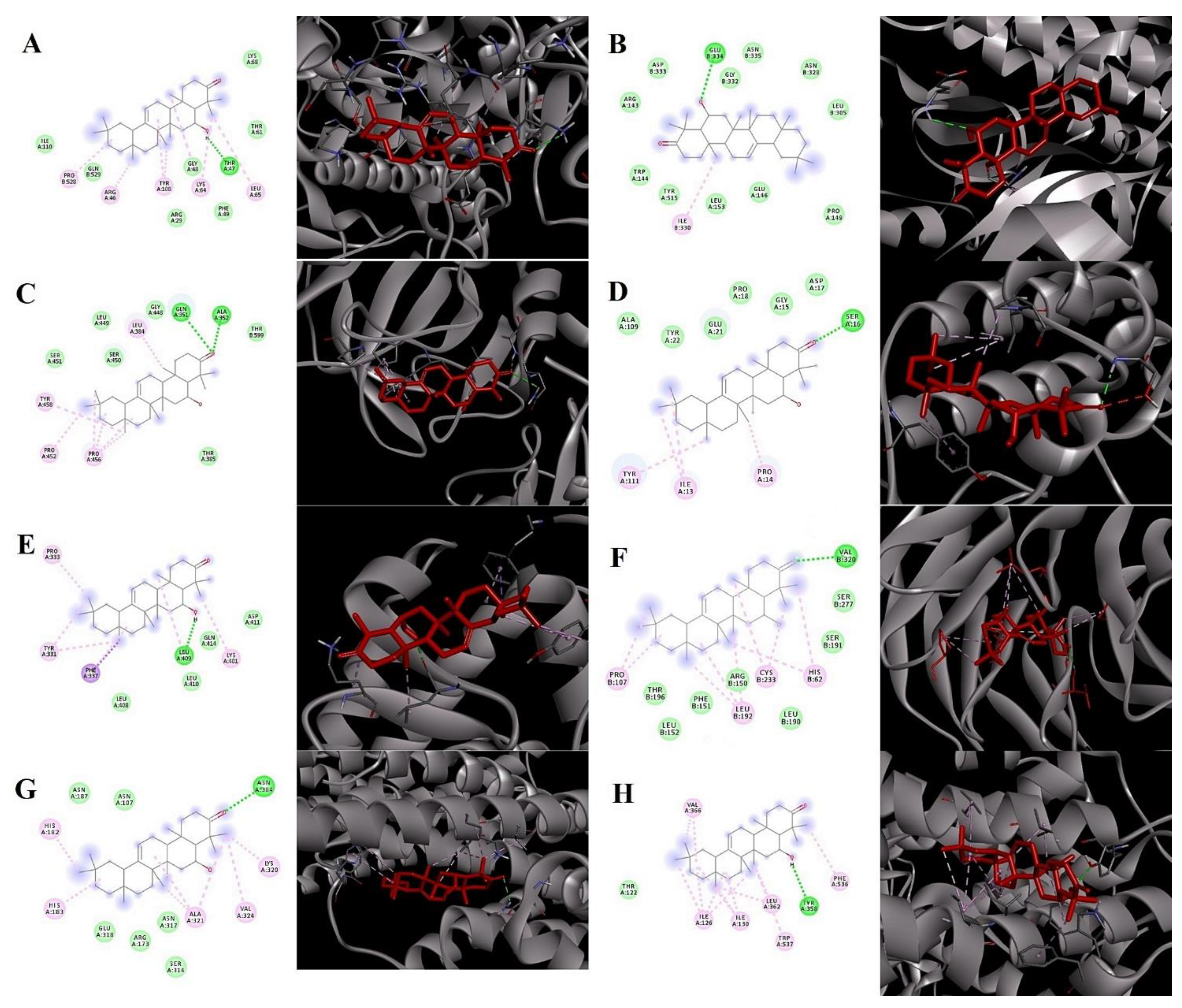
2.5. Molecular Docking Analysis
2.6. In Vitro Cytotoxicity of Daturaolone
2.7. PK, TNF-α Activated NF-κB and NO Production Inhibition Properties of Daturaolone
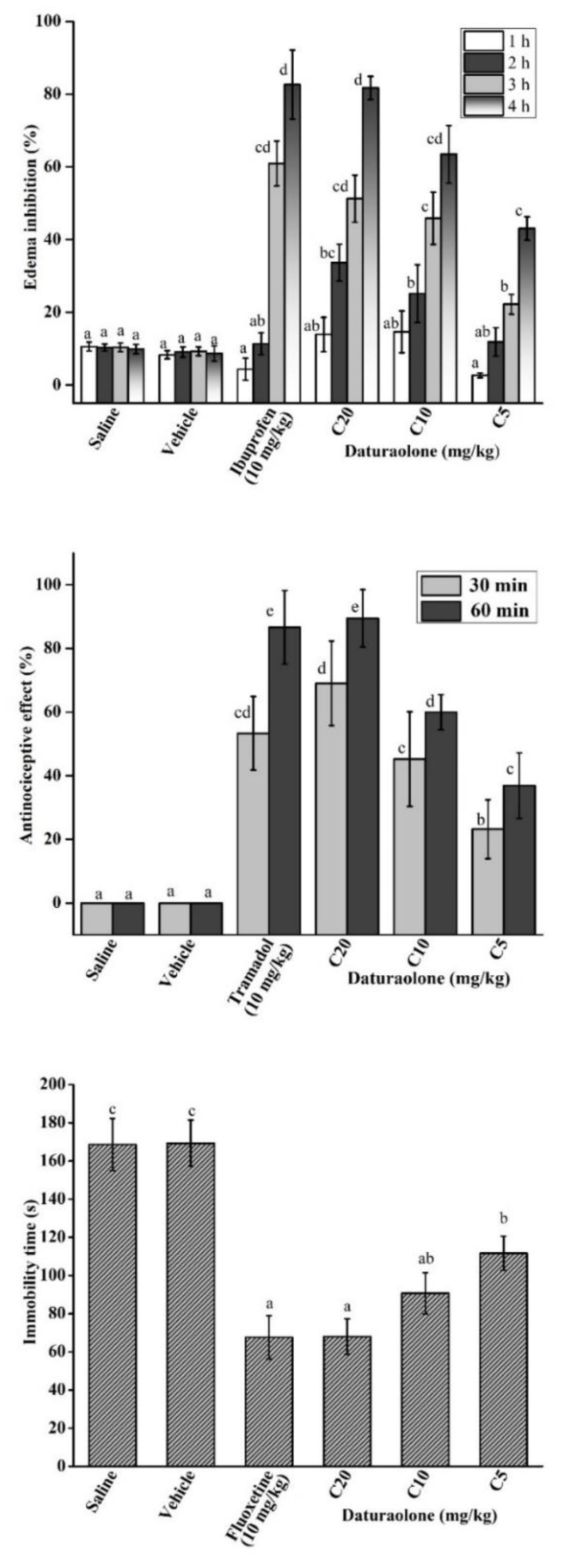
2.8. Daturaolone Reduced Inflammation In Vivo
2.9. Daturaolone Diminished Thermal Pain
2.10. Daturaolone Improved Depression
3. Materials and Methods
3.1. Chemicals and Cell Lines
3.2. Isolation of Daturaolone
3.3. Animals
3.4. In Silico Screening
3.4.1. Drug Likeliness Prediction
3.4.2. Absorption, Distribution, Metabolism, Excretion and Toxicity (ADMET) Profile Prediction
3.4.3. Molecular Target and Cancer Cell Line Cytotoxicity Prediction
3.4.4. Molecular Docking Analysis
3.5. In Vitro Studies
3.5.1. Cytotoxic Activity
3.5.2. Protein Kinase (PK) Inhibition Assay
3.5.3. Inhibition of TNF-α-Activated Nuclear Factor-Kappa B (NF-κB)
3.5.4. Inhibition of NO Production
3.6. In Vivo Studies
3.6.1. Dose Selection and Treatment Groups
3.6.2. Carrageenan-Induced Hind Paw Edema Inflammatory Assay
3.6.3. Hot Plate Antinociceptive Assay
3.6.4. Tail Suspension Antidepressant Assay
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2017, 9, 7204–7218. [Google Scholar] [CrossRef] [Green Version]
- Zelova, H.; Hosek, J. TNF-alpha signalling and inflammation: Interactions between old acquaintances. Inflamm. Res. Off. J. Eur. Histamine Res. Society 2013, 62, 641–651. [Google Scholar] [CrossRef]
- Thoppil, R.J.; Bishayee, A. Terpenoids as potential chemopreventive and therapeutic agents in liver cancer. World J. Hepatol. 2011, 3, 228–249. [Google Scholar] [CrossRef]
- Kocor, M.; Pyrek, J.S.; Atal, C.; Bedi, K.; Sharma, B. Triterpenes of Datura innoxia. Structure of daturadiol and daturaolone. J. Org. Chem. 1973, 38, 3685–3688. [Google Scholar] [CrossRef] [PubMed]
- Rauf, A.; Maione, F.; Uddin, G.; Raza, M.; Siddiqui, B.S.; Muhammad, N.; Shah, S.U.A.; Khan, H.; De Feo, V.; Mascolo, N. Biological evaluation and docking analysis of daturaolone as potential cyclooxygenase inhibitor. Evid.-Based Complementary Altern. Med. 2016, 2016, 4098686. [Google Scholar] [CrossRef] [Green Version]
- Bawazeer, S.; Rauf, A.; Bawazeer, S. Gastrointestinal motility, muscle relaxation, antipyretic and acute toxicity screening of amyrin type triterpenoid (daturaolone) isolated from Datura metel Linnaeus (Angel’s trumpet) fruits. Front. Pharmacol. 2020, 11, 1473. [Google Scholar] [CrossRef] [PubMed]
- Bawazeer, S.; Rauf, A.; Bawazeer, S. Potent In Vitro α-Glucosidase and β-Secretase Inhibition of Amyrin-Type Triterpenoid Isolated from Datura metel Linnaeus (Angel’s Trumpet) Fruits. BioMed Res. Int. 2020, 2020, 8530165. [Google Scholar] [CrossRef]
- Bawazeer, S.; Rauf, A. In Vitro Antibacterial and Antifungal Potential of Amyrin-Type Triterpenoid Isolated from Datura metel Linnaeus. BioMed Res. Int. 2021, 2021, 1543574. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, P. The role of P-glycoprotein in the blood-brain barrier. Einstein. QJ Biol. Med. 2003, 19, 160–165. [Google Scholar]
- Wang, Z.; Sun, W.; Huang, C.-K.; Wang, L.; Ia, M.-M.; Cui, X.; Hu, G.-X.; Wang, Z.-S. Inhibitory effects of curcumin on activity of cytochrome P450 2C9 enzyme in human and 2C11 in rat liver microsomes. Drug Dev. Ind. Pharm. 2015, 41, 613–616. [Google Scholar] [CrossRef]
- Aronov, A.M. Predictive in silico modeling for hERG channel blockers. Drug Discov. Today 2005, 10, 149–155. [Google Scholar] [CrossRef]
- Issa, T.N.; Wathieu, H.; Ojo, A.; Byers, W.S.; Dakshanamurthy, S. Drug metabolism in preclinical drug development: A survey of the discovery process, toxicology, and computational tools. Curr. Drug Metab. 2017, 18, 556–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attiq, A.; Jalil, J.; Husain, K.; Ahmad, W. Raging the war against inflammation with natural products. Front. Pharmacol. 2018, 9, 976. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.J.; Reynell, C. How do antidepressants influence the BOLD signal in the developing brain? Dev. Cogn. Neurosci. 2017, 25, 45–57. [Google Scholar] [CrossRef]
- Hong, J.; Sang, S.; Park, H.-J.; Kwon, S.J.; Suh, N.; Huang, M.-T.; Ho, C.-T.; Yang, C.S. Modulation of arachidonic acid metabolism and nitric oxide synthesis by garcinol and its derivatives. Carcinogenesis 2006, 27, 278–286. [Google Scholar] [CrossRef]
- Vučković, S.; Srebro, D.; Vujović, K.S.; Vučetić, Č.; Prostran, M. Cannabinoids and pain: New insights from old molecules. Front. Pharmacol. 2018, 9, 1259. [Google Scholar] [CrossRef] [Green Version]
- Brahmkshatriya, P.P.; Brahmkshatriya, P.S. Terpenes: Chemistry, Biological Role, and Therapeutic Applications. In Natural Products: Phytochemistry, Botany and Metabolism of Alkaloids, Phenolics and Terpenes; Ramawat, K.G., Mérillon, J.-M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 2665–2691. [Google Scholar] [CrossRef]
- Lagunin, A.A.; Dubovskaja, V.I.; Rudik, A.V.; Pogodin, P.V.; Druzhilovskiy, D.S.; Gloriozova, T.A.; Filimonov, D.A.; Sastry, N.G.; Poroikov, V.V. CLC-Pred: A freely available web-service for in silico prediction of human cell line cytotoxicity for drug-like compounds. PLoS ONE 2018, 13, e0191838. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Zhao, J.; Chen, Z. Insights into the molecular mechanisms of protein-ligand interactions by molecular docking and molecular dynamics simulation: A case of oligopeptide binding protein. Comput. Math. Methods Med. 2018, 2018, 3502514. [Google Scholar] [CrossRef]
- Lin, H.-C.; Lin, T.-H.; Wu, M.-Y.; Chiu, Y.-C.; Tang, C.-H.; Hour, M.-J.; Liou, H.-C.; Tu, H.-J.; Yang, R.-S.; Fu, W.-M. 5-Lipoxygenase inhibitors attenuate TNF-α-induced inflammation in human synovial fibroblasts. PLoS ONE 2014, 9, e107890. [Google Scholar] [CrossRef]
- Karin, M. NF-κB as a critical link between inflammation and cancer. Cold Spring Harb. Perspect. Biol. 2009, 1, a000141. [Google Scholar] [CrossRef]
- Caviedes, A.; Lafourcade, C.; Soto, C.; Wyneken, U. BDNF/NF-κB signaling in the neurobiology of depression. Curr. Pharm. Des. 2017, 23, 3154–3163. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Shishodia, S. Molecular targets of dietary agents for prevention and therapy of cancer. Biochem. Pharmacol. 2006, 71, 1397–1421. [Google Scholar] [CrossRef] [PubMed]
- Yao, G.; Sebisubi, F.M.; Voo, L.Y.C.; Ho, C.C.; Tan, G.T.; Chang, L.C. Citrinin Derivatives from the Soil Filamentous Fungus Penicillium sp. H9318. J. Braz. Chem. Soc. 2011, 22, 1125–1129. [Google Scholar] [CrossRef] [Green Version]
- Barbara, W.; Saxena, G.; Wanggui, Y.; Kau, D.; Wrigley, S.; Stokes, R.; Davies, J. Identifying Protein Kinase Inhibitors Using an Assay Based on Inhibition of Aerial Hyphae Formation in Streptomyces. J. Antibiot. 2002, 55, 407–416. [Google Scholar]
- Salminen, A.; Lehtonen, M.; Suuronen, T.; Kaarniranta, K.; Huuskonen, J. Terpenoids: Natural inhibitors of NF-κB signaling with anti-inflammatory and anticancer potential. Cell. Mol. Life Sci. 2008, 65, 2979–2999. [Google Scholar] [CrossRef]
- Orlowski, R.Z.; Baldwin Jr, A.S. NF-κB as a therapeutic target in cancer. Trends Mol. Med. 2002, 8, 385–389. [Google Scholar] [CrossRef] [Green Version]
- Bradley, J. TNF-mediated inflammatory disease. J. Pathol. 2008, 214, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Haq, I.u.; Youn, U.J.; Chai, X.; Park, E.-J.; Kondratyuk, T.P.; Simmons, C.J.; Borris, R.P.; Mirza, B.; Pezzuto, J.M.; Chang, L.C. Biologically Active Withanolides from Withania coagulans. J. Nat. Prod. 2013, 76, 22–28. [Google Scholar] [CrossRef]
- Posadas, I.; Bucci, M.; Roviezzo, F.; Rossi, A.; Parente, L.; Sautebin, L.; Cirino, G. Carrageenan--induced mouse paw oedema is biphasic, age--weight dependent and displays differential nitric oxide cyclooxygenase--2 expression. Br. J. Pharmacol. 2004, 142, 331–338. [Google Scholar] [CrossRef]
- Walker, A.K.; Kavelaars, A.; Heijnen, C.; Dantzer, R. Neuroinflammation and comorbidity of pain and depression. Pharmacol. Rev. 2014, 66, 80–101. [Google Scholar] [CrossRef] [Green Version]
- Niederberger, E.; Geisslinger, G. The IKK--NF--κB pathway: A source for novel molecular drug targets in pain therapy? FASEB J. 2008, 22, 3432–3442. [Google Scholar] [CrossRef] [PubMed]
- Koch, A.; Zacharowski, K.; Boehm, O.; Stevens, M.; Lipfert, P.; Von Giesen, H.; Wolf, A.; Freynhagen, R. Nitric oxide and pro-inflammatory cytokines correlate with pain intensity in chronic pain patients. Inflamm. Res. 2007, 56, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Lotrich, F.E. Inflammatory cytokine-associated depression. Brain Res. 2015, 1617, 113–125. [Google Scholar] [CrossRef] [Green Version]
- Dhir, A.; Kulkarni, S. Nitric oxide and major depression. Nitric Oxide 2011, 24, 125–131. [Google Scholar] [CrossRef]
- Waseem, D.; Butt, A.F.; Haq, I.-u.; Bhatti, M.H.; Khan, G.M. Carboxylate derivatives of tributyltin (IV) complexes as anticancer and antileishmanial agents. DARU J. Pharm. Sci. 2017, 25, 8. [Google Scholar] [CrossRef] [Green Version]
- de Bruyn Kops, C.; Stork, C.; Šícho, M.; Kochev, N.; Svozil, D.; Jeliazkova, N.; Kirchmair, J. GLORY: Generator of the structures of likely cytochrome P450 metabolites based on predicted sites of metabolism. Front. Chem. 2019, 7, 402. [Google Scholar] [CrossRef]
- Ibrahim, M.T.; Uzairu, A.; Shallangwa, G.A.; Ibrahim, A. In-silico studies of some oxadiazoles derivatives as anti-diabetic compounds. J. King Saud Univ.-Sci. 2020, 32, 423–432. [Google Scholar] [CrossRef]
- Kondratyuk, T.P.; Park, E.-J.; Yu, R.; Van Breemen, R.B.; Asolkar, R.N.; Murphy, B.T.; Fenical, W.; Pezzuto, J.M. Novel marine phenazines as potential cancer chemopreventive and anti-inflammatory agents. Mar. Drugs 2012, 10, 451–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, S.; Shin, E.M.; Choi, R.J.; Jung, Y.H.; Kim, J.; Tosun, A.; Kim, Y.S. Suppression of LPS--induced inflammatory and NF--κB responses by anomalin in RAW 264.7 macrophages. J. Cell. Biochem. 2011, 112, 2179–2188. [Google Scholar] [CrossRef]
- Kayani, W.K.; Dilshad, E.; Ahmed, T.; Ismail, H.; Mirza, B. Evaluation of Ajuga bracteosa for antioxidant, anti-inflammatory, analgesic, antidepressant and anticoagulant activities. BMC complementary Altern. Med. 2016, 16, 375. [Google Scholar] [CrossRef] [Green Version]
- Peng, W.-H.; Lo, K.-L.; Lee, Y.-H.; Hung, T.-H.; Lin, Y.-C. Berberine produces antidepressant-like effects in the forced swim test and in the tail suspension test in mice. Life Sci. 2007, 81, 933–938. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drug Likeliness Profile | |||||||||||
| ab Lipinski’s Rule | Scores | ||||||||||
| MW * | MLogP | N or O | NH or OH | b Bioavailability | c Drug Likeness | ||||||
| 440.7 | 5.89 | ≤10 | ≤5 | 0.55 | 0.33 | ||||||
| ADME Profile | |||||||||||
| a PPB * (%) | b BBB * permeability | b P-gp * substrate | b Lipophilicity consensus LogPo/w | b Water Solubility LogS (ESOL) | a GI abs * (%) | a Caco-2 cells permeability (nm/s) | |||||
| 100 | no | no | 6.34 | Low; −7.29 | High: 96.6 | 34.6 | |||||
| Toxicity Profile | |||||||||||
| a Ames Test | a hERG * inhibition risk | a Carcinogenicity | |||||||||
| Pred * | TA100 * | TA1535 * | Mouse | Rat | |||||||
| +S9 | −S9 | +S9 | −S9 | ||||||||
| Non-mutagen | -ive | -ive | -ive | -ive | Low | No | No | ||||
| a Anti-Inflammatory | ||||||
| a Nuclear factor kappa B | b NO synthase | b Arachidonate-5 lipoxygenase | b Cyclooxygenase 2 | b Phospholipase A2 | ||
| a Antinociceptive | ||||||
| b Prostaglandin E synthase | b Peroxisome proliferator-activated receptor-alpha | b 5-HT receptor 1B | c Estrogen receptor antagonist | b Cannabinoid receptor 1 | ||
| a Antidepressant | ||||||
| b 11-betahydroxysteroid dehydrogenase 2 | b Norepinephrine transporter | c Dopamine 1A receptor antagonist | c GABA transporter 1 antagonist | b Serotonin transporter | ||
| d Cancer cell lines cytotoxicity prediction (Pa > Pi) | ||||||
| Leukemia | Pancreatic carcinoma | Thyroid carcinoma | Gastric carcinoma | Hepatoblastoma | Melanoma | |
| HL-60 | PANC-1 | 8505-C | MKN-7 | HepG2 | SK-MEL-1 | |
| 0.53 > 0.02 | 0.48 > 0.00 | 0.47 > 0.00 | 0.43 > 0.00 | 0.42 > 0.03 | 0.48 > 0.03 | |
| In Vitro Cytotoxicity Assessment in Cancer and Normal Cells | |||||||
|---|---|---|---|---|---|---|---|
| Cell lines | C20 | Vincristine | Cabazitaxel | 1% DMSO | |||
| % Inhibition | IC50 (µg/mL) | % Inhibition | IC50 (µg/mL) | % Inhibition | IC50 (µg/mL) | % Inhibition | |
| Huh7.0 | 38.33 ± 1.58 | >20 | 86.75 ± 0.56 | 6.75 ± 0.79 a | -- | ||
| Huh7.5 | 53.60 ± 3.24 | 17.32 b ± 1.43 | 84.57 ± 0.49 | 5.62 ± 0.72 a | -- | ||
| DU145 | 54.70 ± 2.21 | 18.64 b ± 2.15 | 94.75 ± 0.15 | 4.75 ± 1.12 a | -- | ||
| PC3 | 41.53 ± 2.36 | >20 | 92.76 ± 1.76 | 6.62 ± 2.11 a | -- | ||
| MCF7 | 46.86 ± 4.66 | >20 | 78.55 ± 2.36 | 4.33 ± 1.87 a | -- | ||
| Lymphocytes | 29.13 ± 2.57 | >20 | 67.73 ± 1.70 | 6.86 ± 0.49 a | -- | ||
| Sr. No. | Sample | Protein Kinase Inhibition | TNF-α-Activated NF-κB Inhibition | Inhibition of NO Production | ||||
|---|---|---|---|---|---|---|---|---|
| Clear Zone (mm) | Bald Zone (mm) | MIC (µg/disc) | % Inhibition | IC50 (µg/mL) | % Inhibition | IC50 (µg/mL) | ||
| 1 | C20 (µg/disc) | 10.22 ± 1.74 | 12.37 ± 2.41 b | 20 | ||||
| 2 | C20 (µg/mL) | 92.17 ± 5.1 a | 1.2 ± 0.8 a | 84 ± 2.87 a | 4.51 ± 0.92 a | |||
| 3 | Surfactin | 0 | 28.23 ± 2.63 a | - | ||||
| 4 | TPCK | 87.23 ± 4.38 a | 10.7 ± 1.6 a | |||||
| 5 | Curcumin | 91 ± 3.31 a | 2.94 ± 0.74 a | |||||
| 6 | ≥1% DMSO | - | - | - | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baig, M.W.; Fatima, H.; Akhtar, N.; Hussain, H.; Okla, M.K.; Al-Hashimi, A.; Al-Qahtani, W.H.; AbdElgawad, H.; Haq, I.-u. Anti-Inflammatory Potential of Daturaolone from Datura innoxia Mill.: In Silico, In Vitro and In Vivo Studies. Pharmaceuticals 2021, 14, 1248. https://doi.org/10.3390/ph14121248
Baig MW, Fatima H, Akhtar N, Hussain H, Okla MK, Al-Hashimi A, Al-Qahtani WH, AbdElgawad H, Haq I-u. Anti-Inflammatory Potential of Daturaolone from Datura innoxia Mill.: In Silico, In Vitro and In Vivo Studies. Pharmaceuticals. 2021; 14(12):1248. https://doi.org/10.3390/ph14121248
Chicago/Turabian StyleBaig, Muhammad Waleed, Humaira Fatima, Nosheen Akhtar, Hidayat Hussain, Mohammad K. Okla, Abdulrahman Al-Hashimi, Wahidah H. Al-Qahtani, Hamada AbdElgawad, and Ihsan-ul Haq. 2021. "Anti-Inflammatory Potential of Daturaolone from Datura innoxia Mill.: In Silico, In Vitro and In Vivo Studies" Pharmaceuticals 14, no. 12: 1248. https://doi.org/10.3390/ph14121248
APA StyleBaig, M. W., Fatima, H., Akhtar, N., Hussain, H., Okla, M. K., Al-Hashimi, A., Al-Qahtani, W. H., AbdElgawad, H., & Haq, I. -u. (2021). Anti-Inflammatory Potential of Daturaolone from Datura innoxia Mill.: In Silico, In Vitro and In Vivo Studies. Pharmaceuticals, 14(12), 1248. https://doi.org/10.3390/ph14121248