
Pyrvinium Pamoate and Structural Analogs Are Early Macrofilaricide Leads

 and
and
Abstract
:1. Introduction
2. Results
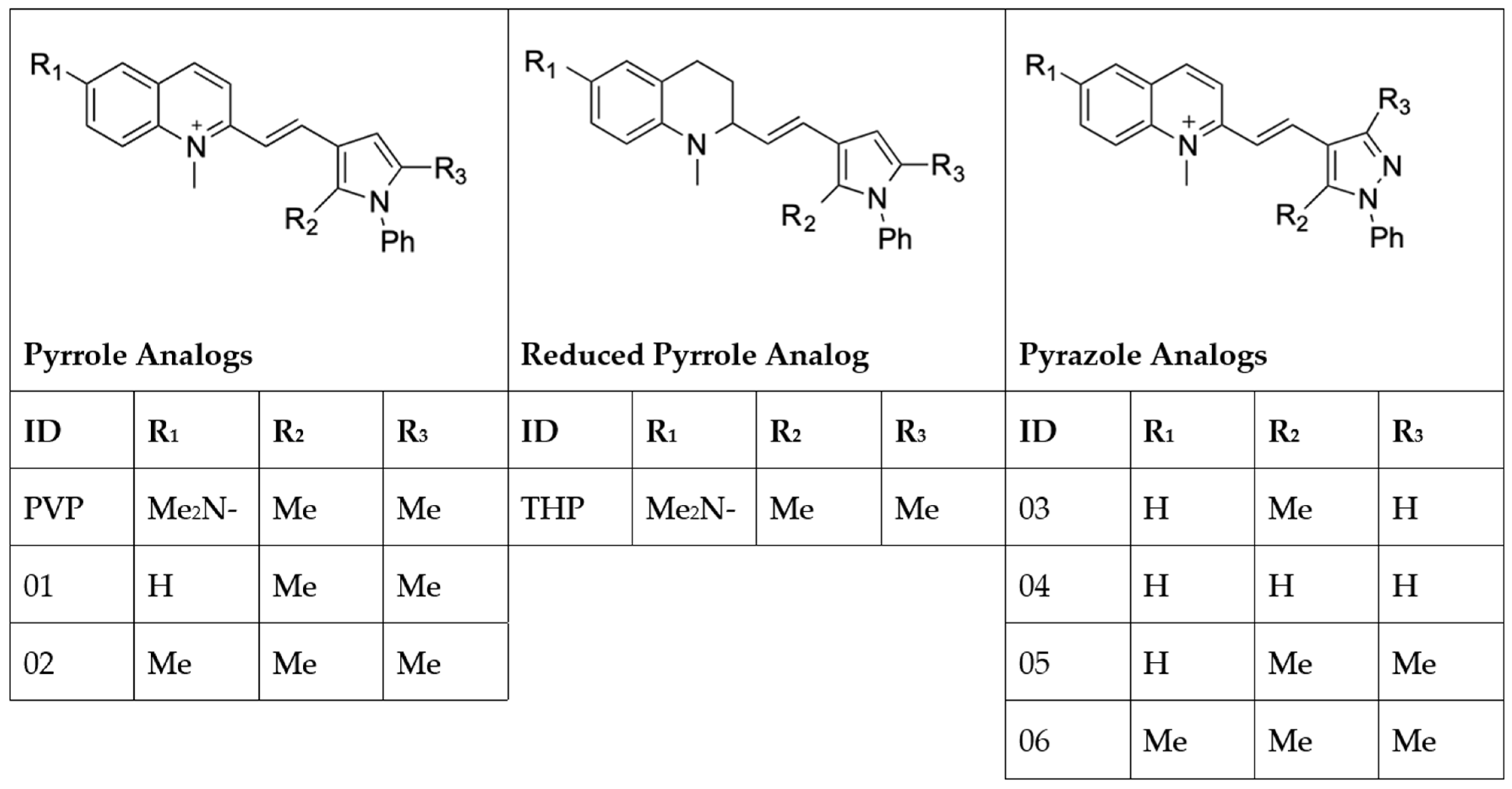
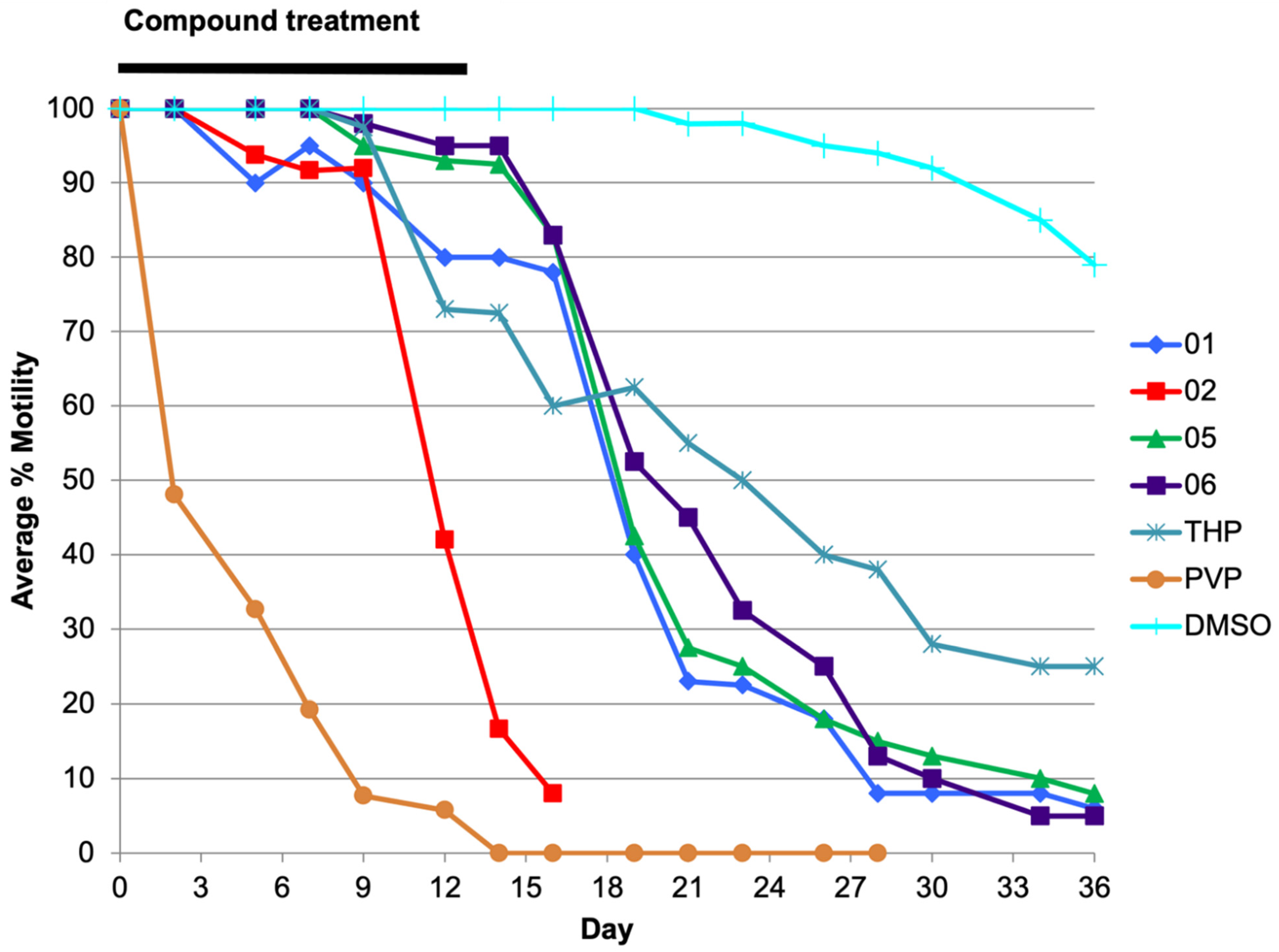
2.1. Analog 06 Shows a Favorable Inhibitory Profile across Parasite Species and Life Stages In Vitro
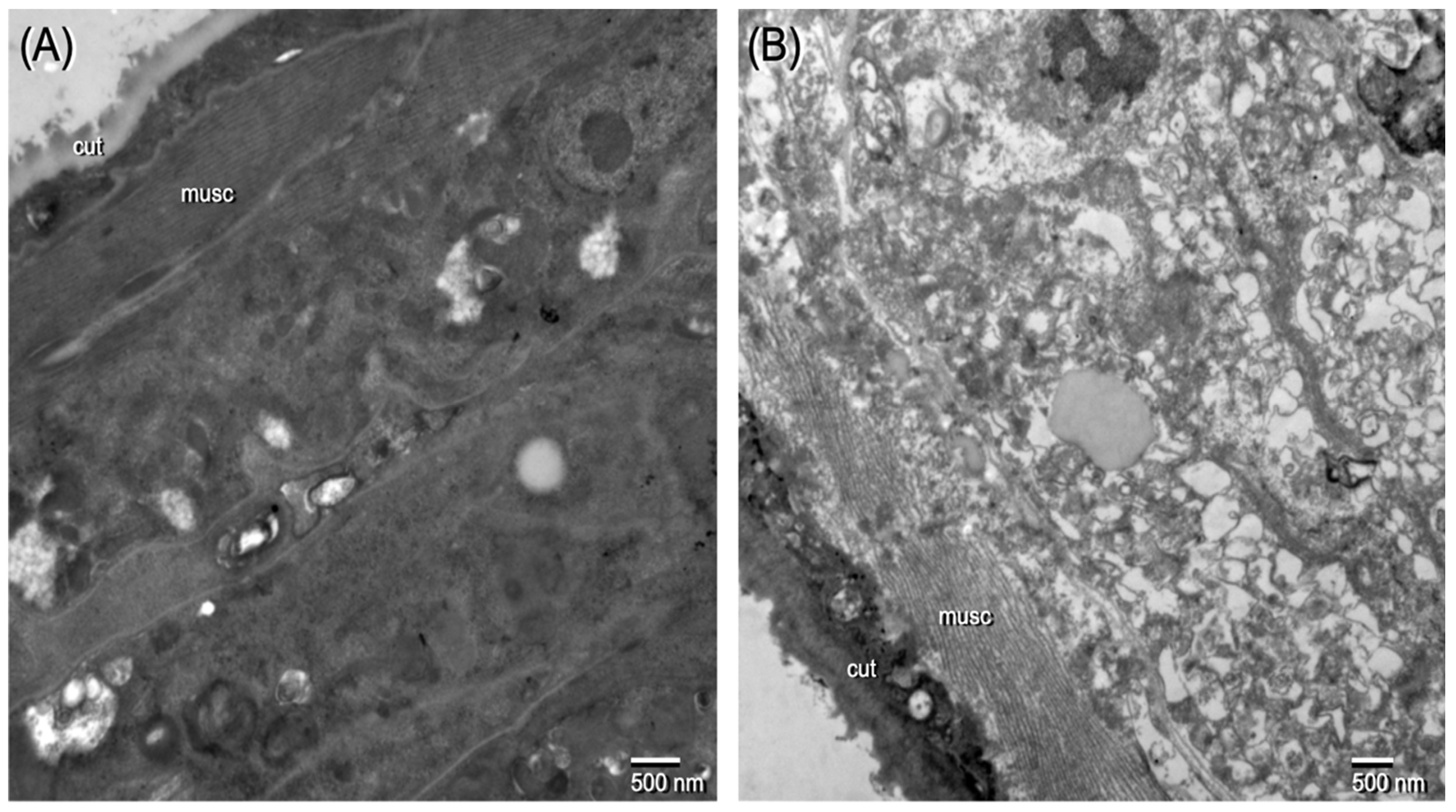
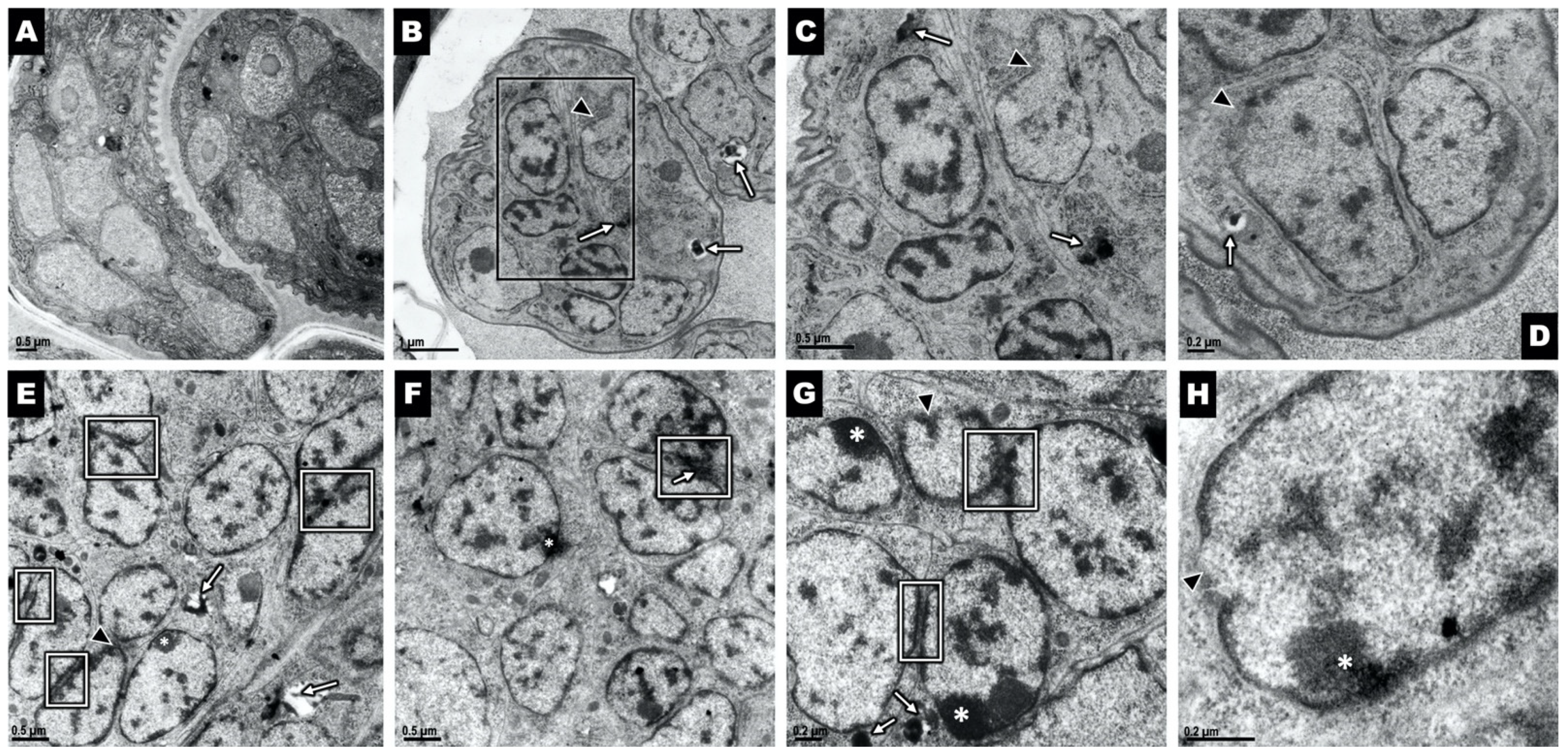
2.2. Ov L5 Treated with 10 µM THP In Vitro Show Ultrastructural Damage
2.3. Analog 06 Demonstrated Acceptable Activity in ADME Studies
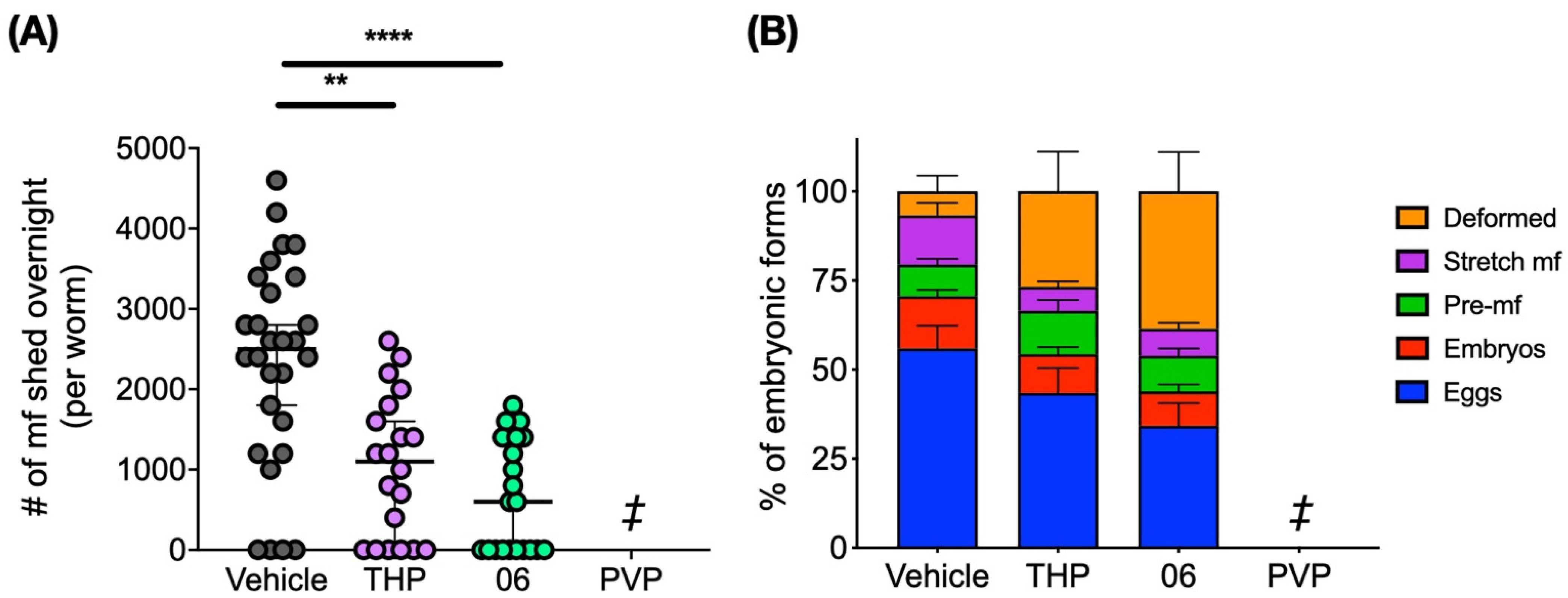
2.4. Pyrvinium, THP and 06 Reduce Female Worm Fecundity in the Brugia/Gerbil Model
2.5. THP and Analog 06 Cause Damage to Female Reproductive Structures
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
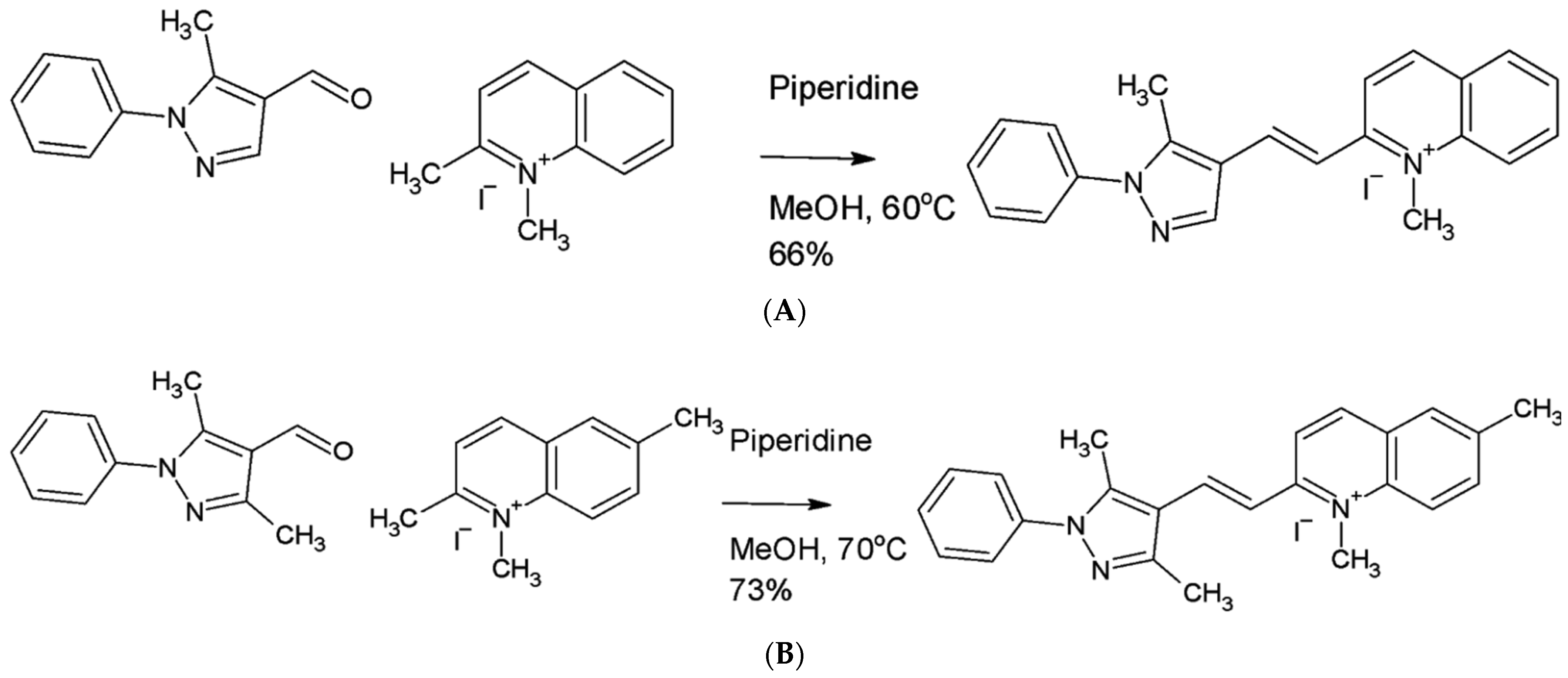
4.2. Synthesis of Pyrvinium Pamoate Analogs
4.3. In Vitro Motility Assays with Adult Female Brugia pahangi
4.4. In Vitro Assays with Adult Onchocerca ochengi and Mf and Loa loa Mf
4.5. Onchocerca volvulus Molting Assay of Third-Stage Larvae (L3) to Fourth-Stage Larvae (L4)
4.6. Onchocerca volvulus Pre-Adult (Ov L5) Assay
4.7. In Vitro Absorption, Distribution, Metabolism and Excretion (ADME)
4.8. In Vivo Studies with the Brugia pahangi/Gerbil Model of Infection
4.9. Ex Vivo Microfilarial Release by Female Brugia pahangi Worms and Embryograms
4.10. qPCR of Wolbachia and Worm DNA in Treated Female Worms
4.11. Statistical Analysis of the In Vivo Study
4.12. Transmission Electron Microscopy of Ov L5 (In Vitro) and Brugia pahangi Adult Females (In Vivo)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- GBD 2017 Disease and Injury Incidence and Prevalence Collaborators. Global, Regional, and National Incidence, Prevalence, and Years Lived with Disability for 354 Diseases and Injuries for 195 Countries and Territories, 1990–2017: A Systematic Analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1789–1858. [Google Scholar] [CrossRef] [Green Version]
- Rajasekaran, S.; Anuradha, R.; Manokaran, G.; Bethunaickan, R. An Overview of Lymphatic Filariasis Lymphedema. Lymphology 2017, 50, 164–182. [Google Scholar]
- WHO. Global Programme to Eliminate Lymphatic Filariasis: Progress Report, 2015. Relev. Epidemiol. Hebd. 2016, 91, 441–455. [Google Scholar]
- Hotez, P.J.; Lo, N.C. 27—Neglected Tropical Diseases: Public Health Control Programs and Mass Drug Administration. In Hunter’s Tropical Medicine and Emerging Infectious Diseases, 10th ed.; Ryan, E.T., Hill, D.R., Solomon, T., Aronson, N.E., Endy, T.P., Eds.; Elsevier: London, UK, 2020; pp. 209–213. [Google Scholar] [CrossRef]
- Gems, D. Longevity and Ageing in Parasitic and Free-Living Nematodes. Biogerontology 2000, 1, 289–307. [Google Scholar] [CrossRef]
- Gardon, J.; Gardon-Wendel, N.; Kamgno, J.; Chippaux, J.P.; Boussinesq, M. Serious Reactions after Mass Treatment of Onchocerciasis with Ivermectin in an Area Endemic for Loa loa Infection. Lancet 1997, 350, 18–22. [Google Scholar] [CrossRef]
- Boussinesq, M.; Gardon, J.; Gardon-Wendel, N.; Chippaux, J.-P. Clinical Picture, Epidemiology and Outcome of Loa-Associated Serious Adverse Events Related to Mass Ivermectin Treatment of Onchocerciasis in Cameroon. Filaria J. 2003, 2, S4. [Google Scholar] [CrossRef] [Green Version]
- Molyneux, D.H.; Hopkins, A.; Bradley, M.H.; Kelly-Hope, L.A. Multidimensional Complexities of Filariasis Control in an Era of Large-Scale Mass Drug Administration Programmes: A Can of Worms. Parasite Vectors 2014, 7, 363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, E.L.; de Vlas, S.J.; Fronterre, C.; Hollingsworth, T.D.; Kontoroupis, P.; Michael, E.; Prada, J.M.; Smith, M.E.; Stolk, W.A.; Touloupou, P. The Roadmap towards Elimination of Lymphatic Filariasis by 2030: Insights from Quantitative and Mathematical Modelling. Gates Open Res. 2019, 3, 1538. [Google Scholar] [CrossRef] [Green Version]
- Basáñez, M.G.; Walker, M.; Hamley, J.I.D.; Milton, P.; Fronterre, C.; de Vlas, S.J.; Stolk, W.A. The World Health Organization 2030 Goals for Onchocerciasis: Insights and Perspectives from Mathematical Modelling. Gates Open Res. 2019, 3, 1545. [Google Scholar] [CrossRef]
- Tyagi, R.; Bulman, C.A.; Cho-Ngwa, F.; Fischer, C.; Marcellino, C.; Arkin, M.R.; McKerrow, J.H.; McNamara, C.W.; Mahoney, M.; Tricoche, N.; et al. An Integrated Approach to Identify New Anti-Filarial Leads to Treat River Blindness, a Neglected Tropical Disease. Pathogens 2021, 10, 71. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.C.; Kinkel, A.W.; Gryczko, C.M.; Goulet, J.R. Absorption of Pyrvinium Pamoate. Clin. Pharmacol. Ther. 1976, 19, 802–806. [Google Scholar] [CrossRef]
- Hamilton, G.; Rath, B. Repurposing of Anthelminthics as Anticancer Drugs. Oncomedicine 2017, 3, 1–8. [Google Scholar] [CrossRef]
- Jones, J.O.; Bolton, E.C.; Huang, Y.; Feau, C.; Guy, R.K.; Yamamoto, K.R.; Hann, B.; Diamond, M.I. Non-Competitive Androgen Receptor Inhibition in vitro and In Vivo. Proc. Natl. Acad. Sci. USA 2009, 106, 7233–7238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diamond, M.I.; Jones, J.O.; Renslo, A.R. Small-Molecule Inhibitors of the Androgen Receptor. U.S. Patent 8,580,773 B2, 12 November 2013. [Google Scholar]
- Beck, J.W.; Saavedra, D.; Antell, G.J.; Tejeiro, B. The Treatment of Pinworm Infections in Humans (Enterobiasis) with Pyrvinium Cloride and Pyrvinium Pamoate. Am. J. Trop. Med. Hyg. 1959, 8, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Beck, J.W. Treatment of Pinworm Infections with Reduced Single Dose of Pyrvinium Pamoate. JAMA J. Am. Med. Assoc. 1964, 189, 511. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.C.; Galli, G.A. Strongyloidiasis Treated with Pyrvinium Pamoate. J. Am. Med. Assoc. 1965, 193, 191–192. [Google Scholar] [CrossRef]
- De Padua Vilela, M.; Zucas, A.W.; Iglesias, J. The Therapy of Trichuriasis with a Combination of Thiabendazole and Pyrvinium Pamoate. J. New Drugs 1965, 5, 86–89. [Google Scholar] [CrossRef]
- Downey, A.S.; Chong, C.R.; Graczyk, T.K.; Sullivan, D.J. Efficacy of Pyrvinium Pamoate against Cryptosporidium parvum Infection in vitro and in a Neonatal Mouse Model. Antimicrob. Agents Chemother. 2008, 52, 3106–3112. [Google Scholar] [CrossRef] [Green Version]
- Downey, A.S.; Graczyk, T.K.; Sullivan, D.J. in vitro Activity of Pyrvinium Pamoate against Entamoeba histolytica and Giardia intestinalis Using Radiolabelled Thymidine Incorporation and an SYBR Green I-Based Fluorescence Assay. J. Antimicrob. Chemother. 2009, 64, 751–754. [Google Scholar] [CrossRef] [Green Version]
- Gaikwad, V.R.; Karale, U.B.; Govindarajalu, G.; Adhikari, N.; Krishna, E.V.; Siva, V.; Misra, S.; Sriram, D.; Sijwali, P.S.; Rode, H.B. Synthesis and Efficacy of Pyrvinium-Inspired Analogs against Tuberculosis and Malaria Pathogens. Biooroganic. Med. Chem. Lett. 2020, 30, 127037. [Google Scholar] [CrossRef]
- Sun, Y.; Gao, L.; Yuan, M.; Yuan, L.; Yang, J.; Zeng, T. In Vitro and In Vivo Study of Antifungal Effect of Pyrvinium Pamoate Alone and in Combination with Azoles Against Exophiala dermatitidis. Front. Cell. Infect. Microbiol. 2020, 10, 576975. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Mulla, W.A.; Kucharavy, A.; Tsai, H.J.; Rubinstein, B.; Conkright, J.; McCroskey, S.; Bradford, W.D.; Weems, L.; Haug, J.S.; et al. Targeting the Adaptability of Heterogeneous Aneuploids. Cell 2015, 160, 771–784. [Google Scholar] [CrossRef] [Green Version]
- Jawahar, S.; Tricoche, N.; Bulman, C.A.; Sakanari, J.; Lustigman, S. Drugs That Target Early Stages of Onchocerca volvulus: A Revisited Means to Facilitate the Elimination Goals for Onchocerciasis. PLoS Negl. Trop. Dis. 2021, 15, e0009064. [Google Scholar] [CrossRef] [PubMed]
- Kamgno, J.; Pion, S.D.; Chesnais, C.B.; Bakalar, M.H.; D’Ambrosio, M.V.; Mackenzie, C.D.; Nana-Djeunga, H.C.; Gounoue-Kamkumo, R.; Njitchouang, G.R.; Nwane, P.; et al. Test-and-Not-Treat Strategy for Onchocerciasis in Loa loa-Endemic Areas. N. Engl. J. Med. 2017, 377, 2044–2052. [Google Scholar] [CrossRef] [Green Version]
- Drugs for Neglected Diseases Initiative (DNDi). Available online: https://dndi.org/research-development/portfolio/ (accessed on 18 December 2021).
- Wanji, S.; Chunda, V.C.; Fombad, F.F.; Njouendou, A.J.; Gandjui, N.V.T.; Ritter, M.; Enyong, P.A.; Mackenzie, C.; Taylor, M.J.; Hoerauf, A.; et al. Advances in Preclinical Platforms of Loa loa for Filarial Neglected Tropical Disease Drug and Diagnostics Research. Front. Trop. Dis. 2021, 2, 778724. [Google Scholar] [CrossRef]
- DNDi. Target Product Profile for River Blindness. Available online: https://dndi.org/diseases/filaria-river-blindness/target-product-profile/ (accessed on 18 December 2021).
- Hoerauf, A.; Nissen-Pähle, K.; Schmetz, C.; Henkle-Dührsen, K.; Blaxter, M.L.; Büttner, D.W.; Gallin, M.Y.; Al-Qaoud, K.M.; Lucius, R.; Fleischer, B. Tetracycline Therapy Targets Intracellular Bacteria in the Filarial Nematode Litomosoides sigmodontis and Results in Filarial Infertility. J. Clin. Investig. 1999, 103, 11–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoerauf, A.; Mand, S.; Fischer, K.; Kruppa, T.; Marfo-Debrekyei, Y.; Debrah, A.Y.; Pfarr, K.M.; Adjei, O.; Büttner, D.W. Doxycycline as a Novel Strategy against Bancroftian Filariasis—Depletion of Wolbachia Endosymbionts from Wuchereria bancrofti and Stop of Microfilaria Production. Med. Microbiol. Immunol. 2003, 192, 211–216. [Google Scholar] [CrossRef]
- Aljayyoussi, G.; Tyrer, H.E.; Ford, L.; Sjoberg, H.; Pionnier, N.; Waterhouse, D.; Davies, J.; Gamble, J.; Metugene, H.; Cook, D.A.N.; et al. Short-Course, High-Dose Rifampicin Achieves Wolbachia Depletion Predictive of Curative Outcomes in Preclinical Models of Lymphatic Filariasis and Onchocerciasis. Sci. Rep. 2017, 7, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, M.J.; Makunde, W.H.; McGarry, H.F.; Turner, J.D.; Mand, S.; Hoerauf, A. Macrofilaricidal Activity after Doxycycline Treatment of Wuchereria bancrofti: A Double-Blind, Randomised Placebo-Controlled Trial. Lancet 2005, 365, 2116–2121. [Google Scholar] [CrossRef]
- Debrah, A.Y.; Mand, S.; Marfo-Debrekyei, Y.; Batsa, L.; Pfarr, K.; Buttner, M.; Adjei, O.; Buttner, D.; Hoerauf, A. Macrofilaricidal Effect of 4 Weeks of Treatment with Doxycycline on Wuchereria bancrofti. Trop. Med. Int. Health 2007, 12, 1433–1441. [Google Scholar] [CrossRef] [PubMed]
- Hoerauf, A.; Specht, S.; Buttner, M.; Pfarr, K.; Mand, S.; Fimmers, R.; Marfo-Debrekyei, Y.; Konadu, P.; Debrah, A.Y.; Bandi, C.; et al. Wolbachia Endobacteria Depletion by Doxycycline as Antifilarial Therapy Has Macrofilaricidal Activity in Onchocerciasis: A Randomized Placebo-Controlled Study. Med. Microbiol. Immunol. 2008, 197, 335. [Google Scholar] [CrossRef] [PubMed]
- Hoerauf, A.; Specht, S.; Marfo-Debrekyei, Y.; Buttner, M.; Debrah, A.Y.; Mand, S.; Batsa, L.; Brattig, N.; Konadu, P.; Bandi, C.; et al. Efficacy of 5-Week Doxycycline Treatment on Adult Onchocerca volvulus. Parasitol. Res. 2009, 104, 437–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Specht, S.; Mand, S.; Marfo-Debrekyei, Y.; Debrah, A.Y.; Konadu, P.; Adjei, O.; Büttner, D.W.; Hoerauf, A. Efficacy of 2- and 4-Week Rifampicin Treatment on the Wolbachia of Onchocerca volvulus. Parasitol. Res. 2008, 103, 1303–1309. [Google Scholar] [CrossRef] [PubMed]
- Sungpradit, S.; Chatsuwan, T.; Nuchprayoon, S. Susceptibility of Wolbachia, an Endosymbiont of Brugia malayi Microfilariae, to Doxycycline Determined by Quantitative PCR Assay. Southeast. Asian J. Trop. Med. Public Health 2012, 43, 841–850. [Google Scholar] [PubMed]
- Debrah, A.Y.; Specht, S.; Klarmann-schulz, U.; Batsa, L.; Mand, S.; Marfo-debrekyei, Y.; Fimmers, R.; Dubben, B.; Kwarteng, A.; Osei-atweneboana, M.; et al. Doxycycline Leads to Sterility and Enhanced Killing of Female Onchocerca volvulus Worms in an Area with Persistent Microfilaridermia After Repeated Ivermectin Treatment: A Randomized, Placebo-Controlled, Double-Blind Trial. Clin. Infect. Dis. 2015, 61, 517–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishii, I.; Harada, Y.; Kasahara, T. Reprofiling a Classical Anthelmintic, Pyrvinium Pamoate, as an Anti-Cancer Drug Targeting Mitochondrial Respiration. Front. Oncol. 2012, 2, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.; Lacerda, L.; Debeb, B.G.; Atkinson, R.L.; Solley, T.N.; Li, L.; Orton, D.; McMurray, J.S.; Hang, B.I.; Lee, E.; et al. The Antihelmintic Drug Pyrvinium Pamoate Targets Aggressive Breast Cancer. PLoS ONE 2013, 8, e71508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Momtazi-Borojeni, A.A.; Abdollahi, E.; Ghasemi, F.; Caraglia, M.; Sahebkar, A. The Novel Role of Pyrvinium in Cancer Therapy. J. Cell. Physiol. 2018, 233, 2871–2881. [Google Scholar] [CrossRef]
- Guan, Q.; Zhan, L.; Liu, Z.H.; Pan, Q.; Chen, X.L.; Xiao, Z.; Qin, C.; Zhang, X.L. Identification of Pyrvinium Pamoate as an Anti-Tuberculosis Agent in vitro and In Vivo by SOSA Approach amongst Known Drugs. Emerg. Microbes Infect. 2020, 9, 302–312. [Google Scholar] [CrossRef]
- Bulman, C.A.; Bidlow, C.M.; Lustigman, S.; Cho-Ngwa, F.; Williams, D.; Rascon, A.A., Jr.; Tricoche, N.; Samje, M.; Bell, A.; Suzuki, B.; et al. Repurposing Auranofin as a Lead Candidate for Treatment of Lymphatic Filariasis and Onchocerciasis. PLoS Negl. Trop. Dis. 2015, 9, e0003534. [Google Scholar] [CrossRef]
- Voronin, D.; Tricoche, N.; Jawahar, S.; Shlossman, M.; Bulman, C.A.; Fischer, C.; Suderman, M.T.; Sakanari, J.A.; Lustigman, S. Development of a Preliminary In Vitro Drug Screening Assay Based on a Newly Established Culturing System for Pre-Adult Fifth-Stage Onchocerca volvulus Worms. PLoS Negl. Trop. Dis. 2019, 13, e0007108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunderson, E.L.; Vogel, I.; Chappell, L.; Bulman, C.A.; Lim, K.C.; Luo, M.; Whitman, J.D.; Franklin, C.; Choi, Y.-J.; Lefoulon, E.; et al. The Endosymbiont Wolbachia Rebounds Following Antibiotic Treatment. PLoS Pathog. 2020, 16, e1008623. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Brugia Adult Females Inhibition of Motility Day 3 @ 1 µM | Brugia Adult Females Inhibition of Motility Day 3 IC50 (µM) | O. volvulus L5 Inhibition of Motility Up to Day 36 | O. volvulus L5 Inhibition of Motility IC50 (µM) | O. volvulus L3 Inhibition of Molting Day 6 IC50 (µM) | O. ochengi Adult Males Inhibition of Motility Day 5 IC50 (µM) | O. ochengi Adult Females Inhibition of Survival Day 7 IC50 (µM) | O. ochengi Mf Inhibition of Motility Day 5 @ 10 µM | Loa loa Mf Inhibition of Motility Day 5 @ 10 µM |
|---|---|---|---|---|---|---|---|---|---|
| 01 | 99% | 0.008 | 94% on Day 36 @ 1 µM | nd | nd | 0.96 | 0.79 | 100% | 100% |
| 02 | 83% | 0.001 | 90% on Day 16 @ 1 µM | nd | nd | 3.08 | 0.70 | 100% | 100% |
| 03 | 3% | nd | 97% on Day 28 @ 10 µM | nd | 0.14 | 1.54 | 5.18 | 0% | 13% |
| 04 | 26% | nd | nd | nd | nd | * 100% @ 10 µM | nd | 0% | 38% |
| 05 | 81% | 0.405 | 92% on Day 36 @ 1 µM | nd | nd | * 86% @ 10 µM | nd | 0% | 38% |
| 06 | 71% | 0.733 | 94% on Day 36 @ 1 µM | 0.22 µM on Day 21 | 0.46 | 2.75 | 2.99 | 46% | 88% |
| THP | 100% | 0.003 | 68% on Day 36 @ 1 µM | <1.0 µM on Day 21 | 0.19 | 0.06 | 0.61 | 100% | 100% |
| PVP | 82% | 0.0003 | 100% on Day 14 @ 1 µM | <0.3 µM on Day 21 | 0.03 | 0.11 | 0.05 | 100% and ** IC50 = 0.3 µM | 100% |
| Compound | Gerbil Liver Microsome T1/2 (min) | CLint (µL/min/mg Protein) | Mean PappA-B (×10−6 cm/s) | Mean PappB-A (×10−6 cm/s) | MDCK Efflux Ratio B-A/A-B | MDCK Recovery Rate (%) |
|---|---|---|---|---|---|---|
| 01 | 2.3 | 610.8 | 0.1 | 61.3 | 531.9 | 77 |
| 02 | 2.6 | 541.8 | 0.1 | 51.2 | 736.7 | 67 |
| 03 | 4.9 | 280.2 | 1.3 | 9.1 | 7.2 | 104 |
| 05 | 7.7 | 179.88 | 0.7 | 6.8 | 9.3 | 96 |
| 06 | 1484.9 | 0.933 | 0.5 | 12.1 | 23.7 | 94 |
| THP | 14.6 | 95.14 | <0.25 | 0.6 | >2.5 | 15 |
| PVP | 5.3 | 261.6 | <0.07 | 6.6 | >94.6 | 69 |
| Dosage of 1 mg/kg | ||||
|---|---|---|---|---|
| Vehicle | THP | 06 | PVP | |
| Number of animals | 8 | 8 | 8 | 7 |
| Adult worms per animal | 18.5 (8.2) | 21.9 (7.6) | 31.3 (11.8) | 0.9 (0.5) |
| Female worms per animal | 8.5 (3.9) | 10.4 (4.2) | 15.9 (5.7) | 0.1 (0.1) |
| Male worms per animal | 10 (4.3) | 11.5 (3.7) | 15.4 (6.1) | 0.7 (0.4) |
| Mf per animal | 1,271,443 (1,092,551) | 652,500 (326,014) | 915,000 (263,235) | 10,429 |
| (5822) | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gunderson, E.L.; Bryant, C.; Bulman, C.A.; Fischer, C.; Luo, M.; Vogel, I.; Lim, K.-C.; Jawahar, S.; Tricoche, N.; Voronin, D.; et al. Pyrvinium Pamoate and Structural Analogs Are Early Macrofilaricide Leads. Pharmaceuticals 2022, 15, 189. https://doi.org/10.3390/ph15020189
Gunderson EL, Bryant C, Bulman CA, Fischer C, Luo M, Vogel I, Lim K-C, Jawahar S, Tricoche N, Voronin D, et al. Pyrvinium Pamoate and Structural Analogs Are Early Macrofilaricide Leads. Pharmaceuticals. 2022; 15(2):189. https://doi.org/10.3390/ph15020189
Chicago/Turabian StyleGunderson, Emma L., Clifford Bryant, Christina A. Bulman, Chelsea Fischer, Mona Luo, Ian Vogel, Kee-Chong Lim, Shabnam Jawahar, Nancy Tricoche, Denis Voronin, and et al. 2022. "Pyrvinium Pamoate and Structural Analogs Are Early Macrofilaricide Leads" Pharmaceuticals 15, no. 2: 189. https://doi.org/10.3390/ph15020189
APA StyleGunderson, E. L., Bryant, C., Bulman, C. A., Fischer, C., Luo, M., Vogel, I., Lim, K. -C., Jawahar, S., Tricoche, N., Voronin, D., Corbo, C., Ayiseh, R. B., Manfo, F. P. T., Mbah, G. E., Cho-Ngwa, F., Beerntsen, B., Renslo, A. R., Lustigman, S., & Sakanari, J. A. (2022). Pyrvinium Pamoate and Structural Analogs Are Early Macrofilaricide Leads. Pharmaceuticals, 15(2), 189. https://doi.org/10.3390/ph15020189