
Asperuloside Prevents Peri-Implantitis via Suppression of NF-κB and ERK1/2 on Rats

 , , and
, , and
Abstract
:1. Introduction
2. Results
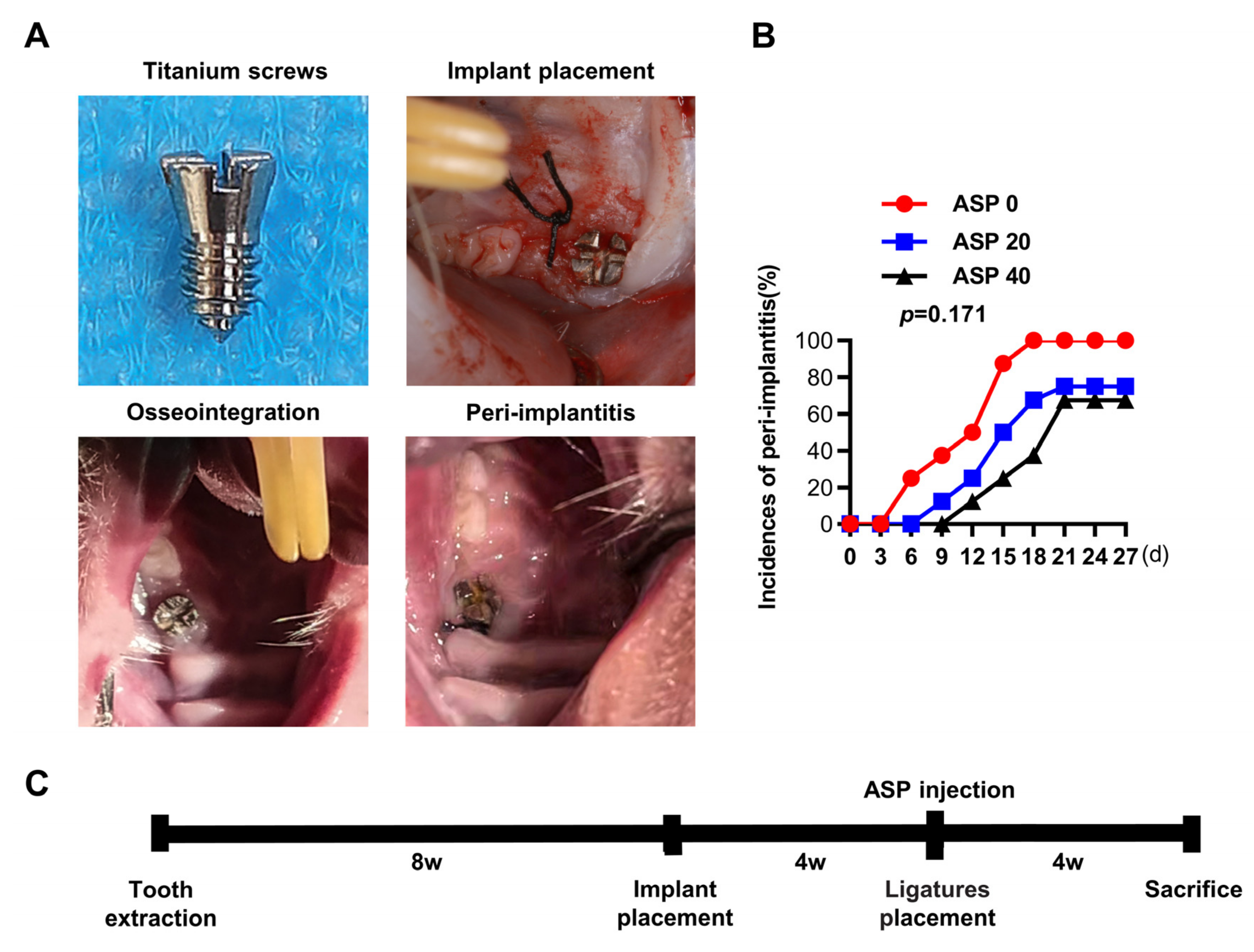
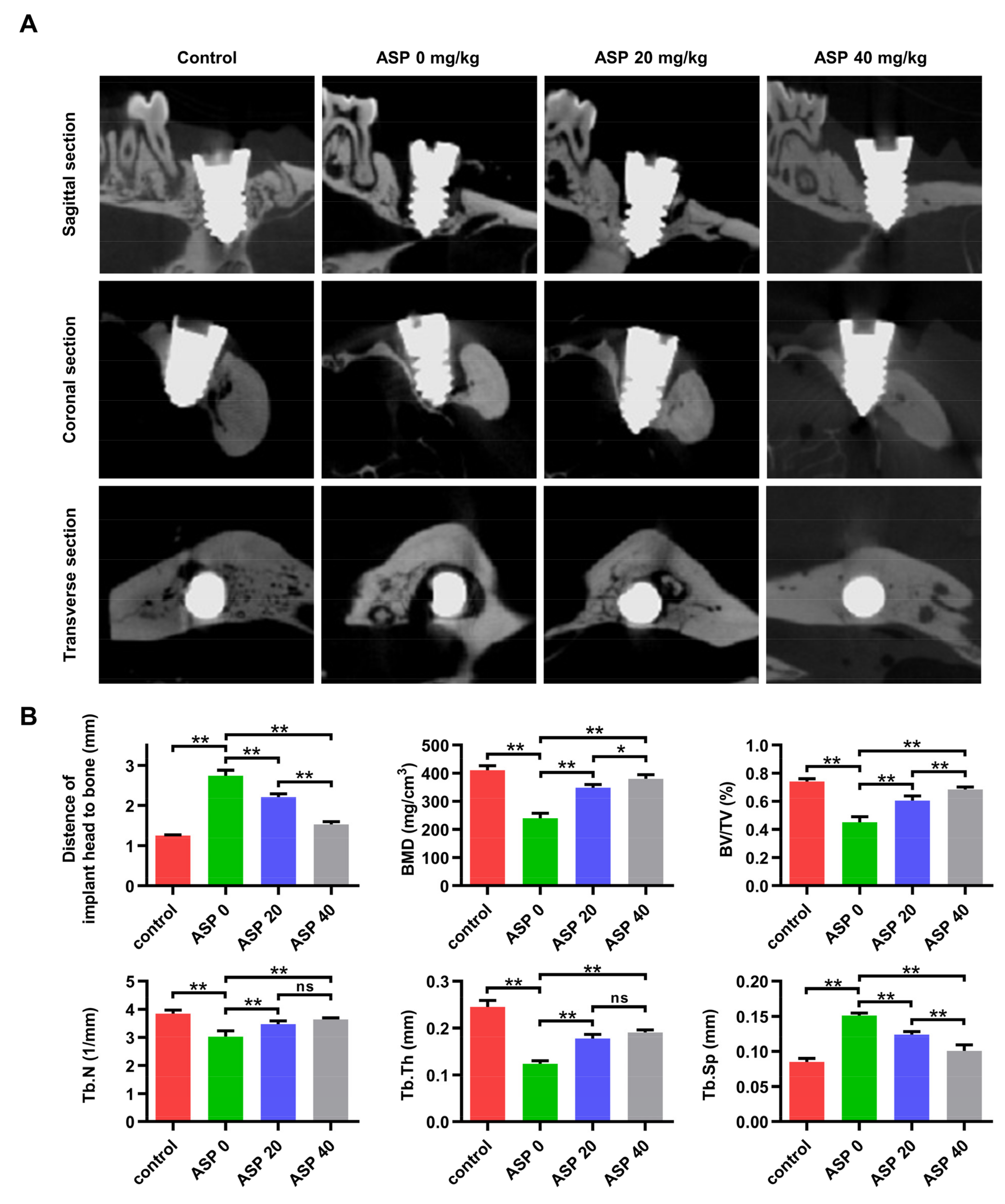
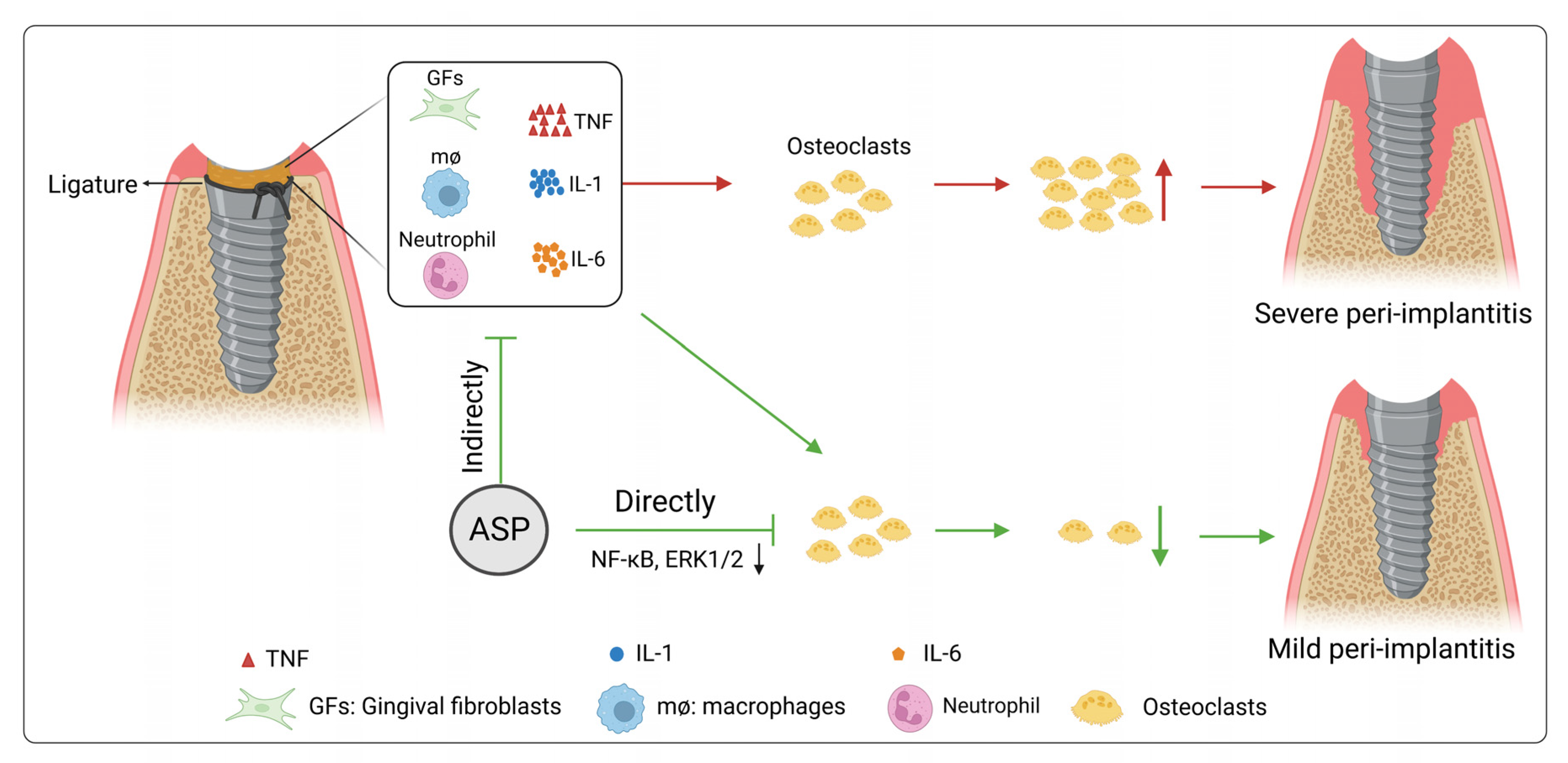
2.1. ASP Caused a Decrease in the Morbidity and Alveolar Bone Resorption of Peri-Implantitis
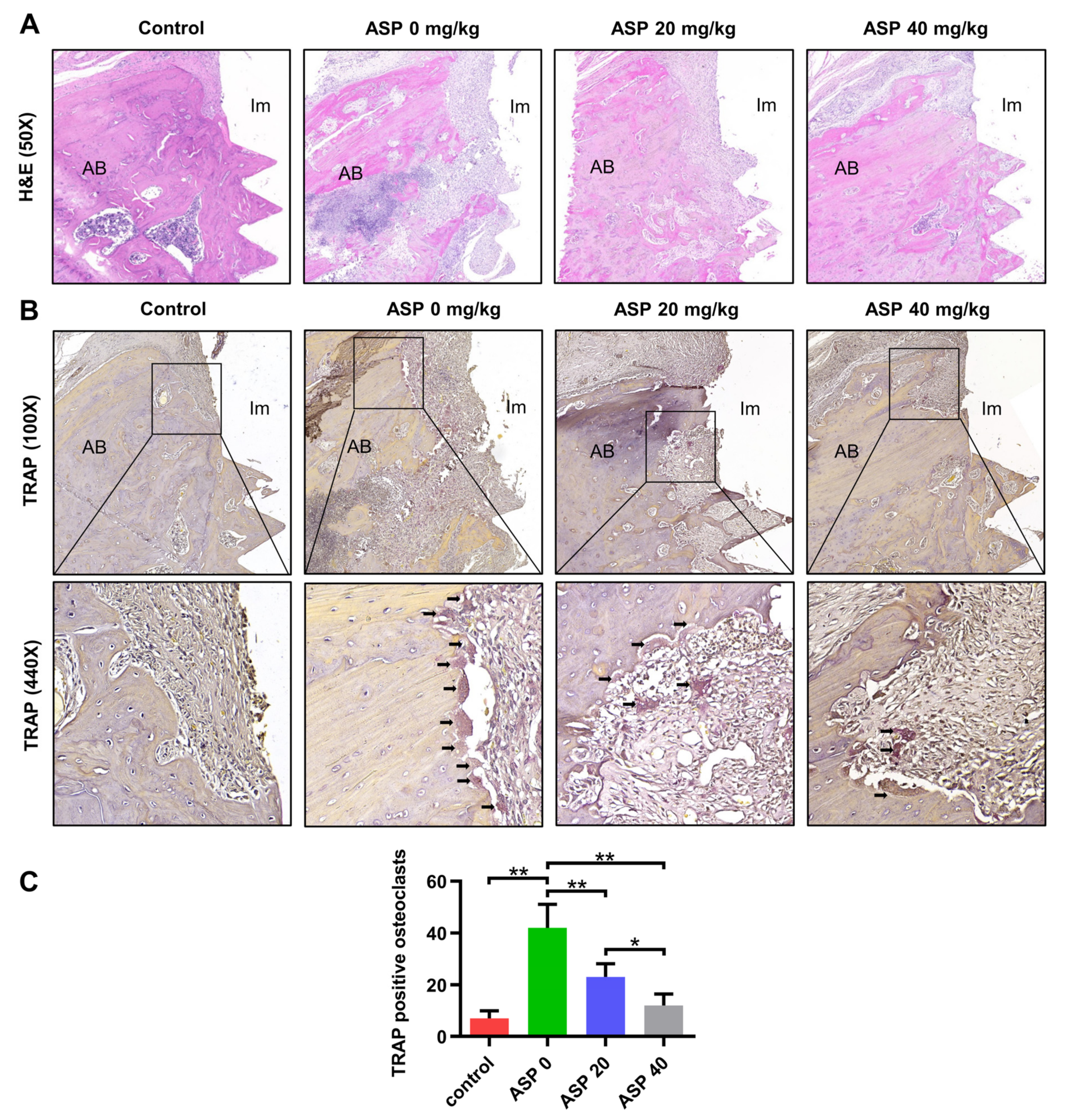
2.2. ASP Decreased the Inflammatory Tissues and the Osteoclast Quantity of Peri-Implantitis
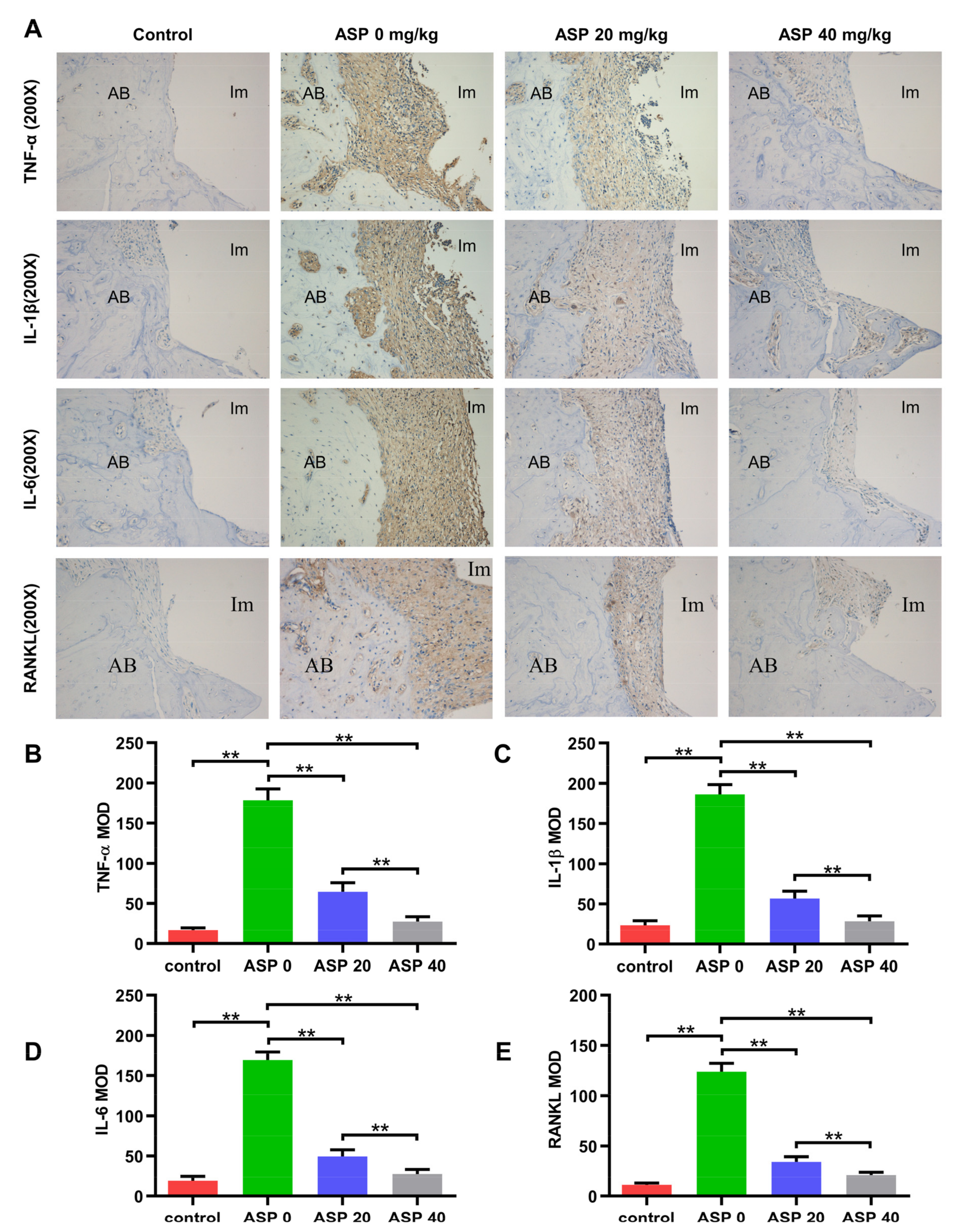
2.3. ASP Caused a Decrease of Pro-Inflammatory Cytokines and RANKL in Peri-Implant Tissues
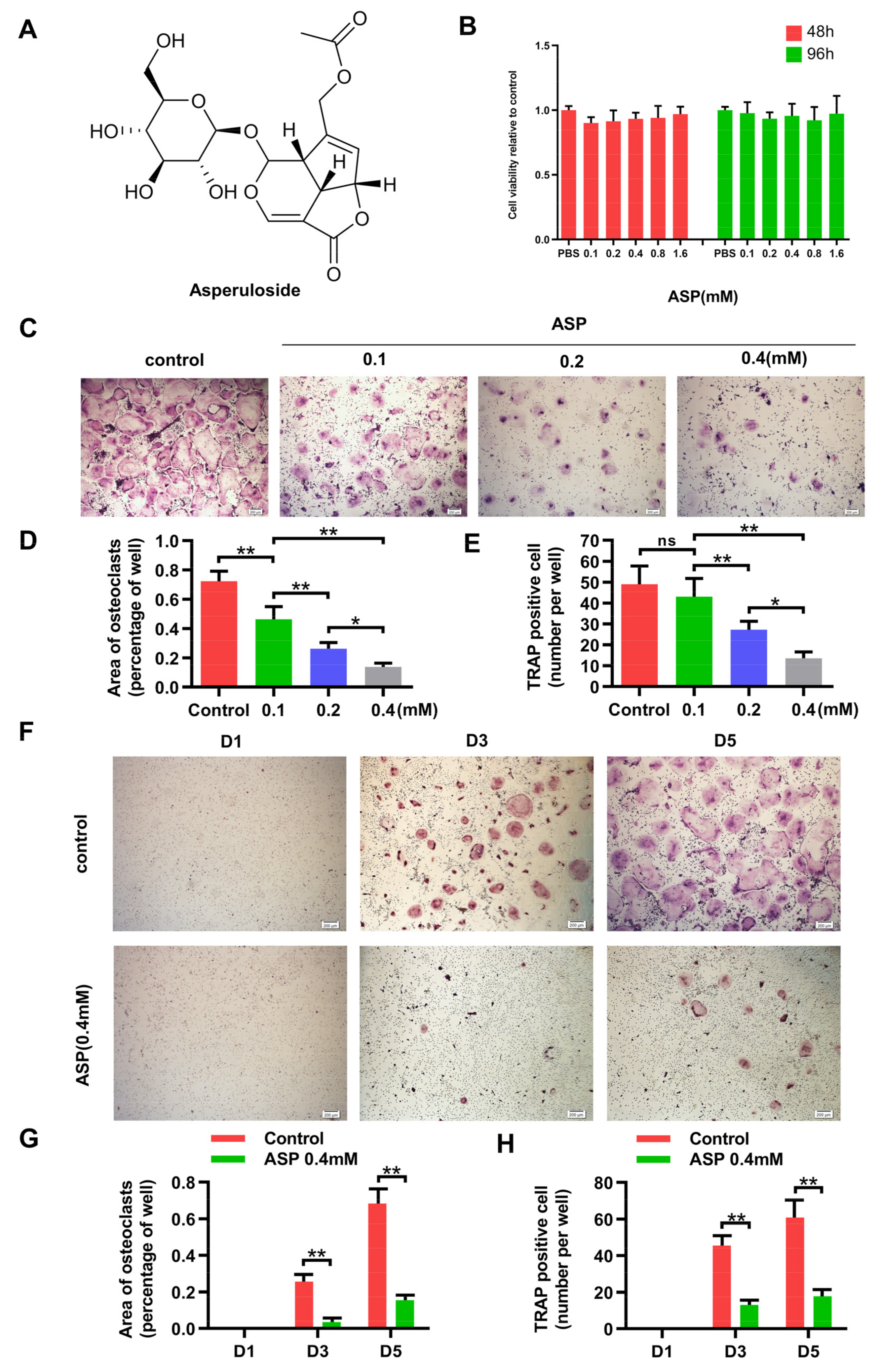
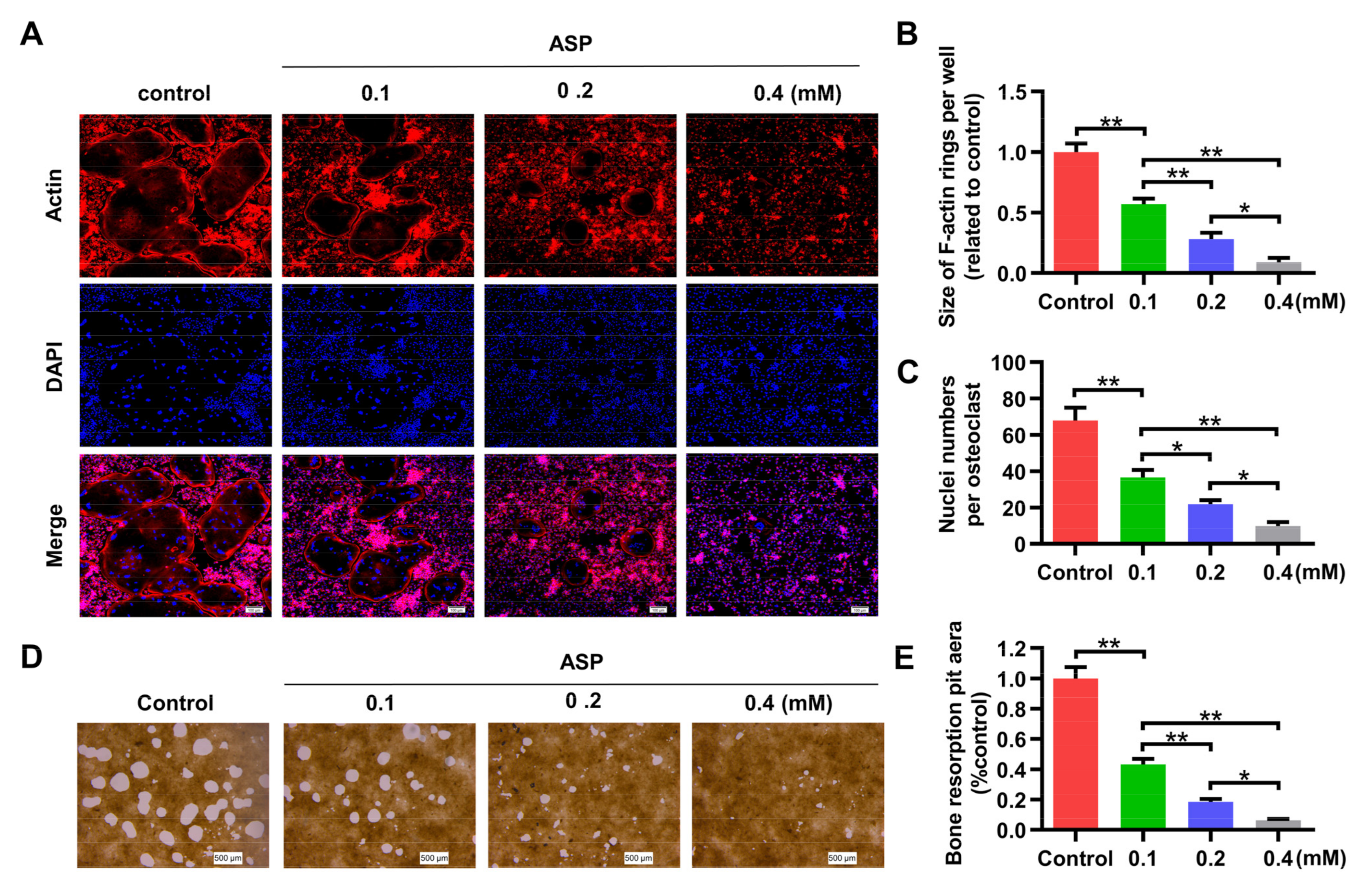
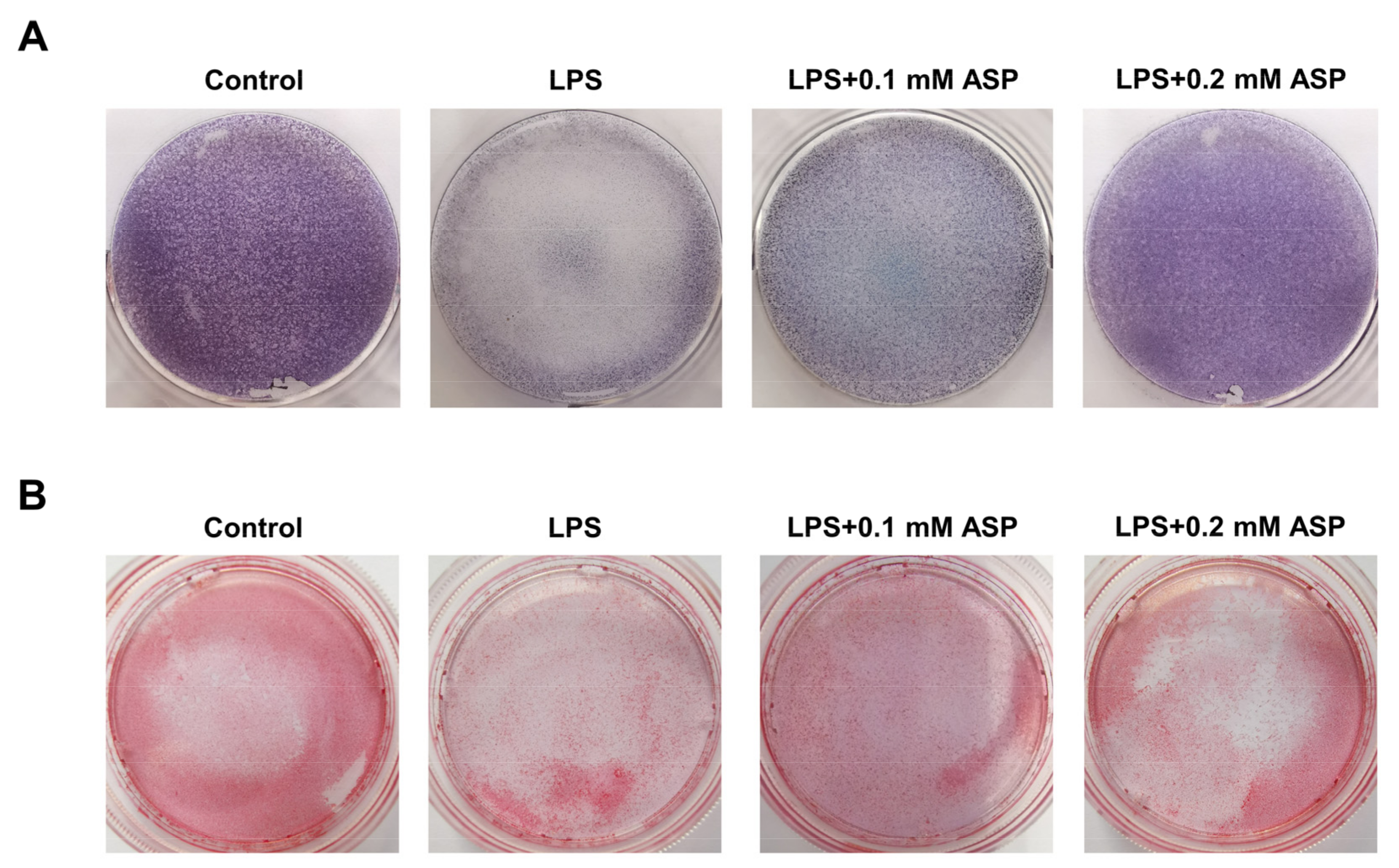
2.4. ASP Inhibited RANKL-Induced Osteoclast Formation In Vitro
2.5. ASP Led to Inhibition of Bone Resorption Function of Osteoclasts In Vitro
2.6. ASP Caused Downregulation of Osteoclast-Specific Gene Expression and Protein Expression
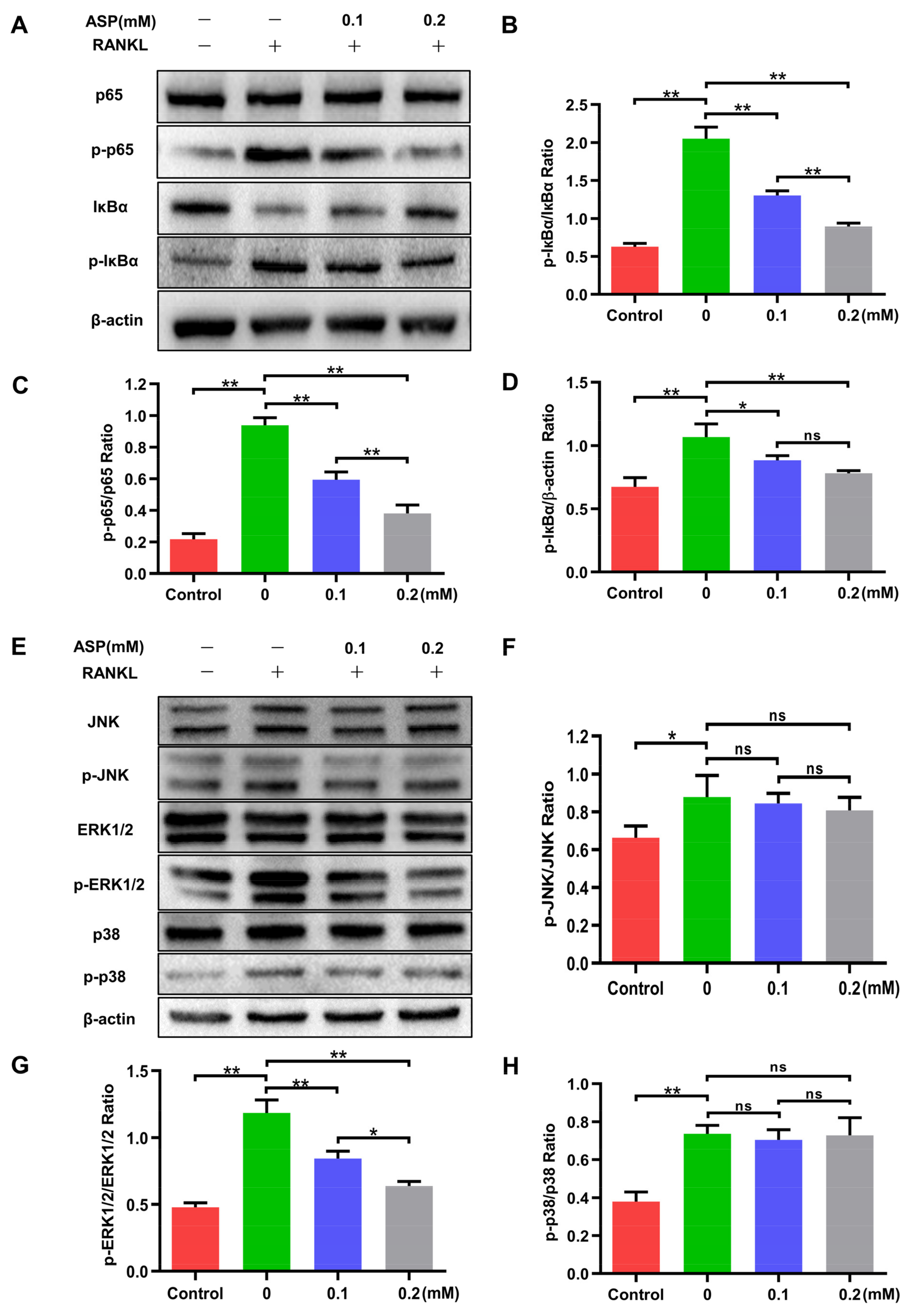
2.7. ASP Attenuated the Activation of RANKL-Induced NF-κB and ERK1/2
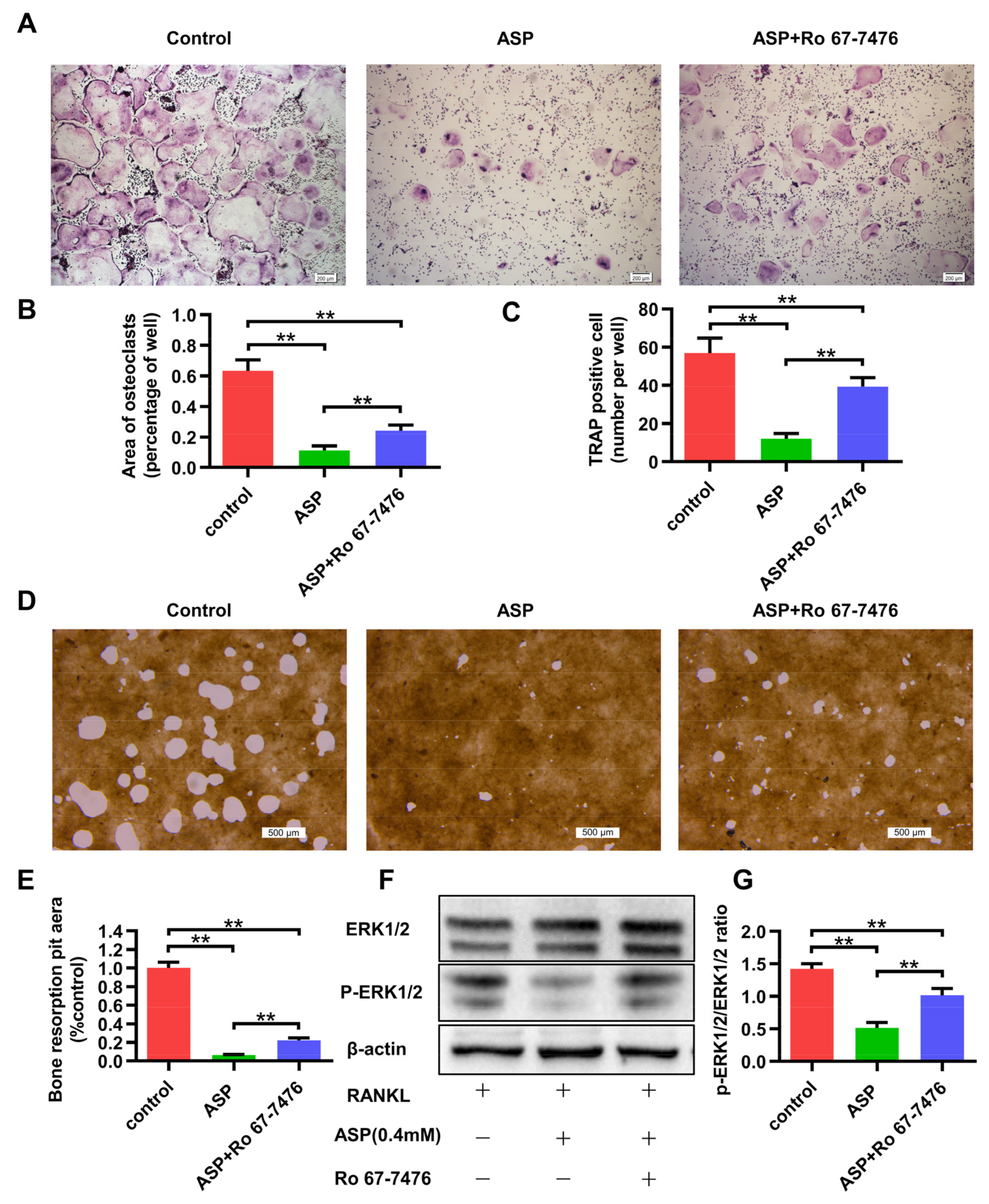
2.8. The ERK1/2 phosphorylation Agonist Caused Partial Reversal of the Inhibiting Effects of ASP on Osteoclastogenesis
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
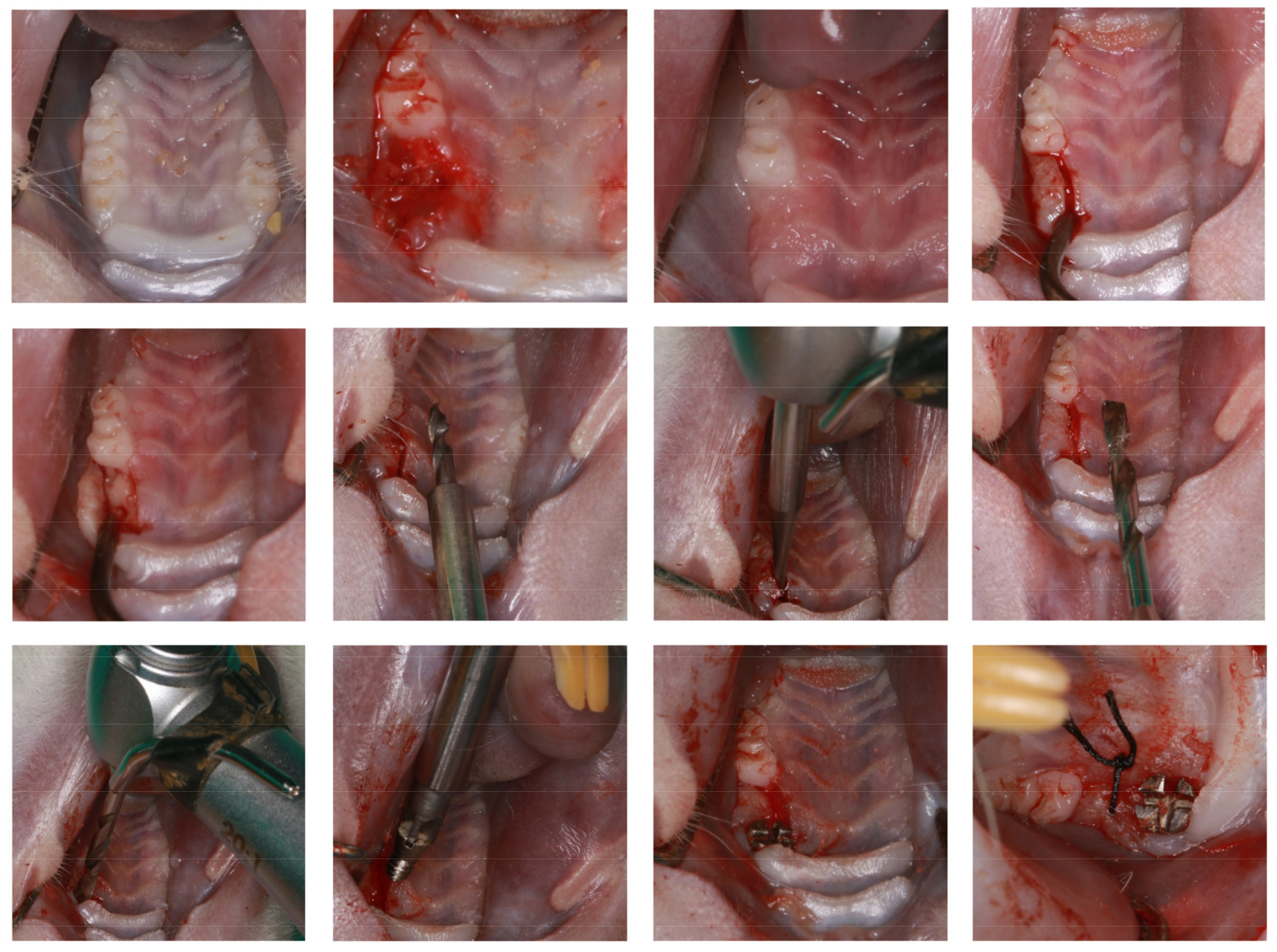
4.2. Animal Experiments
4.3. Micro-CT Scanning
4.4. H&E Staining and TRAP Staining
4.5. Immunohistochemistry Staining
4.6. Bone Marrow-Derived Macrophages (BMMs) Culture and Cell Viability Assay
4.7. Cell Culture and TRAP Staining Assay
4.8. Fibrous Actin (F-Actin) Ring Staining Assay
4.9. In Vitro Hydroxyapatite Resorption Assay
4.10. Real-Time PCR
4.11. Western Blotting Assay
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Karoussis, I.K.; Kotsovilis, S.; Fourmousis, I. A comprehensive and critical review of dental implant prognosis in periodontally compromised partially edentulous patients. Clin. Oral Implant. Res. 2007, 18, 669–679. [Google Scholar] [CrossRef] [PubMed]
- Bornstein, M.M.; Halbritter, S.; Harnisch, H.; Weber, H.P.; Buser, D. A retrospective analysis of patients referred for implant placement to a specialty clinic: Indications, surgical procedures, and early failures. Int. J. Oral Maxillofac. Implants 2008, 23, 1109–1116. [Google Scholar] [CrossRef] [PubMed]
- Berglundh, T.; Armitage, G.; Araujo, M.G.; Avila-Ortiz, G.; Blanco, J.; Camargo, P.M.; Chen, S.; Cochran, D.; Derks, J.; Figuero, E.; et al. Peri-implant diseases and conditions: Consensus report of workgroup 4 of the 2017 World Workshop on the Classification of Periodontal and Peri-Implant Diseases and Conditions. J. Clin. Periodontol. 2018, 45 (Suppl. S20), S286–S291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konstantinidis, I.K.; Kotsakis, G.; Gerdes, S.; Walter, M.H. Cross-sectional study on the prevalence and risk indicators of peri-implant diseases. Eur. J. Oral Implant. 2015, 8, 75–88. [Google Scholar]
- Safioti, L.M.; Kotsakis, G.; Pozhitkov, A.; Chung, W.O.; Daubert, D.M. Increased Levels of Dissolved Titanium Are Associated With Peri-Implantitis—A Cross-Sectional Study. J. Periodontol. 2017, 88, 436–442. [Google Scholar] [CrossRef]
- Sanz, M.; Noguerol, B.; Sanz-Sanchez, I.; Hammerle, C.H.F.; Schliephake, H.; Renouard, F.; Sicilia, A.; Steering Committee; Cordaro, L.; Jung, R.; et al. European Association for Osseointegration Delphi study on the trends in Implant Dentistry in Europe for the year 2030. Clin. Oral Implant. Res. 2019, 30, 476–486. [Google Scholar] [CrossRef]
- Scarano, A.; de Oliveira, P.S.; Leo, L.; Festa, F.; Carinci, F.; Lorusso, F. Evaluation of a new antibacterial coating of the internal chamber of an implant via real time measurement of Volatile Organic Compounds (VOCs). Front. Biosci. 2021, 13, 216. [Google Scholar] [CrossRef]
- Petkovic-Curcin, A.; Matic, S.; Vojvodic, D.; Stamatovic, N.; Todorovic, T. Cytokines in pathogenesis of peri-implantitis. Vojn. Pregl. 2011, 68, 435–440. [Google Scholar] [CrossRef]
- Mordini, L.; Sun, N.; Chang, N.; De Guzman, J.-P.; Generali, L.; Consolo, U. Peri-Implantitis Regenerative Therapy: A Review. Biology 2021, 10, 773. [Google Scholar] [CrossRef]
- Li, H.; Chen, Z.; Zhong, X.; Li, J.; Li, W. Mangiferin alleviates experimental peri-implantitis via suppressing interleukin-6 production and Toll-like receptor 2 signaling pathway. J. Orthop. Surg. Res. 2019, 14, 325. [Google Scholar] [CrossRef] [Green Version]
- Teitelbaum, S.L. Bone Resorption by Osteoclasts. Science 2000, 289, 1504–1508. [Google Scholar] [CrossRef] [PubMed]
- Bar-Shavit, Z. The osteoclast: A multinucleated, hematopoietic-origin, bone-resorbing osteoimmune cell. J. Cell. Biochem. 2007, 102, 1130–1139. [Google Scholar] [CrossRef] [PubMed]
- Pearson, G.; Robinson, F.; Beers Gibson, T.; Xu, B.E.; Karandikar, M.; Berman, K.; Cobb, M.H. Mitogen-Activated Protein (MAP) Kinase Pathways: Regulation and Physiological Functions. Endocr. Rev. 2001, 22, 153–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast differentiation and activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef]
- Baud’Huin, M.; Duplomb, L.; Velasco, C.R.; Fortun, Y.; Heymann, D.; Padrines, M. Key roles of the OPG–RANK–RANKL system in bone oncology. Expert Rev. Anticancer Ther. 2007, 7, 221–232. [Google Scholar] [CrossRef]
- Qu, S.; Bai, Y.; Liu, X.; Fu, R.; Duan, K.; Weng, J. Study on in vitro release and cell response to alendronate sodium-loaded ultrahigh molecular weight polyethylene loaded with alendronate sodium wear particles to treat the particles-induced osteolysis. J. Biomed. Mater. Res. Part A 2013, 101, 394–403. [Google Scholar] [CrossRef]
- Liu, S.; Virdi, A.S.; Sena, K.; Sumner, D.R. Sclerostin antibody prevents particle-induced implant loosening by stimulating bone formation and inhibiting bone resorption in a rat model. Arthritis Care Res. 2012, 64, 4012–4020. [Google Scholar] [CrossRef]
- Liu, X.; Zhu, S.; Cui, J.; Shao, H.; Zhang, W.; Yang, H.; Xu, Y.; Geng, D.; Yu, L. Strontium ranelate inhibits titanium-particle-induced osteolysis by restraining inflammatory osteoclastogenesis in vivo. Acta Biomater. 2014, 10, 4912–4918. [Google Scholar] [CrossRef]
- Ilyas, Z.; Camacho, P.M. Rare adverse effects of bisphosphonate therapy. Curr. Opin. Endocrinol. Diabetes Obes. 2019, 26, 335–338. [Google Scholar] [CrossRef]
- McClung, M.R.; Grauer, A.; Boonen, S.; Bolognese, M.A.; Brown, J.P.; Diez-Perez, A.; Langdahl, B.L.; Reginster, J.-Y.; Zanchetta, J.R.; Wasserman, S.M.; et al. Romosozumab in Postmenopausal Women with Low Bone Mineral Density. N. Engl. J. Med. 2014, 370, 412–420. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.Y.; Shen, M.X.; Lim, Y.L.; Tay, Y.K.; Chan, M.M.F.; Pang, S.M.; Xiao, Z.W.; Ang, S.B.; Ren, E.C. Increased risk of strontium ranelate-related SJS/TEN is associated with HLA. Osteoporos. Int. 2016, 27, 2577–2583. [Google Scholar] [CrossRef]
- Wei, J.; Li, Y.; Liu, Q.; Lan, Y.; Wei, C.; Tian, K.; Wu, L.; Lin, C.; Xu, J.; Zhao, J.; et al. Betulinic Acid Protects From Bone Loss in Ovariectomized Mice and Suppresses RANKL-Associated Osteoclastogenesis by Inhibiting the MAPK and NFATc1 Pathways. Front. Pharmacol. 2020, 11, 1025. [Google Scholar] [CrossRef]
- Qiu, J.; Chi, G.; Wu, Q.; Ren, Y.; Chen, C.; Feng, H. Pretreatment with the compound asperuloside decreases acute lung injury via inhibiting MAPK and NF-κB signaling in a murine model. Int. Immunopharmacol. 2016, 31, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Pang, M.; Dong, L.; Huang, X.; Wang, S.; Zhou, L. Anti-inflammatory and immunomodulatory effects of iridoid glycosides from Paederia scandens (LOUR.) MERRILL (Rubiaceae) on uric acid nephropathy rats. Life Sci. 2012, 91, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Li, B. Application of Aestin in the Preparation of Drugs for Rheumatoid Arthritis. Cn200510019323.6, 15 October 2008. (In Chinese). [Google Scholar]
- He, J.; Lu, X.; Wei, T.; Dong, Y.; Cai, Z.; Tang, L.; Liu, M. Asperuloside and Asperulosidic Acid Exert an Anti-Inflammatory Effect via Suppression of the NF-κB and MAPK Signaling Pathways in LPS-Induced RAW 264.7 Macrophages. Int. J. Mol. Sci. 2018, 19, 2027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, T.; Nakashima, T. Recent advances in osteoclast biology. Histochem. Cell Biol. 2018, 149, 325–341. [Google Scholar] [CrossRef]
- Nakashima, T.; Hayashi, M.; Takayanagi, H. New insights into osteoclastogenic signaling mechanisms. Trends Endocrinol. Metab. 2012, 23, 582–590. [Google Scholar] [CrossRef]
- Xianyuan, L.; Wei, Z.; Yaqian, D.; Dan, Z.; Xueli, T.; Zhanglu, D.; Guanyi, L.; Lan, T.; Menghua, L. Anti-renal fibrosis effect of asperulosidic acid via TGF-β1/smad2/smad3 and NF-κB signaling pathways in a rat model of unilateral ureteral obstruction. Phytomedicine 2019, 53, 274–285. [Google Scholar] [CrossRef]
- Tenuta, M.C.; Deguin, B.; Loizzo, M.R.; Dugay, A.; Acquaviva, R.; Malfa, G.A.; Bonesi, M.; Bouzidi, C.; Tundis, R. Contribution of Flavonoids and Iridoids to the Hypoglycaemic, Antioxidant, and Nitric Oxide (NO) Inhibitory Activities of Arbutus unedo L. Antioxidants 2020, 9, 184. [Google Scholar] [CrossRef] [Green Version]
- Yue, G.G.-L.; Lee, J.K.-M.; Chan, B.C.-L.; Kwok, H.-F.; Hoi, S.W.-H.; Sze, D.M.-Y.; Fung, K.-P.; Leung, P.-C.; Lau, C.B.-S. An innovative anti-cancer Chinese herbal formula exhibited multi-targeted efficacies in metastatic breast cancer mouse model. Chin. Med. 2018, 13, 64. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Xin, P.; Wang, Y.; Zhou, X.; Wei, D.; Deng, C.; Sun, S. Iridoids and sfingolipids from Hedyotis diffusa. Fitoterapia 2018, 124, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, A.; Yokoyama, Y.; Tanaka, K.; Benegiamo, G.; Hirayama, A.; Zhu, Q.; Kitamura, N.; Sugizaki, T.; Morimoto, K.; Itoh, H.; et al. Asperuloside Improves Obesity and Type 2 Diabetes through Modulation of Gut Microbiota and Metabolic Signaling. iScience 2020, 23, 101522. [Google Scholar] [CrossRef] [PubMed]
- Hosoo, S.; Koyama, M.; Watanabe, A.; Ishida, R.; Hirata, T.; Yamaguchi, Y.; Yamasaki, H.; Wada, K.; Higashi, Y.; Nakamura, K. Preventive effect of Eucommia leaf extract on aortic media hypertrophy in Wistar-Kyoto rats fed a high-fat diet. Hypertens. Res. 2017, 40, 546–551. [Google Scholar] [CrossRef] [PubMed]
- Charalampakis, G.; Abrahamsson, I.; Carcuac, O.; Dahlén, G.; Berglundh, T. Microbiota in experimental periodontitis and peri-implantitis in dogs. Clin. Oral Implant. Res. 2014, 25, 1094–1098. [Google Scholar] [CrossRef]
- Pirih, F.Q.; Hiyari, S.; Barroso, A.D.V.; Jorge, A.C.; Perussolo, J.; Atti, E.; Tetradis, S.; Camargo, P.M. Ligature-induced peri-implantitis in mice. J. Periodontal Res. 2015, 50, 519–524. [Google Scholar] [CrossRef] [Green Version]
- NguyenVo, T.N.; Hao, J.; Chou, J.; Oshima, M.; Aoki, K.; Kuroda, S.; Kaboosaya, B.; Kasugai, S. Ligature induced peri-implantitis: Tissue destruction and inflammatory progression in a murine model. Clin. Oral Implant. Res. 2017, 28, 129–136. [Google Scholar] [CrossRef]
- Ghassib, I.; Chen, Z.; Zhu, J.; Wang, H.-L. Use of IL-1 β, IL-6, TNF-α, and MMP-8 biomarkers to distinguish peri-implant diseases: A systematic review and meta-analysis. Clin. Implant Dent. Relat. Res. 2019, 21, 190–207. [Google Scholar] [CrossRef]
- Renvert, S.; Widén, C.; Persson, R.G. Cytokine and microbial profiles in relation to the clinical outcome following treatment of peri-implantitis. Clin. Oral Implant. Res. 2017, 28, 1127–1132. [Google Scholar] [CrossRef]
- Zupan, J.; Jeras, M.; Marc, J. Osteoimmunology and the influence of pro-inflammatory cytokines on osteoclasts. Biochem. Med. 2013, 23, 43–63. [Google Scholar] [CrossRef]
- Marahleh, A.; Kitaura, H.; Ohori, F.; Kishikawa, A.; Ogawa, S.; Shen, W.-R.; Qi, J.; Noguchi, T.; Nara, Y.; Mizoguchi, I. TNF-α Directly Enhances Osteocyte RANKL Expression and Promotes Osteoclast Formation. Front. Immunol. 2019, 10, 2925. [Google Scholar] [CrossRef] [Green Version]
- Deng, S.; Hu, Y.; Zhou, J.; Wang, Y.; Wang, Y.; Li, S.; Huang, G.; Peng, C.; Hu, A.; Yu, Q.; et al. TLR4 mediates alveolar bone resorption in experimental peri-implantitis through regulation of CD45+ cell infiltration, RANKL/OPG ratio, and inflammatory cytokine production. J. Periodontol. 2020, 91, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Son, H.S.; Lee, J.; Lee, H.I.; Kim, N.; Jo, Y.-J.; Lee, G.-R.; Hong, S.-E.; Kwon, M.; Kim, N.Y.; Kim, H.J.; et al. Benzydamine inhibits osteoclast differentiation and bone resorption via down-regulation of interleukin-1 β expression. Acta Pharm. Sin. B 2020, 10, 462–474. [Google Scholar] [CrossRef] [PubMed]
- Takayanagi, H.; Kim, S.; Koga, T.; Nishina, H.; Isshiki, M.; Yoshida, H.; Saiura, A.; Isobe, M.; Yokochi, T.; Inoue, J.-I.; et al. Induction and Activation of the Transcription Factor NFATc1 (NFAT2) Integrate RANKL Signaling in Terminal Differentiation of Osteoclasts. Dev. Cell 2002, 3, 889–901. [Google Scholar] [CrossRef] [Green Version]
- Asagiri, M.; Sato, K.; Usami, T.; Ochi, S.; Nishina, H.; Yoshida, H.; Morita, I.; Wagner, E.F.; Mak, T.W.; Serfling, E.; et al. Autoamplification of NFATc1 expression determines its essential role in bone homeostasis. J. Exp. Med. 2005, 202, 1261–1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.-Y.; Cheon, Y.-H.; Kwak, S.C.; Baek, J.M.; Kim, Y.-C.; Yoon, K.-H.; Oh, J.; Lee, M.S. 9-Hydroxy-6,7-dimethoxydalbergiquinol inhibits osteoclast differentiation through down-regulation of Akt, c-Fos and NFATc1. Int. Immunopharmacol. 2014, 20, 213–220. [Google Scholar] [CrossRef]
- Yagi, M.; Miyamoto, T.; Sawatani, Y.; Iwamoto, K.; Hosogane, N.; Fujita, N.; Morita, K.; Ninomiya, K.; Suzuki, T.; Miyamoto, K.; et al. DC-STAMP is essential for cell–cell fusion in osteoclasts and foreign body giant cells. J. Exp. Med. 2005, 202, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Lee, S.-H.; Ha Kim, J.; Choi, Y.; Kim, N. NFATc1 Induces Osteoclast Fusion Via Up-Regulation of Atp6v0d2 and the Dendritic Cell-Specific Transmembrane Protein (DC-STAMP). Mol. Endocrinol. 2008, 22, 176–185. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, T.; Katagiri, H.; Kanegae, Y.; Takayanagi, H.; Sawada, Y.; Yamamoto, A.; Pando, M.P.; Asano, T.; Verma, I.M.; Oda, H.; et al. Reciprocal role of ERK and NF-kappaB pathways in survival and activation of osteoclasts. J. Cell Biol. 2000, 148, 333–342. [Google Scholar] [CrossRef]
- Nakamura, H.; Hirata, A.; Tsuji, T.; Yamamoto, T. Role of Osteoclast Extracellular Signal-Regulated Kinase (ERK) in Cell Survival and Maintenance of Cell Polarity. J. Bone Miner. Res. 2003, 18, 1198–1205. [Google Scholar] [CrossRef]
- Lee, M.S.; Kim, H.S.; Yeon, J.-T.; Choi, S.-W.; Chun, C.H.; Kwak, H.B.; Oh, J. GM-CSF Regulates Fusion of Mononuclear Osteoclasts into Bone-Resorbing Osteoclasts by Activating the Ras/ERK Pathway. J. Immunol. 2009, 183, 3390–3399. [Google Scholar] [CrossRef] [Green Version]
- Feng, Q.; Yang, P.; Wang, H.; Li, C.; Hasegawa, T.; Liu, Z.; Li, M. ID09, A Newly-Designed Tubulin Inhibitor, Regulating the Proliferation, Migration, EMT Process and Apoptosis of Oral Squamous Cell Carcinoma. Int. J. Biol. Sci. 2022, 18, 473–490. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Sequences 5′–3′ | Reverse Sequences 5′–3′ |
|---|---|---|
| NFATc1 | TCCGTCCATACTGCCAGAGGTG | GGATTGTGCTGGAGAGGTCGTTAC |
| c-FOS | CCTTCACCCTGCCTCTTCTCAATG | AGCCTTCAGCTCCATGTTGCTAATG |
| DC-STAMP | GCCTCTTCCTGAAGCGATTCCTG | GGCACCTCTCCTCTTCATCAAACAG |
| Atp6v0d2 | AGCCAGCCTCCTAACTCAGCAG | GAGCCAGGAAGTTGCCATAGTCAG |
| Cathepsin K | TGGCTGTGGAGGCGGCTATATG | CGGGTAAGCGTCTTCAGAGTCAATG |
| MMP-9 | CCCATGTCACTTTCCCTTCACCTTC | CGATAACCATCCGAGCGACCTTTAG |
| β-actin | AGGAGTACGATGAGTCCGGC | CGCAGCTCAGTAACAGTCCG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Chen, X.; Zhang, Z.; Chen, J.; Ge, Z.; Huang, S.; Wei, H.; Li, D. Asperuloside Prevents Peri-Implantitis via Suppression of NF-κB and ERK1/2 on Rats. Pharmaceuticals 2022, 15, 1027. https://doi.org/10.3390/ph15081027
Wang X, Chen X, Zhang Z, Chen J, Ge Z, Huang S, Wei H, Li D. Asperuloside Prevents Peri-Implantitis via Suppression of NF-κB and ERK1/2 on Rats. Pharmaceuticals. 2022; 15(8):1027. https://doi.org/10.3390/ph15081027
Chicago/Turabian StyleWang, Xinge, Xutao Chen, Zhaoxin Zhang, Ji Chen, Zeyang Ge, Shitou Huang, Hongbo Wei, and Dehua Li. 2022. "Asperuloside Prevents Peri-Implantitis via Suppression of NF-κB and ERK1/2 on Rats" Pharmaceuticals 15, no. 8: 1027. https://doi.org/10.3390/ph15081027
APA StyleWang, X., Chen, X., Zhang, Z., Chen, J., Ge, Z., Huang, S., Wei, H., & Li, D. (2022). Asperuloside Prevents Peri-Implantitis via Suppression of NF-κB and ERK1/2 on Rats. Pharmaceuticals, 15(8), 1027. https://doi.org/10.3390/ph15081027