

Structural Characterization and Anti-Nonalcoholic Fatty Liver Effect of High-Sulfated Ulva pertusa Polysaccharide

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Chemical Components of U and HU
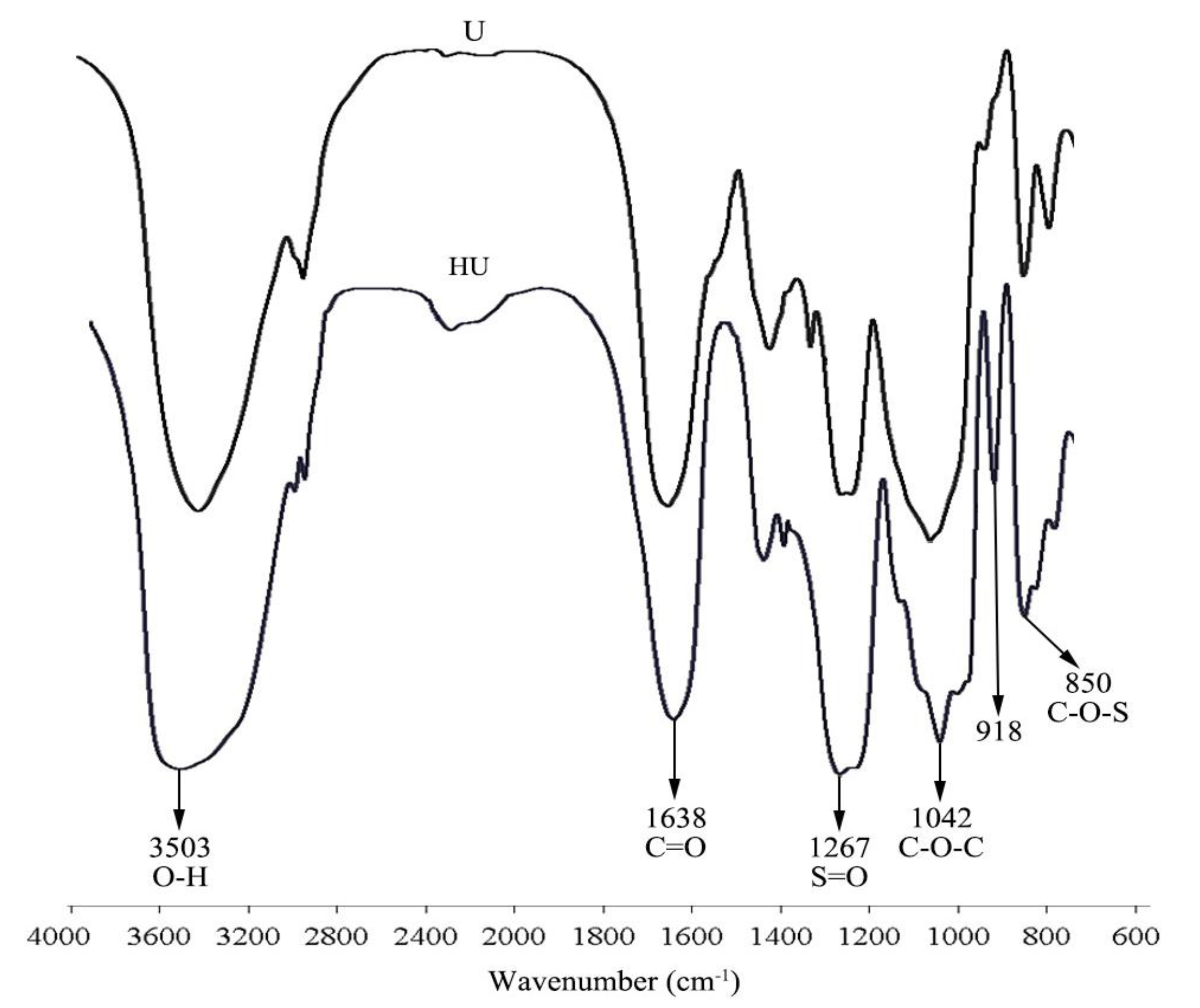
2.2. FT-IR Spectrum Analysis of U and HU
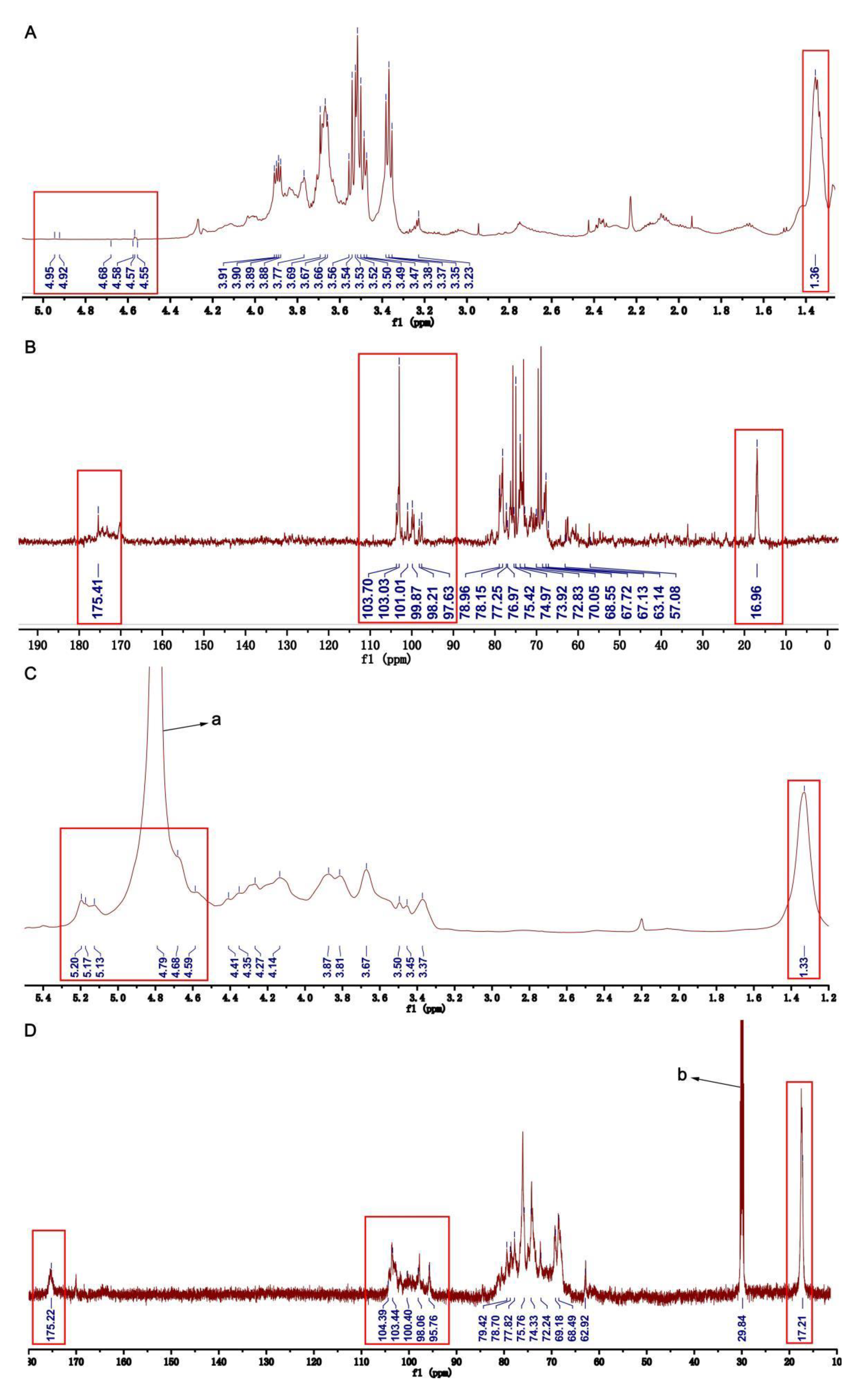
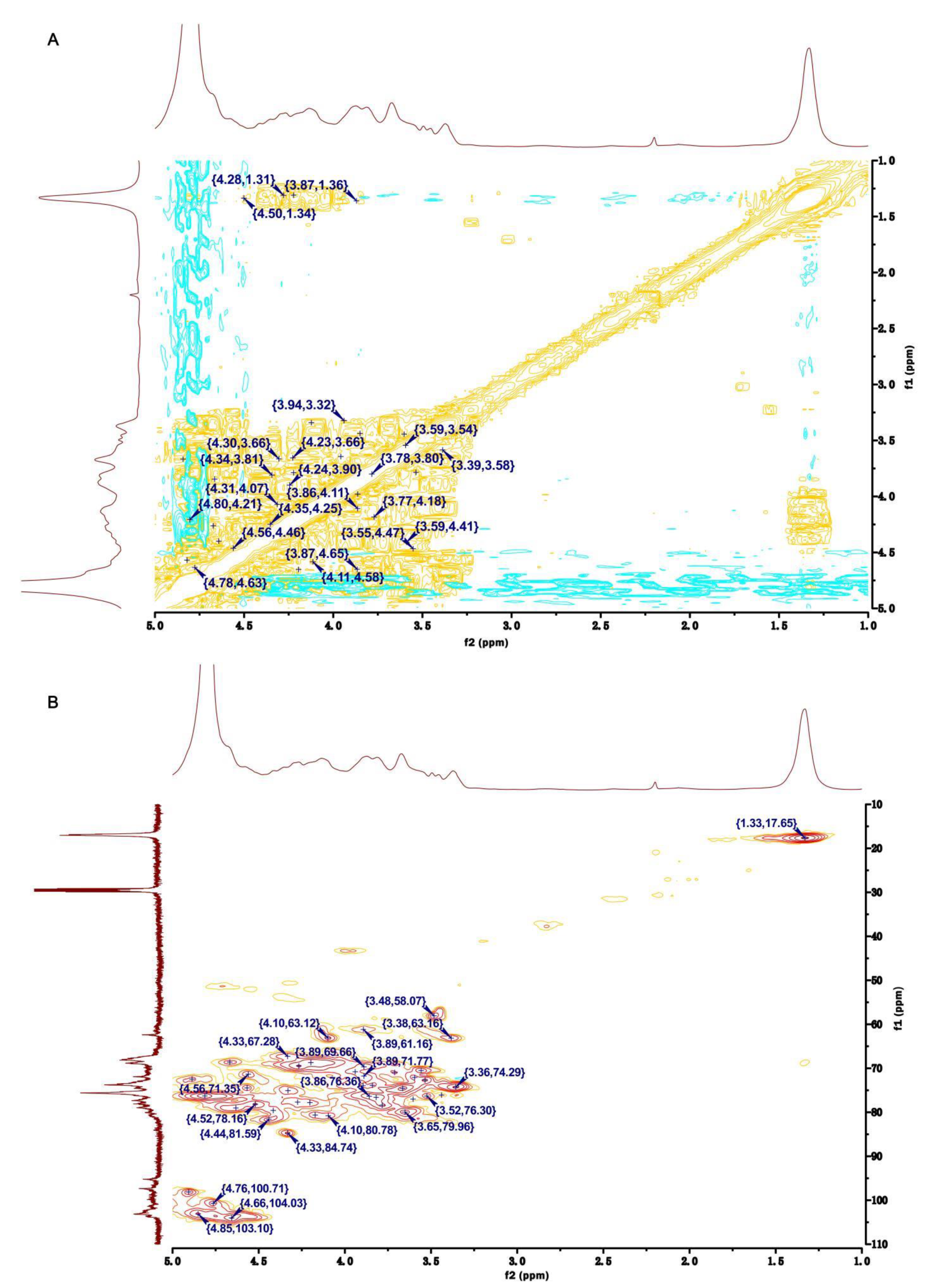
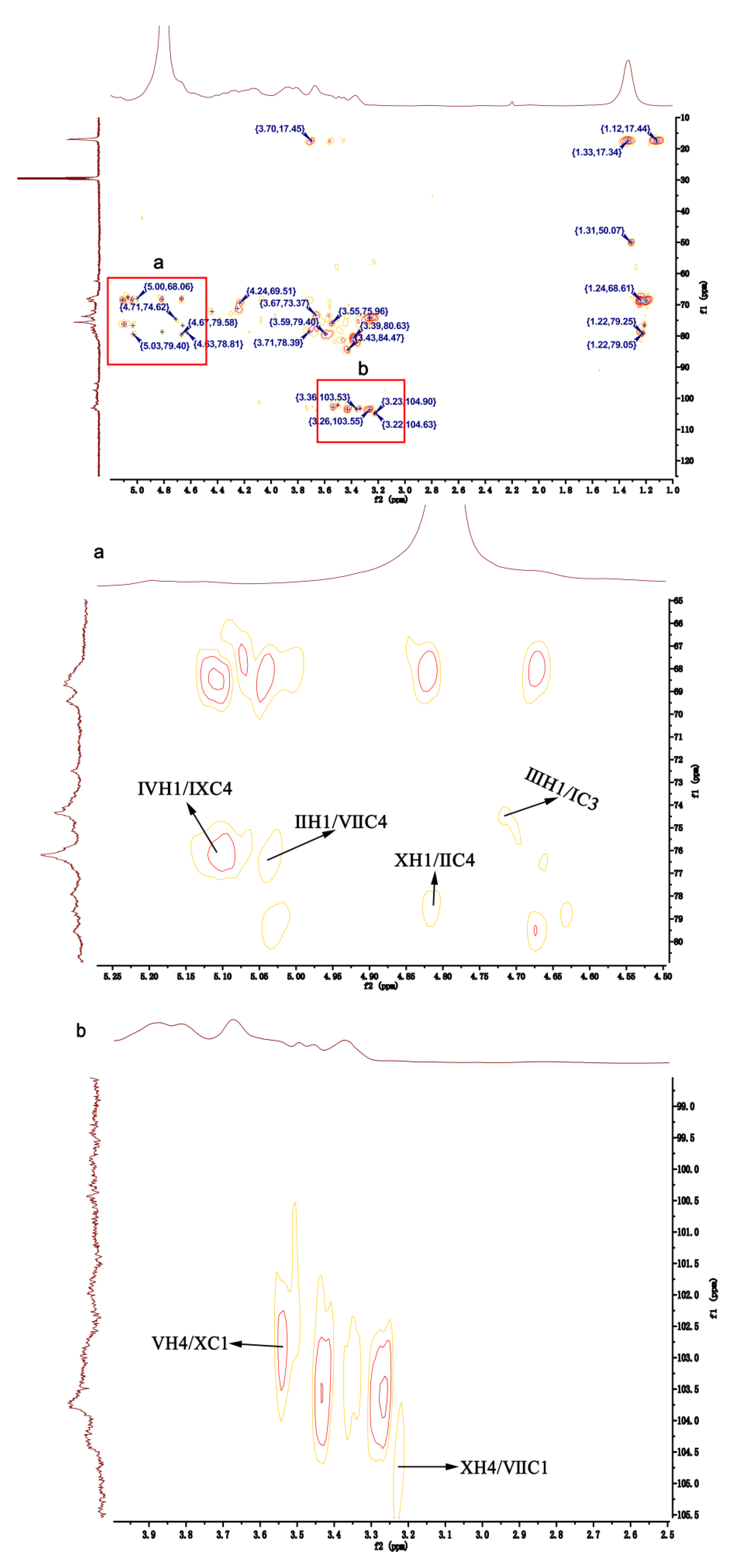
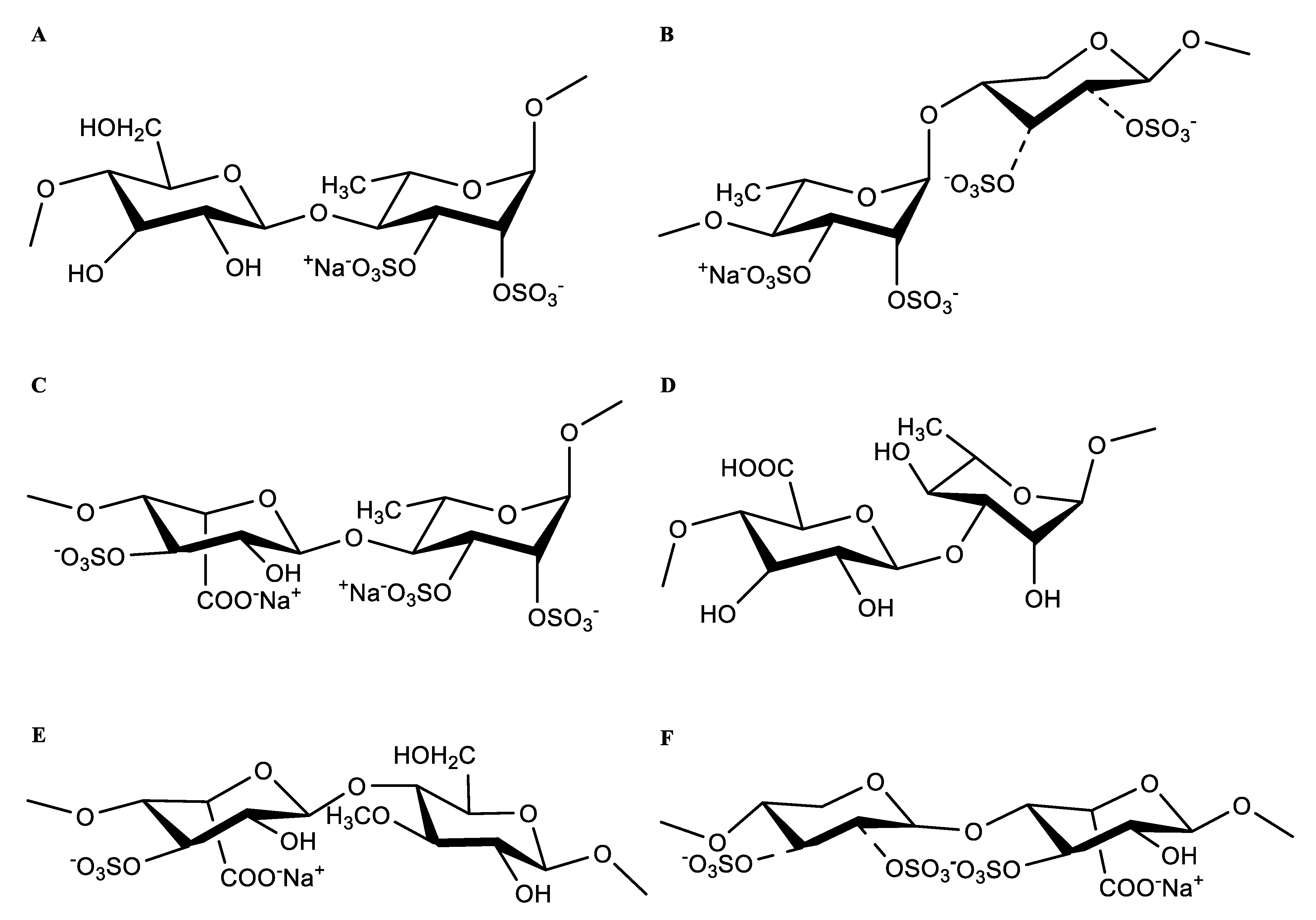
2.3. NMR Analyses of HU and U
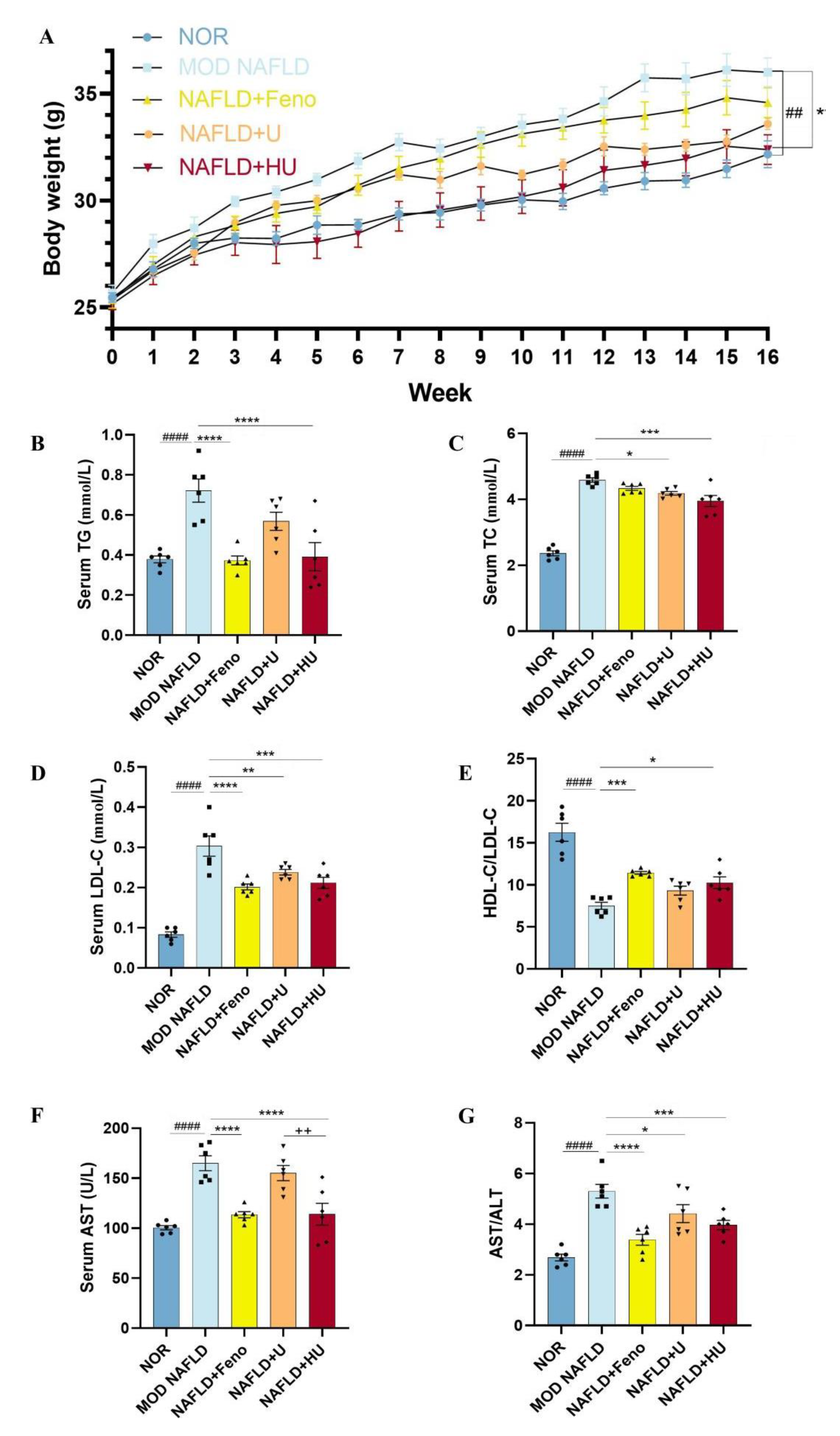
2.4. Effect of HU on Body Weight in Mice with NAFLD
2.5. Effect of HU on the Abnormal Changes in Lipid Metabolism Indicators Caused by NAFLD
2.6. Effect of HU on Liver Function in NAFLD Mice
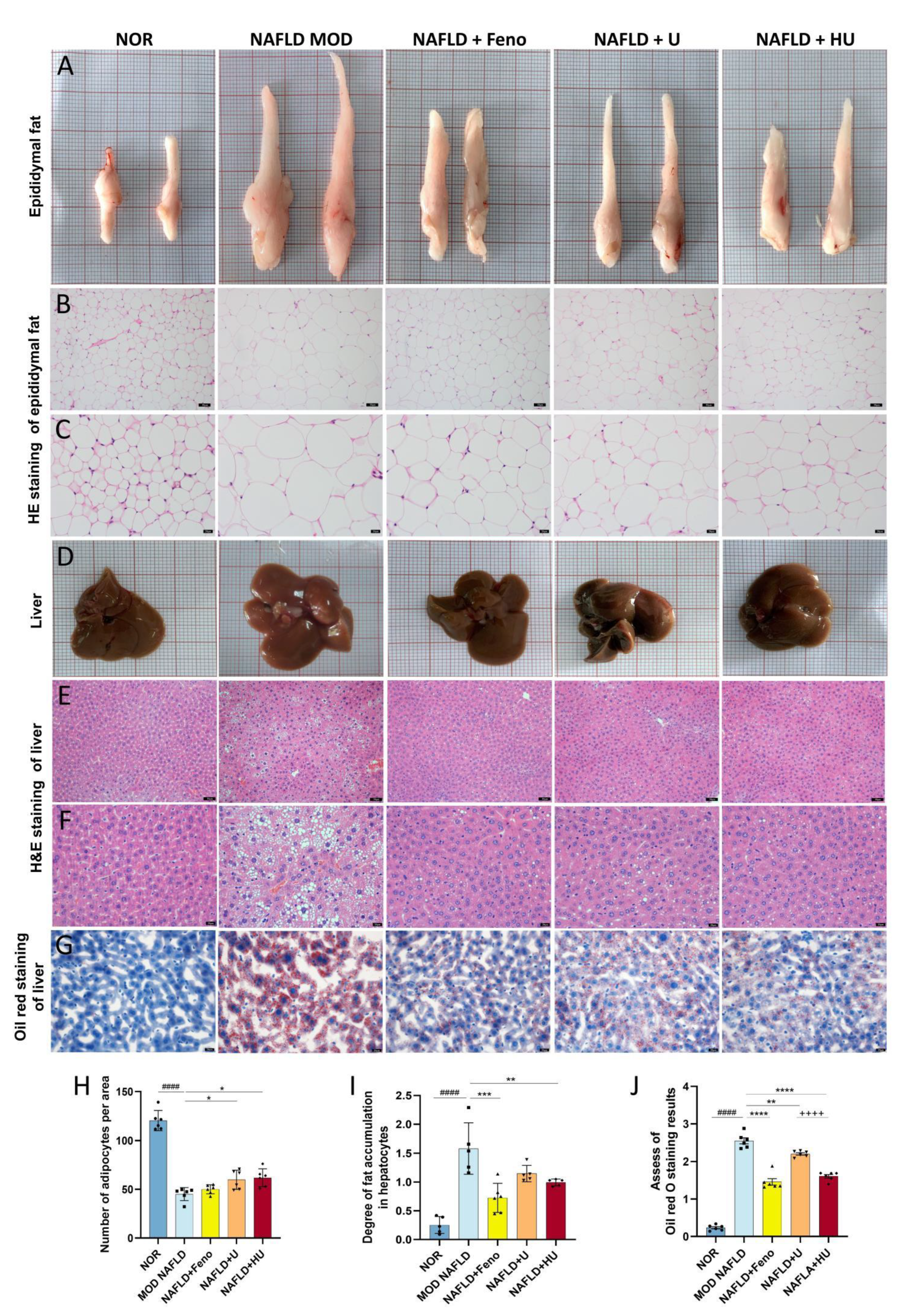
2.7. Effect of HU on Epididymal Adipose Tissue Changes Associated with NAFLD
2.8. Effect of HU on Liver Damage Caused by NAFLD
3. Materials and Methods
3.1. Materials and Reagents
3.2. Isolation of U
3.3. Preparation of HU
3.4. Chemical Analysis of U and HU
3.5. Determination of Molecular Weights of U and HU
3.6. Analysis of Monosaccharide Compositions of U and HU
3.7. Analysis of FT-IR and NMR Spectrum of U and HU
3.8. Animal Models and Experimental Design
3.9. Biochemical Analysis of Blood Samples
3.10. Histopathologic Observations
3.11. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Han, H.B.; Yan, L.; Song, W.; Wang, Z.L.; Zhang, X.L. Complete chloroplast genome of Ulva pertusa, one of the causal species of green macroalgal blooms in the coastal waters of Qinhuangdao, China. Mitochondrial DNA B Resour. 2020, 5, 1084–1086. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Wang, Z.; Song, H.; Fan, S.; Yuan, C.; Fu, M.; Miao, X.; Zhang, X.; Su, R.; Hu, C. An anomalous bi-macroalgal bloom caused by Ulva and Sargassum seaweeds during spring to summer of 2017 in the western Yellow Sea, China. Harmful Algae 2020, 93, 101760. [Google Scholar] [CrossRef] [PubMed]
- Le, B.; Golokhvast, K.S.; Yang, S.H.; Sun, S. Optimization of Microwave-Assisted Extraction of Polysaccharides from Ulva pertusa and Evaluation of Their Antioxidant Activity. Antioxidants 2019, 8, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Wu, Y.; Li, G.; Li, M.; Yan, R.; Xu, Z.; Lei, H.; Sun, Y.; Duan, X.; Hu, L.; et al. Structural characterization and transcript-metabolite correlation network of immunostimulatory effects of sulfated polysaccharides from green alga Ulva pertusa. Food Chem. 2021, 342, 128537. [Google Scholar] [CrossRef] [PubMed]
- Pengzhan, Y.; Ning, L.; Xiguang, L.; Gefei, Z.; Quanbin, Z.; Pengcheng, L. Antihyperlipidemic effects of different molecular weight sulfated polysaccharides from Ulva pertusa (Chlorophyta). Pharmacol. Res. 2003, 48, 543–549. [Google Scholar] [CrossRef]
- Chi, Y.; Zhang, M.; Wang, X.; Fu, X.; Guan, H.; Wang, P. Ulvan lyase assisted structural characterization of ulvan from Ulva pertusa and its antiviral activity against vesicular stomatitis virus. Int. J. Biol. Macromol. 2020, 157, 75–82. [Google Scholar] [CrossRef]
- Shi, J.; Cheng, C.; Zhao, H.; Jing, J.; Gong, N.; Lu, W. In vivo anti-radiation activities of the Ulva pertusa polysaccharides and polysaccharide-iron (III) complex. Int. J. Biol. Macromol. 2013, 60, 341–346. [Google Scholar] [CrossRef]
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease-Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, M.I.; Khan, M.U.; Kodali, S.; Shetty, A.; Bell, S.M.; Victor, D. Hepatocellular Carcinoma Due to Nonalcoholic Fatty Liver Disease: Current Concepts and Future Challenges. J. Hepatocell. Carcinoma. 2022, 9, 477–496. [Google Scholar] [CrossRef]
- Younossi, Z.M.; Corey, K.E.; Lim, J.K. AGA Clinical Practice Update on Lifestyle Modification Using Diet and Exercise to Achieve Weight Loss in the Management of Nonalcoholic Fatty Liver Disease: Expert Review. Gastroenterology 2021, 160, 912–918. [Google Scholar] [CrossRef]
- Schattenberg, J.M. Emerging Pharmacological Treatment in Nonalcoholic Steatohepatitis. Visc. Med. 2020, 36, 411–416. [Google Scholar] [CrossRef]
- Nair, D.G.; Weiskirchen, R.; Al-Musharafi, S.K. The use of marine-derived bioactive compounds as potential hepatoprotective agents. Acta. Pharmacol. Sin. 2015, 36, 158–170. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yang, L.; Zhao, N.; Hong, Z.; Cai, B.; Le, Q.; Yang, T.; Shi, L.; He, J. Soluble Polysaccharide Derived from Laminaria japonica Attenuates Obesity-Related Nonalcoholic Fatty Liver Disease Associated with Gut Microbiota Regulation. Mar. Drugs 2021, 19, 699. [Google Scholar] [CrossRef]
- Ren, R.; Yang, Z.; Zhao, A.; Huang, Y.; Lin, S.; Gong, J.; Chen, J.; Zhu, P.; Huang, F.; Lin, W. Sulfated polysaccharide from Enteromorpha prolifera increases hydrogen sulfide production and attenuates non-alcoholic fatty liver disease in high-fat diet rats. Food Funct. 2018, 9, 4376–4383. [Google Scholar] [CrossRef]
- Liu, X.; Chen, X.; Xie, L.; Xie, J.; Shen, M. Sulfated Chinese yam polysaccharide enhances the immunomodulatory activity of RAW 264.7 cells via the TLR4-MAPK/NF-κB signaling pathway. Food Funct. 2022, 13, 1316–1326. [Google Scholar] [CrossRef]
- Deng, Z.; Wu, N.; Wang, J.; Geng, L.; Yue, Y.; Wang, F.; Zhang, Q. Low molecular weight fucoidan fraction LF2 improves metabolic syndrome via up-regulating PI3K-AKT-mTOR axis and increasing the abundance of Akkermansia muciniphila in the gut microbiota. Int. J. Biol. Macromol. 2021, 193, 789–798. [Google Scholar] [CrossRef]
- Qi, H.; Huang, L.Y.; Liu, X.L.; Liu, D.M.; Zhang, Q.B.; Liu, S.M. Antihyperlipidemic activity of high sulfate content derivative of polysaccharide extracted from Ulva pertusa (Chlorophyta). Carbohydr. Polym. 2012, 87, 1637–1640. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, Y.; Ai, C.; Tian, W.; Wen, C.; Song, S.; Zhu, B. An acidic polysaccharide from Patinopecten yessoensis skirt prevents obesity and improves gut microbiota and metabolism of mice induced by high-fat diet. Food Res. Int. 2022, 154, 110980. [Google Scholar] [CrossRef]
- Cui, Y.; Liu, X.; Li, S.; Hao, L.; Du, J.; Gao, D.; Kang, Q.; Lu, J. Extraction, characterization and biological activity of sulfated polysaccharides from seaweed Dictyopteris divaricata. Int. J. Biol. Macromol. 2018, 117, 256–263. [Google Scholar] [CrossRef]
- Yang, Q.; Jiang, Y.; Fu, S.; Shen, Z.; Zong, W.; Xia, Z.; Zhan, Z.; Jiang, X. Protective Effects of Ulva lactuca Polysaccharide Extract on Oxidative Stress and Kidney Injury Induced by D-Galactose in Mice. Mar. Drugs 2021, 19, 539. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhang, S.; Wang, Q.; Lu, X.; Lin, L.; Tian, Y.; Xiao, J.; Zheng, B. Characterization and hypoglycemic activity of a β-pyran polysaccharides from bamboo shoot (Leleba oldhami Nakal) shells. Carbohydr. Polym. 2016, 144, 438–446. [Google Scholar] [CrossRef] [PubMed]
- Olawuyi, I.F.; Lee, W.Y. Structural characterization, functional properties and antioxidant activities of polysaccharide extract obtained from okra leaves (Abelmoschus esculentus). Food Chem. 2021, 354, 129437. [Google Scholar] [CrossRef] [PubMed]
- Halaj, M.; Matulová, M.; Šutovská, M.; Barboríková, J.; Kazimierová, I.; Fraňová, S.; Přibyl, P.; Cepák, V.; Lukavský, J.; Capek, P. Chemico-physical and pharmacodynamic properties of extracellular Dictyosphaerium chlorelloides biopolymer. Carbohydr. Polym. 2018, 198, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Tu, J.Q.; Liu, H.P.; Wen, Y.H.; Chen, P.; Liu, Z.T. A novel polysaccharide from Hericium erinaceus: Preparation, structural characteristics, thermal stabilities, and antioxidant activities in vitro. J. Food Biochem. 2021, 45, e13871. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.Y.; Wang, J.Q.; Yin, J.Y.; Nie, S.P.; Xie, M.Y. A review of NMR analysis in polysaccharide structure and conformation: Progress, challenge and perspective. Food. Res. Int. 2021, 143, 110290. [Google Scholar] [CrossRef]
- de Carvalho, M.M.; de Freitas, R.A.; Ducatti, D.R.B.; Ferreira, L.G.; Gonçalves, A.G.; Colodi, F.G.; Mazepa, E.; Aranha, E.M.; Noseda, M.D.; Duarte, M.E.R. Modification of ulvans via periodate-chlorite oxidation: Chemical characterization and anticoagulant activity. Carbohydr. Polym. 2018, 197, 631–640. [Google Scholar] [CrossRef]
- Guidara, M.; Yaich, H.; Amor, I.B.; Fakhfakh, J.; Gargouri, J.; Lassoued, S.; Blecker, C.; Richel, A.; Attia, H.; Garna, H. Effect of extraction procedures on the chemical structure, antitumor and anticoagulant properties of ulvan from Ulva lactuca of Tunisia coast. Carbohydr. Polym. 2021, 253, 117283. [Google Scholar] [CrossRef]
- Wang, S.; Wang, W.; Hou, L.; Qin, L.; He, M.; Li, W.; Mao, W. A sulfated glucuronorhamnan from the green seaweed Monostroma nitidum: Characteristics of its structure and antiviral activity. Carbohydr. Polym. 2020, 227, 115280. [Google Scholar] [CrossRef]
- Glasson, C.R.K.; Luiten, C.A.; Carnachan, S.M.; Daines, A.M.; Kidgell, J.T.; Hinkley, S.F.R.; Praeger, C.; Andrade Martinez, M.; Sargison, L.; Magnusson, M.; et al. Structural characterization of ulvans extracted from blade (Ulva ohnoi) and filamentous (Ulva tepida and Ulva prolifera) species of cultivated Ulva. Int. J. Biol. Macromol. 2022, 194, 571–579. [Google Scholar] [CrossRef]
- Liu, W.; Li, W.; Sui, Y.; Li, X.Q.; Liu, C.; Jing, H.; Zhang, H.; Cao, W. Structure characterization and anti-leukemia activity of a novel polysaccharide from Angelica sinensis (Oliv.) Diels. Int. J. Biol. Macromol. 2019, 121, 161–172. [Google Scholar] [CrossRef]
- Gao, X.; Qu, H.; Shan, S.; Song, C.; Baranenko, D.; Li, Y.; Lu, W. A novel polysaccharide isolated from Ulva Pertusa: Structure and physicochemical property. Carbohydr. Polym. 2020, 233, 115849. [Google Scholar] [CrossRef]
- Patova, O.A.; Luаnda, A.; Paderin, N.M.; Popov, S.V.; Makangara, J.J.; Kuznetsov, S.P.; Kalmykova, E.N. Xylogalacturonan-enriched pectin from the fruit pulp of Adansonia digitata: Structural characterization and antidepressant-like effect. Carbohydr. Polym. 2021, 262, 117946. [Google Scholar] [CrossRef]
- Juanola, O.; Martínez-López, S.; Francés, R.; Gómez-Hurtado, I. Non-Alcoholic Fatty Liver Disease: Metabolic, Genetic, Epigenetic and Environmental Risk Factors. Int. J. Environ. Res. Public Health. 2021, 18, 5227. [Google Scholar] [CrossRef]
- Simbrunner, B.; Trauner, M.; Reiberger, T. Review article: Therapeutic aspects of bile acid signalling in the gut-liver axis. Aliment. Pharmacol. Ther. 2021, 54, 1243–1262. [Google Scholar] [CrossRef]
- Staley, C.; Weingarden, A.R.; Khoruts, A.; Sadowsky, M.J. Interaction of gut microbiota with bile acid metabolism and its influence on disease states. Appl. Microbiol. Biotechnol. 2017, 101, 47–64. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Liu, X.; Liu, D.; Yin, Z.; Yang, X.; Zeng, M. Oyster (Crassostrea gigas) Polysaccharide Ameliorates High-Fat-Diet-Induced Oxidative Stress and Inflammation in the Liver via the Bile Acid-FXR-AMPKα Pathway. J. Agric. Food Chem. 2022, 70, 8662–8671. [Google Scholar] [CrossRef]
- Gao, J.; Lin, L.; Sun, B.; Zhao, M. A comparison study on polysaccharides extracted from Laminaria japonica using different methods: Structural characterization and bile acid-binding capacity. Food Funct. 2017, 8, 3043–3052. [Google Scholar] [CrossRef]
- Bennick, R.A.; Nagengast, A.A.; DiAngelo, J.R. The SR proteins SF2 and RBP1 regulate triglyceride storage in the fat body of Drosophila. Biochem. Biophys. Res. Commun. 2019, 516, 928–933. [Google Scholar] [CrossRef]
- Li, Q.; Jiang, S.; Shi, W.; Qi, X.; Song, W.; Mou, J.; Yang, J. Structure characterization, antioxidant and immunoregulatory properties of a novel fucoidan from the sea cucumber Stichopus chloronotus. Carbohydr. Polym. 2020, 231, 115767. [Google Scholar] [CrossRef]
- Mou, J.; Zhuang, D.; Li, Q.; Song, W.; Yang, J. Comparison of chain conformation properties of bio-active fucosylated chondroitin sulfates from two different sea cucumbers. Int. J. Biol. Macromol. 2019, 133, 44–50. [Google Scholar] [CrossRef]
- Arlov, Ø.; Aachmann, F.L.; Feyzi, E.; Sundan, A.; Skjåk-Bræk, G. The Impact of Chain Length and Flexibility in the Interaction between Sulfated Alginates and HGF and FGF-2. Biomacromolecules 2015, 16, 3417–3424. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Wu, W.; Xu, X.; Zhang, L.; Liu, Y.; Wang, K. Chain conformation and anti-tumor activity of derivatives of polysaccharide from Rhizoma Panacis Japonici. Carbohydr. Polym. 2014, 105, 308–316. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Xu, Y.; Chen, H.; Sun, P. Extraction, Structural Characterization, and Potential Antioxidant Activity of the Polysaccharides from Four Seaweeds. Int. J. Mol. Sci. 2016, 17, 1988. [Google Scholar] [CrossRef] [PubMed]
- Jia, R.B.; Wu, J.; Li, Z.R.; Ou, Z.R.; Lin, L.; Sun, B.; Zhao, M. Structural characterization of polysaccharides from three seaweed species and their hypoglycemic and hypolipidemic activities in type 2 diabetic rats. Int. J. Biol. Macromol. 2020, 155, 1040–1049. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Zhu, L.; Li, X.; Jia, J.; Zhang, Y.; Sun, X.; Ma, J.; Liu, Z.; Ma, X. Low-molecular weight fucoidan inhibits the differentiation of osteoclasts and reduces osteoporosis in ovariectomized rats. Mol. Med. Rep. 2017, 15, 890–898. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Liu, T.; Wang, Z.; Xu, Y.; Zhang, Q.; Luo, D. Low molecular weight fucoidan attenuates liver injury via SIRT1/AMPK/PGC1α axis in db/db mice. Int. J. Biol. Macromol. 2018, 112, 929–936. [Google Scholar] [CrossRef]
- O’Hara, A.M.; Shanahan, F. The gut flora as a forgotten organ. EMBO Rep. 2006, 7, 688–693. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.; Wang, X.; Li, L. Human gut microbiome: The second genome of human body. Protein Cell 2010, 1, 718–725. [Google Scholar] [CrossRef] [Green Version]
- Cheong, K.L.; Yu, B.; Chen, J.; Zhong, S. A Comprehensive Review of the Cardioprotective Effect of Marine Algae Polysaccharide on the Gut Microbiota. Foods 2022, 11, 3550. [Google Scholar] [CrossRef]
- Shannon, E.; Conlon, M.; Hayes, M. Seaweed Components as Potential Modulators of the Gut Microbiota. Mar. Drugs 2021, 19, 358. [Google Scholar] [CrossRef]
- Wang, X.; Wang, X.; Jiang, H.; Cai, C.; Li, G.; Hao, J.; Yu, G. Marine polysaccharides attenuate metabolic syndrome by fermentation products and altering gut microbiota: An overview. Carbohydr. Polym. 2018, 195, 601–612. [Google Scholar] [CrossRef]
- Zhang, Y.; Zuo, J.; Yan, L.; Cheng, Y.; Li, Q.; Wu, S.; Chen, L.; Thring, R.W.; Yang, Y.; Gao, Y.; et al. Sargassum fusiforme Fucoidan Alleviates High-Fat Diet-Induced Obesity and Insulin Resistance Associated with the Improvement of Hepatic Oxidative Stress and Gut Microbiota Profile. J. Agric. Food Chem. 2020, 68, 10626–10638. [Google Scholar] [CrossRef]
- Oh, J.K.; Vasquez, R.; Kim, S.H.; Lee, J.H.; Kim, E.J.; Hong, S.K.; Kang, D.K. Neoagaro-Oligosaccharides modulate gut microbiota and alleviate body weight gain and metabolic syndrome in high-fat diet-induced obese rats. J. Funct. Foods 2022, 88, 104869. [Google Scholar] [CrossRef]
- Yue, C.; Li, M.; Li, J.; Han, X.; Zhu, H.; Yu, G.; Cheng, J. Medium-, long- and medium-chain-type structured lipids ameliorate high-fat diet-induced atherosclerosis by regulating inflammation, adipogenesis, and gut microbiota in ApoE(−/−) mice. Food Funct. 2020, 11, 5142–5155. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, Q.; Ma, W.; Tian, F.; Shen, H.; Zhou, M. A combination of quercetin and resveratrol reduces obesity in high-fat diet-fed rats by modulation of gut microbiota. Food Funct. 2017, 8, 4644–4656. [Google Scholar] [CrossRef]
- Pung, H.C.; Lin, W.S.; Lo, Y.C.; Hsu, C.C.; Ho, C.T.; Pan, M.H. Ulva prolifera polysaccharide exerts anti-obesity effects via upregulation of adiponectin expression and gut microbiota modulation in high-fat diet-fed C57BL/6 mice. J. Food Drug Anal. 2022, 30, 46–61. [Google Scholar] [CrossRef]
- Jia, B.; Zou, Y.; Han, X.; Bae, J.W.; Jeon, C.O. Gut microbiome-mediated mechanisms for reducing cholesterol levels: Implications for ameliorating cardiovascular disease. Trends. Microbiol. 2023, 01, 76–91. [Google Scholar] [CrossRef]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids from Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef] [Green Version]
- Canfora, E.E.; Meex, R.C.R.; Venema, K.; Blaak, E.E. Gut microbial metabolites in obesity, NAFLD and T2DM. Nat. Rev. Endocrinol. 2019, 15, 261–273. [Google Scholar] [CrossRef]
- Duparc, T.; Plovier, H.; Marrachelli, V.G.; Van, H.M.; Essaghir, A.; Ståhlman, M.; Matamoros, S.; Geurts, L.; Pardo-Tendero, M.M.; Druart, C.; et al. Hepatocyte MyD88 affects bile acids, gut microbiota and metabolome contributing to regulate glucose and lipid metabolism. Gut 2017, 66, 620–632. [Google Scholar] [CrossRef]
- Fu, X.; Cao, C.; Ren, B.; Zhang, B.; Huang, Q.; Li, C. Structural characterization and in vitro fermentation of a novel polysaccharide from Sargassum thunbergii and its impact on gut microbiota. Carbohydr. Polym. 2018, 183, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.X.; Chen, X.Q.; Cheong, K.L. Current trends in marine algae polysaccharides: The digestive tract, microbial catabolism, and prebiotic potential. Int. J. Biol. Macromol. 2020, 151, 344–354. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, S.; Goodspeed, L.; Wang, S.; Kim, J.; Zeng, L.; Ioannou, G.N.; Haigh, W.G.; Yeh, M.M.; Kowdley, K.V.; O’Brien, K.D.; et al. Dietary cholesterol exacerbates hepatic steatosis and inflammation in obese LDL receptor-deficient mice. J. Lipid Res. 2011, 52, 1626–1635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Den Besten, G.; Bleeker, A.; Gerding, A.; van Eunen, K.; Havinga, R.; van Dijk, T.H.; Oosterveer, M.H.; Jonker, J.W.; Groen, A.K.; Reijngoud, D.J.; et al. Short-Chain Fatty Acids Protect Against High-Fat Diet-Induced Obesity via a PPARγ-Dependent Switch from Lipogenesis to Fat Oxidation. Diabetes 2015, 64, 2398–2408. [Google Scholar] [CrossRef] [Green Version]
- Dubois, M.; Gilles, K.; Hamilton, J.K.; Rebers, P.A.; Smith, F. A colorimetric method for the determination of sugars. Nature 1951, 168, 167. [Google Scholar] [CrossRef]
- Kawai, Y.; Seno, N.; Anno, K. A modified method for chondrosulfatase assay. Anal. Biochem. 1969, 32, 314–321. [Google Scholar] [CrossRef]
- Wen, M.; Cui, Y.; Dong, C.X.; Zhang, L. Quantitative changes in monosaccharides of Keemun black tea and qualitative analysis of theaflavins-glucose adducts during processing. Food Res. Int. 2021, 148, 110588. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, H.; Li, X.; Wang, Q.; Yan, M.; Zhang, H.; Zhao, T.; Zhang, N.; Zhang, P.; Peng, L.; et al. Formononetin alleviates hepatic steatosis by facilitating TFEB-mediated lysosome biogenesis and lipophagy. J. Nutr. Biochem. 2019, 73, 108214. [Google Scholar] [CrossRef]
- Castera, L.; Friedrich-Rust, M.; Loomba, R. Noninvasive Assessment of Liver Disease in Patients with Nonalcoholic Fatty Liver Disease. Gastroenterology 2019, 156, 1264–1281.e4. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scheme | Total Sugar (%) | Sulfate (%) | Average Molecular Weights (kDa) | Monosaccharide Components (mol%) | |||||
|---|---|---|---|---|---|---|---|---|---|
| Rhap | Xylp | Glcp | GlcpA | Fucp | Manp | ||||
| U | 45.2 | 19.9 | 143.5 | 31.3 | 19.9 | 6.7 | 5.5 | 0.5 | -- |
| HU | 54.1 | 34.7 | 22.6 | 30.7 | 18.1 | 6.5 | 5.4 | 0.6 | 0.5 |
| Polysaccharides | Name | Sugar Residues | Chemical Shifts (ppm) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| H1/C1 | H2/C2 | H3/C3 | H4/C4 | H5/C5 | H6/C6 | -OMe | |||
| U | Ⅰ | -3)-α-L-Rhap(1- | 4.94/97.64 | 4.26/68.99 | 3.93/75.50 | 3.68/70.72 | 4.03/72.41 | 1.36/16.96 | |
| Ⅱ | -4)-α-L-Rhap3S(1- | 4.80/99.99 | 4.27/64.38 | 4.63/78.32 | 3.80/78.20 | 4.21/68.01 | 1.34/16.96 | ||
| Ⅲ | -4)-β-D-GlcpA(1- | 4.95/98.35 | 4.36/75.42 | 3.91/73.92 | 4.16/78.40 | 3.58/69.85 | --/175.41 | ||
| Ⅳ | -4)-β-D-Glcp(1- | 4.90/101.09 | 4.20/70.00 | 3.77/74.35 | 3.67/75.01 | 3.99/73.48 | 3.69/62.67 | ||
| Ⅴ | -4)-β-D-Glcp3Me(1- | 4.90/101.09 | 4.26/68.97 | 4.04/78.82 | 3.69/77.01 | 3.90/76.47 | 3.41/62.52 | 3.47/57.33 | |
| Ⅵ | -4)-β-D-Xylp(1- | 4.56/103.10 | 3.66/71.77 | 3.53/75.74 | 4.16/78.38 | 3.41/62.52 | --/-- | ||
| Ⅶ | -4)-α-L-IdopA(1- | 4.66/103.34 | 3.38/73.85 | 3.87/71.37 | 3.67/74.00 | 4.27/64.40 | --/175.41 | ||
| HU | Ⅰ | -3)-α-L-Rhap(1- | 5.19/95.99 | 4.19/69.73 | 3.78/75.34 | 3.84/73.86 | 3.53/72.70 | 1.33/17.21 | |
| Ⅱ | -4)-α-L-Rhap3S(1- | 4.91/98.13 | 4.07/66.53 | 4.33/75.08 | 3.65/79.96 | 4.26/69.66 | 1.33/17.21 | ||
| Ⅲ | -4)-β-D-GlcpA(1- | 4.76/100.71 | 4.63/78.97 | 3.89/71.77 | 4.10/80.78 | 4.52/78.16 | --/175.22 | ||
| Ⅳ | -4)-β-D-Glcp(1- | 5.11/98.13 | 4.63/79.02 | 4.42/79.58 | 3.60/77.00 | 3.44/76.09 | 3.89/61.16 | ||
| Ⅴ | -4)-β-D-Glcp3Me(1- | 5.11/98.13 | 4.57/74.46 | 4.44/81.59 | 3.52/76.32 | 3.59/72.09 | 3.38/63.16 | 3.48/58.07 | |
| Ⅵ | -4)-β-D-Xylp(1- | 4.66/104.03 | 3.89/73.95 | 4.20/77.80 | 3.80/76.58 | 3.71/70.99 | --/-- | ||
| Ⅶ | -4)-β-D-Xylp2,3S(1- | 4.66/104.03 | 4.27/77.69 | 4.33/84.74 | 3.85/76.36 | 3.89/69.66 | --/-- | ||
| Ⅷ | -4)-α-L-IdopA(1- | 4.85/103.10 | 3.66/74.59 | 3.94/70.79 | 3.36/74.17 | 4.10/63.12 | --/175.22 | ||
| Ⅸ | -4)-α-L-Rhap2,3S(1- | 5.15/100.24 | 4.67/68.66 | 4.17/80.62 | 4.81/76.29 | 4.56/71.35 | 1.33/17.21 | ||
| Ⅹ | -4)-α-L-IdopA3S(1- | 4.85/103.10 | 3.66/74.59 | 4.89/72.45 | 3.36/74.17 | 4.10/63.12 | --/175.22 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wan, Y.; Liu, L.; Zhang, B.; Wang, S.; Wang, X.; Chen, K.; Li, Y.; Zhao, T.; Qi, H. Structural Characterization and Anti-Nonalcoholic Fatty Liver Effect of High-Sulfated Ulva pertusa Polysaccharide. Pharmaceuticals 2023, 16, 62. https://doi.org/10.3390/ph16010062
Wan Y, Liu L, Zhang B, Wang S, Wang X, Chen K, Li Y, Zhao T, Qi H. Structural Characterization and Anti-Nonalcoholic Fatty Liver Effect of High-Sulfated Ulva pertusa Polysaccharide. Pharmaceuticals. 2023; 16(1):62. https://doi.org/10.3390/ph16010062
Chicago/Turabian StyleWan, Yuzhou, Lin Liu, Bo Zhang, Shaopeng Wang, Xiaoqian Wang, Kexu Chen, Yuxi Li, Tingting Zhao, and Huimin Qi. 2023. "Structural Characterization and Anti-Nonalcoholic Fatty Liver Effect of High-Sulfated Ulva pertusa Polysaccharide" Pharmaceuticals 16, no. 1: 62. https://doi.org/10.3390/ph16010062
APA StyleWan, Y., Liu, L., Zhang, B., Wang, S., Wang, X., Chen, K., Li, Y., Zhao, T., & Qi, H. (2023). Structural Characterization and Anti-Nonalcoholic Fatty Liver Effect of High-Sulfated Ulva pertusa Polysaccharide. Pharmaceuticals, 16(1), 62. https://doi.org/10.3390/ph16010062