


Sargassum horneri (Turner) C. Agardh Extract Regulates Neuroinflammation In Vitro and In Vivo
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Culture and Extraction of Sample
2.3. UPLC-HR-MS/MS Analysis
2.4. Cell Viability Assay
2.5. NO Assay
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Western Blot Analysis
2.8. In Vivo Experiment
2.9. Immunohistochemistry
2.10. Statistical Analysis
3. Results
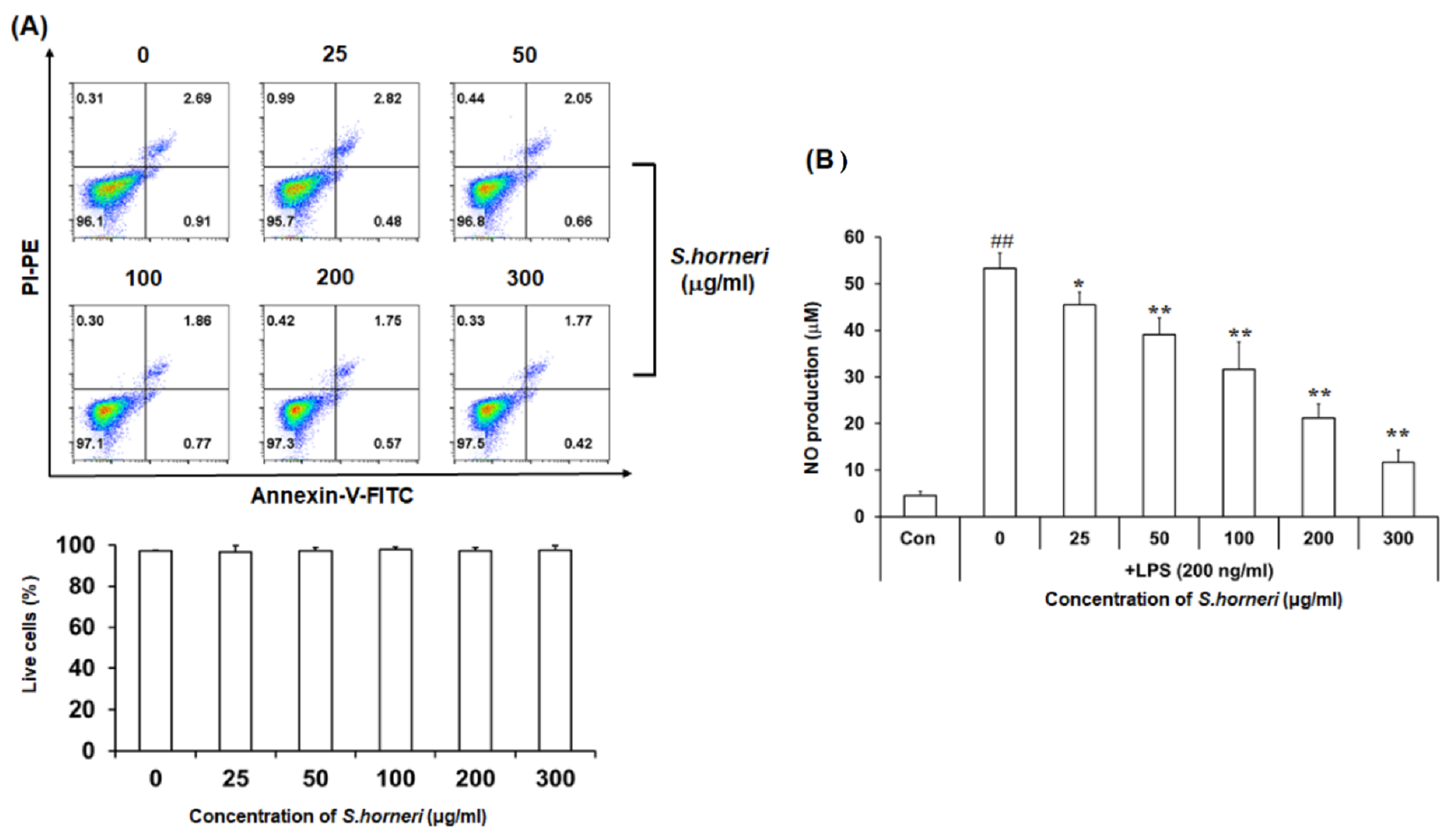
3.1. S. horneri Extract Inhibits NO Production in LPS-Stimulated BV-2 Microglial Cells
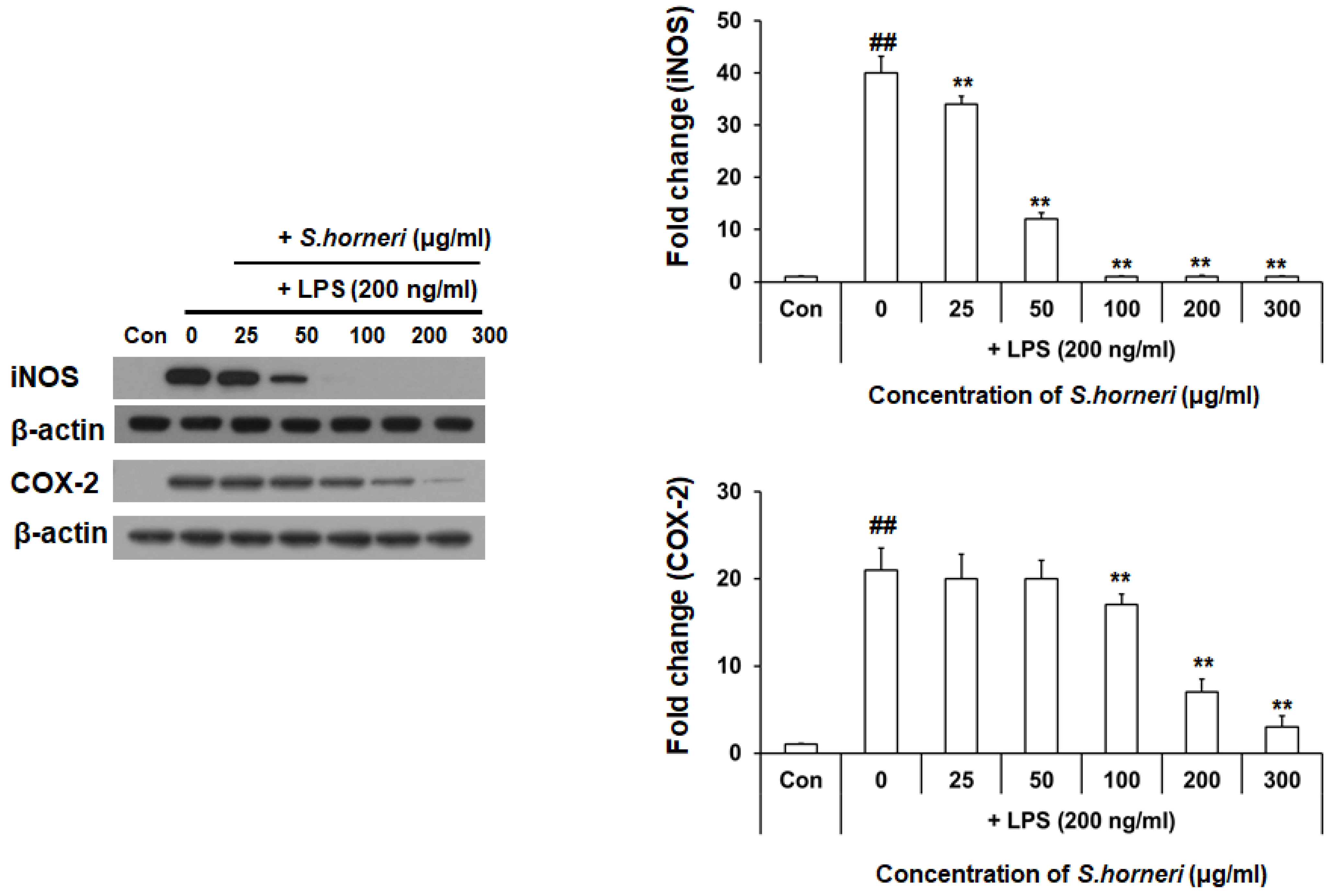
3.2. S. horneri Extract Reduced LPS-Induced iNOS and COX-2 Protein Expression
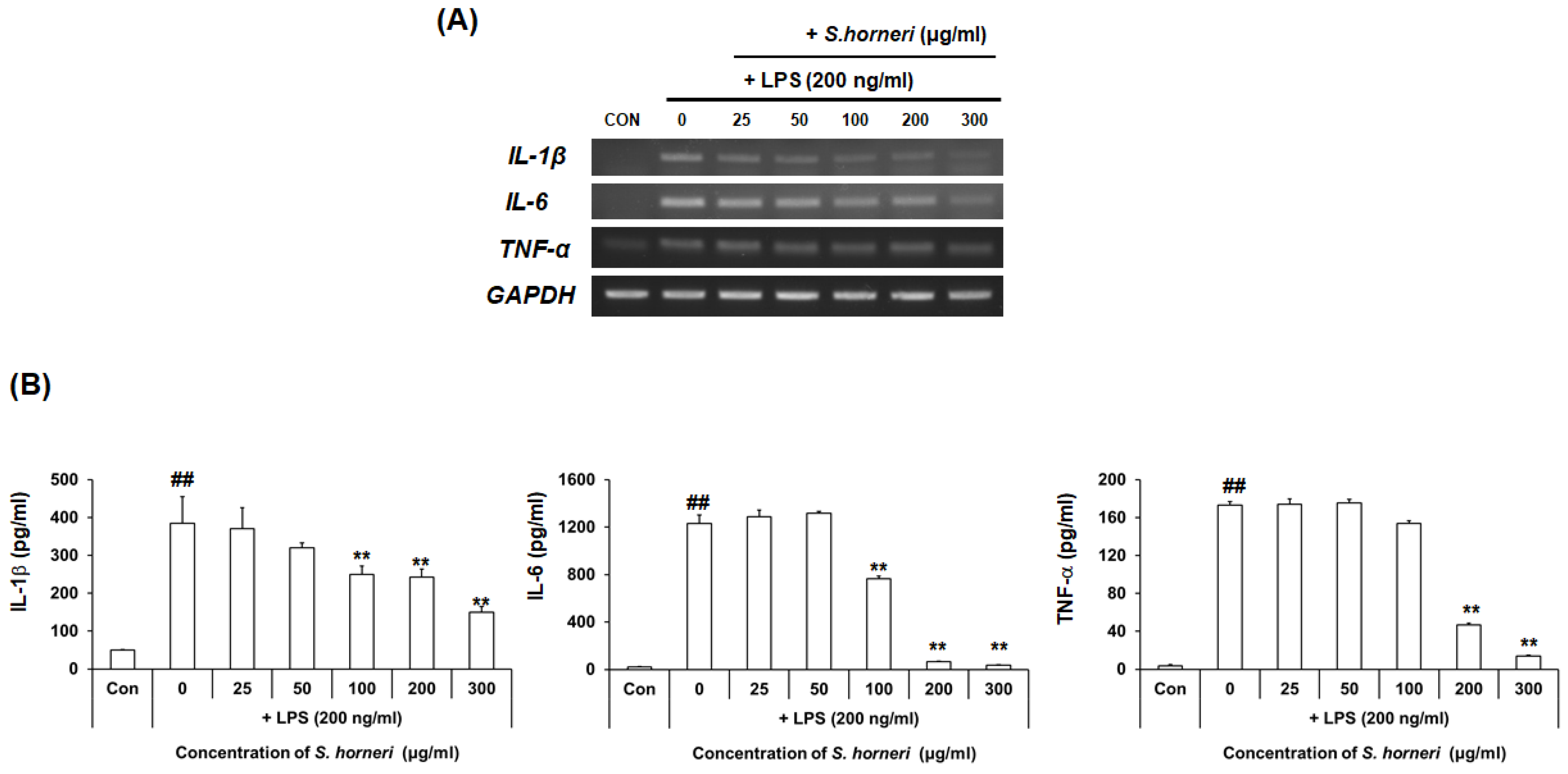
3.3. S. horneri Extract Reduced Neuroinflammatory Factors in LPS-Stimulated BV-2 Microglial Cells
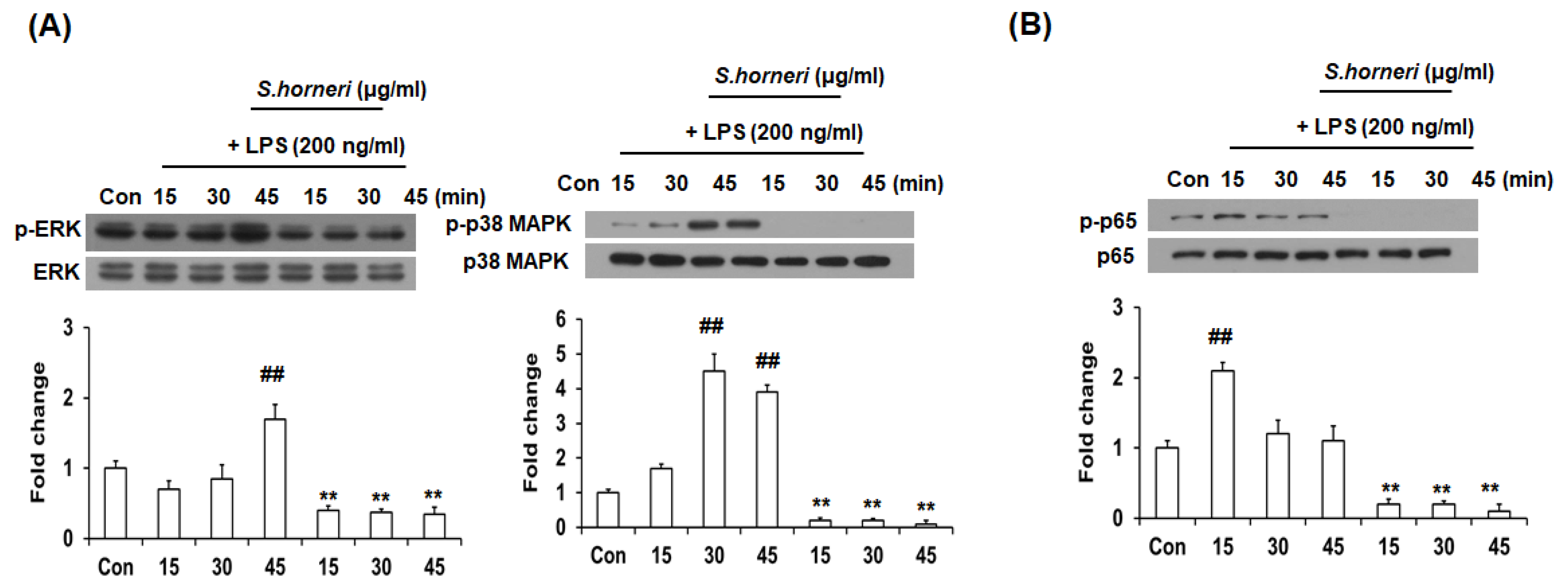
3.4. S. horneri Extract Inhibited Phosphorylation of p38 MAPK, ERK and NF-κB
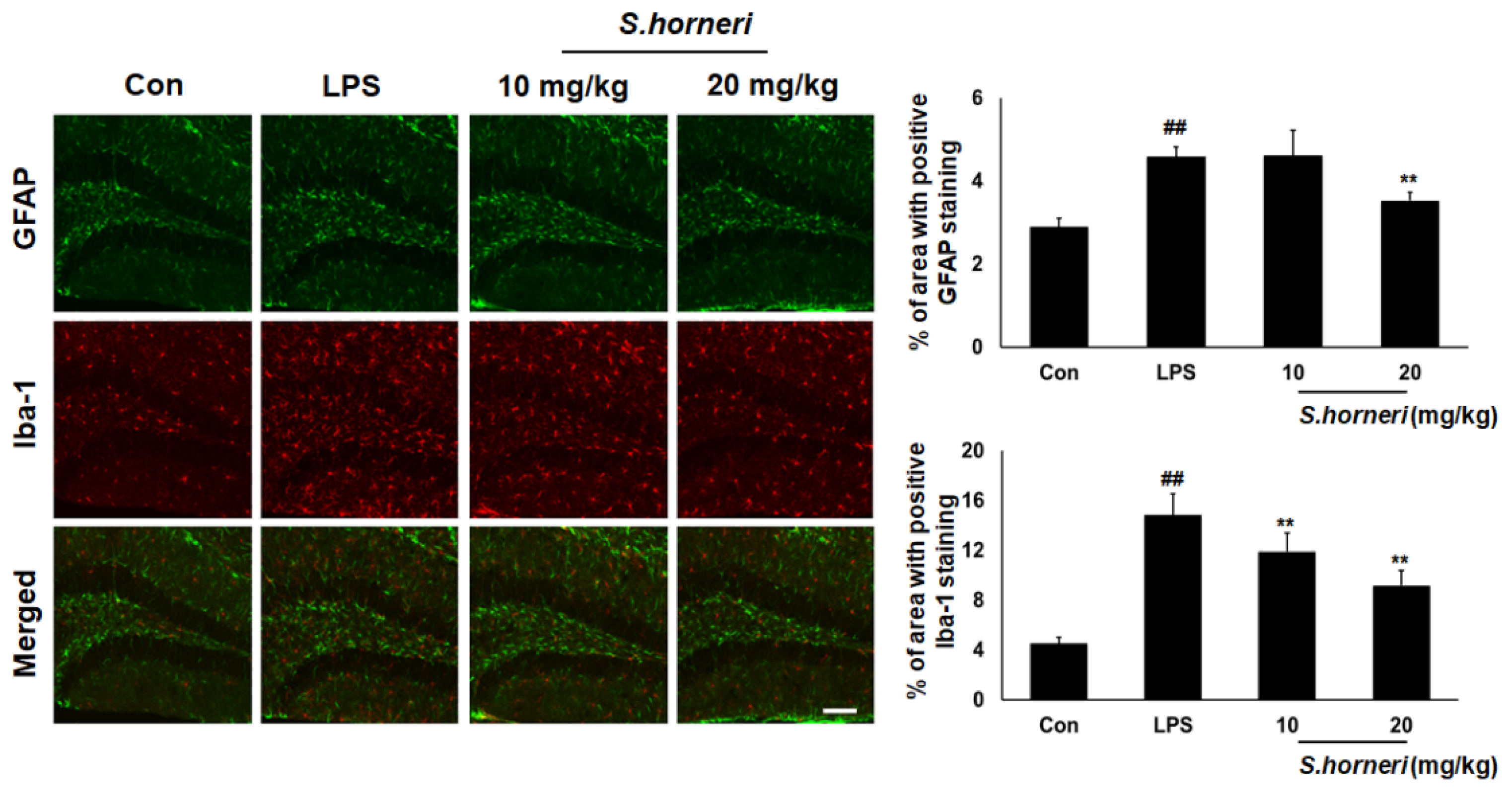
3.5. S. horneri Extract Attenuates Astrocyte and Microglia Activation in the Mice Brain
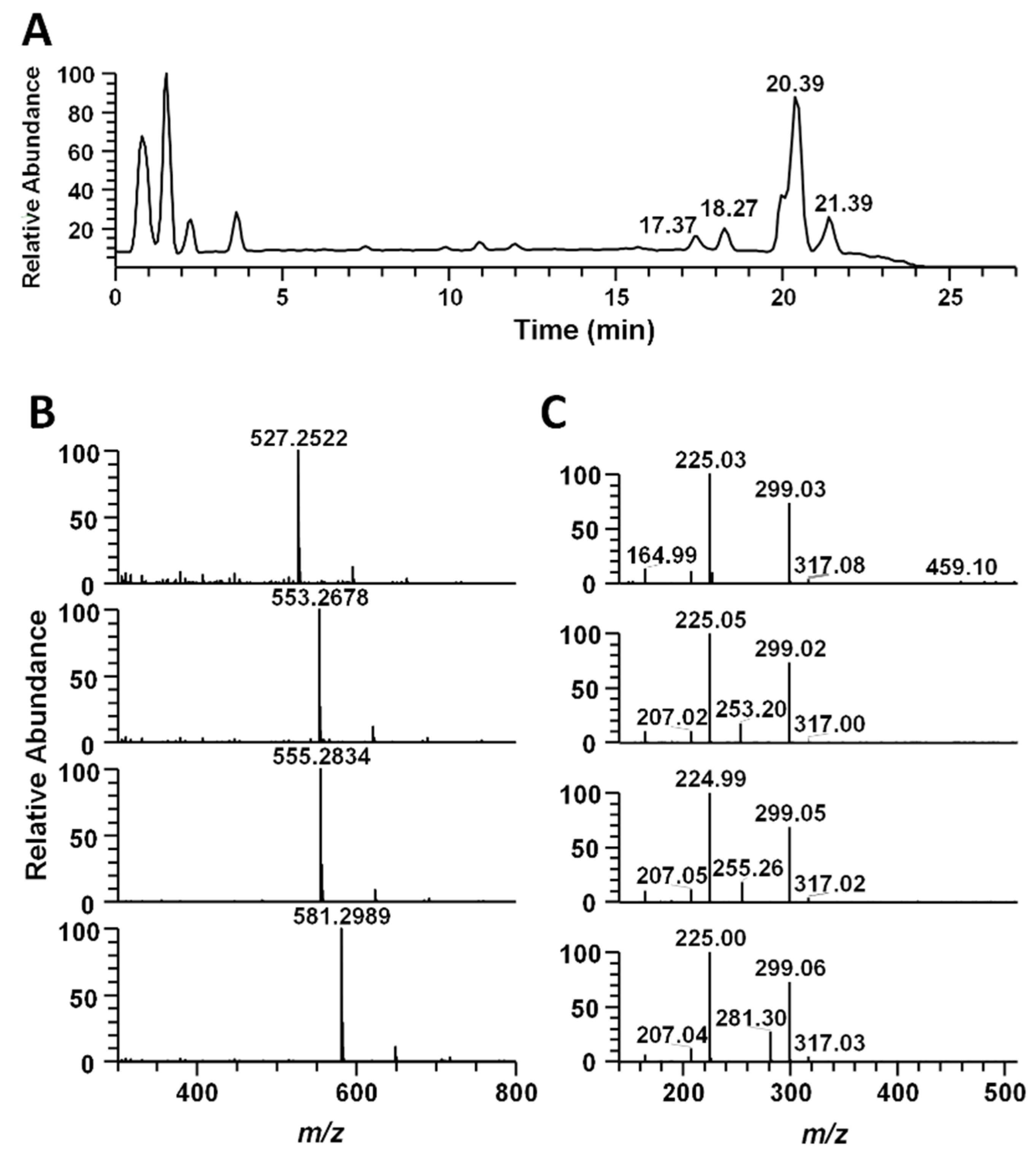
3.6. Identification of S. horneri Extract Using LC-MS/MS Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Morganti-Kossmann, M.C.; Satgunaseelan, L.; Bye, N.; Kossmann, T. Modulation of immune response by head injury. Injury 2007, 38, 1392–1400. [Google Scholar] [CrossRef]
- Sorrenti, V.; Giusti, P.; Zusso, M. A Model of Systemic Inflammation to Study Neuroinflammation. Methods Mol. Biol. 2018, 1727, 361–372. [Google Scholar] [PubMed]
- Broux, B.; Gowing, E.; Prat, A. Glial regulation of the blood-brain barrier in health and disease. Semin. Immunopathol. 2015, 37, 577–590. [Google Scholar] [CrossRef]
- Hung, T.H.; Shyue, S.K.; Wu, C.H.; Chen, C.C.; Lin, C.C.; Chang, C.F.; Chen, S.F. Deletion or inhibition of soluble epoxide hydrolase protects against brain damage and reduces microglia-mediated neuroinflammation in traumatic brain injury. Oncotarget 2017, 8, 103236–103260. [Google Scholar] [CrossRef] [Green Version]
- Shastri, A.; Bonifati, D.M.; Kishore, U. Innate immunity and neuroinflammation. Mediat. Inflamm. 2013, 2013, 342931. [Google Scholar] [CrossRef] [PubMed]
- Lehnardt, S. Innate immunity and neuroinflammation in the CNS: The role of microglia in Toll-like receptor-mediated neuronal injury. Glia 2010, 58, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, A. Microglia and brain macrophages: An update. Neuropathology 2016, 37, 452–464. [Google Scholar] [CrossRef] [PubMed]
- Skaper, S.D.; Facci, L.; Zusso, M.; Giusti, P. Neuroinflammation, Mast Cells, and Glia: Dangerous Liaisons. Neuroscientist 2017, 23, 478–498. [Google Scholar] [CrossRef]
- Fu, R.; Shen, Q.; Xu, P.; Luo, J.J.; Tang, Y. Phagocytosis of microglia in the central nervous system diseases. Mol. Neurobiol. 2014, 49, 1422–1434. [Google Scholar] [CrossRef] [Green Version]
- Block, M.L.; Zecca, L.; Hong, J.S. Microglia-mediated neurotoxicity: Uncovering the molecular mechanisms. Nat. Rev. Neurosci. 2007, 8, 57–69. [Google Scholar] [CrossRef]
- Chan, C.K.; Tan, L.T.; Andy, S.N.; Kamarudin, M.N.A.; Goh, B.H.; Kadir, H.A. Anti-neuroinflammatory Activity of Elephantopus scaber L. via Activation of Nrf2/HO-1 Signaling and Inhibition of p38 MAPK Pathway in LPS-Induced Microglia BV-2 Cells. Front. Pharm. 2017, 8, 397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geng, Y.; Zhu, S.; Cheng, P.; Lu, Z.M.; Xu, H.Y.; Shi, J.S.; Xu, Z.H. Bioassay-guided fractionation of ethyl acetate extract from Armillaria mellea attenuates inflammatory response in lipopolysaccharide (LPS) stimulated BV-2 microglia. Phytomedicine 2017, 26, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Sanjeewa, K.K.; Fernando, I.P.; Kim, E.A.; Ahn, G.; Jee, Y.; Jeon, Y.J. Anti-inflammatory activity of a sulfated polysaccharide isolated from an enzymatic digest of brown seaweed Sargassum horneri in RAW 264.7 cells. Nutr. Res. Pract. 2017, 11, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.E.; Jung, Y.C.; Jung, I.; Lee, H.W.; Youn, H.Y.; Lee, J.S. Anti-inflammatory effects of ethanolic extract from Sargassum horneri (Turner) C. Agardh on lipopolysaccharide-stimulated macrophage activation via NF-kappaB pathway regulation. Immunol. Investig. 2015, 44, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M. Regulatory mechanism of food factors in bone metabolism and prevention of osteoporosis. Yakugaku Zasshi 2006, 126, 1117–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Yang, R.; Li, M.; Zhou, Q.; Liang, X.; Elmada, Z.C. Effects of dietary fucoidan on the blood constituents, anti-oxidation and innate immunity of juvenile yellow catfish (Pelteobagrus fulvidraco). Fish Shellfish Immunol. 2014, 41, 264–270. [Google Scholar] [CrossRef]
- Kim, J.A.; Ahn, B.N.; Kong, C.S.; Kim, S.K. Protective effect of chromene isolated from Sargassum horneri against UV-A-induced damage in skin dermal fibroblasts. Exp. Dermatol. 2012, 21, 630–631. [Google Scholar] [CrossRef]
- Fujiwara, T.; Nishida, N.; Nota, J.; Kitani, T.; Aoishi, K.; Takahashi, H.; Sugahara, T.; Hato, N. Efficacy of chlorophyll c2 for seasonal allergic rhinitis: Single-center double-blind randomized control trial. Eur. Arch. Otorhinolaryngol. 2016, 273, 4289–4294. [Google Scholar] [CrossRef]
- Kim, M.E.; Jung, I.; Lee, J.S.; Na, J.Y.; Kim, W.J.; Kim, Y.O.; Park, Y.D.; Lee, J.S. Pseudane-VII Isolated from Pseudoalteromonas sp. M2 Ameliorates LPS-Induced Inflammatory Response In Vitro and In Vivo. Mar. Drugs 2017, 15, 336. [Google Scholar] [CrossRef] [Green Version]
- Qin, L.; Wu, X.; Block, M.L.; Liu, Y.; Breese, G.R.; Hong, J.S.; Knapp, D.J.; Crews, F.T. Systemic LPS causes chronic neuroinflammation and progressive neurodegeneration. Glia 2007, 55, 453–462. [Google Scholar] [CrossRef]
- Cui, Z.; Li, Y.S.; Liu, H.B.; Yuan, D.; Lu, B.R. Sulfoglycolipid from the marine brown alga Sargassum Hemiphyllum. J. Asian Nat. Prod. Res. 2001, 3, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Vanhoutte, P.M.; Leung, S.W. Vascular nitric oxide: Beyond eNOS. J. Pharm. Sci. 2015, 129, 83–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picon-Pages, P.; Garcia-Buendia, J.; Munoz, F.J. Functions and dysfunctions of nitric oxide in brain. Biochim. Biophys Acta Mol. Basis Dis. 2019, 1865, 1949–1967. [Google Scholar] [CrossRef]
- Madhu, B.P.; Singh, K.P.; Saminathan, M.; Singh, R.; Shivasharanappa, N.; Sharma, A.K.; Malik, Y.S.; Dhama, K.; Manjunatha, V. Role of nitric oxide in the regulation of immune responses during rabies virus infection in mice. Virusdisease 2016, 27, 387–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jo, S.H.; Kim, M.E.; Cho, J.H.; Lee, Y.; Lee, J.; Park, Y.D.; Lee, J.S. Hesperetin inhibits neuroinflammation on microglia by suppressing inflammatory cytokines and MAPK pathways. Arch. Pharm. Res. 2019, 42, 695–703. [Google Scholar] [CrossRef]
- Kim, M.E.; Park, P.R.; Na, J.Y.; Jung, I.; Cho, J.H.; Lee, J.S. Anti-neuroinflammatory effects of galangin in LPS-stimulated BV-2 microglia through regulation of IL-1beta production and the NF-kappaB signaling pathways. Mol. Cell Biochem. 2019, 451, 145–153. [Google Scholar] [CrossRef]
- D’Ignazio, L.; Bandarra, D.; Rocha, S. NF-kappaB and HIF crosstalk in immune responses. FEBS J. 2016, 283, 413–424. [Google Scholar] [CrossRef] [Green Version]
- El-Daly, S.M.; Omara, E.A.; Hussein, J.; Youness, E.R.; El-Khayat, Z. Differential expression of miRNAs regulating NF-kappaB and STAT3 crosstalk during colitis-associated tumorigenesis. Mol. Cell Probes 2019, 47, 101442. [Google Scholar] [CrossRef]
- Kumar, V. A sting to inflammation and autoimmunity. J. Leukoc. Biol. 2019, 106, 171–185. [Google Scholar] [CrossRef]
- Ahmad, S.F.; Ansari, M.A.; Zoheir, K.M.; Bakheet, S.A.; Korashy, H.M.; Nadeem, A.; Ashour, A.E.; Attia, S.M. Regulation of TNF-alpha and NF-kappaB activation through the JAK/STAT signaling pathway downstream of histamine 4 receptor in a rat model of LPS-induced joint inflammation. Immunobiology 2015, 220, 889–898. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RT (min) | m/z ([M-H]) | Formula | Δppm | Compound Identification |
|---|---|---|---|---|
| 17.37 | 527.2522 | C23H43O11S | −1.775 | 1-O-Tetradecanoyl-3-O-(6′-sulfo-a-D-quinovopyranosyl) glycerol |
| 18.27 | 553.2678 | C25H45O11S | −1.782 | 1-O-(11-Hexadecenoyl)-3-O-(6′-sulfo-a-D-quinovopyranosyl) glycerol |
| 20.39 | 555.2834 | C25H47O11S | −1.938 | 1-O-Hexadecanoyl-3-O-(6′-sulfo-a-D-quinovopyranosyl) glycerol |
| 21.39 | 581.2989 | C27H49O11S | −2.092 | 1-O-(9-Octadecenoyl)-3-O-(6′-sulfo-a-D-quinovopyranosyl) glycerol |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, J.H.; Kim, D.H.; Lee, J.S.; Seo, M.-S.; Kim, M.E.; Lee, J.S. Sargassum horneri (Turner) C. Agardh Extract Regulates Neuroinflammation In Vitro and In Vivo. Curr. Issues Mol. Biol. 2022, 44, 5416-5426. https://doi.org/10.3390/cimb44110367
Cho JH, Kim DH, Lee JS, Seo M-S, Kim ME, Lee JS. Sargassum horneri (Turner) C. Agardh Extract Regulates Neuroinflammation In Vitro and In Vivo. Current Issues in Molecular Biology. 2022; 44(11):5416-5426. https://doi.org/10.3390/cimb44110367
Chicago/Turabian StyleCho, Jun Hwi, Dae Hyun Kim, Jong Suk Lee, Mi-Suk Seo, Mi Eun Kim, and Jun Sik Lee. 2022. "Sargassum horneri (Turner) C. Agardh Extract Regulates Neuroinflammation In Vitro and In Vivo" Current Issues in Molecular Biology 44, no. 11: 5416-5426. https://doi.org/10.3390/cimb44110367
APA StyleCho, J. H., Kim, D. H., Lee, J. S., Seo, M. -S., Kim, M. E., & Lee, J. S. (2022). Sargassum horneri (Turner) C. Agardh Extract Regulates Neuroinflammation In Vitro and In Vivo. Current Issues in Molecular Biology, 44(11), 5416-5426. https://doi.org/10.3390/cimb44110367