

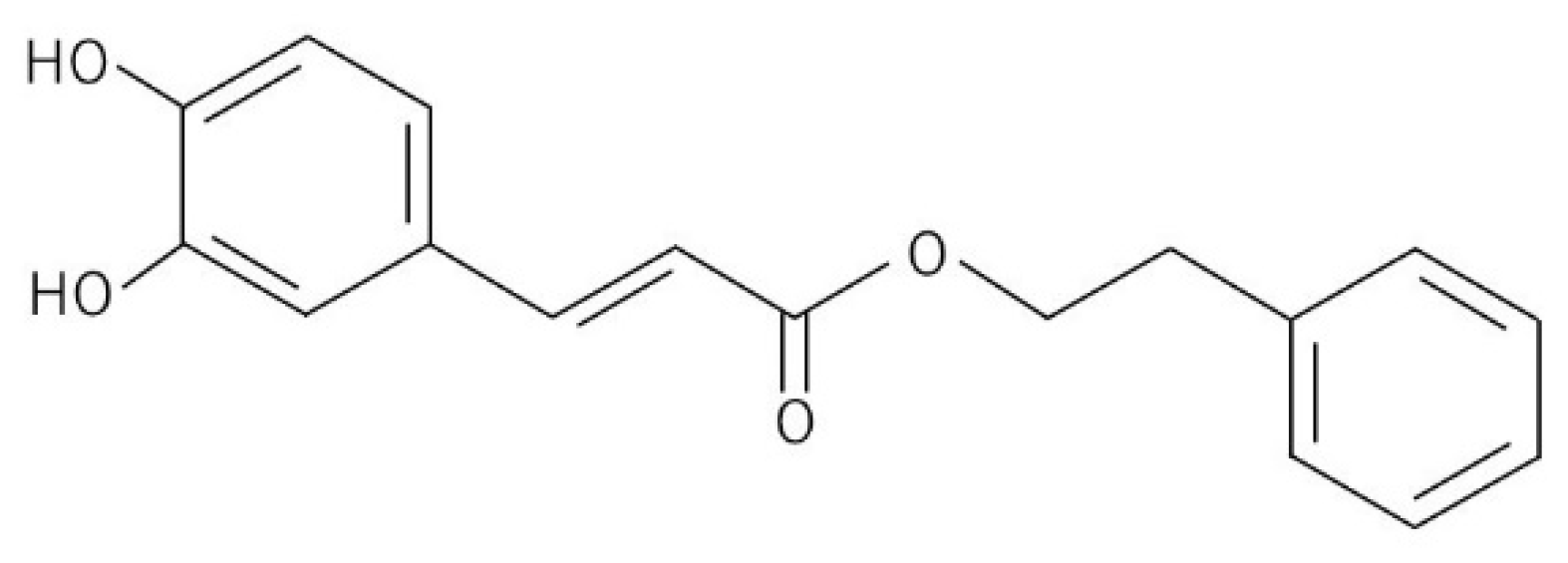
Caffeic Acid Phenethyl Ester Induces Vascular Endothelial Growth Factor Production and Inhibits CXCL10 Production in Human Dental Pulp Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Reagents
2.3. Cell Proliferation Assay
2.4. Enzyme-Linked Immunosorbent Assay (ELISA)
2.5. Western Blot Analysis
2.6. Statistical Analysis
3. Results
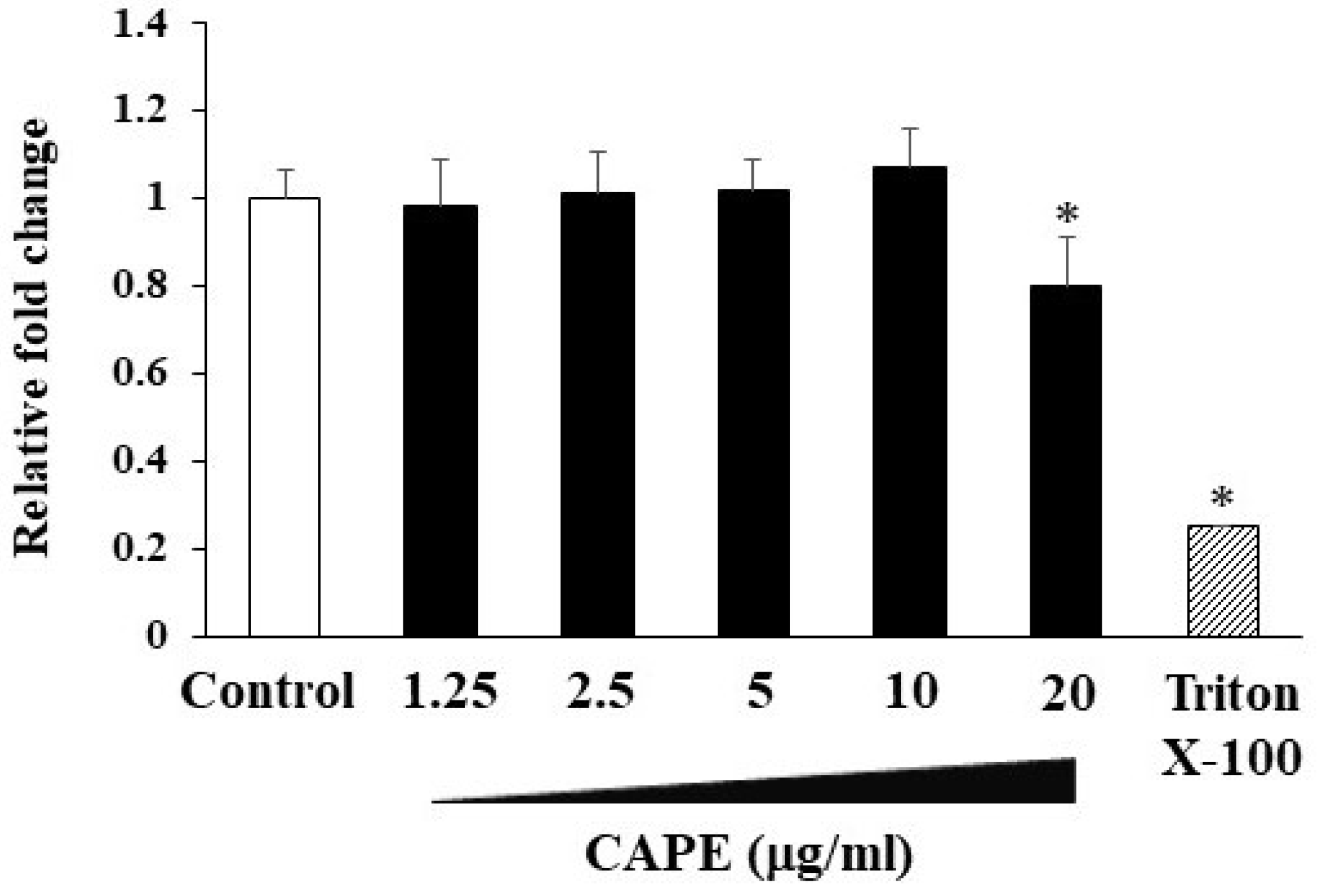
3.1. The Effects of CAPE on Cell Proliferation in HDPCs
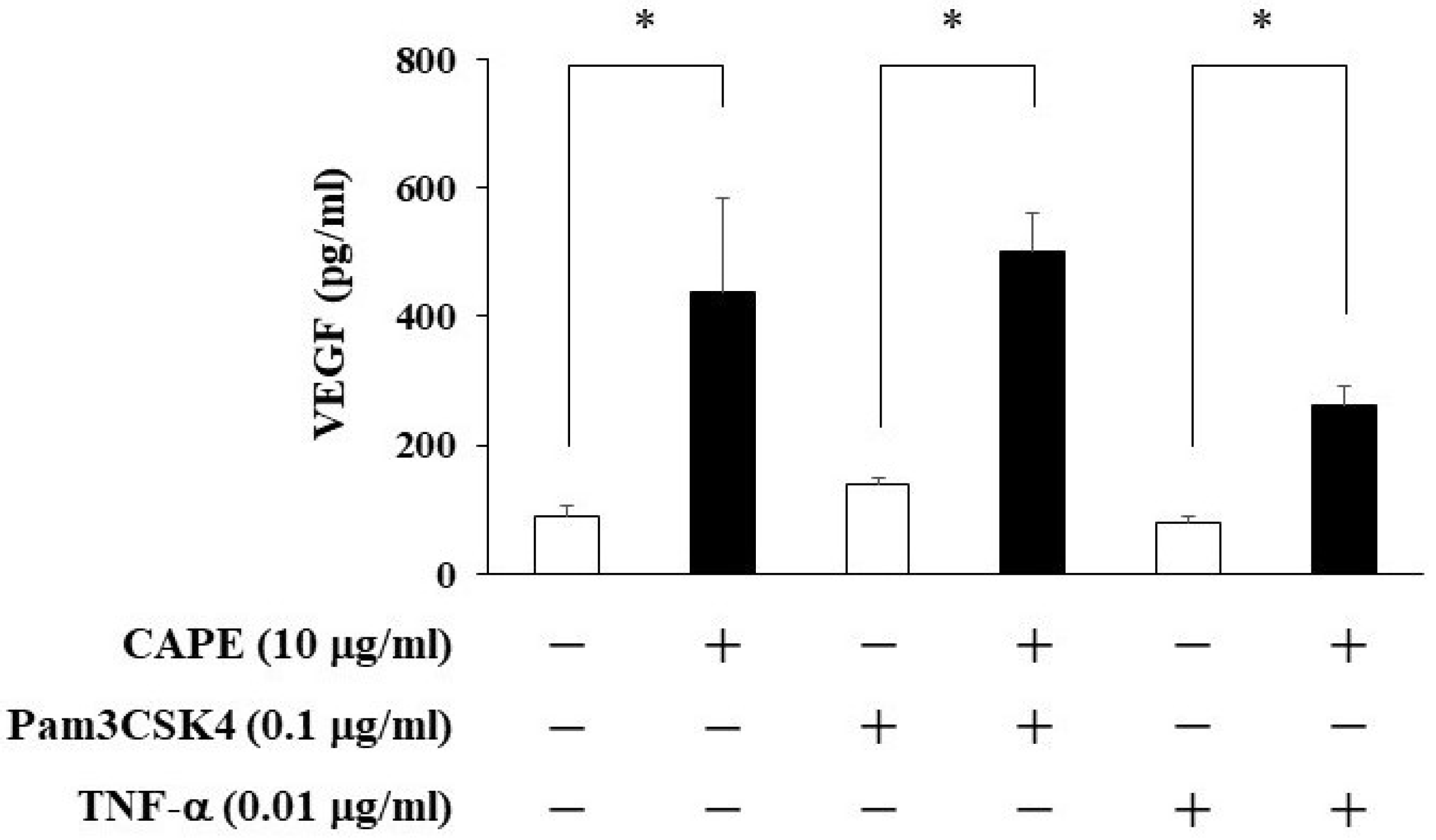
3.2. The Effects of CAPE on VEGF Production from HDPCs
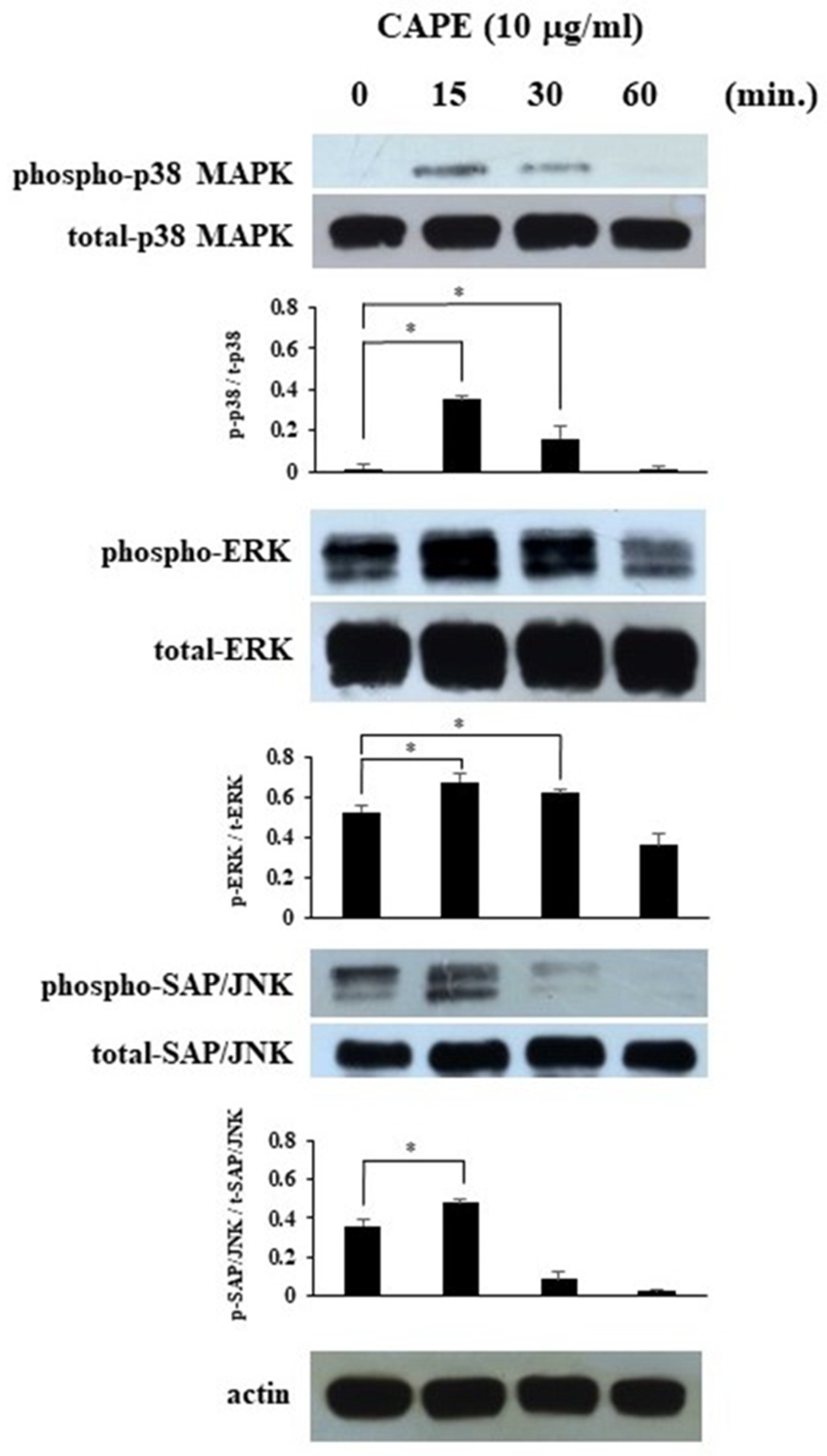
3.3. The Effects of CAPE on MAPKs’ Phosphorylation in HDPCs
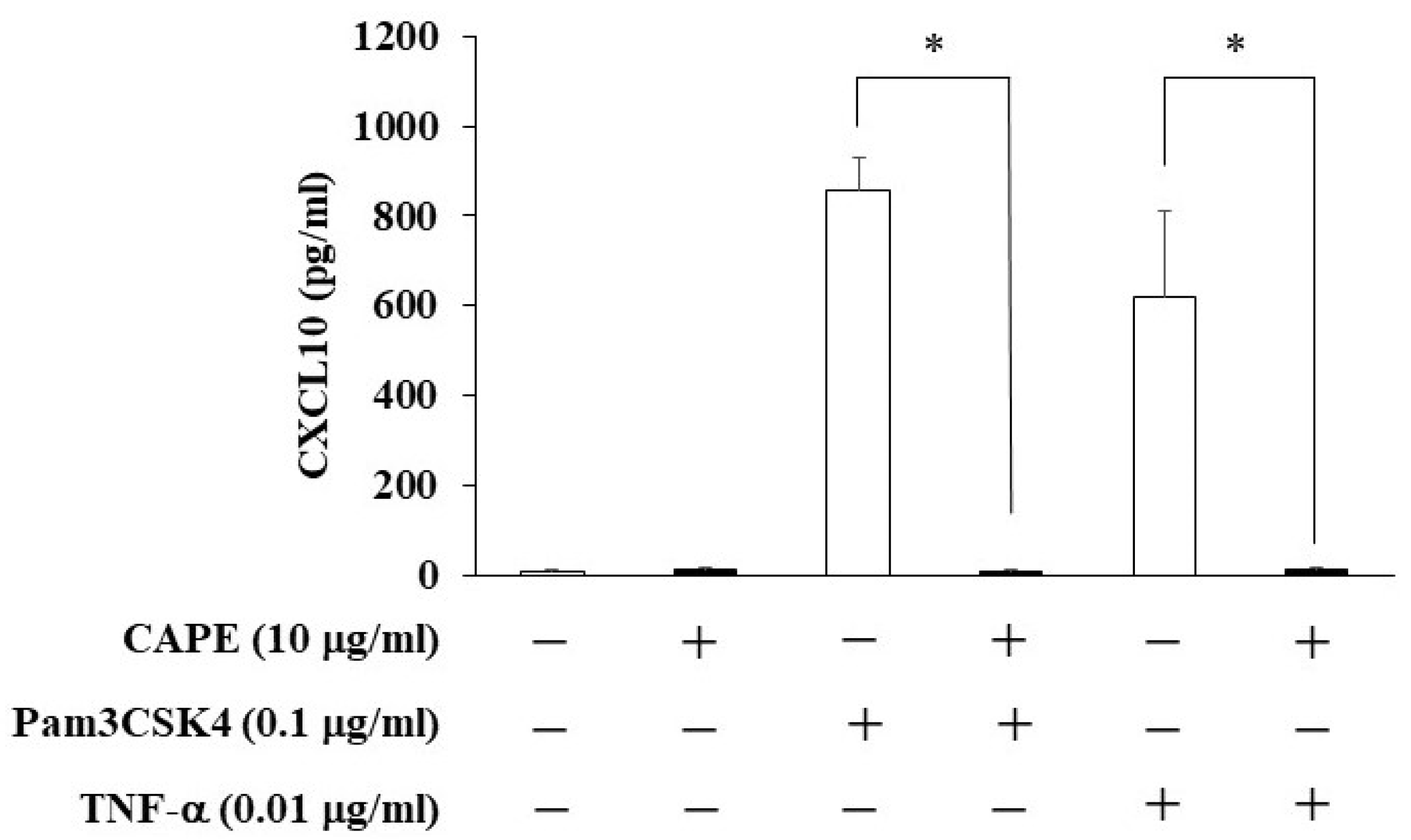
3.4. The Effects of CAPE on CXCL10 Production from HDPCs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Caplan, D.J.; Cai, J.; Yin, G.; White, B.A. Root canal filled versus non-root canal filled teeth: A retrospective comparison of survival times. J. Public Health Dent. 2005, 65, 90–96. [Google Scholar] [CrossRef]
- Leong, D.J.X.; Yap, A.U. Vital pulp therapy in carious pulp-exposed permanent teeth: An umbrella review. Clin. Oral Investig. 2021, 25, 6743–6756. [Google Scholar] [CrossRef]
- Manaspon, C.; Jongwannasiri, C.; Chumprasert, S.; Sa-Ard-Iam, N.; Mahanonda, R.; Pavasant, P.; Porntaveetus, T.; Osathanon, T. Human dental pulp stem cell responses to different dental pulp capping materials. BMC Oral Health 2021, 21, 209. [Google Scholar] [CrossRef] [PubMed]
- Awawdeh, L.; Al-Qudah, A.; Hamouri, H.; Chakra, R.J. Outcomes of Vital Pulp Therapy Using Mineral Trioxide Aggregate or Biodentine: A Prospective Randomized Clinical Trial. J. Endod. 2018, 44, 1603–1609. [Google Scholar] [CrossRef] [PubMed]
- Parirokh, M.; Torabinejad, M. Mineral trioxide aggregate: A comprehensive literature review—Part I: Chemical, physical, and antibacterial properties. J. Endod. 2010, 36, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Torabinejad, M.; Parirokh, M.; Dummer, P.M.H. Mineral trioxide aggregate and other bioactive endodontic cements: An updated overview—Part II: Other clinical applications and complications. Int. Endod. J. 2018, 51, 284–317. [Google Scholar] [CrossRef]
- Paula, A.B.; Laranjo, M.; Marto, C.M.; Paulo, S.; Abrantes, A.M.; Casalta-Lopes, J.; Marques-Ferreira, M.; Botelho, M.F.; Carrilho, E. Direct Pulp Capping: What is the Most Effective Therapy?-Systematic Review and Meta-Analysis. J. Evid.-Based Dent. Pract. 2018, 18, 298–314. [Google Scholar] [CrossRef] [PubMed]
- Lambrichts, I.; Driesen, R.B.; Dillen, Y.; Gervois, P.; Ratajczak, J.; Vangansewinkel, T.; Wolfs, E.; Bronckaers, A.; Hilkens, P. Dental Pulp Stem Cells: Their Potential in Reinnervation and Angiogenesis by Using Scaffolds. J. Endod. 2017, 43, S12–S16. [Google Scholar] [CrossRef]
- Tran-Hung, L.; Laurent, P.; Camps, J.; About, I. Quantification of angiogenic growth factors released by human dental cells after injury. Arch. Oral Biol. 2008, 53, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, X.; Yu, W.; Zhang, Y.; Shi, C.; Ni, S.; Liu, Q.; Li, X.; Sun, Y.; Zheng, C.; et al. Effects of human vascular endothelial growth factor on reparative dentin formation. Mol. Med. Rep. 2016, 13, 705–712. [Google Scholar] [CrossRef]
- Artese, L.; Rubini, C.; Ferrero, G.; Fioroni, M.; Santinelli, A.; Piattelli, A. Vascular endothelial growth factor (VEGF) expression in healthy and inflamed human dental pulps. J. Endod. 2002, 28, 20–23. [Google Scholar] [CrossRef] [PubMed]
- Hirao, K.; Yumoto, H.; Takahashi, K.; Mukai, K.; Nakanishi, T.; Matsuo, T. Roles of TLR2, TLR4, NOD2, and NOD1 in pulp fibroblasts. J. Dent. Res. 2009, 88, 762–767. [Google Scholar] [CrossRef] [PubMed]
- Adachi, T.; Nakanishi, T.; Yumoto, H.; Hirao, K.; Takahashi, K.; Mukai, K.; Nakae, H.; Matsuo, T. Caries-related bacteria and cytokines induce CXCL10 in dental pulp. J. Dent. Res. 2007, 86, 1217–1222. [Google Scholar] [CrossRef]
- Olgierd, B.; Kamila, Ż.; Anna, B.; Emilia, M. The Pluripotent Activities of Caffeic Acid Phenethyl Ester. Molecules 2021, 26, 1335. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.Y.; Jim, W.T.; Su, L.C.; Chung, C.J.; Lin, C.Y.; Huo, C.; Tseng, J.C.; Huang, S.H.; Lai, C.J.; Chen, B.C.; et al. Caffeic Acid phenethyl ester is a potential therapeutic agent for oral cancer. Int. J. Mol. Sci. 2015, 16, 10748–10766. [Google Scholar] [CrossRef] [Green Version]
- Karaboğa, İ. Caffeic acid phenethyl ester ameliorates pulmonary inflammation and apoptosis reducing Nf-κβ activation in blunt pulmonary contusion model. Ulus. Travma Ve Acil Cerrahi Derg. = Turk. J. Trauma Emerg. Surg. TJTES 2019, 25, 433–439. [Google Scholar] [CrossRef]
- Alfarrayeh, I.; Pollák, E.; Czéh, Á.; Vida, A.; Das, S.; Papp, G. Antifungal and Anti-Biofilm Effects of Caffeic Acid Phenethyl Ester on Different Candida Species. Antibiotics 2021, 10, 1359. [Google Scholar] [CrossRef]
- Mapesa, J.O.; Waldschmitt, N.; Schmoeller, I.; Blume, C.; Hofmann, T.; Mahungu, S.; Clavel, T.; Haller, D. Catechols in caffeic acid phenethyl ester are essential for inhibition of TNF-mediated IP-10 expression through NF-κB-dependent but HO-1- and p38-independent mechanisms in mouse intestinal epithelial cells. Mol. Nutr. Food Res. 2011, 55, 1850–1861. [Google Scholar] [CrossRef]
- Kuramoto, H.; Hirao, K.; Yumoto, H.; Hosokawa, Y.; Nakanishi, T.; Takegawa, D.; Washio, A.; Kitamura, C.; Matsuo, T. Caffeic Acid Phenethyl Ester (CAPE) Induces VEGF Expression and Production in Rat Odontoblastic Cells. BioMed Res. Int. 2019, 2019, 5390720. [Google Scholar] [CrossRef]
- Nakanishi, T.; Mukai, K.; Hosokawa, Y.; Takegawa, D.; Matsuo, T. Catechins inhibit vascular endothelial growth factor production and cyclooxygenase-2 expression in human dental pulp cells. Int. Endod. J. 2015, 48, 277–282. [Google Scholar] [CrossRef]
- Shin, M.R.; Kang, S.K.; Kim, Y.S.; Lee, S.Y.; Hong, S.C.; Kim, E.C. TNF-α and LPS activate angiogenesis via VEGF and SIRT1 signalling in human dental pulp cells. Int. Endod. J. 2015, 48, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Xie, X.; Liu, Z.; Yang, J.; Zuo, D.; Xu, S. Neu5Ac Induces Human Dental Pulp Stem Cell Osteo-/Odontoblastic Differentiation by Enhancing MAPK/ERK Pathway Activation. Stem Cells Int. 2021, 2021, 5560872. [Google Scholar] [CrossRef] [PubMed]
- Cargnello, M.; Roux, P.P. Activation and function of the MAPKs and their substrates, the MAPK-activated protein kinases. Microbiol. Mol. Biol. Rev. MMBR 2011, 75, 50–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.J.; Kuo, H.C.; Chu, C.Y.; Wang, C.J.; Lin, W.C.; Tseng, T.H. Involvement of tumor suppressor protein p53 and p38 MAPK in caffeic acid phenethyl ester-induced apoptosis of C6 glioma cells. Biochem. Pharmacol. 2003, 66, 2281–2289. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.P.; Tsui, K.H.; Chang, K.S.; Sung, H.C.; Hsu, S.Y.; Lin, Y.H.; Yang, P.S.; Chen, C.L.; Feng, T.H.; Juang, H.H. Caffeic acid phenethyl ester inhibits the growth of bladder carcinoma cells by upregulating growth differentiation factor 15. Biomed. J. 2022, 45, 763–775. [Google Scholar] [CrossRef]
- Owuor, E.D.; Kong, A.N. Antioxidants and oxidants regulated signal transduction pathways. Biochem. Pharmacol. 2002, 64, 765–770. [Google Scholar] [CrossRef]
- Hahn, C.L.; Liewehr, F.R. Relationships between caries bacteria, host responses, and clinical signs and symptoms of pulpitis. J. Endod. 2007, 33, 213–219. [Google Scholar] [CrossRef]
- Nagaoka, S.; Tokuda, M.; Sakuta, T.; Taketoshi, Y.; Tamura, M.; Takada, H.; Kawagoe, M. Interleukin-8 gene expression by human dental pulp fibroblast in cultures stimulated with Prevotella intermedia lipopolysaccharide. J. Endod. 1996, 22, 9–12. [Google Scholar] [CrossRef]
- Nakanishi, T.; Matsuo, T.; Ebisu, S. Quantitative analysis of immunoglobulins and inflammatory factors in human pulpal blood from exposed pulps. J. Endod. 1995, 21, 131–136. [Google Scholar] [CrossRef]
- Yang, J.W.; Jung, W.K.; Lee, C.M.; Yea, S.S.; Choi, Y.H.; Kim, G.Y.; Lee, D.S.; Na, G.; Park, S.G.; Seo, S.K.; et al. Caffeic acid phenethyl ester inhibits the inflammatory effects of interleukin-1β in human corneal fibroblasts. Immunopharmacol. Immunotoxicol. 2014, 36, 371–377. [Google Scholar] [CrossRef]
- Niu, Y.; Wang, K.; Zheng, S.; Wang, Y.; Ren, Q.; Li, H.; Ding, L.; Li, W.; Zhang, L. Antibacterial Effect of Caffeic Acid Phenethyl Ester on Cariogenic Bacteria and Streptococcus mutans Biofilms. Antimicrob. Agents Chemother. 2020, 64, e00251-20. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuramoto, H.; Nakanishi, T.; Takegawa, D.; Mieda, K.; Hosaka, K. Caffeic Acid Phenethyl Ester Induces Vascular Endothelial Growth Factor Production and Inhibits CXCL10 Production in Human Dental Pulp Cells. Curr. Issues Mol. Biol. 2022, 44, 5691-5699. https://doi.org/10.3390/cimb44110385
Kuramoto H, Nakanishi T, Takegawa D, Mieda K, Hosaka K. Caffeic Acid Phenethyl Ester Induces Vascular Endothelial Growth Factor Production and Inhibits CXCL10 Production in Human Dental Pulp Cells. Current Issues in Molecular Biology. 2022; 44(11):5691-5699. https://doi.org/10.3390/cimb44110385
Chicago/Turabian StyleKuramoto, Hitomi, Tadashi Nakanishi, Daisuke Takegawa, Katsuhiro Mieda, and Keiichi Hosaka. 2022. "Caffeic Acid Phenethyl Ester Induces Vascular Endothelial Growth Factor Production and Inhibits CXCL10 Production in Human Dental Pulp Cells" Current Issues in Molecular Biology 44, no. 11: 5691-5699. https://doi.org/10.3390/cimb44110385
APA StyleKuramoto, H., Nakanishi, T., Takegawa, D., Mieda, K., & Hosaka, K. (2022). Caffeic Acid Phenethyl Ester Induces Vascular Endothelial Growth Factor Production and Inhibits CXCL10 Production in Human Dental Pulp Cells. Current Issues in Molecular Biology, 44(11), 5691-5699. https://doi.org/10.3390/cimb44110385