
The Crosstalk between the Blood–Brain Barrier Dysfunction and Neuroinflammation after General Anaesthesia


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
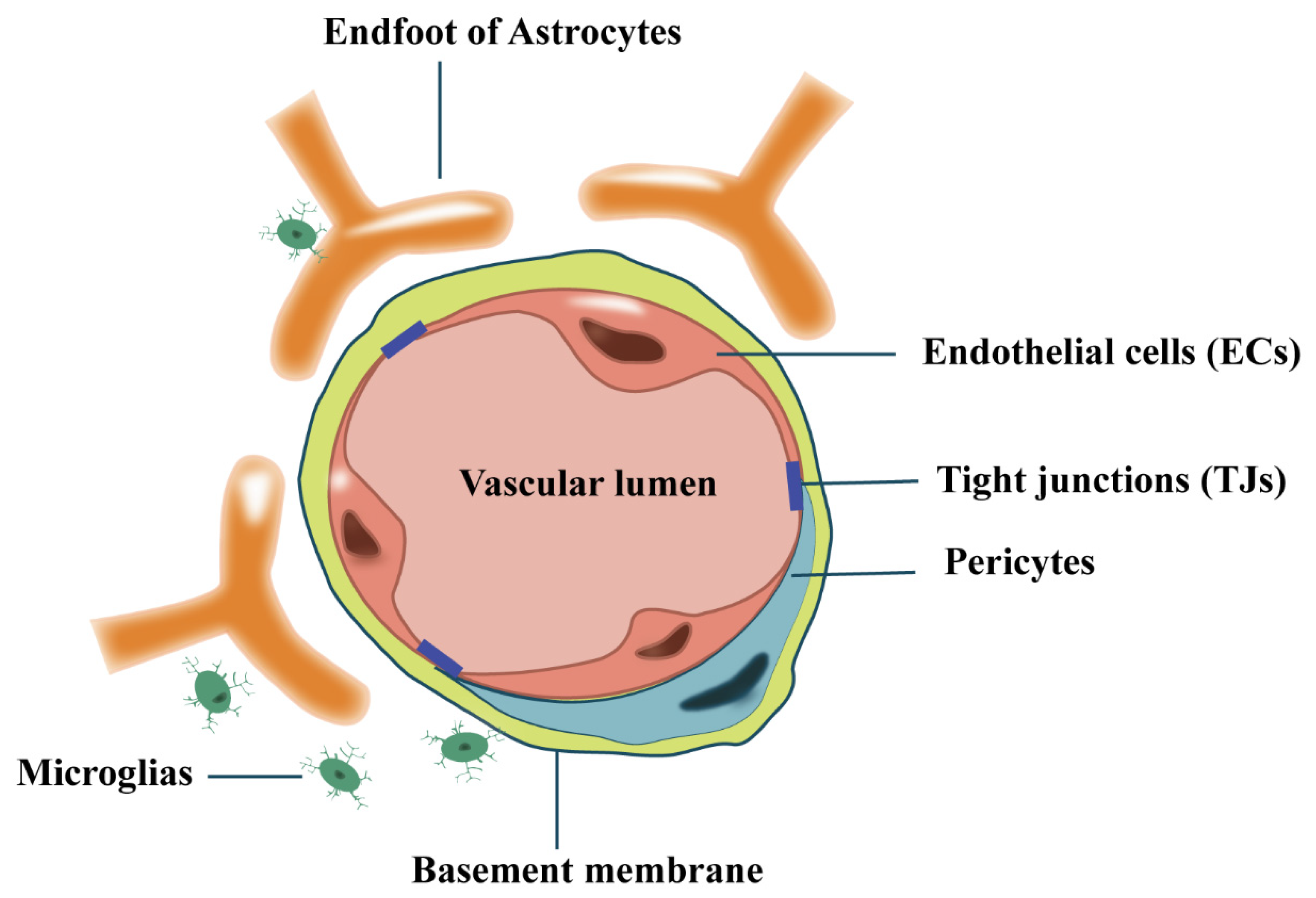
2. The Blood–Brain-Barrier (BBB)
2.1. The Generation of BBB
2.2. Endothelial Cells (ECs)
2.3. Tight Junctions (TJs)
2.4. Pericytes
2.5. Astrocytes
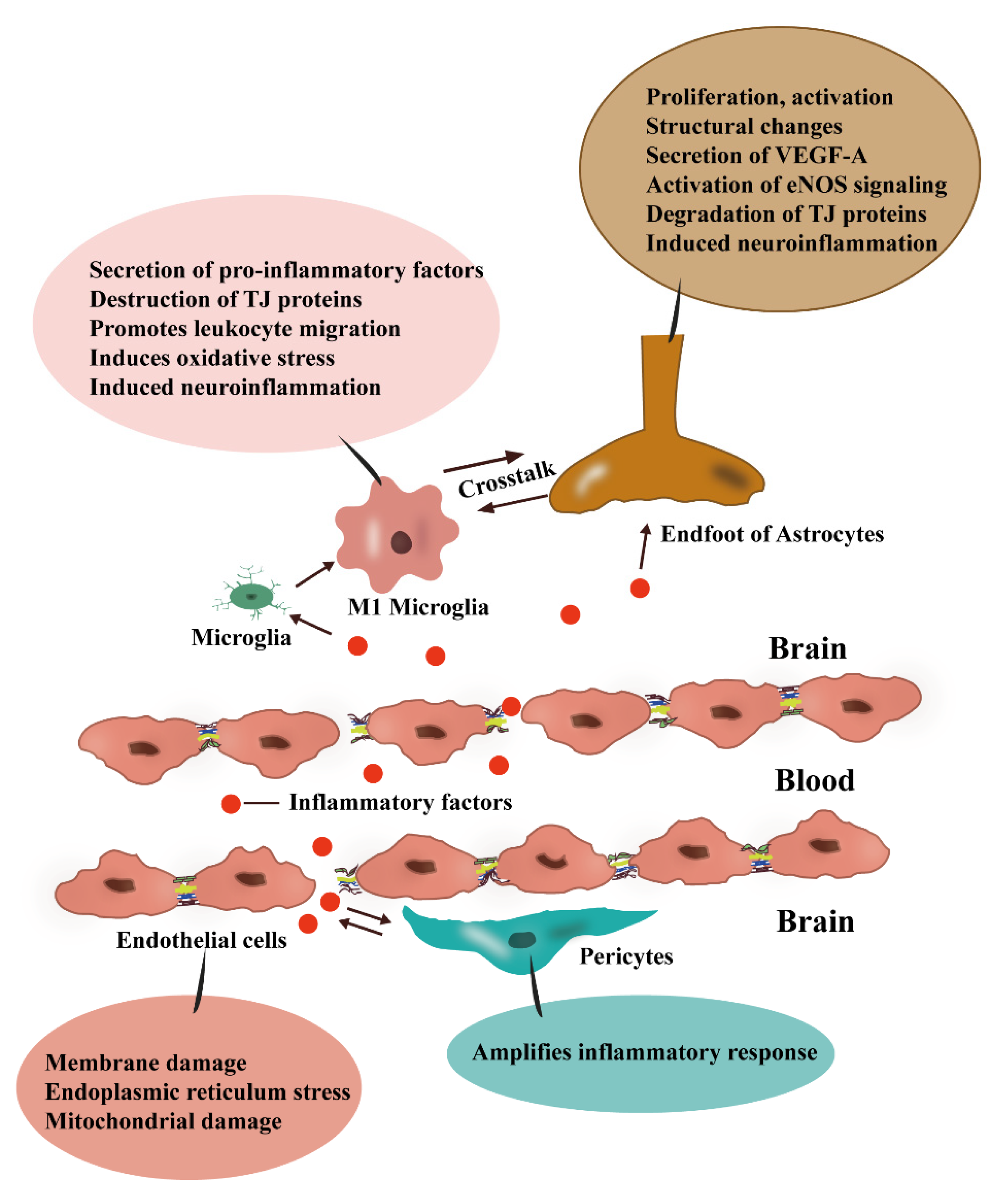
3. BBB and Inflammation
3.1. Damage to ECs Is a Key Component of Neuroinflammation
3.2. Inflammation Disrupts the Components of TJs
3.3. Pericytes Expand the Inflammatory Response
4. Glia Cells and Neuroinflammation
4.1. Astrocytes as Mediators between Peripheral Inflammation and Neuroinflammation
4.2. M1-Type Microglia Can Facilitate the Development of Neuroinflammation and Can also Disrupt the BBB
4.3. Crosstalk between Microglia and Astrocytes
5. Anaesthetics
5.1. Propofol
5.2. Inhalation Anaesthesia
5.3. Opioids
5.4. Different α2-Agonists
6. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moody, O.A.; Zhang, E.R.; Vincent, K.F.; Kato, R.; Melonakos, E.D.; Nehs, C.J.; Solt, K. The Neural Circuits Underlying General Anesthesia and Sleep. Anesth. Analg. 2021, 132, 1254–1264. [Google Scholar] [CrossRef] [PubMed]
- Vutskits, L.; Xie, Z. Lasting impact of general anaesthesia on the brain: Mechanisms and relevance. Nat. Rev. Neurosci. 2016, 17, 705–717. [Google Scholar] [CrossRef] [PubMed]
- Fodale, V.; Tripodi, V.F.; Penna, O.; Fama, F.; Squadrito, F.; Mondello, E.; David, A. An update on anesthetics and impact on the brain. Expert. Opin. Drug. Saf. 2017, 16, 997–1008. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Ji, J.; Zhao, G.Q. General anesthesia affecting on developing brain: Evidence from animal to clinical research. J. Anesth. 2020, 34, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Hao, L.J.; Zhang, Y.T.; Zhang, Y.; Zhang, C. Influence of general anesthetic exposure in developing brain on cognition and the underlying mechanisms. Sheng Li Xue Bao 2019, 71, 749–759. [Google Scholar] [PubMed]
- Fodale, V.; Ritchie, K.; Rasmussen, L.S.; Mandal, P.K. Anesthetics and Alzheimer’s disease: Background and research. J. Alzheimer’s Dis. 2010, 22 (Suppl. S3), 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.; Kang, Z.; Li, S.; Zeng, J.; Zhao, J. Sarm1 is Essential for Anesthesia-Induced Neuroinflammation and Cognitive Impairment in Aged Mice. Cell Mol. Neurobiol. 2022, 42, 1465–1476. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, X.; Li, S.; Li, W.; Teng, Y.; Zhou, Y.; Xiong, H. Carnosol alleviates sevoflurane-induced cognitive dysfunction by mediating NF-kappaB pathway in aged rats. Drug Dev. Res. 2022, 83, 1342–1350. [Google Scholar] [CrossRef]
- Peng, L.; Liu, S.; Xu, J.; Xie, W.; Fang, X.; Xia, T.; Gu, X. Metformin alleviates prolonged isoflurane inhalation induced cognitive decline via reducing neuroinflammation in adult mice. Int. Immunopharmacol. 2022, 109, 108903. [Google Scholar] [CrossRef]
- Daiello, L.A.; Racine, A.M.; Yun Gou, R.; Marcantonio, E.R.; Xie, Z.; Kunze, L.J.; Vlassakov, K.V.; Inouye, S.K.; Jones, R.N.; Alsop, D.; et al. Postoperative Delirium and Postoperative Cognitive Dysfunction: Overlap and Divergence. Anesthesiology 2019, 131, 477–491. [Google Scholar] [CrossRef]
- Calsolaro, V.; Edison, P. Neuroinflammation in Alzheimer’s disease: Current evidence and future directions. Alzheimer’s Dement. 2016, 12, 719–732. [Google Scholar] [CrossRef] [PubMed]
- El Idrissi, F.; Gressier, B.; Devos, D.; Belarbi, K. A Computational Exploration of the Molecular Network Associated to Neuroinflammation in Alzheimer’s Disease. Front. Pharmacol. 2021, 12, 630003. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Chopp, M.; Chen, J. Multifaceted roles of pericytes in central nervous system homeostasis and disease. J. Cereb. Blood Flow Metab. 2020, 40, 1381–1401. [Google Scholar] [CrossRef] [PubMed]
- Zlokovic, B.V. The blood-brain barrier in health and chronic neurodegenerative disorders. Neuron 2008, 57, 178–201. [Google Scholar] [CrossRef] [Green Version]
- Hughes, J.; Neese, O.; Bieber, D.; Lewis, K.; Ahmadi, L.; Parsons, D.; Canfield, S. The Effects of Propofol on a Human In Vitro Blood-Brain Barrier Model. Front. Cell. Neurosci. 2022, 16, 835649. [Google Scholar] [CrossRef]
- Spieth, L.; Berghoff, S.; Stumpf, S.; Winchenbach, J.; Michaelis, T.; Watanabe, T.; Gerndt, N.; Düking, T.; Hofer, S.; Ruhwedel, T.; et al. Anesthesia triggers drug delivery to experimental glioma in mice by hijacking caveolar transport. Neuro-Oncol. Adv. 2021, 3, vdab140. [Google Scholar] [CrossRef]
- Zhang, Q.; Zheng, M.; Betancourt, C.; Liu, L.; Sitikov, A.; Sladojevic, N.; Zhao, Q.; Zhang, J.; Liao, J.; Wu, R.J.O.m.; et al. Increase in Blood-Brain Barrier (BBB) Permeability Is Regulated by MMP3 via the ERK Signaling Pathway. Oxidative Med. Cell. Longev. 2021, 2021, 6655122. [Google Scholar] [CrossRef]
- Acharya, N.; Goldwaser, E.; Forsberg, M.; Godsey, G.; Johnson, C.; Sarkar, A.; DeMarshall, C.; Kosciuk, M.; Dash, J.; Hale, C.; et al. Sevoflurane and Isoflurane induce structural changes in brain vascular endothelial cells and increase blood-brain barrier permeability: Possible link to postoperative delirium and cognitive decline. Brain Res. 2015, 1620, 29–41. [Google Scholar] [CrossRef]
- Langen, U.H.; Ayloo, S.; Gu, C. Development and Cell Biology of the Blood-Brain Barrier. Annu. Rev. Cell Dev. Biol. 2019, 35, 591–613. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.; Kwon, H.J.; Huang, Z. A functional requirement for astroglia in promoting blood vessel development in the early postnatal brain. PLoS ONE 2012, 7, e48001. [Google Scholar] [CrossRef]
- Nakamura, K.; Ikeuchi, T.; Nara, K.; Rhodes, C.S.; Zhang, P.; Chiba, Y.; Kazuno, S.; Miura, Y.; Ago, T.; Arikawa-Hirasawa, E.; et al. Perlecan regulates pericyte dynamics in the maintenance and repair of the blood-brain barrier. J. Cell Biol. 2019, 218, 3506–3525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez, J.I.; Dodelet-Devillers, A.; Kebir, H.; Ifergan, I.; Fabre, P.J.; Terouz, S.; Sabbagh, M.; Wosik, K.; Bourbonniere, L.; Bernard, M.; et al. The Hedgehog pathway promotes blood-brain barrier integrity and CNS immune quiescence. Science 2011, 334, 1727–1731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, F.; Kanda, T. Pericytes of the Nervous System: Physiological and Pathological Role. Brain Nerve 2020, 72, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Reed, M.J.; Damodarasamy, M.; Banks, W.A. The extracellular matrix of the blood-brain barrier: Structural and functional roles in health, aging, and Alzheimer’s disease. Tissue Barriers 2019, 7, 1651157. [Google Scholar] [CrossRef]
- Augustin, H.G.; Koh, G.Y.; Thurston, G.; Alitalo, K. Control of vascular morphogenesis and homeostasis through the angiopoietin-Tie system. Nat. Rev. Mol. Cell Biol. 2009, 10, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Cano, F.; Hernandez-Kelly, L.C.; Ortega, A. The Blood-Brain Barrier: Much More Than a Selective Access to the Brain. Neurotox. Res. 2021, 39, 2154–2174. [Google Scholar] [CrossRef] [PubMed]
- Sigaud, R.; Dussault, N.; Berenguer-Daize, C.; Vellutini, C.; Benyahia, Z.; Cayol, M.; Parat, F.; Mabrouk, K.; Vazquez, R.; Riveiro, M.E.; et al. Role of the Tyrosine Phosphatase SHP-2 in Mediating Adrenomedullin Proangiogenic Activity in Solid Tumors. Front. Oncol. 2021, 11, 753244. [Google Scholar] [CrossRef]
- Sonar, S.A.; Shaikh, S.; Joshi, N.; Atre, A.N.; Lal, G. IFN-gamma promotes transendothelial migration of CD4(+) T cells across the blood-brain barrier. Immunol. Cell Biol. 2017, 95, 843–853. [Google Scholar] [CrossRef]
- Hermann, D.M.; ElAli, A. The abluminal endothelial membrane in neurovascular remodeling in health and disease. Sci. Signal. 2012, 5, re4. [Google Scholar] [CrossRef]
- Harilal, S.; Jose, J.; Parambi, D.G.T.; Kumar, R.; Unnikrishnan, M.K.; Uddin, M.S.; Mathew, G.E.; Pratap, R.; Marathakam, A.; Mathew, B. Revisiting the blood-brain barrier: A hard nut to crack in the transportation of drug molecules. Brain Res. Bull. 2020, 160, 121–140. [Google Scholar] [CrossRef]
- Lee, M.J.; Jang, Y.; Han, J.; Kim, S.J.; Ju, X.; Lee, Y.L.; Cui, J.; Zhu, J.; Ryu, M.J.; Choi, S.Y.; et al. Endothelial-specific Crif1 deletion induces BBB maturation and disruption via the alteration of actin dynamics by impaired mitochondrial respiration. J. Cereb. Blood Flow Metab. 2020, 40, 1546–1561. [Google Scholar] [CrossRef] [PubMed]
- Daneman, R.; Zhou, L.; Agalliu, D.; Cahoy, J.D.; Kaushal, A.; Barres, B.A. The mouse blood-brain barrier transcriptome: A new resource for understanding the development and function of brain endothelial cells. PLoS ONE 2010, 5, e13741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vestweber, D. Adhesion and signaling molecules controlling the transmigration of leukocytes through endothelium. Immunol. Rev. 2007, 218, 178–196. [Google Scholar] [CrossRef] [PubMed]
- Giannotta, M.; Trani, M.; Dejana, E. VE-cadherin and endothelial adherens junctions: Active guardians of vascular integrity. Dev. Cell 2013, 26, 441–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, Y.; Chen, J.; Freeman, W.; Dong, L.J.; Zhang, Z.H.; Xu, M.; Qiu, F.; Du, Y.; Liu, J.; Li, X.R.; et al. Canonical Wnt Signaling Promotes Neovascularization Through Determination of Endothelial Progenitor Cell Fate via Metabolic Profile Regulation. Stem Cells 2019, 37, 1331–1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Li, Y.; Han, Y.; Wang, S.; Yang, H.; Zhao, Y.; Li, P.; Wang, Y. Notoginsenoside R1 intervenes degradation and redistribution of tight junctions to ameliorate blood-brain barrier permeability by Caveolin-1/MMP2/9 pathway after acute ischemic stroke. Phytomedicine 2021, 90, 153660. [Google Scholar] [CrossRef]
- McConnell, H.L.; Mishra, A. Cells of the Blood-Brain Barrier: An Overview of the Neurovascular Unit in Health and Disease. Methods Mol. Biol. 2022, 2492, 3–24. [Google Scholar] [CrossRef]
- Cong, X.; Kong, W. Endothelial tight junctions and their regulatory signaling pathways in vascular homeostasis and disease. Cell. Signal. 2020, 66, 109485. [Google Scholar] [CrossRef]
- Liebner, S.; Fischmann, A.; Rascher, G.; Duffner, F.; Grote, E.H.; Kalbacher, H.; Wolburg, H. Claudin-1 and claudin-5 expression and tight junction morphology are altered in blood vessels of human glioblastoma multiforme. Acta Neuropathol. 2000, 100, 323–331. [Google Scholar] [CrossRef]
- Leon-Rivera, R.; Veenstra, M.; Donoso, M.; Tell, E.; Eugenin, E.A.; Morgello, S.; Berman, J.W. Central Nervous System (CNS) Viral Seeding by Mature Monocytes and Potential Therapies To Reduce CNS Viral Reservoirs in the cART Era. mBio 2021, 12, e03633-20. [Google Scholar] [CrossRef]
- Nagasawa, K.; Chiba, H.; Fujita, H.; Kojima, T.; Saito, T.; Endo, T.; Sawada, N. Possible involvement of gap junctions in the barrier function of tight junctions of brain and lung endothelial cells. J. Cell. Physiol. 2006, 208, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, C.; Schwietzer, Y.A.; Otani, T.; Furuse, M.; Ebnet, K. Physiological functions of junctional adhesion molecules (JAMs) in tight junctions. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183299. [Google Scholar] [CrossRef] [PubMed]
- Tornavaca, O.; Chia, M.; Dufton, N.; Almagro, L.O.; Conway, D.E.; Randi, A.M.; Schwartz, M.A.; Matter, K.; Balda, M.S. ZO-1 controls endothelial adherens junctions, cell-cell tension, angiogenesis, and barrier formation. J. Cell Biol. 2015, 208, 821–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanning, A.S.; Anderson, J.M. Zonula occludens-1 and -2 are cytosolic scaffolds that regulate the assembly of cellular junctions. Ann. N. Y. Acad. Sci. 2009, 1165, 113–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itoh, M.; Furuse, M.; Morita, K.; Kubota, K.; Saitou, M.; Tsukita, S. Direct binding of three tight junction-associated MAGUKs, ZO-1, ZO-2, and ZO-3, with the COOH termini of claudins. J. Cell Biol. 1999, 147, 1351–1363. [Google Scholar] [CrossRef] [Green Version]
- Ebnet, K.; Schulz, C.U.; Meyer Zu Brickwedde, M.K.; Pendl, G.G.; Vestweber, D. Junctional adhesion molecule interacts with the PDZ domain-containing proteins AF-6 and ZO-1. J. Biol. Chem. 2000, 275, 27979–27988. [Google Scholar] [CrossRef] [Green Version]
- Beazley-Long, N.; Durrant, A.M.; Swift, M.N.; Donaldson, L.F. The physiological functions of central nervous system pericytes and a potential role in pain. F1000Research 2018, 7, 341. [Google Scholar] [CrossRef] [Green Version]
- ten Dijke, P.; Arthur, H.M. Extracellular control of TGFbeta signalling in vascular development and disease. Nat. Rev. Mol. Cell Biol. 2007, 8, 857–869. [Google Scholar] [CrossRef]
- Antonelli-Orlidge, A.; Saunders, K.B.; Smith, S.R.; D’Amore, P.A. An activated form of transforming growth factor beta is produced by cocultures of endothelial cells and pericytes. Proc. Natl. Acad. Sci. USA 1989, 86, 4544–4548. [Google Scholar] [CrossRef] [Green Version]
- Hill, J.; Rom, S.; Ramirez, S.H.; Persidsky, Y. Emerging roles of pericytes in the regulation of the neurovascular unit in health and disease. J. Neuroimmune Pharm. 2014, 9, 591–605. [Google Scholar] [CrossRef]
- Dickson, M.C.; Martin, J.S.; Cousins, F.M.; Kulkarni, A.B.; Karlsson, S.; Akhurst, R.J. Defective haematopoiesis and vasculogenesis in transforming growth factor-beta 1 knock out mice. Development 1995, 121, 1845–1854. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Yuan, Y.; Wang, D.; Su, Z. Heterogeneous astrocytes: Active players in CNS. Brain Res. Bull. 2016, 125, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Broux, B.; Gowing, E.; Prat, A. Glial regulation of the blood-brain barrier in health and disease. Semin. Immunopathol. 2015, 37, 577–590. [Google Scholar] [CrossRef] [PubMed]
- Abbott, N.J.; Ronnback, L.; Hansson, E. Astrocyte-endothelial interactions at the blood-brain barrier. Nat. Rev. Neurosci. 2006, 7, 41–53. [Google Scholar] [CrossRef]
- Wang, Y.; Jin, S.; Sonobe, Y.; Cheng, Y.; Horiuchi, H.; Parajuli, B.; Kawanokuchi, J.; Mizuno, T.; Takeuchi, H.; Suzumura, A. Interleukin-1beta induces blood-brain barrier disruption by downregulating Sonic hedgehog in astrocytes. PLoS ONE 2014, 9, e110024. [Google Scholar] [CrossRef] [Green Version]
- Fontijn, R.D.; Volger, O.L.; Fledderus, J.O.; Reijerkerk, A.; de Vries, H.E.; Horrevoets, A.J. SOX-18 controls endothelial-specific claudin-5 gene expression and barrier function. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, H891–H900. [Google Scholar] [CrossRef]
- Podjaski, C.; Alvarez, J.I.; Bourbonniere, L.; Larouche, S.; Terouz, S.; Bin, J.M.; Lecuyer, M.A.; Saint-Laurent, O.; Larochelle, C.; Darlington, P.J.; et al. Netrin 1 regulates blood-brain barrier function and neuroinflammation. Brain 2015, 138, 1598–1612. [Google Scholar] [CrossRef] [Green Version]
- Suri, C.; Jones, P.F.; Patan, S.; Bartunkova, S.; Maisonpierre, P.C.; Davis, S.; Sato, T.N.; Yancopoulos, G.D. Requisite role of angiopoietin-1, a ligand for the TIE2 receptor, during embryonic angiogenesis. Cell 1996, 87, 1171–1180. [Google Scholar] [CrossRef] [Green Version]
- Lane-Donovan, C.; Herz, J. The ApoE receptors Vldlr and Apoer2 in central nervous system function and disease. J. Lipid Res. 2017, 58, 1036–1043. [Google Scholar] [CrossRef] [Green Version]
- Lane-Donovan, C.; Philips, G.T.; Herz, J. More than cholesterol transporters: Lipoprotein receptors in CNS function and neurodegeneration. Neuron 2014, 83, 771–787. [Google Scholar] [CrossRef]
- Zhang, J.; Sadowska, G.B.; Chen, X.; Park, S.Y.; Kim, J.E.; Bodge, C.A.; Cummings, E.; Lim, Y.P.; Makeyev, O.; Besio, W.G.; et al. Anti-IL-6 neutralizing antibody modulates blood-brain barrier function in the ovine fetus. FASEB J. 2015, 29, 1739–1753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Hussain, B.; Chang, J. Peripheral inflammation and blood-brain barrier disruption: Effects and mechanisms. CNS Neurosci. Ther. 2021, 27, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Subramaniyan, S.; Terrando, N. Neuroinflammation and Perioperative Neurocognitive Disorders. Anesth. Analg. 2019, 128, 781–788. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.; Stone, K.P.; Hsuchou, H.; Manda, V.K.; Zhang, Y.; Kastin, A.J. Cytokine signaling modulates blood-brain barrier function. Curr. Pharm. Des. 2011, 17, 3729–3740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso, F.L.; Kittel, A.; Veszelka, S.; Palmela, I.; Toth, A.; Brites, D.; Deli, M.A.; Brito, M.A. Exposure to lipopolysaccharide and/or unconjugated bilirubin impair the integrity and function of brain microvascular endothelial cells. PLoS ONE 2012, 7, e35919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernhart, E.; Kogelnik, N.; Prasch, J.; Gottschalk, B.; Goeritzer, M.; Depaoli, M.R.; Reicher, H.; Nusshold, C.; Plastira, I.; Hammer, A.; et al. 2-Chlorohexadecanoic acid induces ER stress and mitochondrial dysfunction in brain microvascular endothelial cells. Redox Biol. 2018, 15, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Labus, J.; Woltje, K.; Stolte, K.N.; Hackel, S.; Kim, K.S.; Hildmann, A.; Danker, K. IL-1beta promotes transendothelial migration of PBMCs by upregulation of the FN/alpha5beta1 signalling pathway in immortalised human brain microvascular endothelial cells. Exp. Cell Res. 2018, 373, 99–111. [Google Scholar] [CrossRef]
- Zheng, X.; Liang, Y.; Kang, A.; Ma, S.J.; Xing, L.; Zhou, Y.Y.; Dai, C.; Xie, H.; Xie, L.; Wang, G.J.; et al. Peripheral immunomodulation with ginsenoside Rg1 ameliorates neuroinflammation-induced behavioral deficits in rats. Neuroscience 2014, 256, 210–222. [Google Scholar] [CrossRef]
- Tan, S.; Shan, Y.; Lin, Y.; Liao, S.; Zhang, B.; Zeng, Q.; Wang, Y.; Deng, Z.; Chen, C.; Hu, X.; et al. Neutralization of interleukin-9 ameliorates experimental stroke by repairing the blood-brain barrier via down-regulation of astrocyte-derived vascular endothelial growth factor-A. FASEB J. 2019, 33, 4376–4387. [Google Scholar] [CrossRef]
- Menard, C.; Pfau, M.L.; Hodes, G.E.; Kana, V.; Wang, V.X.; Bouchard, S.; Takahashi, A.; Flanigan, M.E.; Aleyasin, H.; LeClair, K.B.; et al. Social stress induces neurovascular pathology promoting depression. Nat. Neurosci. 2017, 20, 1752–1760. [Google Scholar] [CrossRef]
- Chen, W.; Ju, X.Z.; Lu, Y.; Ding, X.W.; Miao, C.H.; Chen, J.W. Propofol improved hypoxia-impaired integrity of blood-brain barrier via modulating the expression and phosphorylation of zonula occludens-1. CNS Neurosci. Ther. 2019, 25, 704–713. [Google Scholar] [CrossRef] [PubMed]
- Nitta, T.; Hata, M.; Gotoh, S.; Seo, Y.; Sasaki, H.; Hashimoto, N.; Furuse, M.; Tsukita, S. Size-selective loosening of the blood-brain barrier in claudin-5-deficient mice. J. Cell Biol. 2003, 161, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xue, G.X.; Liu, W.C.; Shu, H.; Wang, M.; Sun, Y.; Liu, X.; Sun, Y.E.; Liu, C.F.; Liu, J.; et al. Melatonin alleviates lipopolysaccharide-compromised integrity of blood-brain barrier through activating AMP-activated protein kinase in old mice. Aging Cell 2017, 16, 414–421. [Google Scholar] [CrossRef]
- Piro, J.R.; Suidan, G.L.; Quan, J.; Pi, Y.; O’Neill, S.M.; Ilardi, M.; Pozdnyakov, N.; Lanz, T.A.; Xi, H.; Bell, R.D.; et al. Inhibition of 2-AG hydrolysis differentially regulates blood brain barrier permeability after injury. J. Neuroinflammat. 2018, 15, 142. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Wang, X.; Zhang, R.; Jiao, Y.; Yu, W.; Su, D.; Zhao, Y.; Tian, J. Mice with pre-existing tumors are vulnerable to postoperative cognitive dysfunction. Brain Res. 2020, 1732, 146650. [Google Scholar] [CrossRef] [PubMed]
- Sil, S.; Niu, F.; Tom, E.; Liao, K.; Periyasamy, P.; Buch, S. Cocaine Mediated Neuroinflammation: Role of Dysregulated Autophagy in Pericytes. Mol. NeuroBiol. 2019, 56, 3576–3590. [Google Scholar] [CrossRef] [PubMed]
- Medina-Flores, F.; Hurtado-Alvarado, G.; Deli, M.A.; Gomez-Gonzalez, B. The Active Role of Pericytes During Neuroinflammation in the Adult Brain. Cell Mol. Neurobiol. 2022. [Google Scholar] [CrossRef]
- Dohgu, S.; Takata, F.; Matsumoto, J.; Kimura, I.; Yamauchi, A.; Kataoka, Y. Monomeric alpha-synuclein induces blood-brain barrier dysfunction through activated brain pericytes releasing inflammatory mediators in vitro. Microvasc. Res. 2019, 124, 61–66. [Google Scholar] [CrossRef]
- Jansson, D.; Rustenhoven, J.; Feng, S.; Hurley, D.; Oldfield, R.L.; Bergin, P.S.; Mee, E.W.; Faull, R.L.; Dragunow, M. A role for human brain pericytes in neuroinflammation. J. Neuroinflammat. 2014, 11, 104. [Google Scholar] [CrossRef] [Green Version]
- Kan, M.H.; Yang, T.; Fu, H.Q.; Fan, L.; Wu, Y.; Terrando, N.; Wang, T.L. Pyrrolidine Dithiocarbamate Prevents Neuroinflammation and Cognitive Dysfunction after Endotoxemia in Rats. Front. Aging Neurosci. 2016, 8, 175. [Google Scholar] [CrossRef]
- Yang, T.; Velagapudi, R.; Terrando, N. Neuroinflammation after surgery: From mechanisms to therapeutic targets. Nat. Immunol. 2020, 21, 1319–1326. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.C.B.; Dammer, E.B.; Duong, D.M.; Ping, L.; Zhou, M.; Yin, L.; Higginbotham, L.A.; Guajardo, A.; White, B.; Troncoso, J.C.; et al. Large-scale proteomic analysis of Alzheimer’s disease brain and cerebrospinal fluid reveals early changes in energy metabolism associated with microglia and astrocyte activation. Nat. Med. 2020, 26, 769–780. [Google Scholar] [CrossRef] [PubMed]
- Kam, T.I.; Hinkle, J.T.; Dawson, T.M.; Dawson, V.L. Microglia and astrocyte dysfunction in parkinson’s disease. Neurobiol. Dis. 2020, 144, 105028. [Google Scholar] [CrossRef]
- Donato, R.; Cannon, B.R.; Sorci, G.; Riuzzi, F.; Hsu, K.; Weber, D.J.; Geczy, C.L. Functions of S100 proteins. Curr. Mol. Med. 2013, 13, 24–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farfara, D.; Feierman, E.; Richards, A.; Revenko, A.S.; MacLeod, R.A.; Norris, E.H.; Strickland, S. Knockdown of circulating C1 inhibitor induces neurovascular impairment, glial cell activation, neuroinflammation, and behavioral deficits. Glia 2019, 67, 1359–1373. [Google Scholar] [CrossRef]
- Biesmans, S.; Meert, T.F.; Bouwknecht, J.A.; Acton, P.D.; Davoodi, N.; De Haes, P.; Kuijlaars, J.; Langlois, X.; Matthews, L.J.; Ver Donck, L.; et al. Systemic immune activation leads to neuroinflammation and sickness behavior in mice. Mediat. Inflamm. 2013, 2013, 271359. [Google Scholar] [CrossRef]
- Zamanian, J.L.; Xu, L.; Foo, L.C.; Nouri, N.; Zhou, L.; Giffard, R.G.; Barres, B.A. Genomic analysis of reactive astrogliosis. J. Neurosci. 2012, 32, 6391–6410. [Google Scholar] [CrossRef] [Green Version]
- Argaw, A.T.; Asp, L.; Zhang, J.; Navrazhina, K.; Pham, T.; Mariani, J.N.; Mahase, S.; Dutta, D.J.; Seto, J.; Kramer, E.G.; et al. Astrocyte-derived VEGF-A drives blood-brain barrier disruption in CNS inflammatory disease. J. Clin. Investig. 2012, 122, 2454–2468. [Google Scholar] [CrossRef] [Green Version]
- Arcuri, C.; Bianchi, R.; Brozzi, F.; Donato, R. S100B increases proliferation in PC12 neuronal cells and reduces their responsiveness to nerve growth factor via Akt activation. J. Biol. Chem. 2005, 280, 4402–4414. [Google Scholar] [CrossRef] [Green Version]
- Sorci, G.; Bianchi, R.; Riuzzi, F.; Tubaro, C.; Arcuri, C.; Giambanco, I.; Donato, R. S100B Protein, A Damage-Associated Molecular Pattern Protein in the Brain and Heart, and Beyond. Cardiovasc. Psychiatry Neurol. 2010, 2010. [Google Scholar] [CrossRef] [Green Version]
- Petrova, T.V.; Hu, J.; Van Eldik, L.J. Modulation of glial activation by astrocyte-derived protein S100B: Differential responses of astrocyte and microglial cultures. Brain Res. 2000, 853, 74–80. [Google Scholar] [CrossRef]
- Adami, C.; Bianchi, R.; Pula, G.; Donato, R. S100B-stimulated NO production by BV-2 microglia is independent of RAGE transducing activity but dependent on RAGE extracellular domain. Biochim. Biophys. Acta 2004, 1742, 169–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchi, R.; Kastrisianaki, E.; Giambanco, I.; Donato, R. S100B protein stimulates microglia migration via RAGE-dependent up-regulation of chemokine expression and release. J. Biol. Chem. 2011, 286, 7214–7226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Wang, Y.; Wu, H.; Lei, L.; Xu, S.; Shen, X.; Guo, X.; Shen, R.; Xia, X.; Liu, Y.; et al. Postoperative cognitive dysfunction: Current developments in mechanism and prevention. Med. Sci. Monit. 2014, 20, 1908–1912. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.S.; Suh, Y.H. Minocycline and neurodegenerative diseases. Behav. Brain Res. 2009, 196, 168–179. [Google Scholar] [CrossRef]
- Stojiljkovic, M.R.; Schmeer, C.; Witte, O.W. Pharmacological Depletion of Microglia Leads to a Dose-Dependent Reduction in Inflammation and Senescence in the Aged Murine Brain. Neuroscience 2022, 488, 1–9. [Google Scholar] [CrossRef]
- O’Neil, S.M.; Hans, E.E.; Jiang, S.; Wangler, L.M.; Godbout, J.P. Astrocyte immunosenescence and deficits in interleukin 10 signaling in the aged brain disrupt the regulation of microglia following innate immune activation. Glia 2022, 70, 913–934. [Google Scholar] [CrossRef]
- Ronaldson, P.T.; Davis, T.P. Regulation of blood-brain barrier integrity by microglia in health and disease: A therapeutic opportunity. J. Cereb. Blood Flow Metab. 2020, 40, S6–S24. [Google Scholar] [CrossRef]
- Da Fonseca, A.C.; Matias, D.; Garcia, C.; Amaral, R.; Geraldo, L.H.; Freitas, C.; Lima, F.R. The impact of microglial activation on blood-brain barrier in brain diseases. Front. Cell. Neurosci. 2014, 8, 362. [Google Scholar] [CrossRef] [Green Version]
- Haruwaka, K.; Ikegami, A.; Tachibana, Y.; Ohno, N.; Konishi, H.; Hashimoto, A.; Matsumoto, M.; Kato, D.; Ono, R.; Kiyama, H.; et al. Dual microglia effects on blood brain barrier permeability induced by systemic inflammation. Nat. Commun. 2019, 10, 5816. [Google Scholar] [CrossRef]
- Guo, Z.N.; Liu, J.; Chang, J.; Zhang, P.; Jin, H.; Sun, X.; Yang, Y. GAS6/Axl Signaling Modulates Blood-Brain Barrier Function Following Intravenous Thrombolysis in Acute Ischemic Stroke. Front. Immunol. 2021, 12, 742359. [Google Scholar] [CrossRef] [PubMed]
- Rochfort, K.D.; Cummins, P.M. The blood-brain barrier endothelium: A target for pro-inflammatory cytokines. Biochem. Soc. Trans. 2015, 43, 702–706. [Google Scholar] [CrossRef] [PubMed]
- Kayki-Mutlu, G.; Koch, W.J. Nitric Oxide and S-Nitrosylation in Cardiac Regulation: G Protein-Coupled Receptor Kinase-2 and beta-Arrestins as Targets. Int. J. Mol. Sci. 2021, 22, 521. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Dhammu, T.S.; Sakakima, H.; Shunmugavel, A.; Gilg, A.G.; Singh, A.K.; Singh, I. The inhibitory effect of S-nitrosoglutathione on blood-brain barrier disruption and peroxynitrite formation in a rat model of experimental stroke. J. Neurochem. 2012, 123 (Suppl. S2), 86–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.D.; Yang, D.I.; Lin, T.K.; Shaw, F.Z.; Liou, C.W.; Chuang, Y.C. Roles of oxidative stress, apoptosis, PGC-1alpha and mitochondrial biogenesis in cerebral ischemia. Int. J. Mol. Sci. 2011, 12, 7199–7215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logsdon, A.F.; Meabon, J.S.; Cline, M.M.; Bullock, K.M.; Raskind, M.A.; Peskind, E.R.; Banks, W.A.; Cook, D.G. Blast exposure elicits blood-brain barrier disruption and repair mediated by tight junction integrity and nitric oxide dependent processes. Sci. Rep. 2018, 8, 11344. [Google Scholar] [CrossRef] [Green Version]
- Subramaniam, S.R.; Federoff, H.J. Targeting Microglial Activation States as a Therapeutic Avenue in Parkinson’s Disease. Front. Aging Neurosci. 2017, 9, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.J.; Ran, Y.Y.; Qie, S.Y.; Gong, W.J.; Gao, F.H.; Ding, Z.T.; Xi, J.N. Melatonin protects against ischemic stroke by modulating microglia/macrophage polarization toward anti-inflammatory phenotype through STAT3 pathway. CNS Neurosci. Ther. 2019, 25, 1353–1362. [Google Scholar] [CrossRef] [Green Version]
- Garcia, J.M.; Stillings, S.A.; Leclerc, J.L.; Phillips, H.; Edwards, N.J.; Robicsek, S.A.; Hoh, B.L.; Blackburn, S.; Dore, S. Role of Interleukin-10 in Acute Brain Injuries. Front. Neurol. 2017, 8, 244. [Google Scholar] [CrossRef] [Green Version]
- Mori, S.; Maher, P.; Conti, B. Neuroimmunology of the Interleukins 13 and 4. Brain Sci. 2016, 6, 18. [Google Scholar] [CrossRef]
- Rocken, M.; Racke, M.; Shevach, E.M. IL-4-induced immune deviation as antigen-specific therapy for inflammatory autoimmune disease. Immunol. Today 1996, 17, 225–231. [Google Scholar] [CrossRef]
- Raphael, I.; Nalawade, S.; Eagar, T.N.; Forsthuber, T.G. T cell subsets and their signature cytokines in autoimmune and inflammatory diseases. Cytokine 2015, 74, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Dong, H.; Qian, Q.; Zhang, X.; Wang, Y.; Jin, W.; Qian, Y. Astrocyte-derived CCL2 participates in surgery-induced cognitive dysfunction and neuroinflammation via evoking microglia activation. Behav. Brain Res. 2017, 332, 145–153. [Google Scholar] [CrossRef]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Munch, A.E.; Chung, W.S.; Peterson, T.C.; et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef] [Green Version]
- Pekny, M.; Wilhelmsson, U.; Tatlisumak, T.; Pekna, M. Astrocyte activation and reactive gliosis-A new target in stroke? Neurosci. Lett. 2019, 689, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Litvinchuk, A.; Wan, Y.W.; Swartzlander, D.B.; Chen, F.; Cole, A.; Propson, N.E.; Wang, Q.; Zhang, B.; Liu, Z.; Zheng, H. Complement C3aR Inactivation Attenuates Tau Pathology and Reverses an Immune Network Deregulated in Tauopathy Models and Alzheimer’s Disease. Neuron 2018, 100, 1337–1353.e1335. [Google Scholar] [CrossRef]
- Chen, T.; Lennon, V.A.; Liu, Y.U.; Bosco, D.B.; Li, Y.; Yi, M.H.; Zhu, J.; Wei, S.; Wu, L.J. Astrocyte-microglia interaction drives evolving neuromyelitis optica lesion. J. Clin. Investig. 2020, 130, 4025–4038. [Google Scholar] [CrossRef]
- Li, D.; Chen, M.; Meng, T.; Fei, J. Hippocampal microglial activation triggers a neurotoxic-specific astrocyte response and mediates etomidate-induced long-term synaptic inhibition. J. Neuroinflammat. 2020, 17, 109. [Google Scholar] [CrossRef] [Green Version]
- Xing, C.; Li, W.; Deng, W.; Ning, M.; Lo, E.H. A potential gliovascular mechanism for microglial activation: Differential phenotypic switching of microglia by endothelium versus astrocytes. J. Neuroinflammat. 2018, 15, 143. [Google Scholar] [CrossRef] [Green Version]
- Turan, A.; Duncan, A.; Leung, S.; Karimi, N.; Fang, J.; Mao, G.; Hargrave, J.; Gillinov, M.; Trombetta, C.; Ayad, S.; et al. Dexmedetomidine for reduction of atrial fibrillation and delirium after cardiac surgery (DECADE): A randomised placebo-controlled trial. Lancet 2020, 396, 177–185. [Google Scholar] [CrossRef]
- Zhang, D.F.; Su, X.; Meng, Z.T.; Li, H.L.; Wang, D.X.; Xue-Ying, L.; Maze, M.; Ma, D. Impact of Dexmedetomidine on Long-term Outcomes After Noncardiac Surgery in Elderly: 3-Year Follow-up of a Randomized Controlled Trial. Ann. Surg. 2019, 270, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Evered, L.A.; Silbert, B.S. Postoperative Cognitive Dysfunction and Noncardiac Surgery. Anesth. Analg. 2018, 127, 496–505. [Google Scholar] [CrossRef] [PubMed]
- Balon, R. Benzodiazepines revisited. Psychother. Psychosom. 2013, 82, 353–354. [Google Scholar] [CrossRef] [PubMed]
- McPherson, C.; Grunau, R.E. Neonatal pain control and neurologic effects of anesthetics and sedatives in preterm infants. Clin. Perinatol. 2014, 41, 209–227. [Google Scholar] [CrossRef] [Green Version]
- Volkow, N.D.; Baler, R.D. Addiction science: Uncovering neurobiological complexity. Neuropharmacology 2014, 76, 235–249. [Google Scholar] [CrossRef] [Green Version]
- Doyle, H.H.; Eidson, L.N.; Sinkiewicz, D.M.; Murphy, A.Z. Sex Differences in Microglia Activity within the Periaqueductal Gray of the Rat: A Potential Mechanism Driving the Dimorphic Effects of Morphine. J. Neurosci. 2017, 37, 3202–3214. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Zhao, S.; Qiao, H.; Li, T.; Mi, W.; Xu, Z.; Xue, X. Does propofol definitely improve postoperative cognitive dysfunction?-a review of propofol-related cognitive impairment. Acta Biochim. Biophys. Sin. 2022, 54, 875–881. [Google Scholar] [CrossRef]
- Vinckier, F.; Gaillard, R.; Taylor, G.; Murray, G.K.; Plaze, M.; Bourdillon, P.; Perin-Dureau, F. Acute psychosis following propofol in a patient with Parkinson disease: Effects of a gamma-aminobutyric acid-dopamine imbalance. Psychiatry Clin. Neurosci. 2022, 76, 273–274. [Google Scholar] [CrossRef]
- Liu, C.; Shi, F.; Fu, B.; Luo, T.; Zhang, L.; Zhang, Y.; Zhang, Y.; Yu, S.; Yu, T. GABAA receptors in the basal forebrain mediates emergence from propofol anaesthesia in rats. Int. J. Neurosci. 2022, 132, 802–814. [Google Scholar] [CrossRef]
- Song, F.; Lv, X.; Meng, J. Propofol Induces Postoperative Depression and Inhibits Microglial Function in Mice. Mediat. Inflamm. 2019, 2019, 7651383. [Google Scholar] [CrossRef]
- Sun, W.; Pei, L. Dexmedetomidine attenuates propofol-induced apoptosis of neonatal hippocampal astrocytes by inhibiting the Bcl2l1 signalling pathway. Eur. J. Neurosci. 2021, 54, 7775–7789. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Huang, B.; Wei, Q.; Su, X.; Li, X.; Qin, S.; Huang, W. The Protective Effects of Benzbromarone Against Propofol-Induced Inflammation and Injury in Human Brain Microvascular Endothelial Cells (HBMVECs). Neurotox. Res. 2021, 39, 1449–1458. [Google Scholar] [CrossRef] [PubMed]
- Langrich, T.; Bork, K.; Horstkorte, R.; Weber, V.; Hofmann, B.; Fuszard, M.; Olzscha, H. Disturbance of Key Cellular Subproteomes upon Propofol Treatment Is Associated with Increased Permeability of the Blood-Brain Barrier. Proteomes 2022, 10, 28. [Google Scholar] [CrossRef] [PubMed]
- Sharma, H.S.; Ponten, E.; Gordh, T.; Eriksson, P.; Fredriksson, A.; Sharma, A. Propofol promotes blood-brain barrier breakdown and heat shock protein (HSP 72 kd) activation in the developing mouse brain. CNS Neurol. Disord. Drug Targets 2014, 13, 1595–1603. [Google Scholar] [CrossRef]
- Ji, F.T.; Liang, J.J.; Miao, L.P.; Wu, Q.; Cao, M.H. Propofol post-conditioning protects the blood brain barrier by decreasing matrix metalloproteinase-9 and aquaporin-4 expression and improves the neurobehavioral outcome in a rat model of focal cerebral ischemia-reperfusion injury. Mol. Med. Rep. 2015, 12, 2049–2055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altay, O.; Suzuki, H.; Hasegawa, Y.; Caner, B.; Krafft, P.R.; Fujii, M.; Tang, J.; Zhang, J.H. Isoflurane attenuates blood-brain barrier disruption in ipsilateral hemisphere after subarachnoid hemorrhage in mice. Stroke 2012, 43, 2513–2516. [Google Scholar] [CrossRef] [Green Version]
- Thal, S.C.; Luh, C.; Schaible, E.V.; Timaru-Kast, R.; Hedrich, J.; Luhmann, H.J.; Engelhard, K.; Zehendner, C.M. Volatile anesthetics influence blood-brain barrier integrity by modulation of tight junction protein expression in traumatic brain injury. PLoS ONE 2012, 7, e50752. [Google Scholar] [CrossRef] [Green Version]
- Hu, N.; Guo, D.; Wang, H.; Xie, K.; Wang, C.; Li, Y.; Wang, C.; Wang, C.; Yu, Y.; Wang, G. Involvement of the blood-brain barrier opening in cognitive decline in aged rats following orthopedic surgery and high concentration of sevoflurane inhalation. Brain Res. 2014, 1551, 13–24. [Google Scholar] [CrossRef]
- Patel, J.; Chowdhury, E.A.; Noorani, B.; Bickel, U.; Huang, J. Isoflurane increases cell membrane fluidity significantly at clinical concentrations. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183140. [Google Scholar] [CrossRef]
- Cao, Y.; Ni, C.; Li, Z.; Li, L.; Liu, Y.; Wang, C.; Zhong, Y.; Cui, D.; Guo, X. Isoflurane anesthesia results in reversible ultrastructure and occludin tight junction protein expression changes in hippocampal blood-brain barrier in aged rats. Neurosci. Lett. 2015, 587, 51–56. [Google Scholar] [CrossRef]
- Wang, N.; Wang, M. Dexmedetomidine suppresses sevoflurane anesthesia-induced neuroinflammation through activation of the PI3K/Akt/mTOR pathway. BMC Anesthesiol. 2019, 19, 134. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Sun, Y.M.; Huang, H.; Chen, C.; Wan, J.; Ma, L.H.; Sun, Y.Y.; Miao, H.H.; Wu, Y.Q. Sirtuin 3 protects against anesthesia/surgery-induced cognitive decline in aged mice by suppressing hippocampal neuroinflammation. J. Neuroinflamm. 2021, 18, 41. [Google Scholar] [CrossRef] [PubMed]
- Xie, N.; Li, H.; Wei, D.; LeSage, G.; Chen, L.; Wang, S.; Zhang, Y.; Chi, L.; Ferslew, K.; He, L.; et al. Glycogen synthase kinase-3 and p38 MAPK are required for opioid-induced microglia apoptosis. Neuropharmacology 2010, 59, 444–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merighi, S.; Gessi, S.; Varani, K.; Fazzi, D.; Stefanelli, A.; Borea, P.A. Morphine mediates a proinflammatory phenotype via mu-opioid receptor-PKCvarepsilon-Akt-ERK1/2 signaling pathway in activated microglial cells. Biochem. Pharmacol. 2013, 86, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Chu, H.; Jiang, Y.; Yuan, L. Morphine enhances IL-1beta release through toll-like receptor 4-mediated endocytic pathway in microglia. Purinergic Signal. 2016, 12, 637–645. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, J.M.; Bilbo, S.D. Adolescent morphine exposure affects long-term microglial function and later-life relapse liability in a model of addiction. J. Neurosci. 2013, 33, 961–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.; Kong, H.; Pan, Y.B.; Jiang, L.; Pan, X.X.; Hu, L.; Qian, Y.N.; Jiang, C.Y.; Liu, W.T. Procyanidins alleviates morphine tolerance by inhibiting activation of NLRP3 inflammasome in microglia. J. Neuroinflamm. 2016, 13, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grace, P.M.; Strand, K.A.; Galer, E.L.; Urban, D.J.; Wang, X.; Baratta, M.V.; Fabisiak, T.J.; Anderson, N.D.; Cheng, K.; Greene, L.I.; et al. Morphine paradoxically prolongs neuropathic pain in rats by amplifying spinal NLRP3 inflammasome activation. Proc. Natl. Acad. Sci. USA 2016, 113, E3441–E3450. [Google Scholar] [CrossRef] [Green Version]
- Grace, P.M.; Strand, K.A.; Galer, E.L.; Rice, K.C.; Maier, S.F.; Watkins, L.R. Protraction of neuropathic pain by morphine is mediated by spinal damage associated molecular patterns (DAMPs) in male rats. Brain. Behav. Immun. 2018, 72, 45–50. [Google Scholar] [CrossRef]
- Zaidi, S.A.H.; Thakore, N.; Singh, S.; Guzman, W.; Mehrotra, S.; Gangaraju, V.; Husain, S. Histone Deacetylases Regulation by delta-Opioids in Human Optic Nerve Head Astrocytes. Investig. Ophthalmol. Vis. Sci. 2020, 61, 17. [Google Scholar] [CrossRef]
- Ikeda, H.; Miyatake, M.; Koshikawa, N.; Ochiai, K.; Yamada, K.; Kiss, A.; Donlin, M.J.; Panneton, W.M.; Churchill, J.D.; Green, M.; et al. Morphine modulation of thrombospondin levels in astrocytes and its implications for neurite outgrowth and synapse formation. J. Biol. Chem. 2010, 285, 38415–38427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peyravian, N.; Dikici, E.; Deo, S.; Toborek, M.; Daunert, S. Opioid antagonists as potential therapeutics for ischemic stroke. Prog. Neurobiol. 2019, 182, 101679. [Google Scholar] [CrossRef] [PubMed]
- Piazza, O.; Staiano, R.I.; De Robertis, E.; Conti, G.; Di Crescenzo, V.; Loffredo, S.; Marone, G.; Marinosci, G.Z.; Cataldi, M.M. Effect of alpha2-Adrenergic Agonists and Antagonists on Cytokine Release from Human Lung Macrophages Cultured In Vitro. Transl. Med. UniSa 2016, 15, 67–73. [Google Scholar]
- Bao, Y.; Zhu, Y.; He, G.; Ni, H.; Liu, C.; Ma, L.; Zhang, L.; Shi, D. Dexmedetomidine Attenuates Neuroinflammation In LPS-Stimulated BV2 Microglia Cells Through Upregulation Of miR-340. Drug Des. Devel. Ther. 2019, 13, 3465–3475. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Hu, S.; Li, Y.; Xue, D.; Wu, X. Dexmedetomidine, an Alpha 2a Adrenergic Receptor Agonist, Mitigates Experimental Autoimmune Encephalomyelitis by Desensitization of CXCR7 in Microglia. Biochemistry 2018, 57, 4197–4205. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Dai, H.; Ding, T.; Dai, A.; Zhang, F.; Yu, L.; Chen, G.; Chen, Z. Effects of dexmedetomidine on the release of glial cell line-derived neurotrophic factor from rat astrocyte cells. Neurochem. Int. 2011, 58, 549–557. [Google Scholar] [CrossRef]
- Degos, V.; Charpentier, T.L.; Chhor, V.; Brissaud, O.; Lebon, S.; Schwendimann, L.; Bednareck, N.; Passemard, S.; Mantz, J.; Gressens, P. Neuroprotective effects of dexmedetomidine against glutamate agonist-induced neuronal cell death are related to increased astrocyte brain-derived neurotrophic factor expression. Anesthesiology 2013, 118, 1123–1132. [Google Scholar] [CrossRef]
- Ning, Q.; Liu, Z.; Wang, X.; Zhang, R.; Zhang, J.; Yang, M.; Sun, H.; Han, F.; Zhao, W.; Zhang, X. Neurodegenerative changes and neuroapoptosis induced by systemic lipopolysaccharide administration are reversed by dexmedetomidine treatment in mice. Neurol. Res. 2017, 39, 357–366. [Google Scholar] [CrossRef]
- Wang, Y.L.; Zhang, Y.; Cai, D.S. Dexmedetomidine Ameliorates Postoperative Cognitive Dysfunction via the MicroRNA-381-Mediated EGR1/p53 Axis. Mol. Neurobiol. 2021, 58, 5052–5066. [Google Scholar] [CrossRef]
- Guo, Y.; Wu, Y.; Li, N.; Wang, Z. Up-regulation of miRNA-151-3p enhanced the neuroprotective effect of dexmedetomidine against beta-amyloid by targeting DAPK-1 and TP53. Exp. Mol. Pathol. 2021, 118, 104587. [Google Scholar] [CrossRef]
- Yang, J.J.; Zhao, Y.H.; Yin, K.W.; Zhang, X.Q.; Liu, J. Dexmedetomidine inhibits inflammatory response and oxidative stress through regulating miR-205-5p by targeting HMGB1 in cerebral ischemic/reperfusion. Immunopharmacol. Immunotoxicol. 2021, 43, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Xu, T.; Jiang, W. Dexmedetomidine protects PC12 cells from ropivacaine injury through miR-381/LRRC4/SDF-1/CXCR4 signaling pathway. Regen. Ther. 2020, 14, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Ramirez, M.A.; Wu, D.; Pryce, G.; Simpson, J.E.; Reijerkerk, A.; King-Robson, J.; Kay, O.; de Vries, H.E.; Hirst, M.C.; Sharrack, B.; et al. MicroRNA-155 negatively affects blood-brain barrier function during neuroinflammation. FASEB J. 2014, 28, 2551–2565. [Google Scholar] [CrossRef] [PubMed]
- Paeschke, N.; von Haefen, C.; Endesfelder, S.; Sifringer, M.; Spies, C.D. Dexmedetomidine Prevents Lipopolysaccharide-Induced MicroRNA Expression in the Adult Rat Brain. Int. J. Mol. Sci. 2017, 18, 1830. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Chen, X. The Crosstalk between the Blood–Brain Barrier Dysfunction and Neuroinflammation after General Anaesthesia. Curr. Issues Mol. Biol. 2022, 44, 5700-5717. https://doi.org/10.3390/cimb44110386
Yang X, Chen X. The Crosstalk between the Blood–Brain Barrier Dysfunction and Neuroinflammation after General Anaesthesia. Current Issues in Molecular Biology. 2022; 44(11):5700-5717. https://doi.org/10.3390/cimb44110386
Chicago/Turabian StyleYang, Xinxin, and Xiangdong Chen. 2022. "The Crosstalk between the Blood–Brain Barrier Dysfunction and Neuroinflammation after General Anaesthesia" Current Issues in Molecular Biology 44, no. 11: 5700-5717. https://doi.org/10.3390/cimb44110386
APA StyleYang, X., & Chen, X. (2022). The Crosstalk between the Blood–Brain Barrier Dysfunction and Neuroinflammation after General Anaesthesia. Current Issues in Molecular Biology, 44(11), 5700-5717. https://doi.org/10.3390/cimb44110386