
Antifibrosis Efficacy of Apo-9-Fucoxanthinone-Contained Sargassum horneri Ethanol Extract on Nasal Polyp: An In Vitro and Ex Vivo Organ Culture Assay

 and
and 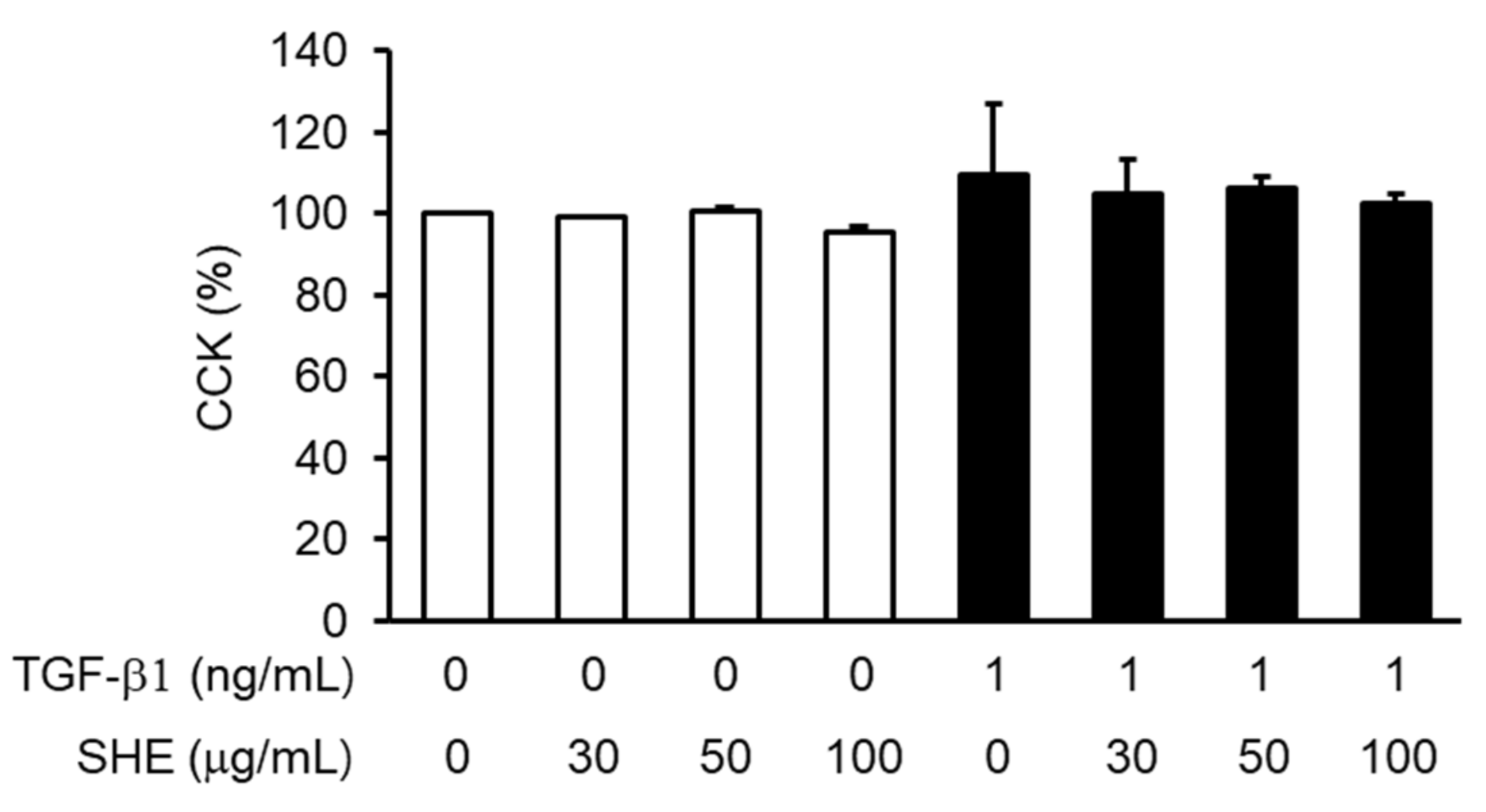{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Preparation of SHE
2.3. NPDF Culture and Cell Viability Assay
2.4. Western Blotting
2.5. Preparation of Nuclear Extracts and EMSA
2.6. Collagen Gel Contraction Assay
2.7. Organ Culture of NPs
2.8. Statistical Analysis
3. Results
3.1. Effects of SHE on the Viability of NPDFs
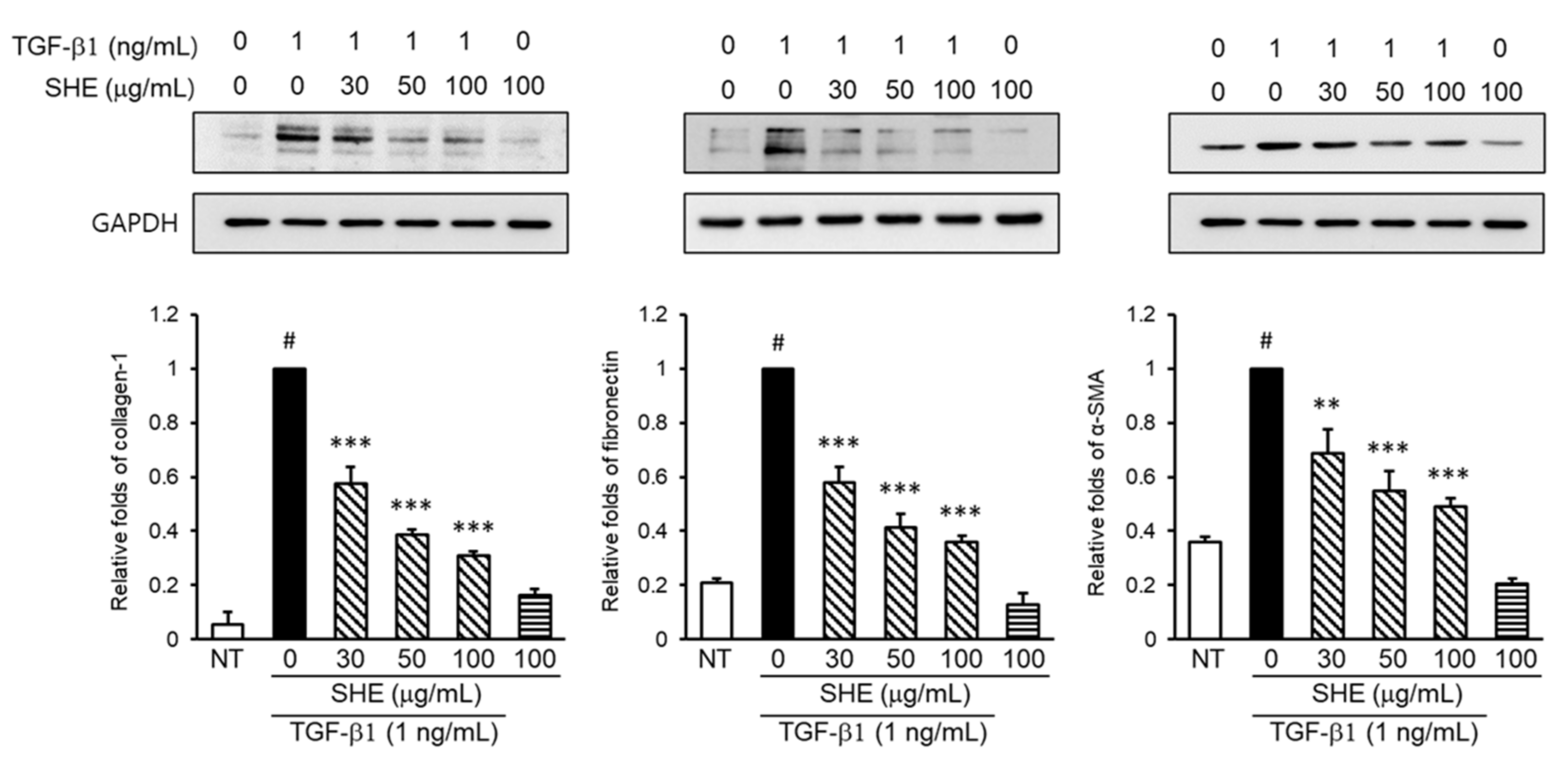
3.2. Effect of SHE on Fibronectin, Collagen-1, and α-SMA Protein Expression in TGF-β1-Stimulated NPDFs
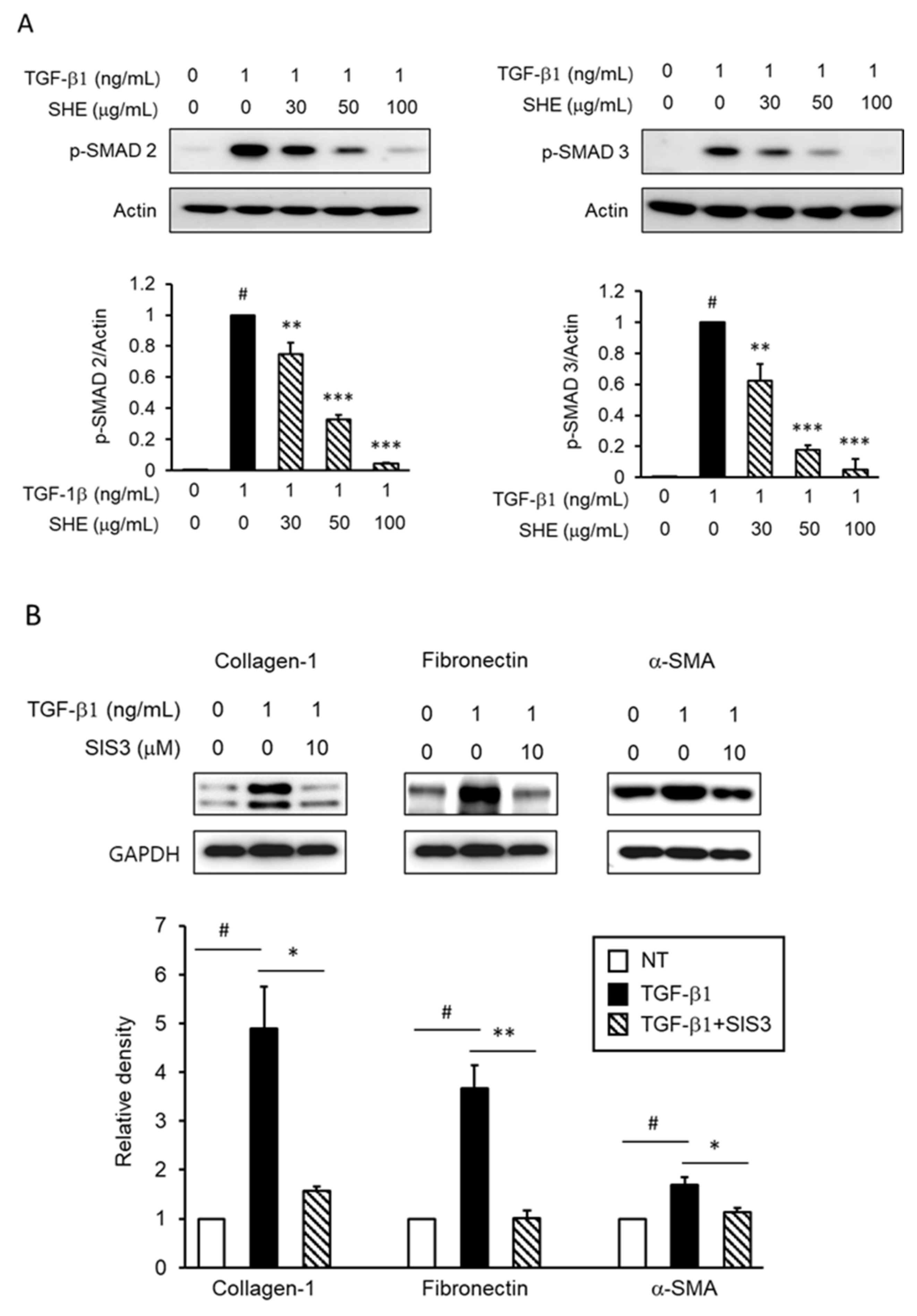
3.3. SHE Inhibits TGF-β1-Stimulated Smad 2/3-Dependent Signaling Pathways
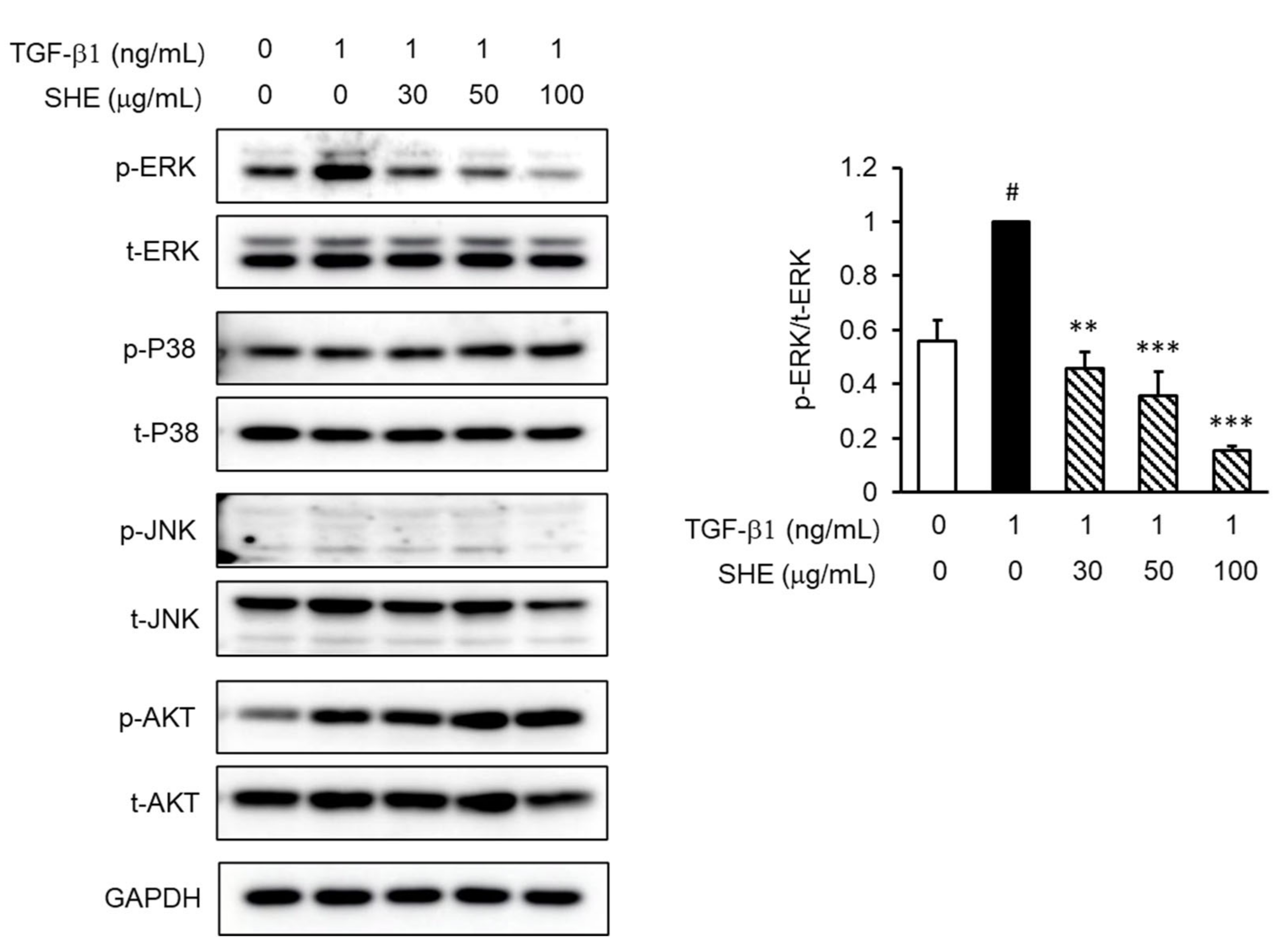
3.4. SHE Inhibits TGF-β1-Stimulated Smad 2/3-Independent Signaling Pathways
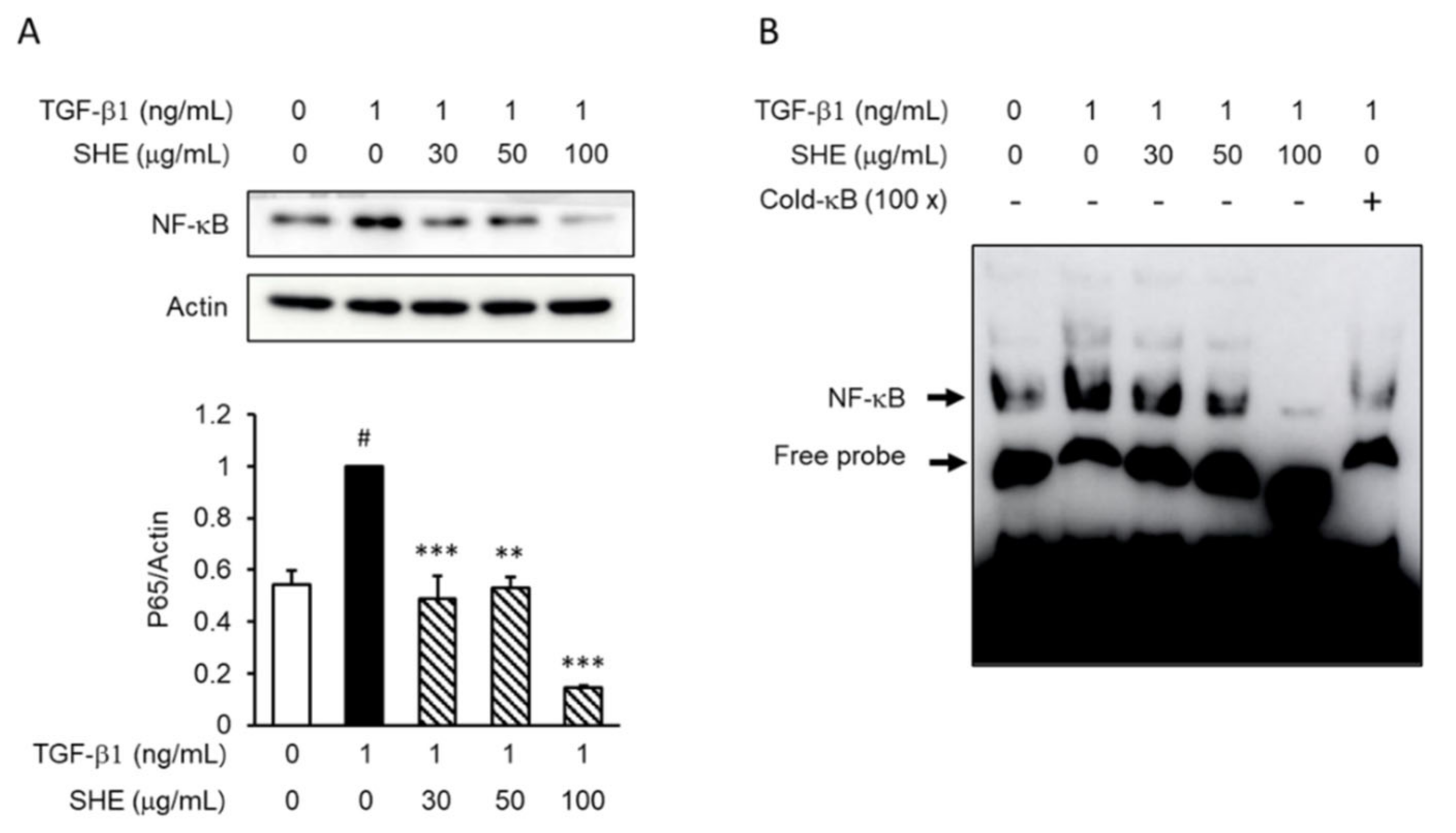
3.5. SHE Inhibits TGF-β1-Induced Nuclear Factor Kappa B (NF-κB) Activation in NPDFs
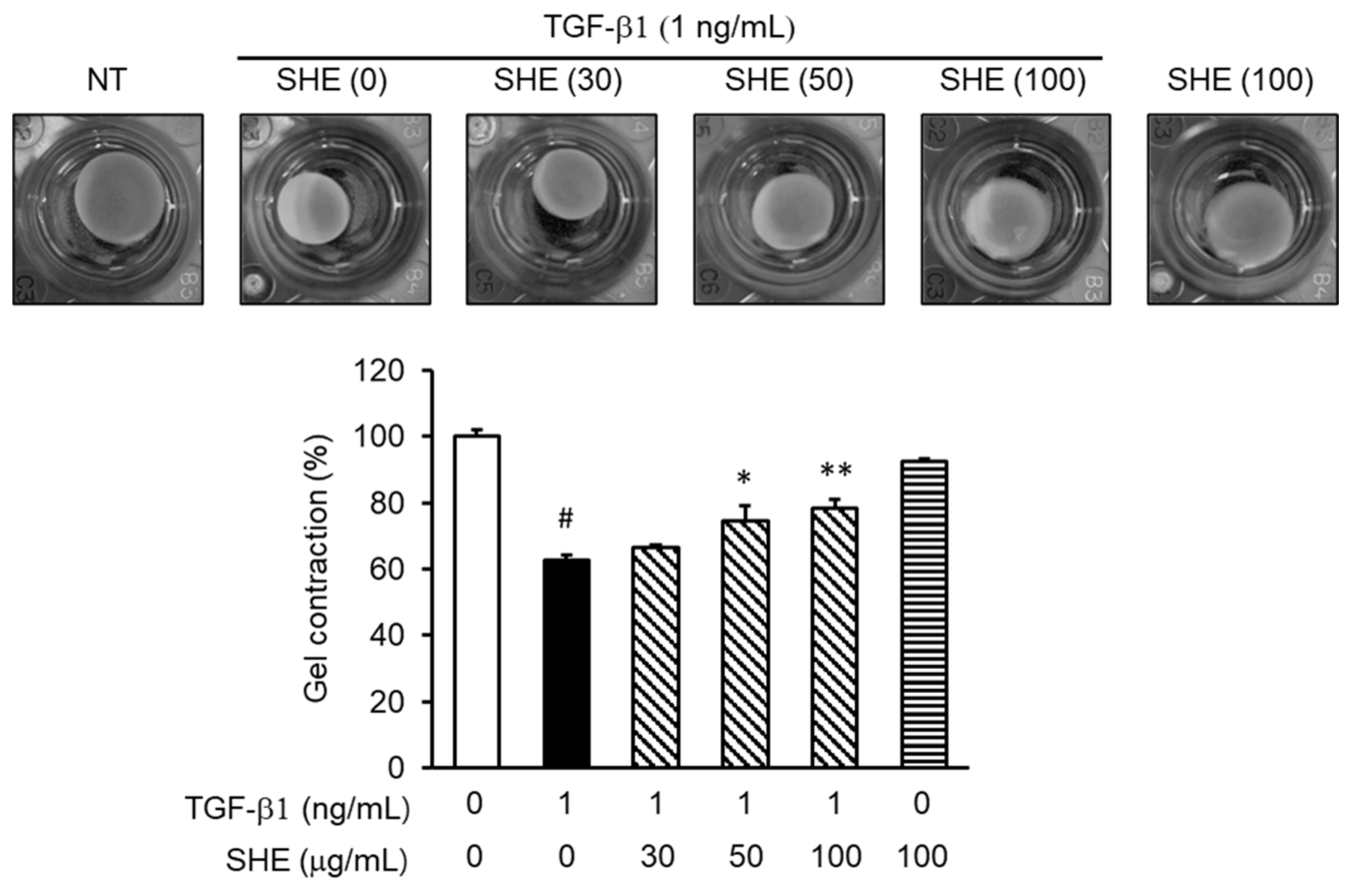
3.6. SHE Inhibits TGF-β1-Stimulated Fibroblast Contractile Activity
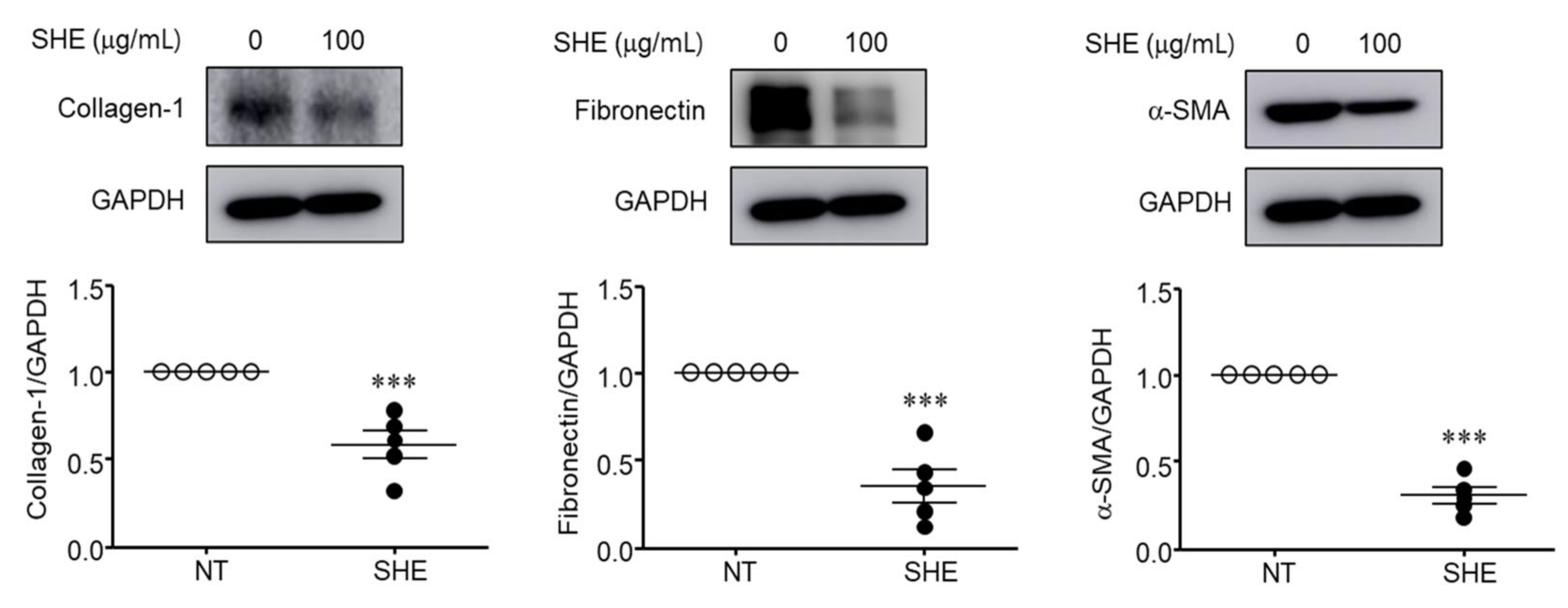
3.7. SHE Inhibits α-SMA, Collagen-1, and Fibronectin Protein Expression in Nasal Polyps Organ Culture
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Naclerio, R.M.; Bachert, C.; Baraniuk, J.N. Pathophysiology of nasal congestion. Int. J. Gen. Med. 2010, 3, 47–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, I.H.; Park, S.J.; Cho, J.S.; Moon, Y.M.; Kim, T.H.; Lee, S.H.; Lee, H.M. Role of reactive oxygen species in transforming growth factor beta1-induced alpha smooth-muscle actin and collagen production in nasal polyp-derived fibroblasts. Int. Arch. Allergy Immunol. 2012, 159, 278–286. [Google Scholar] [CrossRef] [PubMed]
- El Banhawy, O.; Al Abri, R.; Khalil, Y.; Shafy, I.; Fayaz, F. Update of pathogenesis and management of nasal polyposis. Menoufia Med. J. 2016, 29, 469–477. [Google Scholar] [CrossRef]
- Zaravinos, A.; Soufla, G.; Bizakis, J.; Spandidos, D.A. Expression analysis of VEGFA, FGF2, TGFbeta1, EGF and IGF1 in human nasal polyposis. Oncol. Rep. 2008, 19, 385–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soy, F.K.; Pinar, E.; Imre, A.; Calli, C.; Calli, A.; Oncel, S. Histopathologic parameters in chronic rhinosinusitis with nasal polyposis: Impact on quality of life outcomes. Int. Forum Allergy Rhinol. 2013, 3, 828–833. [Google Scholar] [CrossRef]
- Enache, I.; IoniȚĂ, E.; Mitroi, M.; Anghelina, F.; Mogoant, Ă.C.; Ciolofan, S.; CĂpitĂnescu, A.; Stepan, A.; Simionescu, C. Histopathological features of chronic rhinosinusitis with nasal allergic polyps. Curr. Health Sci. J. 2020, 46, 66–71. [Google Scholar] [CrossRef]
- Klingberg, F.; Hinz, B.; White, E.S. The myofibroblast matrix: Implications for tissue repair and fibrosis. J. Pathol. 2013, 229, 298–309. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.S.; Han, I.H.; Lee, H.R.; Lee, H.M. Prostaglandin E2 induces IL-6 and IL-8 production by the EP receptors/Akt/NF-κB pathways in nasal polyp-derived fibroblasts. Allergy Asthma Immunol. Res. 2014, 6, 449–457. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, T.; Yamane, H.; Nakai, Y.; Shigeta, T.; Takashima, T.; Takeda, Z. Comparative assessment of cell proliferation and accumulation of extracellular matrix in nasal polyps. Acta Otolaryngol. 1998, 538, 205–208. [Google Scholar] [CrossRef]
- Ravikanth, M.; Soujanya, P.; Manjunath, K.; Saraswathi, T.R.; Ramachandran, C.R. Heterogenecity of fibroblasts. J. Oral Maxillofac. Pathol. 2011, 15, 247–250. [Google Scholar] [CrossRef]
- Darby, I.A.; Laverdet, B.; Bonté, F.; Desmoulière, A. Fibroblasts and myofibroblasts in wound healing. Clin. Cosmet. Investig. Dermatol. 2014, 7, 301–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suleria, H.A.; Osborne, S.; Masci, P.; Gobe, G. Marine-based nutraceuticals: An innovative trend in the food and supplement industries. Mar. Drugs 2015, 13, 6336–6351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.C.; Hou, M.F.; Huang, H.W.; Chang, F.R.; Yeh, C.C.; Tang, J.Y.; Chang, H.W. Marine algal natural products with anti-oxidative, anti-inflammatory, and anti-cancer properties. Cancer Cell Int. 2013, 13, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijesinghe, W.A.; Jeon, Y.J. Exploiting biological activities of brown seaweed Ecklonia cava for potential industrial applications: A review. Int. J. Food Sci. Nutr. 2012, 63, 225–235. [Google Scholar] [CrossRef]
- Jung, H.A.; Jin, S.E.; Ahn, B.R.; Lee, C.M.; Choi, J.S. Anti-inflammatory activity of edible brown alga Eisenia bicyclis and its constituents fucosterol and phlorotannins in LPS-stimulated RAW264.7 macrophages. Food Chem. Toxicol. 2013, 59, 199–206. [Google Scholar] [CrossRef]
- Kim, H.-S.; Sanjeewa, K.; Fernando, I.; Ryu, B.; Yang, H.-W.; Ahn, G.; Kang, M.C.; Heo, S.-J.; Je, J.-G.; Jeon, Y.-J. A comparative study of Sargassum horneri Korea and China strains collected along the coast of Jeju Island South Korea: Its components and bioactive properties. Algae 2018, 33, 341–349. [Google Scholar] [CrossRef]
- Yamaguchi, M. Regulatory mechanism of food factors in bone metabolism and prevention of osteoporosis. Yakugaku Zasshi 2006, 126, 1117–1137. [Google Scholar] [CrossRef] [Green Version]
- Han, E.J.; Kim, H.S.; Sanjeewa, K.K.A.; Jung, K.; Jee, Y.; Jeon, Y.J.; Fernando, I.P.S.; Ahn, G. Sargassum horneri as a functional food ameliorated IgE/BSA-induced mast cell activation and passive cutaneous anaphylaxis in mice. Mar. Drugs 2020, 18, 594. [Google Scholar] [CrossRef]
- Yoshioka, H.; Ishida, M.; Nishi, K.; Oda, H.; Toyohara, H.; Sugahara, T. Studies on anti-allergic activity of Sargassum horneri extract. J. Funct. Foods 2014, 10, 154–160. [Google Scholar] [CrossRef]
- Rasin, A.B.; Silchenko, A.S.; Kusaykin, M.I.; Malyarenko, O.S.; Zueva, A.O.; Kalinovsky, A.I.; Airong, J.; Surits, V.V.; Ermakova, S.P. Enzymatic transformation and anti-tumor activity of Sargassum horneri fucoidan. Carbohydr. Polym. 2020, 246, 116635. [Google Scholar] [CrossRef]
- Athukorala, Y.; Lee, K.W.; Kim, S.K.; Jeon, Y.J. Anticoagulant activity of marine green and brown algae collected from Jeju Island in Korea. Bioresour. Technol. 2007, 98, 1711–1716. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.S.; Choi, I.W.; Han, M.H.; Hong, S.H.; Kim, S.O.; Kim, G.Y.; Hwang, H.J.; Kim, B.W.; Choi, B.T.; Kim, C.M.; et al. Sargassum horneri methanol extract rescues C2C12 murine skeletal muscle cells from oxidative stress-induced cytotoxicity through Nrf2-mediated upregulation of heme oxygenase-1. BMC Complement. Altern. Med. 2015, 15, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preeprame, S.; Hayashi, K.; Lee, J.B.; Sankawa, U.; Hayashi, T. A novel antivirally active fucan sulfate derived from an edible brown alga, Sargassum horneri. Chem. Pharm. Bull. 2001, 49, 484–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, E.K.; Lee, S.J.; Ha, D.S.; Park, C.S. Sargassum Golden Tides in the Shinan-gun and Jeju Island, Korea. Korean J. Fish Aquat. Sci. 2016, 49, 689–693. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.S.; Lee, C.M.; Park, S.K.; Yim, M.J.; Lee, J.M.; Choi, G.; Yoo, J.S.; Jung, W.K.; Park, S.; Seo, S.K.; et al. Anti-inhibitory potential of an ethanolic extract of Distromium decumbens on pro-inflammatory cytokine production in Pseudomonas aeruginosa lipopolysaccharide-stimulated nasal polyp-derived fibroblasts. Int. J. Mol. Med. 2017, 40, 1950–1956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, H.; Lee, D.S.; Park, S.K.; Choi, J.S.; Jung, W.K.; Park, W.S.; Choi, I.W. Fucoxanthin inhibits myofibroblast differentiation and extracellular matrix production in nasal polyp-derived fibroblasts via modulation of Smad-dependent and Smad-independent signaling pathways. Mar. Drugs 2018, 16, 323. [Google Scholar] [CrossRef] [Green Version]
- Park, S.K.; Lee, W.J.; Yang, Y.I. Organ culture at the air-liquid interface maintains structural and functional integrities of inflammatory and fibrovascular cells of nasal polyps. Am. J. Rhinol. 2007, 21, 402–407. [Google Scholar] [CrossRef]
- Pohlers, D.; Brenmoehl, J.; Löffler, I.; Müller, C.K.; Leipner, C.; Schultze-Mosgau, S.; Stallmach, A.; Kinne, R.W.; Wolf, G. TGF-beta and fibrosis in different organs—Molecular pathway imprints. Biochim. Biophys. Acta 2009, 1792, 746–756. [Google Scholar] [CrossRef] [Green Version]
- Sugiura, H.; Ichikawa, T.; Liu, X.; Kobayashi, T.; Wang, X.Q.; Kawasaki, S.; Togo, S.; Kamio, K.; Mao, L.; Ann, Y.; et al. N-acetyl-l-cysteine inhibits TGF-β1-induced profibrotic responses in fibroblasts. Pulm. Pharmacol. Ther. 2009, 22, 487–491. [Google Scholar] [CrossRef]
- Prud’homme, G.J. Pathobiology of transforming growth factor β in cancer, fibrosis and immunologic disease, and therapeutic considerations. Lab. Investig. 2007, 87, 1077–1091. [Google Scholar] [CrossRef]
- Malakpour-Permlid, A.; Buzzi, I.; Hegardt, C.; Johansson, F.; Oredsson, S. Identification of extracellular matrix proteins secreted by human dermal fibroblasts cultured in 3D electrospun scaffolds. Sci. Rep. 2021, 11, 6655. [Google Scholar] [CrossRef] [PubMed]
- Coste, A.; Lefaucheur, J.P.; Wang, Q.P.; Lesprit, E.; Poron, F.; Peynegre, R.; Escudier, E. Expression of the Transforming Growth Factor β Isoforms in Inflammatory Cells of Nasal Polyps. Arch. Otolaryngol. Head Neck Surg. 1998, 124, 1361–1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walton, K.L.; Johnson, K.E.; Harrison, C.A. Targeting TGF-β mediated SMAD signaling for the prevention of fibrosis. Front. Pharmacol. 2017, 8, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Dong, X.; Wang, L.; Shan, L.; Li, T.; Xu, W.; Ding, Y.; Lai, M.; Lin, X.; Dai, M.; et al. Casticin attenuates liver fibrosis and hepatic stellate cell activation by blocking TGF-β/Smad signaling pathway. Oncotarget 2017, 8, 56267–56280. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.S.; Kang, J.H.; Shin, J.M.; Park, I.H.; Lee, H.M. Inhibitory effect of delphinidin on extracellular matrix production via the MAPK/NF-κB pathway in nasal polyp-derived fibroblasts. Allergy Asthma Immunol. Res. 2015, 7, 276–282. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.J.; Zhang, Y.L.; Kim, D.K.; Rhee, C.S.; Kim, D.Y. The role of NF-kappa B in chronic rhinosinusitis with nasal polyps. Allergy Asthma Immunol. Res. 2019, 11, 806–817. [Google Scholar] [CrossRef]
- Shinde, A.V.; Humeres, C.; Frangogiannis, N.G. The role of α-smooth muscle actin in fibroblast-mediated matrix contraction and remodeling. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 298–309. [Google Scholar] [CrossRef]
- Zhu, Y.K.; Umino, T.; Liu, X.D.; Wang, H.J.; Romberger, D.J.; Spurzem, J.R.; Rennard, S.I. Contraction of fibroblast-containing collagen gels: Initial collagen concentration regulates the degree of contraction and cell survival. In Vitro Cell. Dev. Biol. Anim. 2001, 37, 10–16. [Google Scholar] [CrossRef]
- Kim, H.-S.; Wang, L.; Jayawardena, T.U.; Kim, E.A.; Heo, S.J.; Shanura, I.P.S.; Lee, J.H.; Jeon, Y.J. High-performance centrifugal partition chromatography (HPCPC) for efficient isolation of diphlorethohydroxycarmalol (DPHC) and screening of its antioxidant activity in a zebrafish model. Process Biochem. 2020, 88, 189–196. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yim, M.-J.; Lee, J.M.; Ko, S.-C.; Kim, H.-S.; Kim, J.-Y.; Park, S.K.; Lee, D.-S.; Choi, I.-W. Antifibrosis Efficacy of Apo-9-Fucoxanthinone-Contained Sargassum horneri Ethanol Extract on Nasal Polyp: An In Vitro and Ex Vivo Organ Culture Assay. Curr. Issues Mol. Biol. 2022, 44, 5815-5826. https://doi.org/10.3390/cimb44110395
Yim M-J, Lee JM, Ko S-C, Kim H-S, Kim J-Y, Park SK, Lee D-S, Choi I-W. Antifibrosis Efficacy of Apo-9-Fucoxanthinone-Contained Sargassum horneri Ethanol Extract on Nasal Polyp: An In Vitro and Ex Vivo Organ Culture Assay. Current Issues in Molecular Biology. 2022; 44(11):5815-5826. https://doi.org/10.3390/cimb44110395
Chicago/Turabian StyleYim, Mi-Jin, Jeong Min Lee, Seok-Chun Ko, Hyun-Soo Kim, Ji-Yul Kim, Seong Kook Park, Dae-Sung Lee, and Il-Whan Choi. 2022. "Antifibrosis Efficacy of Apo-9-Fucoxanthinone-Contained Sargassum horneri Ethanol Extract on Nasal Polyp: An In Vitro and Ex Vivo Organ Culture Assay" Current Issues in Molecular Biology 44, no. 11: 5815-5826. https://doi.org/10.3390/cimb44110395
APA StyleYim, M. -J., Lee, J. M., Ko, S. -C., Kim, H. -S., Kim, J. -Y., Park, S. K., Lee, D. -S., & Choi, I. -W. (2022). Antifibrosis Efficacy of Apo-9-Fucoxanthinone-Contained Sargassum horneri Ethanol Extract on Nasal Polyp: An In Vitro and Ex Vivo Organ Culture Assay. Current Issues in Molecular Biology, 44(11), 5815-5826. https://doi.org/10.3390/cimb44110395