_Kim.png)
Phosphodiesterase 5 Inhibitor Potentiates Epigallocatechin 3-O-Gallate-Induced Apoptotic Cell Death via Activation of the cGMP Signaling Pathway in Caco-2 Cells

 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Cell Viability
2.2. Analyzed Apoptotic Cell Death
2.3. The siRNAs Targeting PDE5
2.4. Western Blotting Analysis
2.5. Statistical Analysis
3. Results
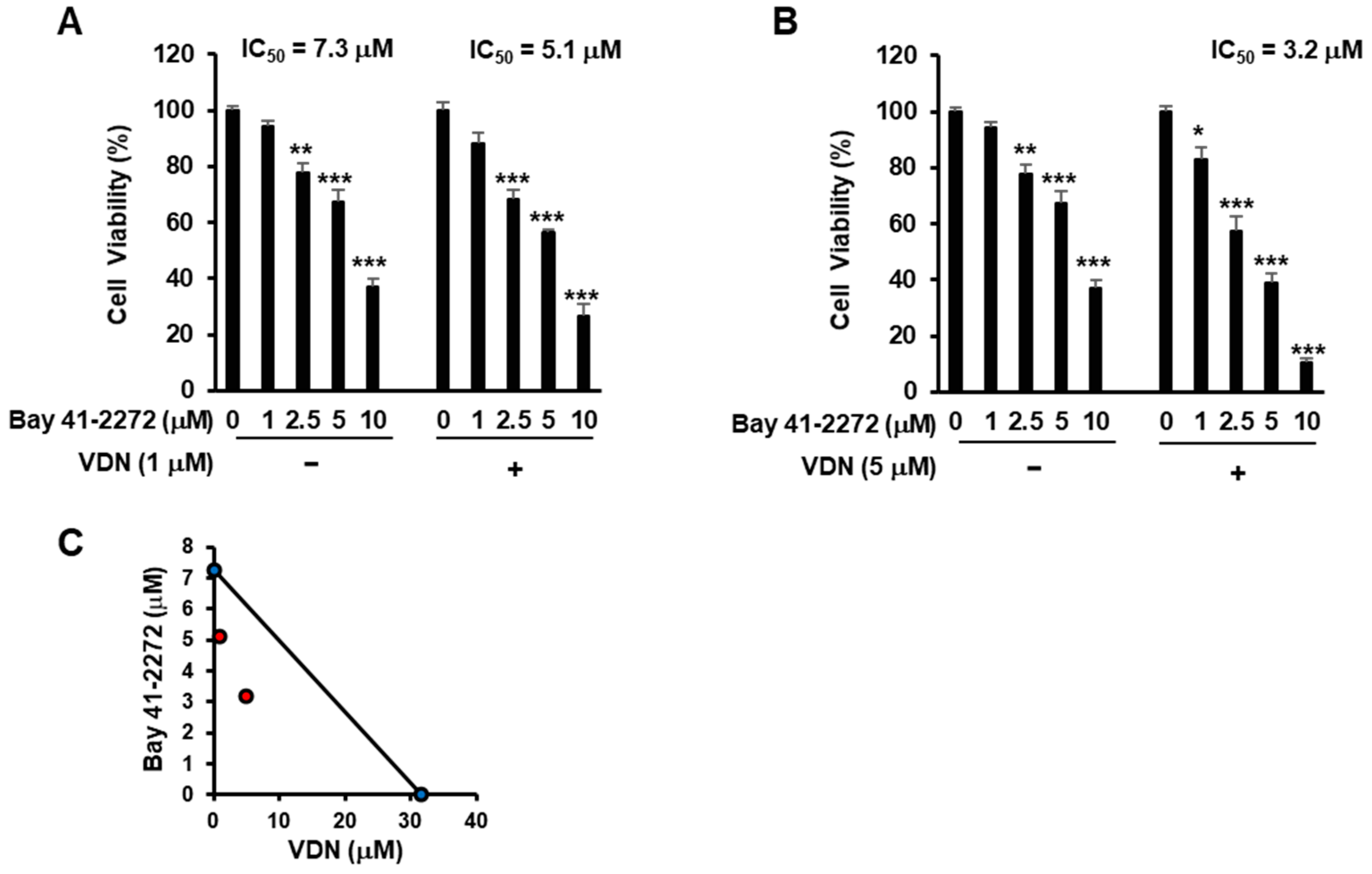
3.1. Combination of EGCG and VDN Synergistically Induces Cell Death in Colorectal Adenocarcinoma
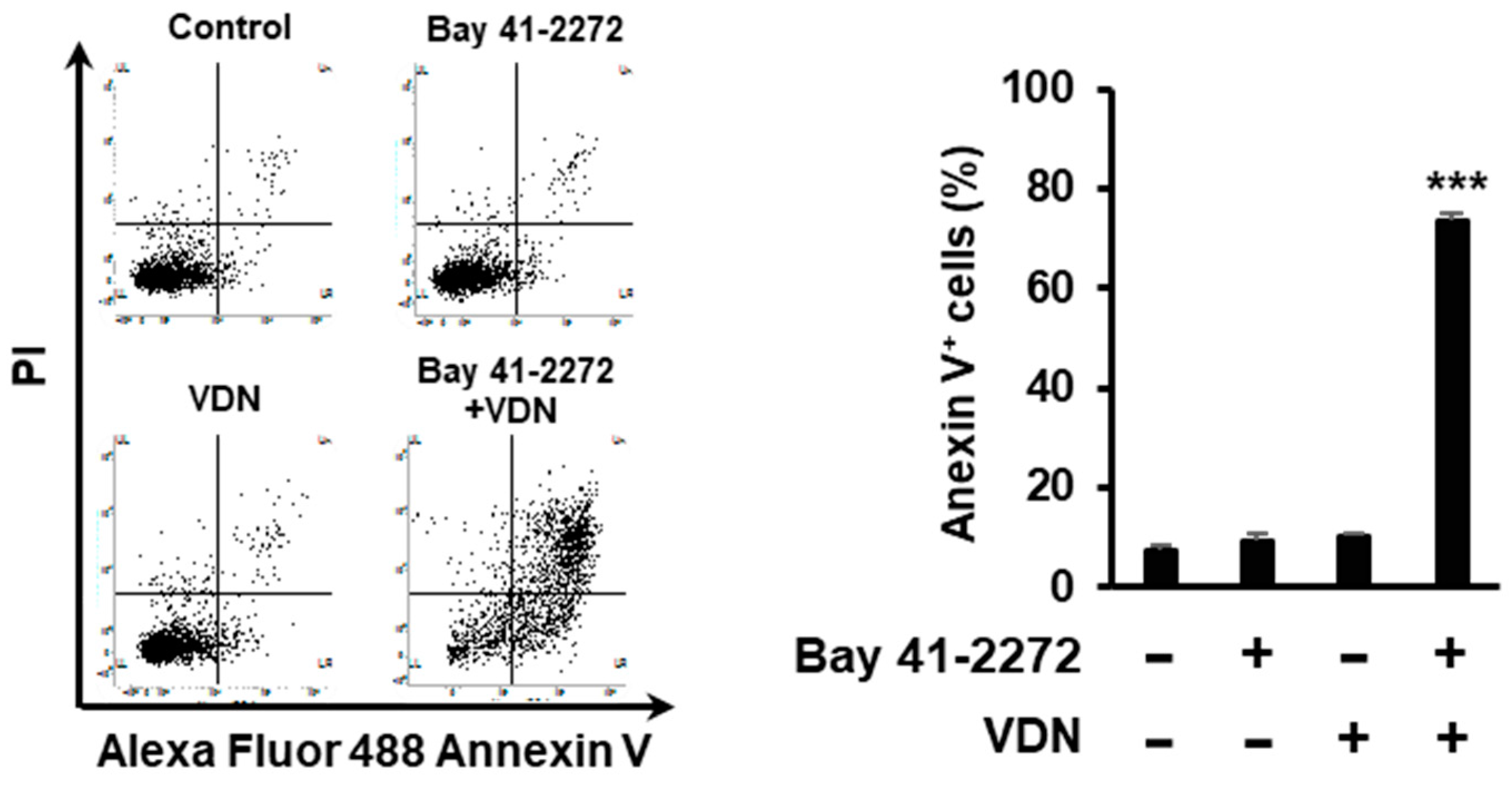
3.2. Combination of EGCG and VDN Induces Apoptosis in Colorectal Adenocarcinoma
3.3. Expression of PDE5 Attenuates EGCG-Induced Apoptotic Cell Death in Colorectal Adenocarcinoma
3.4. Activation of cGMP with Inhibition of PDE5 Activity Induces Cell Death in Colorectal Adenocarcinoma
3.5. Inhibition of PDE5 Potentiates cGMP-Mediated Apoptosis in Colorectal Adenocarcinoma
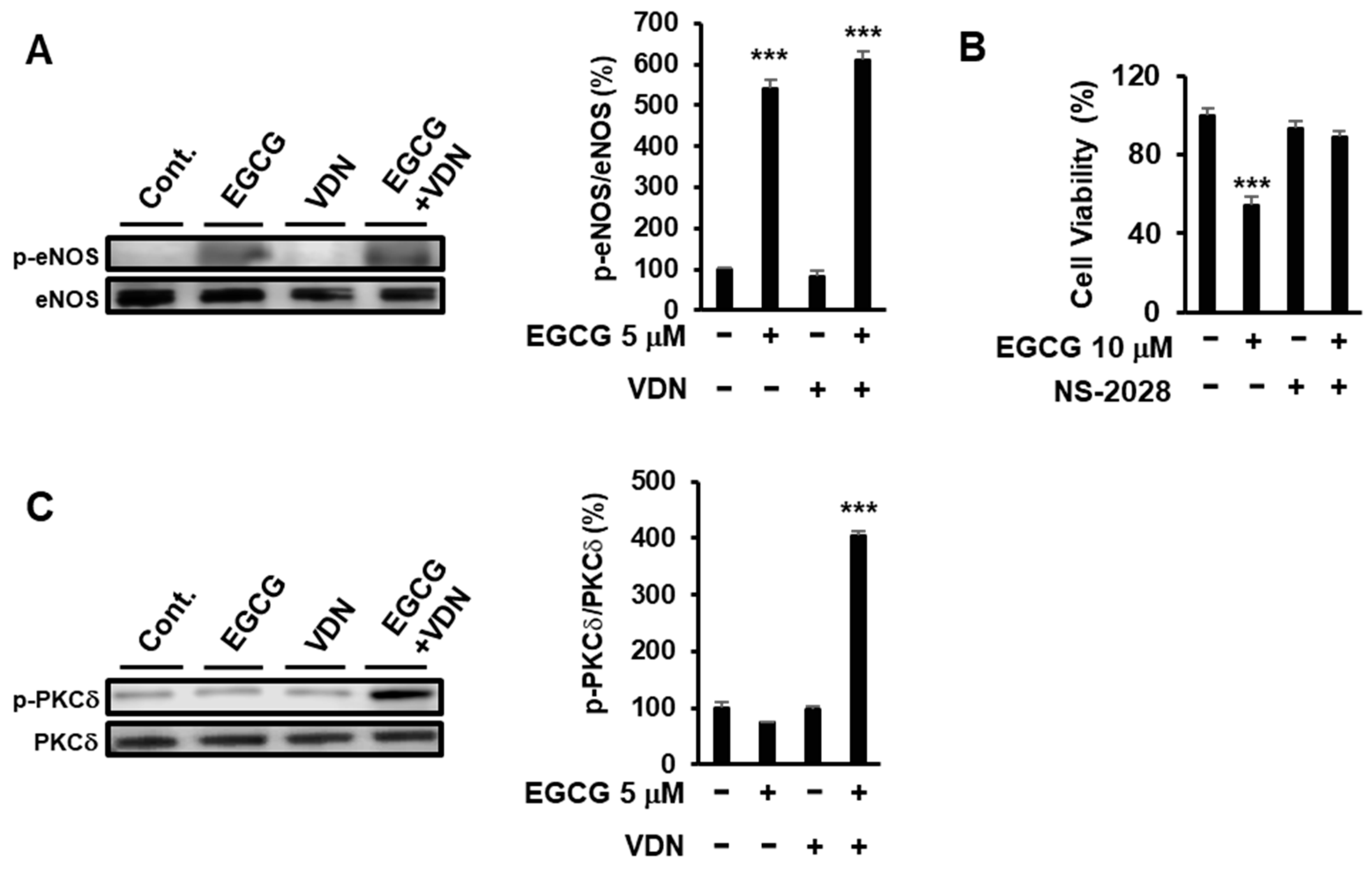
3.6. Inhibition of PDE5 Potentiates EGCG-Induced Apoptotic Cell Death via the eNOS/cGMP/PKCδ Signaling Pathway
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Khan, N.; Mukhtar, H. Tea polyphenols for health promotion. Life Sci. 2007, 81, 519–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tachibana, H.; Koga, K.; Fujimura, Y.; Yamada, K. A receptor for green tea polyphenol EGCG. Nat. Struct. Mol. Biol. 2004, 11, 380–381. [Google Scholar] [CrossRef] [PubMed]
- Kumazoe, M.; Sugihara, K.; Tsukamoto, S.; Huang, Y.; Tsurudome, Y.; Suzuki, T.; Suemasu, Y.; Ueda, N.; Yamashita, S.; Kim, Y.; et al. 67-kDa laminin receptor increases cGMP to induce cancer-selective apoptosis. J. Clin. Investig. 2013, 123, 787–799. [Google Scholar] [CrossRef] [PubMed]
- Shanafelt, T.D.; Call, T.G.; Zent, C.S.; LaPlant, B.; Bowen, D.A.; Roos, M.; Secreto, C.R.; Ghosh, A.K.; Kabat, B.F.; Lee, M.J.; et al. Phase I trial of daily oral polyphenon E in patients with asymptomatic rai stage 0 to II chronic lymphocytic leukemia. J. Clin. Oncol. 2009, 27, 3808–3814. [Google Scholar] [CrossRef]
- Álvarez, E.; Campos-Toimil, M.; Justiniano-Basaran, H.; Lugnier, C.; Orallo, F. Study of the mechanisms involved in the vasorelaxation induced by (-)-epigallocatechin-3-gallate in rat aorta. Br. J. Pharmacol. 2006, 147, 269–280. [Google Scholar] [CrossRef] [Green Version]
- Lugnier, C. Cyclic nucleotide phosphodiesterase (PDE) superfamily: A new target for the development of specific therapeutic agents. Pharmacol. Ther. 2006, 109, 366–398. [Google Scholar] [CrossRef]
- Keravis, T.; Lugnier, C. Cyclic nucleotide phosphodiesterase (PDE) isozymes as targets of the intracellular signalling network: Benefits of PDE inhibitors in various diseases and perspectives for future therapeutic developments. Br. J. Pharmacol. 2012, 165, 1288–1305. [Google Scholar] [CrossRef] [Green Version]
- Porst, H.; Rosen, R.; Padma-Nathan, H.; Goldstein, I.; Giuliano, F.; Ulbrich, E. The efficacy and tolerability of vardenafil, a new, oral, selective phosphodiesterase type 5 inhibitor, in patients with erectile dysfunction: The first at-home clinical trial. Int. J. Impot. Res. 2001, 13, 192–199. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.A.; Lie, J.D. Phosphodiesterase-5 (PDE5) inhibitors in the management of erectile dysfunction. Pharm. Ther. 2013, 38, 407. [Google Scholar]
- Linder, A.E.; McCluskey, L.P.; Cole, K.R.; Lanning, K.M.; Webb, R.C. Dynamic association of nitric oxide downstream signaling molecules with endothelial Caveolin-1 in rat aorta. J. Pharmacol. Exp. Ther. 2005, 314, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Russwurm, M.; Wittau, N.; Koesling, D. Guanylyl cyclase/PSD-95 interaction: Targeting of the nitric oxide-sensitive α2β1 guanylyl cyclase to synaptic membranes. J. Biol. Chem. 2001, 276, 44647–44652. [Google Scholar] [CrossRef] [PubMed]
- Schoser, B.G.H.; Behrends, S. Soluble guanylyl cyclase is localized at the neuromuscular junction in human skeletal muscle. Neuroreport 2001, 12, 979–981. [Google Scholar] [CrossRef] [PubMed]
- Zabel, U.; Kleinschnitz, C.; Oh, P.; Nedvetsky, P.; Smolenski, A.; Müller, H.; Kronich, P.; Kugler, P.; Walters, U.; Schnitzer, J.E.; et al. Calcium-dependent membrane association sensitizes soluble guanylyl cyclase to nitric oxide. Nat. Cell Biol. 2002, 4, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, S.; Hirotsu, K.; Kumazoe, M.; Goto, Y.; Sugihara, K.; Suda, T.; Tsurudome, Y.; Suzuki, T.; Yamashita, S.; Kim, Y.; et al. Green tea polyphenol EGCG induces lipid-raft clustering and apoptotic cell death by activating protein kinase Cδ and acid sphingomyelinase through a 67 kDa laminin receptor in multiple myeloma cells. Biochem. J. 2012, 443, 525–534. [Google Scholar] [CrossRef]
- Naganuma, T.; Kuriyama, S.; Kakizaki, M.; Sone, T.; Nakaya, N.; Ohmori-Matsuda, K.; Hozawa, A.; Nishino, Y.; Tsuji, I. Green tea consumption and hematologic malignancies in Japan: The Ohsaki study. Am. J. Epidemiol. 2009, 170, 30–738. [Google Scholar] [CrossRef] [Green Version]
- Umeda, D.; Yano, S.; Yamada, K.; Tachibana, H. Involvement of 67-kDa laminin receptor-mediated myosin phosphatase activation in antiproliferative effect of epigallocatechin-3-O-gallate at a physiological concentration on Caco-2 colon cancer cells. Biochem. Biophys. Res. Commun. 2008, 371, 172–176. [Google Scholar] [CrossRef]
- Bae, J.; Kumazoe, M.; Murata, K.; Fujimura, Y.; Tachibana, H. Procyanidin C1 Inhibits Melanoma Cell Growth by Activating 67-kDa Laminin Receptor Signaling. Mol. Nutr. Food Res. 2020, 64, e1900986. [Google Scholar] [CrossRef]
- McLarty, J.; Bigelow, R.L.H.; Smith, M.; Elmajian, D.; Ankem, M.; Cardelli, J.A. Tea polyphenols decrease serum levels of prostate-specific antigen, hepatocyte growth factor, and vascular endothelial growth factor in prostate cancer patients and inhibit production of hepatocyte growth factor and vascular endothelial growth factor in vitro. Cancer Prev. Res. 2009, 2, 673–682. [Google Scholar]
- Wilkens, H.; Guth, A.; König, J.; Forestier, N.; Cremers, B.; Hennen, B.; Böhm, M.; Sybrecht, G.W. Effect of inhaled iloprost plus oral sildenafil in patients with primary pulmonary hypertension. Circulation 2001, 104, 1218–1222. [Google Scholar] [CrossRef] [Green Version]
- Guazzi, M.; Vicenzi, M.; Arena, R.; Guazzi, M.D. PDE5 inhibition with sildenafil improves left ventricular diastolic function, cardiac geometry, and clinical status in patients with stable systolic heart failure: Result of a 1-year, prospective, randomized, placebo-controlled study. Circ. Heart Fail. 2011, 4, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Francis, S.H.; Morris, G.Z.; Corbin, J.D. Molecular mechanisms that could contribute to prolonged effectiveness of PDE5 inhibitors to improve erectile function. Int. J. Impot. Res. 2008, 20, 333–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pantziarka, P.; Sukhatme, V.; Crispino, S.; Bouche, G.; Meheus, L.; Sukhatme, V.P. Repurposing drugs in oncology (ReDO)-selective PDE5 inhibitors as anti-cancer agents. Ecancermedicalscience 2018, 12, 824. [Google Scholar] [CrossRef] [PubMed]
- Tinsley, H.N.; Gary, B.D.; Thaiparambil, J.; Li, N.; Lu, W.; Li, Y.; Maxuitenko, Y.Y.; Keeton, A.B.; Piazza, G.A. Colon tumor cell growth-inhibitory activity of sulindac sulfide and other nonsteroidal anti-inflammatory drugs is associated with phosphodiesterase 5 inhibition. Cancer Prev. Res. 2010, 3, 1303–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitt, J.D.; Li, N.; Tinsley, H.N.; Chen, X.; Zhang, W.; Li, Y.; Gary, B.D.; Keeton, A.B.; Xi, Y.; Abadi, A.H.; et al. A novel sulindac derivative that potently suppresses colon tumor cell growth by inhibiting cGMP phosphodiesterase and β-catenin transcriptional activity. Cancer Prev. Res. 2012, 5, 822–833. [Google Scholar] [CrossRef] [Green Version]
- Kumazoe, M.; Kim, Y.; Bae, J.; Takai, M.; Murata, M.; Suemasu, Y.; Sugihara, K.; Yamashita, S.; Tsukamoto, S.; Huang, Y.; et al. Phosphodiesterase 5 inhibitor acts as a potent agent sensitizing acute myeloid leukemia cells to 67-kDa laminin receptor-dependent apoptosis. FEBS Lett. 2013, 587, 3052–3057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, J.; Kumazoe, M.; Yamashita, S.; Tachibana, H. Hydrogen sulphide donors selectively potentiate a green tea polyphenol EGCG-induced apoptosis of multiple myeloma cells. Sci. Rep. 2017, 7, 6665. [Google Scholar] [CrossRef]
- Fajardo, A.M.; Piazza, G.A.; Tinsley, H.N. The role of cyclic nucleotide signaling pathways in cancer: Targets for prevention and treatment. Cancers 2014, 6, 436–458. [Google Scholar] [CrossRef] [Green Version]
- Kumazoe, M.; Takai, M.; Bae, J.; Hiroi, S.; Huang, Y.; Takamatsu, K.; Won, Y.; Yamashita, M.; Hidaka, S.; Yamashita, S.; et al. FOXO3 is essential for CD44 expression in pancreatic cancer cells. Oncogene 2017, 36, 2643–2654. [Google Scholar] [CrossRef]
- Jaumann, M.; Dettling, J.; Gubelt, M.; Zimmermann, U.; Gerling, A.; Paquet-Durand, F.; Feil, S.; Wolpert, S.; Franz, C.; Varakina, K.; et al. CGMP-Prkg1 signaling and Pde5 inhibition shelter cochlear hair cells and hearing function. Nat Med. 2012, 18, 252–259. [Google Scholar] [CrossRef]
- Lambert, J.D.; Kennett, M.J.; Sang, S.; Reuhl, K.R.; Ju, J.; Yang, C.S. Hepatotoxicity of high oral dose (-)-epigallocatechin-3-gallate in mice. Food Chem. Toxicol. 2010, 48, 409–416. [Google Scholar] [CrossRef] [Green Version]
- Jackson, D.N.; Foster, D.A. The enigmatic protein kinase Cδ: Complex roles in cell proliferation and survival. FASEB J. 2004, 18, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Hung, J.H.; Lu, Y.S.; Wang, Y.C.; Ma, Y.H.; Wang, D.S.; Kulp, S.K.; Muthusamy, N.; Byrd, J.C.; Cheng, A.L.; Chen, C.S. FTY720 induces apoptosis in hepatocellular carcinoma cells through activation of protein kinase C δ signaling. Cancer Res. 2008, 68, 1204–1212. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bae, J.; Lee, K.; Park, J.-S.; Jung, J.; Tachibana, H.; Fujimura, Y.; Kumazoe, M.; Lim, J.S.; Cho, Y.-C.; Lee, S.-J.; et al. Phosphodiesterase 5 Inhibitor Potentiates Epigallocatechin 3-O-Gallate-Induced Apoptotic Cell Death via Activation of the cGMP Signaling Pathway in Caco-2 Cells. Curr. Issues Mol. Biol. 2022, 44, 6247-6256. https://doi.org/10.3390/cimb44120426
Bae J, Lee K, Park J-S, Jung J, Tachibana H, Fujimura Y, Kumazoe M, Lim JS, Cho Y-C, Lee S-J, et al. Phosphodiesterase 5 Inhibitor Potentiates Epigallocatechin 3-O-Gallate-Induced Apoptotic Cell Death via Activation of the cGMP Signaling Pathway in Caco-2 Cells. Current Issues in Molecular Biology. 2022; 44(12):6247-6256. https://doi.org/10.3390/cimb44120426
Chicago/Turabian StyleBae, Jaehoon, Kwanwoo Lee, Ji-Sun Park, Jinseok Jung, Hirofumi Tachibana, Yoshinori Fujimura, Motofumi Kumazoe, Jae Sung Lim, Young-Chang Cho, Seung-Jae Lee, and et al. 2022. "Phosphodiesterase 5 Inhibitor Potentiates Epigallocatechin 3-O-Gallate-Induced Apoptotic Cell Death via Activation of the cGMP Signaling Pathway in Caco-2 Cells" Current Issues in Molecular Biology 44, no. 12: 6247-6256. https://doi.org/10.3390/cimb44120426
APA StyleBae, J., Lee, K., Park, J. -S., Jung, J., Tachibana, H., Fujimura, Y., Kumazoe, M., Lim, J. S., Cho, Y. -C., Lee, S. -J., & Park, S. -J. (2022). Phosphodiesterase 5 Inhibitor Potentiates Epigallocatechin 3-O-Gallate-Induced Apoptotic Cell Death via Activation of the cGMP Signaling Pathway in Caco-2 Cells. Current Issues in Molecular Biology, 44(12), 6247-6256. https://doi.org/10.3390/cimb44120426