
Effects of Luteolin on Human Breast Cancer Using Gene Expression Array: Inferring Novel Genes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Reagent
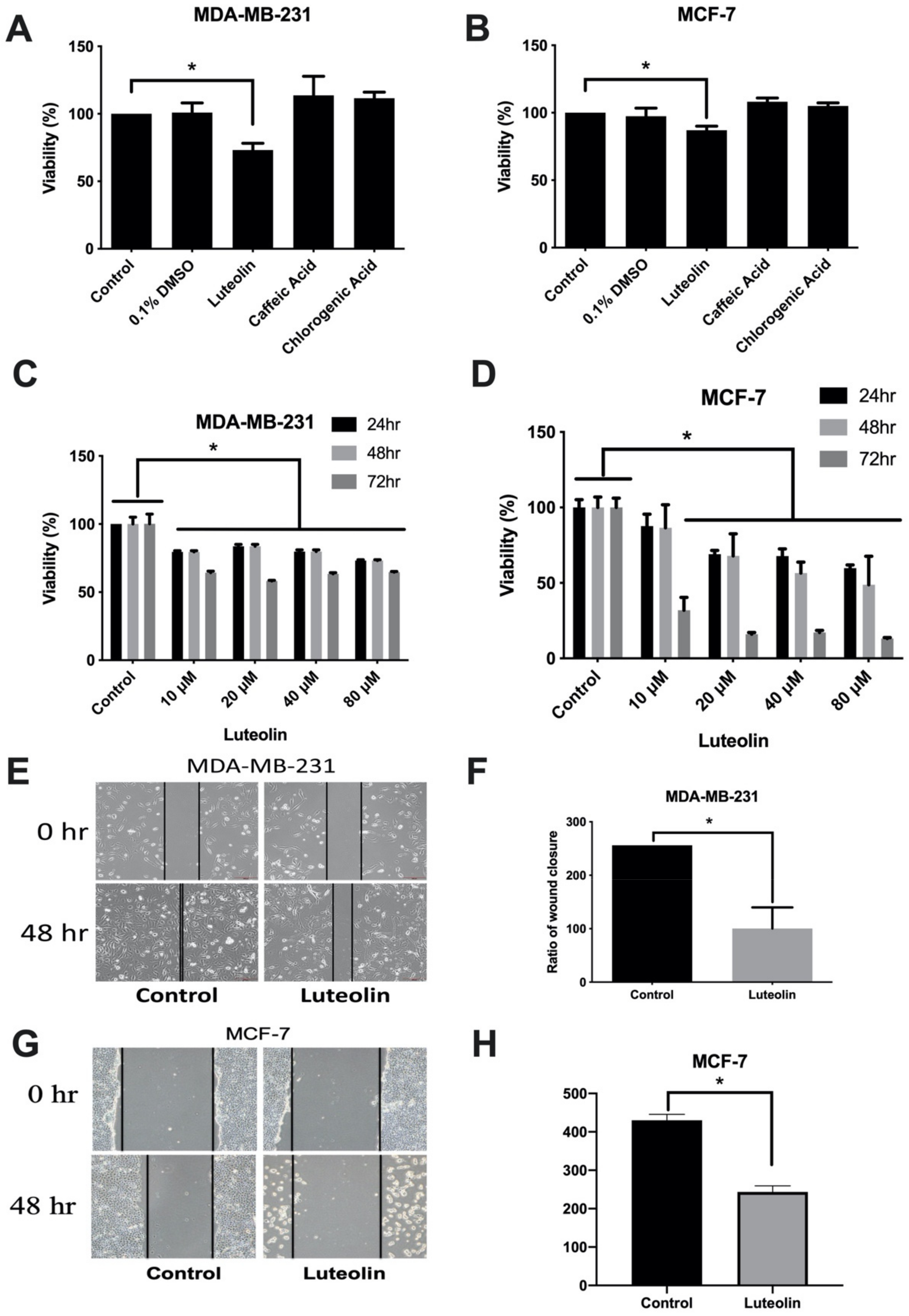
2.2. Cell Viability Assay
2.3. Wound Healing Assay
2.4. RNA Extraction and Reverse Transcription-Quantitative
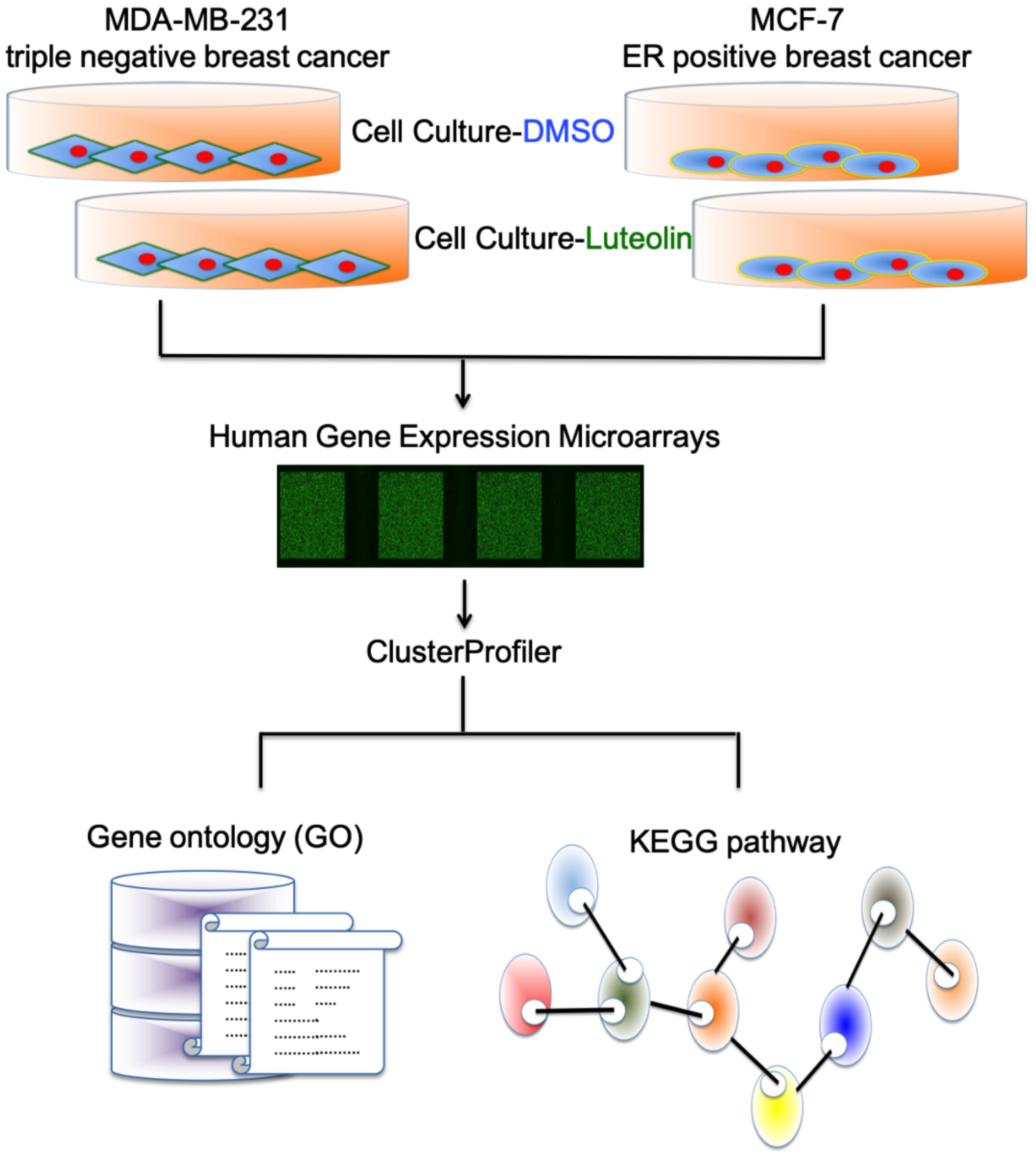
2.5. Array Hybridization and Data Analysis
2.6. Statistical Analysis
3. Results
3.1. Luteolin Inhibits the Viability and Wound Healing Ability of Human Breast Cancer Cells
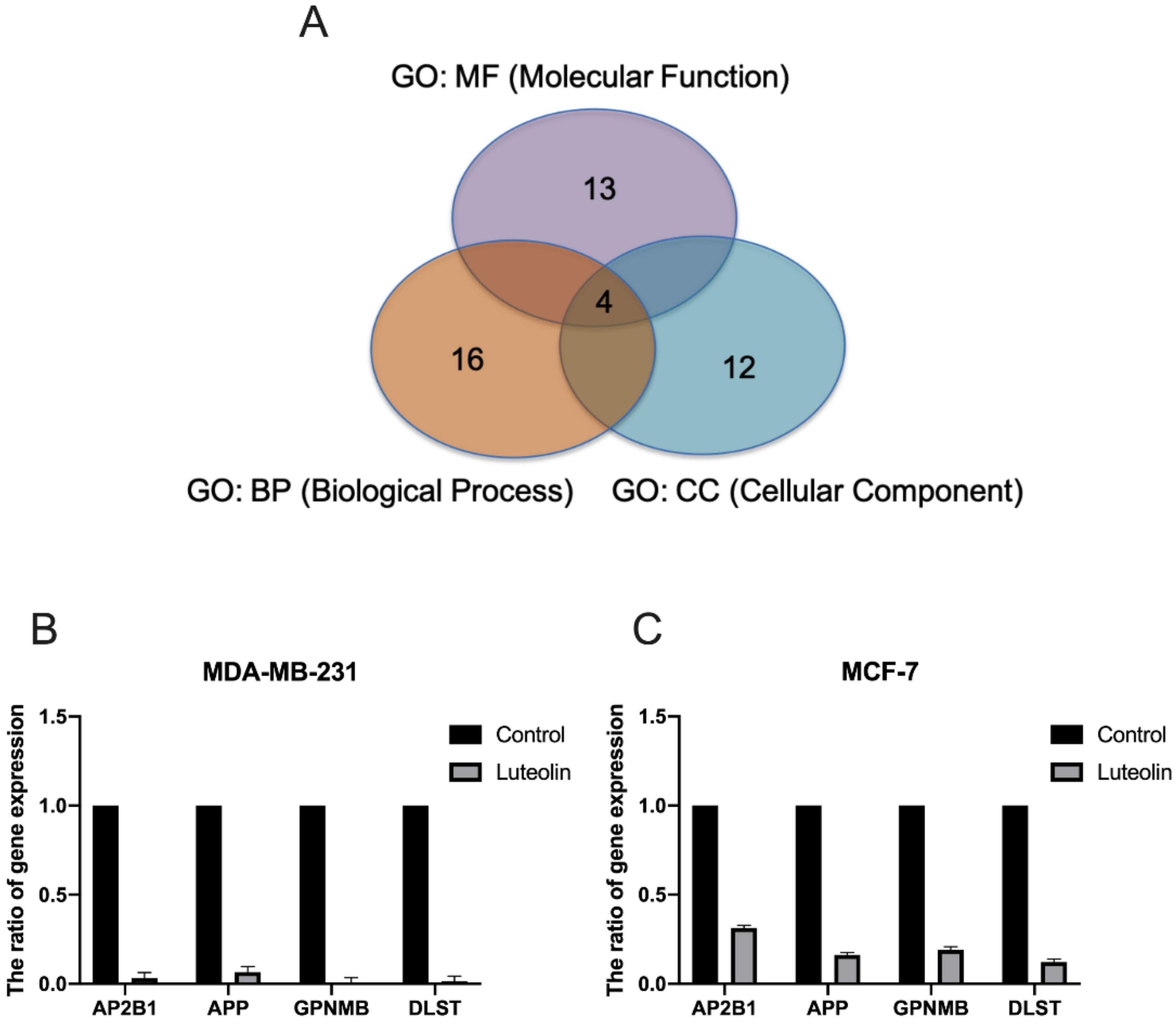
3.2. Identification of Potential Gene Expression Alterations in Luteolin-Treated Breast Cancer Cells
3.3. Validation of Differentially Expressed Genes (DEGs) in Response to Luteolin
3.4. External Validation of AP2B1, APP, GPNMB and DLST in Cell Viability, Human Protein Atlas (HPA) Databases and KEGG Pathways
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Azamjah, N.; Soltan-Zadeh, Y.; Zayeri, F. Global Trend of Breast Cancer Mortality Rate: A 25-Year Study. Asian Pac. J. Cancer Prev. 2019, 20, 2015–2020. [Google Scholar] [CrossRef] [PubMed]
- Duffy, S.W.; Dibden, A.; Michalopoulos, D.; Offman, J.; Parmar, D.; Jenkins, J.; Collins, B.; Robson, T.; Scorfield, S.; Green, K.; et al. Screen detection of ductal carcinoma in situ and subsequent incidence of invasive interval breast cancers: A retrospective population-based study. Lancet Oncol. 2016, 17, 109–114. [Google Scholar] [CrossRef] [Green Version]
- Narkhede, A.A.; Shevde, L.A.; Rao, S.S. Biomimetic strategies to recapitulate organ specific microenvironments for studying breast cancer metastasis. Int. J. Cancer 2017, 141, 1091–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, R.M.; Kuhl, C.K.; Moy, L. Contrast-enhanced MRI for breast cancer screening. J. Magn. Reson. Imaging 2019, 50, 377–390. [Google Scholar] [CrossRef]
- Weng, C.H.; Chen, Y.H.; Lin, C.H.; Luo, X.; Lin, T.H. Thyroid disorders and breast cancer risk in Asian population: A nationwide population-based case-control study in Taiwan. BMJ Open 2018, 8, e020194. [Google Scholar] [CrossRef] [Green Version]
- DeSantis, C.E.; Fedewa, S.A.; Goding Sauer, A.; Kramer, J.L.; Smith, R.A.; Jemal, A. Breast cancer statistics, 2015: Convergence of incidence rates between black and white women. CA Cancer J. Clin. 2016, 66, 31–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drukteinis, J.S.; Mooney, B.P.; Flowers, C.I.; Gatenby, R.A. Beyond mammography: New frontiers in breast cancer screening. Am. J. Med. 2013, 126, 472–479. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.A.; Goldstone, F.; Greenham, J. Flavonoids, cinnamic acids and coumarins from the different tissues and medicinal preparations of Taraxacum officinale. Phytochemistry 1996, 42, 121–127. [Google Scholar] [CrossRef]
- Rudenskaya, G.N.; Bogacheva, A.M.; Preusser, A.; Kuznetsova, A.V.; Dunaevsky Ya, E.; Golovkin, B.N.; Stepanov, V.M. Taraxalisin—A serine proteinase from dandelion Taraxacum officinale Webb s.l. FEBS Lett. 1998, 437, 237–240. [Google Scholar] [CrossRef] [Green Version]
- Omur, A.D.; Kandemir, F.M.; Apaydin Yildirim, B.; Akman, O.; Aktas Senocak, E.; Eldutar, E.; Aksu, E.H. Protective Effect of Dandelion (Taraxacum officinale) Extract Against Gentamicin-Induced Reproductive Damage in Male Rats. Kafkas Univ. Vet. Fak. Derg. 2016, 22, 929–936. [Google Scholar] [CrossRef]
- Jeon, D.; Kim, S.J.; Kim, H.S. Anti-inflammatory evaluation of the methanolic extract of Taraxacum officinale in LPS-stimulated human umbilical vein endothelial cells. BMC Complement. Altern. Med. 2017, 17, 508. [Google Scholar] [CrossRef] [PubMed]
- You, Y.; Yoo, S.; Yoon, H.G.; Park, J.; Lee, Y.H.; Kim, S.; Oh, K.T.; Lee, J.; Cho, H.Y.; Jun, W. In vitro and in vivo hepatoprotective effects of the aqueous extract from Taraxacum officinale (dandelion) root against alcohol-induced oxidative stress. Food Chem. Toxicol. 2010, 48, 1632–1637. [Google Scholar] [CrossRef] [PubMed]
- Verhoeven, K.J.F.; Biere, A. Geographic parthenogenesis and plant-enemy interactions in the common dandelion. BMC Evol. Biol. 2013, 13, 23. [Google Scholar] [CrossRef] [Green Version]
- Tahtamouni, L.H.; Alqurna, N.M.; Al-Hudhud, M.Y.; Al-Hajj, H.A. Dandelion (Taraxacum officinale) decreases male rat fertility in vivo. J. Ethnopharmacol. 2011, 135, 102–109. [Google Scholar] [CrossRef]
- Pereira, C.; Barros, L.; Ferreira, I.C.F.R. Extraction, identification, fractionation and isolation of phenolic compounds in plants with hepatoprotective effects. J. Sci. Food Agric. 2016, 96, 1068–1084. [Google Scholar] [CrossRef] [PubMed]
- Ata, S.; Farooq, F.; Javed, S. Elemental profile of 24 common medicinal plants of Pakistan and its direct link with traditional uses. J. Med. Plants Res. 2011, 5, 6164–6168. [Google Scholar] [CrossRef]
- Lin, Y.; Shi, R.; Wang, X.; Shen, H.M. Luteolin, a flavonoid with potential for cancer prevention and therapy. Curr. Cancer Drug Targets 2008, 8, 634–646. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.Y.; Wang, C.J.; Chen, N.F.; Ho, W.H.; Lu, F.J.; Tseng, T.H. Luteolin enhances paclitaxel-induced apoptosis in human breast cancer MDA-MB-231 cells by blocking STAT3. Chem. Biol. Interact. 2014, 213, 60–68. [Google Scholar] [CrossRef]
- Rosendahl, A.H.; Perks, C.M.; Zeng, L.; Markkula, A.; Simonsson, M.; Rose, C.; Ingvar, C.; Holly, J.M.; Jernstrom, H. Caffeine and Caffeic Acid Inhibit Growth and Modify Estrogen Receptor and Insulin-like Growth Factor I Receptor Levels in Human Breast Cancer. Clin. Cancer Res. 2015, 21, 1877–1887. [Google Scholar] [CrossRef] [Green Version]
- Yamagata, K.; Izawa, Y.; Onodera, D.; Tagami, M. Chlorogenic acid regulates apoptosis and stem cell marker-related gene expression in A549 human lung cancer cells. Mol. Cell. Biochem. 2018, 441, 9–19. [Google Scholar] [CrossRef]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the last 25 years. J. Nat. Prod. 2007, 70, 461–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Kim, D.S.; Kang, S.; Kim, H.J. The combination of luteolin and l-theanine improved Alzheimer disease-like symptoms by potentiating hippocampal insulin signaling and decreasing neuroinflammation and norepinephrine degradation in amyloid-beta-infused rats. Nutr. Res. 2018, 60, 116–131. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wang, R.; Tian, J.; Song, M.; Zhao, R.; Liu, K.; Zhu, F.; Shim, J.H.; Dong, Z.; Lee, M.H. Targeting LIMK1 with luteolin inhibits the growth of lung cancer in vitro and in vivo. J. Cell. Mol. Med. 2021, 25, 5560–5571. [Google Scholar] [CrossRef]
- Saafan, H.; Foerster, S.; Parra-Guillen, Z.P.; Hammer, E.; Michaelis, M.; Cinatl, J., Jr.; Volker, U.; Frohlich, H.; Kloft, C.; Ritter, C.A. Utilising the EGFR interactome to identify mechanisms of drug resistance in non-small cell lung cancer-Proof of concept towards a systems pharmacology approach. Eur. J. Pharm. Sci. 2016, 94, 20–32. [Google Scholar] [CrossRef]
- Wu, H.T.; Liu, Y.E.; Hsu, K.W.; Wang, Y.F.; Chan, Y.C.; Chen, Y.; Chen, D.R. MLL3 Induced by Luteolin Causes Apoptosis in Tamoxifen-Resistant Breast Cancer Cells through H3K4 Monomethylation and Suppression of the PI3K/AKT/mTOR Pathway. Am. J. Chin. Med. 2020, 48, 1221–1241. [Google Scholar] [CrossRef]
- Takagi, K.; Ito, S.; Miyazaki, T.; Miki, Y.; Shibahara, Y.; Ishida, T.; Watanabe, M.; Inoue, S.; Sasano, H.; Suzuki, T. Amyloid precursor protein in human breast cancer: An androgen-induced gene associated with cell proliferation. Cancer Sci. 2013, 104, 1532–1538. [Google Scholar] [CrossRef]
- Lim, S.; Yoo, B.K.; Kim, H.S.; Gilmore, H.L.; Lee, Y.; Lee, H.P.; Kim, S.J.; Letterio, J.; Lee, H.G. Amyloid-beta precursor protein promotes cell proliferation and motility of advanced breast cancer. BMC Cancer 2014, 14, 928. [Google Scholar] [CrossRef] [Green Version]
- Meng, J.Y.; Kataoka, H.; Itoh, H.; Koono, M. Amyloid beta protein precursor is involved in the growth of human colon carcinoma cell in vitro and in vivo. Int. J. Cancer 2001, 92, 31–39. [Google Scholar] [CrossRef]
- Venkataramani, V.; Rossner, C.; Iffland, L.; Schweyer, S.; Tamboli, I.Y.; Walter, J.; Wirths, O.; Bayer, T.A. Histone deacetylase inhibitor valproic acid inhibits cancer cell proliferation via down-regulation of the alzheimer amyloid precursor protein. J. Biol. Chem. 2010, 285, 10678–10689. [Google Scholar] [CrossRef] [Green Version]
- Sobol, A.; Galluzzo, P.; Weber, M.J.; Alani, S.; Bocchetta, M. Depletion of Amyloid Precursor Protein (APP) causes G0 arrest in non-small cell lung cancer (NSCLC) cells. J. Cell. Physiol. 2015, 230, 1332–1341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, H.L.; Tuli, A.; Wang, X.; Liu, C.; Pan, Z.; Ouellette, M.M.; Hollingsworth, M.A.; Macdonald, R.G.; Solheim, J.C. Relevance of amyloid precursor-like protein 2 C-terminal fragments in pancreatic cancer cells. Int. J. Oncol. 2012, 41, 1464–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansel, D.E.; Rahman, A.; Wehner, S.; Herzog, V.; Yeo, C.J.; Maitra, A. Increased expression and processing of the Alzheimer amyloid precursor protein in pancreatic cancer may influence cellular proliferation. Cancer Res. 2003, 63, 7032–7037. [Google Scholar] [PubMed]
- Liguori, M.; Digifico, E.; Vacchini, A.; Avigni, R.; Colombo, F.S.; Borroni, E.M.; Farina, F.M.; Milanesi, S.; Castagna, A.; Mannarino, L.; et al. The soluble glycoprotein NMB (GPNMB) produced by macrophages induces cancer stemness and metastasis via CD44 and IL-33. Cell. Mol. Immunol. 2021, 18, 711–722. [Google Scholar] [CrossRef]
- Rose, A.A.; Grosset, A.A.; Dong, Z.; Russo, C.; Macdonald, P.A.; Bertos, N.R.; St-Pierre, Y.; Simantov, R.; Hallett, M.; Park, M.; et al. Glycoprotein nonmetastatic B is an independent prognostic indicator of recurrence and a novel therapeutic target in breast cancer. Clin. Cancer Res. 2010, 16, 2147–2156. [Google Scholar] [CrossRef] [Green Version]
- Ruan, W.M.; Li, Y.L.; Nie, G.; Zhou, W.X.; Zou, X.M. Differential expression of glycoprotein non-metastatic melanoma protein B (GPNMB) involved in trichostatin A-induced apoptosis in gastric cancer. Int. J. Clin. Exp. Med. 2014, 7, 4857–4866. [Google Scholar]
- Anderson, N.M.; Li, D.; Peng, H.L.; Laroche, F.J.; Mansour, M.R.; Gjini, E.; Aioub, M.; Helman, D.J.; Roderick, J.E.; Cheng, T.; et al. The TCA cycle transferase DLST is important for MYC-mediated leukemogenesis. Leukemia 2016, 30, 1365–1374. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Xu, Y.; Cao, M.; Huan, Y.; Zhu, L.; Jiang, Y.; Shen, W.; Zhu, G. Luteolin Induces Apoptosis and Autophagy in Mouse Macrophage ANA-1 Cells via the Bcl-2 Pathway. J. Immunol. Res. 2018, 2018, 4623919. [Google Scholar] [CrossRef]
- Wang, S.W.; Chen, Y.R.; Chow, J.M.; Chien, M.H.; Yang, S.F.; Wen, Y.C.; Lee, W.J.; Tseng, T.H. Stimulation of Fas/FasL-mediated apoptosis by luteolin through enhancement of histone H3 acetylation and c-Jun activation in HL-60 leukemia cells. Mol. Carcinog. 2018, 57, 866–877. [Google Scholar] [CrossRef]
- Dong, J.; Xu, O.; Wang, J.; Shan, C.; Ren, X. Luteolin ameliorates inflammation and Th1/Th2 imbalance via regulating the TLR4/NF-kappaB pathway in allergic rhinitis rats. Immunopharmacol. Immunotoxicol. 2021, 43, 319–327. [Google Scholar] [CrossRef]
- Xia, N.; Chen, G.; Liu, M.; Ye, X.; Pan, Y.; Ge, J.; Mao, Y.; Wang, H.; Wang, J.; Xie, S. Anti-inflammatory effects of luteolin on experimental autoimmune thyroiditis in mice. Exp. Ther. Med. 2016, 12, 4049–4054. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Official Gene Symbol | Gene Name | MCF-7 Fold Change (Luteolin/Control) | MDA-MB-231 Fold Change (Luteolin/Control) | Gene Expression |
|---|---|---|---|---|
| ENST00000438070 | Homo sapiens cDNA 5′, mRNA sequence [CB265963] | 4.127925665 | 7.321832 | up |
| TMEM59L | transmembrane protein 59-like | 2.667702366 | 4.777713 | up |
| Lnc-CHID1-1 | cDNA FLJ44677 fis, clone | 2.310667214 | 3.432382 | up |
| VPREB3 | pre-B lymphocyte 3 | 2.229125449 | 3.137973 | up |
| Lnc-PPP1R32-1 | linc|LNCipedia lincRNA | 2.14699303 | 3.129121 | up |
| FCGBP | Fc fragment of IgG binding protein | 2.125306475 | 2.639865 | up |
| A_33_P3223860 | Unknown | 2.088463106 | 2.586989 | up |
| ENST00000426023 | Homo sapiens cDNA [FLJ32335] | −2.037285761 | −2.6294 | down |
| A_19_P00803499 | Unknown | −2.041876695 | −2.14134 | down |
| GPNMB | glycoprotein (transmembrane) nmb | −2.065156727 | −2.75712 | down |
| RAB5C | RAB5C, member RAS oncogene family | −2.088751541 | −2.03856 | down |
| COL12A1 | collagen, type XII, alpha 1 | −2.11607192 | −2.6307 | down |
| LINC01116 | long intergenic non-protein coding RNA 1116 | −2.121113679 | −2.09347 | down |
| PID1 | phosphotyrosine interaction domain containing 1 | −2.132012086 | −2.88883 | down |
| PPP3R1 | protein phosphatase 3, regulatory subunit B, alpha | −2.145744297 | −2.30164 | down |
| OXTR | oxytocin receptor | −2.195093084 | −3.77586 | down |
| INCENP | inner centromere protein antigen 135/155 kDa | −2.263751545 | −2.45544 | down |
| SERPINA1 | serpin peptidase inhibitor, clade A (alpha-1 antiproteinase, antitrypsin), member 1 | −2.322611225 | −2.05576 | down |
| HLA-DPB1 | major histocompatibility complex, class II, DP beta 1 | −2.352085002 | −2.60705 | down |
| FAM155A | family with sequence similarity 155, member A | −2.37484673 | −2.73719 | down |
| HMOX1 | heme oxygenase (decycling) 1 | −2.383757494 | −2.25384 | down |
| HLA-DRA | major histocompatibility complex, class II, DR alpha | −2.388556651 | −4.32678 | down |
| APOL6 | apolipoprotein L, 6 | −2.486493415 | −6.89045 | down |
| LMNB1 | lamin B1 | −2.542284469 | −5.82738 | down |
| APH1A | APH1A gamma secretase subunit | −2.561979585 | −2.86107 | down |
| MIR503HG | MIR503 host gene (non-protein-coding) | −2.673050357 | −3.10265 | down |
| SNORA79 | small nucleolar RNA, H/ACA box 79 | −2.6991916 | −2.00797 | down |
| ARHGEF9 | Cdc42 guanine nucleotide exchange factor (GEF) 9 | −2.706861919 | −2.17995 | down |
| UBE2E1 | Ubiquitin-conjugating enzyme E2E 1 | −2.715830339 | −6.74322 | down |
| SNORA12 | Homo sapiens cDNA 5′ end, mRNA sequence [AA378382] | −2.960722177 | −2.14457 | down |
| ERF | Ets2 repressor factor | −3.038398994 | −2.22017 | down |
| APP | amyloid beta (A4) precursor protein | −3.405819858 | −4.2484 | down |
| SLC39A7 | solute carrier family 39 (zinc transporter), member 7 | −3.491704458 | −3.31685 | down |
| DLST | dihydrolipoamide S-succinyltransferase (E2 component of 2-oxo-glutarate complex) | −3.719897742 | −2.18842 | down |
| CCDC71 | Coiled-coil domain containing 71 | −3.727339467 | −5.58472 | down |
| AP2B1 | adaptor-related protein complex 2, beta 1 subunit | −3.866732244 | −2.69467 | down |
| ATF6B | activating transcription factor 6 beta | −4.607165158 | −3.32893 | down |
| RNF121 | ring finger protein 121 | −4.680226478 | −2.12748 | down |
| A_33_P3241184 | Unknown | −5.616014032 | −5.53105 | down |
| AKT2 | v-akt murine thymoma viral oncogene homolog 2 | −5.756814436 | −3.69298 | down |
| TAF15 | TAF15 RNA polymerase II, TATA box binding protein (TBP)-associated factor | −7.381370191 | −4.74989 | down |
| Network | Predominant Diseases and Functions | Number of Genes | Molecules in Network |
|---|---|---|---|
| GO: MF (Molecular Function) | peptide binding, serine-type endopeptidase inhibitor activity, clathrin adaptor activity, cAMP response element binding, mechanosensitive ion channel activity, phospholipase binding, heparin binding, zinc ion transmembrane transporter activity, S-acyltransferase activity | 13 | AP2B1/HLA-DRA/PPP3R1/HLA-DPB1/OXTR/SERPINA1/APP/ATF6B/FAM155A/LMNB1/GPNMB/SLC39A7/DLST |
| GO: BP (Biological Process) | positive regulation of chemokine biosynthetic process, neuron death, suckling behavior, regulation of protein localization to membrane, cellular transition metal ion homeostasis, amyloid-beta formation, endoplasmic reticulum unfolded protein response, lysine metabolic process, growth plate cartilage chondrocyte morphogenesis, regulation of mitotic cell cycle phase transition | 16 | HMOX1/AP2B1/AKT2/APP/GPNMB/OXTR/PID1/PPP3R1/SLC39A7/APH1A/ATF6B/RNF121/DLST/COL12A1/UBE2E1/GPNMB |
| GO: CC (Cellular Component) | COPII-coated ER to Golgi transport vesicle, clathrin-coated endocytic vesicle membrane, endosome membrane, nuclear lamina, dihydrolipoyl dehydrogenase complex, lateral element | 12 | SERPINA1/HLA-DRA/HLA-DPB1/AP2B1/APH1A/RAB5C/AKT2/APP/GPNMB/LMNB1/DLST/INCENP |
| Description | Count | p-Value | Genes |
|---|---|---|---|
| Tuberculosis | 5 | 0.00003 | RAB5C/HLA-DRA/PPP3R1/HLA-DPB1/AKT2 |
| Human T-cell leukemia virus 1 infection | 5 | 0.00007 | HLA-DRA/PPP3R1/HLA-DPB1/AKT2/ATF6B |
| Th1 and Th2 cell differentiation | 3 | 0.0009 | HLA-DRA/PPP3R1/HLA-DPB1 |
| Th17 cell differentiation | 3 | 0.0014 | HLA-DRA/PPP3R1/HLA-DPB1 |
| Toxoplasmosis | 3 | 0.0016 | HLA-DRA/HLA-DPB1/AKT2 |
| Asthma | 2 | 0.0019 | HLA-DRA/HLA-DPB1 |
| Allograft rejection | 2 | 0.0028 | HLA-DRA/HLA-DPB1 |
| Graft-versus-host disease | 2 | 0.0033 | HLA-DRA/HLA-DPB1 |
| Type I diabetes mellitus | 2 | 0.0036 | HLA-DRA/HLA-DPB1 |
| Phagosome | 3 | 0.0037 | RAB5C/HLA-DRA/HLA-DPB1 |
| Alzheimer’s disease | 4 | 0.0045 | APH1A/PPP3R1/AKT2/APP |
| Intestinal immune network for IgA production | 2 | 0.0047 | HLA-DRA/HLA-DPB1 |
| cGMP-PKG signaling pathway | 3 | 0.0048 | PPP3R1/AKT2/ATF6B |
| Influenza A | 3 | 0.0051 | HLA-DRA/HLA-DPB1/AKT2 |
| Autoimmune thyroid disease | 2 | 0.0054 | HLA-DRA/HLA-DPB1 |
| VEGF signaling pathway | 2 | 0.0067 | PPP3R1/AKT2 |
| Viral myocarditis | 2 | 0.0069 | HLA-DRA/HLA-DPB1 |
| Epstein–Barr virus infection | 3 | 0.0081 | HLA-DRA/HLA-DPB1/AKT2 |
| Inflammatory bowel disease (IBD) | 2 | 0.0081 | HLA-DRA/HLA-DPB1 |
| Amphetamine addiction | 2 | 0.0091 | PPP3R1/ATF6B |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.-H.; Wu, C.-H.; Tsai, C.-C.; Chen, T.-Y.; Tsai, K.-J.; Hung, C.-M.; Hsu, C.-Y.; Wu, C.-W.; Hsieh, T.-H. Effects of Luteolin on Human Breast Cancer Using Gene Expression Array: Inferring Novel Genes. Curr. Issues Mol. Biol. 2022, 44, 2107-2121. https://doi.org/10.3390/cimb44050142
Wang S-H, Wu C-H, Tsai C-C, Chen T-Y, Tsai K-J, Hung C-M, Hsu C-Y, Wu C-W, Hsieh T-H. Effects of Luteolin on Human Breast Cancer Using Gene Expression Array: Inferring Novel Genes. Current Issues in Molecular Biology. 2022; 44(5):2107-2121. https://doi.org/10.3390/cimb44050142
Chicago/Turabian StyleWang, Shih-Ho, Chin-Hu Wu, Chin-Chuan Tsai, Tai-Yu Chen, Kuen-Jang Tsai, Chao-Ming Hung, Chia-Yi Hsu, Chia-Wei Wu, and Tsung-Hua Hsieh. 2022. "Effects of Luteolin on Human Breast Cancer Using Gene Expression Array: Inferring Novel Genes" Current Issues in Molecular Biology 44, no. 5: 2107-2121. https://doi.org/10.3390/cimb44050142
APA StyleWang, S. -H., Wu, C. -H., Tsai, C. -C., Chen, T. -Y., Tsai, K. -J., Hung, C. -M., Hsu, C. -Y., Wu, C. -W., & Hsieh, T. -H. (2022). Effects of Luteolin on Human Breast Cancer Using Gene Expression Array: Inferring Novel Genes. Current Issues in Molecular Biology, 44(5), 2107-2121. https://doi.org/10.3390/cimb44050142