
Using Next-Generation Sequencing and Bioinformatic Methods to Predict New Genes That May Be Regulated by CD47 in Oral Squamous Cell Carcinoma

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Plasmid Transfection
2.3. RNA Extraction and qPCR
2.4. RNA-Seq Quantification
2.5. Ingenuity Pathway Analysis
2.6. Database for Annotation, Visualization and Integrated Discovery Analysis
2.7. Cell Viability Assay
2.8. Statistical Analysis
3. Results
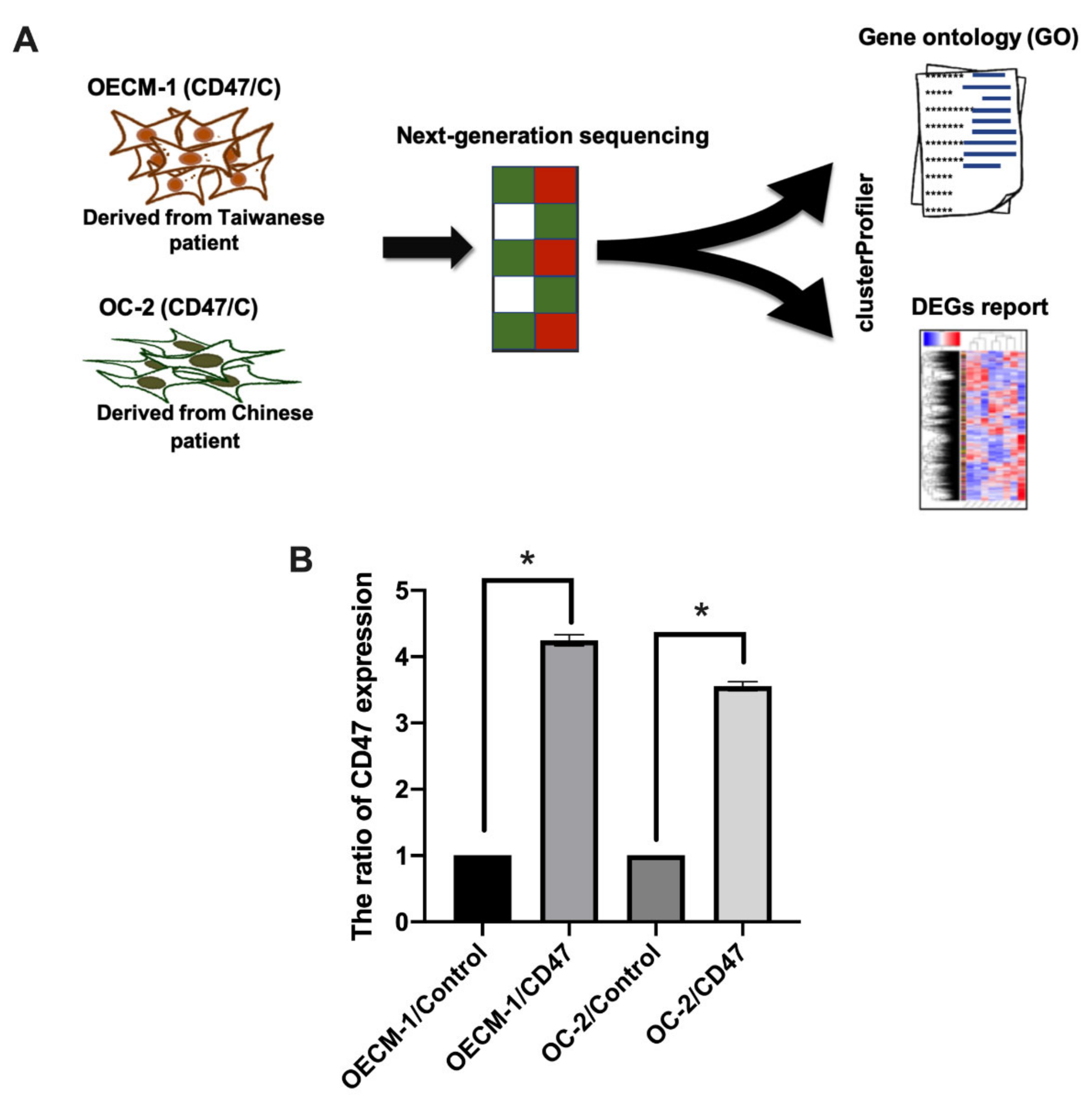
3.1. Gene Expression Profiling and Ovexpression of CD47 in OSCC Cell Lies
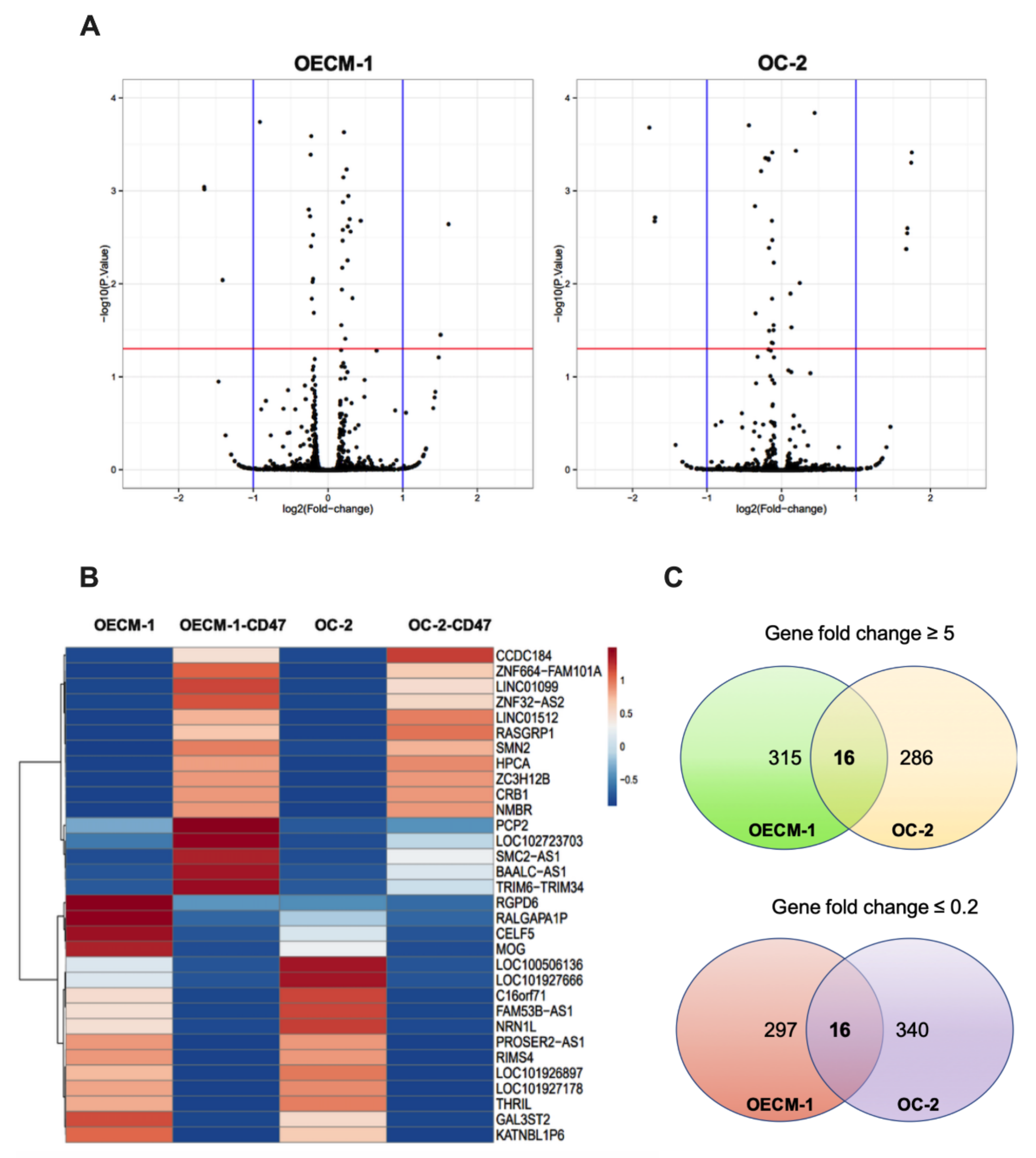
3.2. Gene Expression Profile in CD47-Overexpressing Oral Cancer
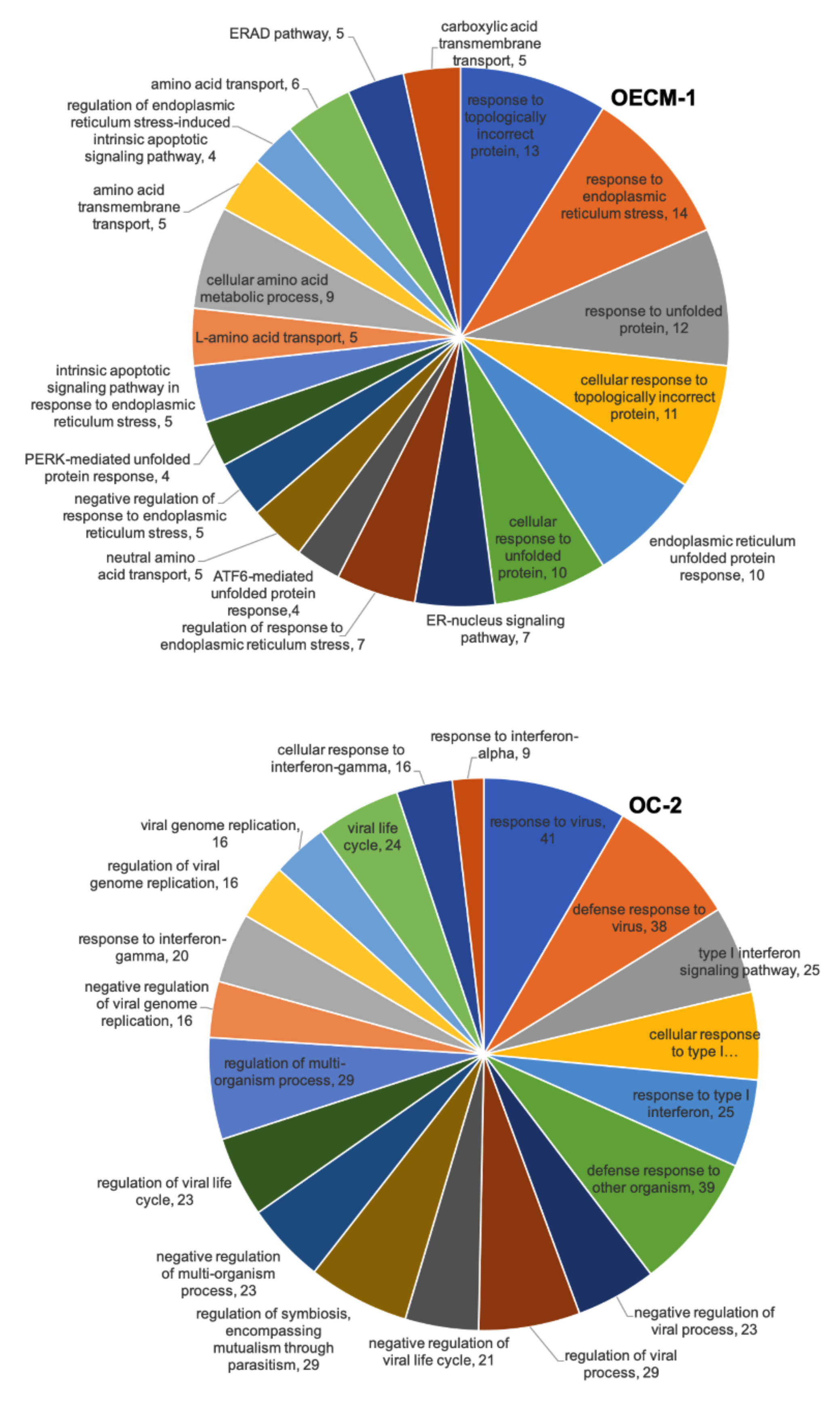
3.3. Biological Process Analysis of Differentially Expressed Genes in CD47-Overexpressing Oral Squamous Cell Carcinoma
3.4. The DAVID Analyses of Differentially Expressed Genes following CD47-Overexpression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hung, L.C.; Kung, P.T.; Lung, C.H.; Tsai, M.H.; Liu, S.A.; Chiu, L.T.; Huang, K.H.; Tsai, W.C. Assessment of the Risk of Oral Cancer Incidence in A High-Risk Population and Establishment of A Predictive Model for Oral Cancer Incidence Using A Population-Based Cohort in Taiwan. Int. J. Environ. Res. Public Health 2020, 17, 665. [Google Scholar] [CrossRef] [Green Version]
- Dhanuthai, K.; Rojanawatsirivej, S.; Thosaporn, W.; Kintarak, S.; Subarnbhesaj, A.; Darling, M.; Kryshtalskyj, E.; Chiang, C.P.; Shin, H.I.; Choi, S.Y.; et al. Oral cancer: A multicenter study. Med. Oral. Patol. Oral Cir. Bucal. 2018, 23, e23–e29. [Google Scholar] [CrossRef]
- Manikandan, M.; Deva Magendhra Rao, A.K.; Arunkumar, G.; Manickavasagam, M.; Rajkumar, K.S.; Rajaraman, R.; Munirajan, A.K. Oral squamous cell carcinoma: MicroRNA expression profiling and integrative analyses for elucidation of tumourigenesis mechanism. Mol. Cancer 2016, 15, 28. [Google Scholar] [CrossRef] [Green Version]
- Pignon, J.P.; le Maitre, A.; Maillard, E.; Bourhis, J.; Group, M.-N.C. Meta-analysis of chemotherapy in head and neck cancer (MACH-NC): An update on 93 randomised trials and 17,346 patients. Radiother. Oncol. 2009, 92, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Dzebo, S.; Mahmutovic, J.; Erkocevic, H.; Foco, F. Frequency of Depression and its Correlation with Quality of Life of Patients with Oral Cavity Cancer. Mater. Sociomed. 2017, 29, 97–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manasa, V.G.; Kannan, S. Impact of microRNA dynamics on cancer hallmarks: An oral cancer scenario. Tumour. Biol. 2017, 39, 1010428317695920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yete, S.; D’Souza, W.; Saranath, D. High-Risk Human Papillomavirus in Oral Cancer: Clinical Implications. Oncology 2018, 94, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Santos, H.B.; dos Santos, T.K.; Paz, A.R.; Cavalcanti, Y.W.; Nonaka, C.F.; Godoy, G.P.; Alves, P.M. Clinical findings and risk factors to oral squamous cell carcinoma in young patients: A 12-year retrospective analysis. Med. Oral. Patol. Oral Cir. Bucal. 2016, 21, e151–e156. [Google Scholar] [CrossRef] [PubMed]
- Polesel, J.; Franceschi, S.; Talamini, R.; Negri, E.; Barzan, L.; Montella, M.; Libra, M.; Vaccher, E.; Franchin, G.; La Vecchia, C.; et al. Tobacco smoking, alcohol drinking, and the risk of different histological types of nasopharyngeal cancer in a low-risk population. Oral Oncol. 2011, 47, 541–545. [Google Scholar] [CrossRef]
- Brown, E.J.; Frazier, W.A. Integrin-associated protein (CD47) and its ligands. Trends Cell. Biol. 2001, 11, 130–135. [Google Scholar] [CrossRef]
- Van Beek, E.M.; Cochrane, F.; Barclay, A.N.; van den Berg, T.K. Signal regulatory proteins in the immune system. J. Immunol. 2005, 175, 7781–7787. [Google Scholar] [CrossRef] [Green Version]
- Kiss, B.; van den Berg, N.S.; Ertsey, R.; McKenna, K.; Mach, K.E.; Zhang, C.A.; Volkmer, J.P.; Weissman, I.L.; Rosenthal, E.L.; Liao, J.C. CD47-Targeted Near-Infrared Photoimmunotherapy for Human Bladder Cancer. Clin. Cancer Res. 2019, 25, 3561–3571. [Google Scholar] [CrossRef] [Green Version]
- Nakaishi, A.; Hirose, M.; Yoshimura, M.; Oneyama, C.; Saito, K.; Kuki, N.; Matsuda, M.; Honma, N.; Ohnishi, H.; Matozaki, T.; et al. Structural insight into the specific interaction between murine SHPS-1/SIRP alpha and its ligand CD47. J. Mol. Biol. 2008, 375, 650–660. [Google Scholar] [CrossRef]
- Oldenborg, P.A.; Sundqvist, I.M.; Sehlin, J. Different effects of glucose on extracellular and intracellular respiratory burst response in normal human neutrophils activated with the soluble agonist fMet-Leu-Phe. Diabet. Med. 2000, 17, 532–537. [Google Scholar] [CrossRef]
- Bornstein, P. Diversity of function is inherent in matricellular proteins: An appraisal of thrombospondin 1. J. Cell. Biol. 1995, 130, 503–506. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, S.; Jamieson, C.H.; Pang, W.W.; Park, C.Y.; Chao, M.P.; Majeti, R.; Traver, D.; van Rooijen, N.; Weissman, I.L. CD47 is upregulated on circulating hematopoietic stem cells and leukemia cells to avoid phagocytosis. Cell 2009, 138, 271–285. [Google Scholar] [CrossRef] [Green Version]
- Imam, R.; Chang, Q.; Black, M.; Yu, C.; Cao, W. CD47 expression and CD163(+) macrophages correlated with prognosis of pancreatic neuroendocrine tumor. BMC Cancer 2021, 21, 320. [Google Scholar] [CrossRef]
- Yuan, J.; Shi, X.; Chen, C.; He, H.; Liu, L.; Wu, J.; Yan, H. High expression of CD47 in triple negative breast cancer is associated with epithelial-mesenchymal transition and poor prognosis. Oncol. Lett. 2019, 18, 3249–3255. [Google Scholar] [CrossRef] [Green Version]
- Barrera, L.; Montes-Servin, E.; Hernandez-Martinez, J.M.; Garcia-Vicente, M.L.A.; Montes-Servin, E.; Herrera-Martinez, M.; Crispin, J.C.; Borbolla-Escoboza, J.R.; Arrieta, O. CD47 overexpression is associated with decreased neutrophil apoptosis/phagocytosis and poor prognosis in non-small-cell lung cancer patients. Br. J. Cancer 2017, 117, 385–397. [Google Scholar] [CrossRef] [Green Version]
- Willingham, S.B.; Volkmer, J.P.; Gentles, A.J.; Sahoo, D.; Dalerba, P.; Mitra, S.S.; Wang, J.; Contreras-Trujillo, H.; Martin, R.; Cohen, J.D.; et al. The CD47-signal regulatory protein alpha (SIRPa) interaction is a therapeutic target for human solid tumors. Proc. Natl. Acad. Sci. USA 2012, 109, 6662–6667. [Google Scholar] [CrossRef] [Green Version]
- Golubovskaya, V.; Berahovich, R.; Zhou, H.; Xu, S.; Harto, H.; Li, L.; Chao, C.C.; Mao, M.M.; Wu, L. CD47-CAR-T Cells Effectively Kill Target Cancer Cells and Block Pancreatic Tumor Growth. Cancers 2017, 9, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Candas-Green, D.; Xie, B.; Huang, J.; Fan, M.; Wang, A.; Menaa, C.; Zhang, Y.; Zhang, L.; Jing, D.; Azghadi, S.; et al. Dual blockade of CD47 and HER2 eliminates radioresistant breast cancer cells. Nat. Commun. 2020, 11, 4591. [Google Scholar] [CrossRef] [PubMed]
- Pai, S.; Bamodu, O.A.; Lin, Y.K.; Lin, C.S.; Chu, P.Y.; Chien, M.H.; Wang, L.S.; Hsiao, M.; Yeh, C.T.; Tsai, J.T. CD47-SIRPalpha Signaling Induces Epithelial-Mesenchymal Transition and Cancer Stemness and Links to a Poor Prognosis in Patients with Oral Squamous Cell Carcinoma. Cells 2019, 8, 1658. [Google Scholar] [CrossRef] [Green Version]
- Wong, D.Y.; Chang, K.W.; Chen, C.F.; Chang, R.C. Characterization of two new cell lines derived from oral cavity human squamous cell carcinomas--OC1 and OC2. J. Oral. Maxillofac. Surg. 1990, 48, 385–390. [Google Scholar] [CrossRef]
- Yang, C.Y.; Meng, C.L. Regulation of PG synthase by EGF and PDGF in human oral, breast, stomach, and fibrosarcoma cancer cell lines. J. Dent. Res. 1994, 73, 1407–1415. [Google Scholar] [CrossRef] [PubMed]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Singh, P.; Rai, A.; Verma, A.K.; Alsahli, M.A.; Rahmani, A.H.; Almatroodi, S.A.; Alrumaihi, F.; Dev, K.; Sinha, A.; Sankhwar, S.; et al. Survival-Based Biomarker Module Identification Associated with Oral Squamous Cell Carcinoma (OSCC). Biology 2021, 10, 760. [Google Scholar] [CrossRef]
- Sakakura, K.; Takahashi, H.; Kaira, K.; Toyoda, M.; Murata, T.; Ohnishi, H.; Oyama, T.; Chikamatsu, K. Relationship between tumor-associated macrophage subsets and CD47 expression in squamous cell carcinoma of the head and neck in the tumor microenvironment. Lab. Invest. 2016, 96, 994–1003. [Google Scholar] [CrossRef]
- Vaeteewoottacharn, K.; Kariya, R.; Pothipan, P.; Fujikawa, S.; Pairojkul, C.; Waraasawapati, S.; Kuwahara, K.; Wongkham, C.; Wongkham, S.; Okada, S. Attenuation of CD47-SIRPalpha Signal in Cholangiocarcinoma Potentiates Tumor-Associated Macrophage-Mediated Phagocytosis and Suppresses Intrahepatic Metastasis. Transl. Oncol. 2019, 12, 217–225. [Google Scholar] [CrossRef]
- Lee, A.S. Glucose-regulated proteins in cancer: Molecular mechanisms and therapeutic potential. Nat. Rev. Cancer 2014, 14, 263–276. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.W.; Tseng, C.F.; Wang, M.Y.; Chang, W.C.; Lee, C.C.; Chen, L.T.; Hung, M.C.; Su, J.L. Deacetylation of HSPA5 by HDAC6 leads to GP78-mediated HSPA5 ubiquitination at K447 and suppresses metastasis of breast cancer. Oncogene 2016, 35, 1517–1528. [Google Scholar] [CrossRef]
- Luo, X.; Yao, J.; Nie, P.; Yang, Z.; Feng, H.; Chen, P.; Shi, X.; Zou, Z. FOXM1 promotes invasion and migration of colorectal cancer cells partially dependent on HSPA5 transactivation. Oncotarget 2016, 7, 26480–26495. [Google Scholar] [CrossRef]
- Lee, A.S. GRP78 induction in cancer: Therapeutic and prognostic implications. Cancer Res. 2007, 67, 3496–3499. [Google Scholar] [CrossRef] [Green Version]
- Luo, S.; Mao, C.; Lee, B.; Lee, A.S. GRP78/BiP is required for cell proliferation and protecting the inner cell mass from apoptosis during early mouse embryonic development. Mol. Cell. Biol. 2006, 26, 5688–5697. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Jiang, Y.; Jia, Z.; Li, Q.; Gong, W.; Wang, L.; Wei, D.; Yao, J.; Fang, S.; Xie, K. Association of elevated GRP78 expression with increased lymph node metastasis and poor prognosis in patients with gastric cancer. Clin. Exp. Metastasis 2006, 23, 401–410. [Google Scholar] [CrossRef]
- Stojadinovic, A.; Hooke, J.A.; Shriver, C.D.; Nissan, A.; Kovatich, A.J.; Kao, T.C.; Ponniah, S.; Peoples, G.E.; Moroni, M. HYOU1/Orp150 expression in breast cancer. Med. Sci. Monit. 2007, 13, BR231–BR239. [Google Scholar]
- Slaby, O.; Sobkova, K.; Svoboda, M.; Garajova, I.; Fabian, P.; Hrstka, R.; Nenutil, R.; Sachlova, M.; Kocakova, I.; Michalek, J.; et al. Significant overexpression of Hsp110 gene during colorectal cancer progression. Oncol. Rep. 2009, 21, 1235–1241. [Google Scholar] [CrossRef] [Green Version]
- Kusaczuk, M.; Cechowska-Pasko, M. Molecular chaperone ORP150 in ER stress-related diseases. Curr. Pharm. Des. 2013, 19, 2807–2818. [Google Scholar] [CrossRef]
- Fagone, P.; Di Rosa, M.; Palumbo, M.; De Gregorio, C.; Nicoletti, F.; Malaguarnera, L. Modulation of heat shock proteins during macrophage differentiation. Inflamm. Res. 2012, 61, 1131–1139. [Google Scholar] [CrossRef]
- Zhang, M.; Hutter, G.; Kahn, S.A.; Azad, T.D.; Gholamin, S.; Xu, C.Y.; Liu, J.; Achrol, A.S.; Richard, C.; Sommerkamp, P.; et al. Anti-CD47 Treatment Stimulates Phagocytosis of Glioblastoma by M1 and M2 Polarized Macrophages and Promotes M1 Polarized Macrophages In Vivo. PLoS ONE 2016, 11, e0153550. [Google Scholar] [CrossRef]
- Zhang, Y.; Sime, W.; Juhas, M.; Sjolander, A. Crosstalk between colon cancer cells and macrophages via inflammatory mediators and CD47 promotes tumour cell migration. Eur. J. Cancer 2013, 49, 3320–3334. [Google Scholar] [CrossRef]
- Liu, R.; Wei, H.; Gao, P.; Yu, H.; Wang, K.; Fu, Z.; Ju, B.; Zhao, M.; Dong, S.; Li, Z.; et al. CD47 promotes ovarian cancer progression by inhibiting macrophage phagocytosis. Oncotarget 2017, 8, 39021–39032. [Google Scholar] [CrossRef] [Green Version]
- Rastgoo, N.; Wu, J.; Liu, A.; Pourabdollah, M.; Atenafu, E.G.; Reece, D.; Chen, W.; Chang, H. Targeting CD47/TNFAIP8 by miR-155 overcomes drug resistance and inhibits tumor growth through induction of phagocytosis and apoptosis in multiple myeloma. Haematologica 2020, 105, 2813–2823. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Li, J.; Tong, B.; Chen, M.; Liu, X.; Zhong, W.; Zhao, J.; Wang, M. Positive tumour CD47 expression is an independent prognostic factor for recurrence in resected non-small cell lung cancer. ESMO Open 2020, 5, e000823. [Google Scholar] [CrossRef] [PubMed]
- Soto-Pantoja, D.R.; Kaur, S.; Roberts, D.D. CD47 signaling pathways controlling cellular differentiation and responses to stress. Crit. Rev. Biochem. Mol. Biol. 2015, 50, 212–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, K.L.; Soto-Pantoja, D.R. “UPRegulation” of CD47 by the endoplasmic reticulum stress pathway controls anti-tumor immune responses. Biomark Res. 2017, 5, 26. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, H.; Cheng, Q. PDIA4: The basic characteristics, functions and its potential connection with cancer. Biomed. Pharmacother. 2020, 122, 109688. [Google Scholar] [CrossRef]
- Chanjiao, Y.; Chunyan, C.; Xiaoxin, Q.; Youjian, H. MicroRNA-378a-3p contributes to ovarian cancer progression through downregulating PDIA4. Immun. Inflamm. Dis. 2021, 9, 108–119. [Google Scholar] [CrossRef]
- Gagnon, E.; Duclos, S.; Rondeau, C.; Chevet, E.; Cameron, P.H.; Steele-Mortimer, O.; Paiement, J.; Bergeron, J.J.; Desjardins, M. Endoplasmic reticulum-mediated phagocytosis is a mechanism of entry into macrophages. Cell 2002, 110, 119–131. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Official Gene Symbol | Gene Name | OECM-1-CD47/OECM-1-Empty Control FOLD-Change | OC2-CD47/OC2-Empty Control Fold-Change | GENE Expression |
|---|---|---|---|---|
| BAALC-AS1 | BAALC antisense RNA 1 | 12.15 | 6.26 | Up |
| CCDC184 | Coiled–coil domain containing 184 | 5.46 | 8.88 | Up |
| CRB1 | Crumbs 1, cell polarity complex component | 5.46 | 6.26 | Up |
| HPCA | Hippocalcin | 7.69 | 8.88 | Up |
| LINC01099 | Long intergenic non-protein coding RNA 1099 | 7.73 | 6.26 | Up |
| LINC01512 | Long intergenic non-protein coding RNA 1512 | 5.46 | 6.26 | Up |
| LOC102723703 | Uncharacterized LOC102723703 | 7.55 | 8.88 | Up |
| NMBR | Neuromedin B receptor | 5.46 | 6.26 | Up |
| PCP2 | Purkinje cell protein 2 | 5.05 | 6.26 | Up |
| RASGRP1 | RAS guanyl releasing protein 1 | 5.46 | 8.88 | Up |
| SMC2-AS1 | SMC2 antisense RNA 1 (head to head) | 9.92 | 6.26 | Up |
| SMN2 | Survival of motor neuron 2, centromeric | 190.25 | 22.01 | Up |
| TRIM6-TRIM34 | TRIM6-TRIM34 readthrough | 117.78 | 48.70 | Up |
| ZC3H12B | Zinc finger CCCH-type containing 12B | 5.48 | 6.26 | Up |
| ZNF32-AS2 | ZNF32 antisense RNA 2 | 7.69 | 6.26 | Up |
| ZNF664-FAM101A | NA | 10.03 | 8.99 | Up |
| C16orf71 | Chromosome 16 open reading frame 71 | 0.19 | 0.12 | Down |
| CELF5 | CUGBP Elav-like family member 5 | 0.14 | 0.16 | Down |
| FAM53B-AS1 | FAM53B antisense RNA 1 | 0.19 | 0.12 | Down |
| GAL3ST2 | Galactose-3-O-sulfotransferase 2 | 0.14 | 0.16 | Down |
| KATNBL1P6 | Katanin regulatory subunit B1-like 1 pseudogene 6 | 0.11 | 0.11 | Down |
| LOC100506136 | NA | 0.14 | 0.05 | Down |
| LOC101926897 | NA | 0.20 | 0.16 | Down |
| LOC101927178 | Uncharacterized LOC101927178 | 0.15 | 0.12 | Down |
| LOC101927666 | Uncharacterized LOC101927666 | 0.20 | 0.09 | Down |
| MOG | Myelin oligodendrocyte glycoprotein | 0.11 | 0.16 | Down |
| NRN1L | Neuritin 1 like | 0.19 | 0.12 | Down |
| PROSER2-AS1 | PROSER2 antisense RNA 1 | 0.14 | 0.12 | Down |
| RALGAPA1P | Ral GTPase activating protein catalytic alpha subunit 1 pseudogene 1 | 0.01 | 0.02 | Down |
| RGPD6 | RANBP2-like and GRIP domain containing 6 | 0.12 | 0.20 | Down |
| RIMS4 | Regulating synaptic membrane exocytosis 4 | 0.19 | 0.16 | Down |
| THRIL | TNF- and HNRNPL-related immunoregulatory long non-coding RNA | 0.19 | 0.16 | Down |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tseng, C.-C.; Tsou, C.-H.; Huang, S.-Y.; Wu, C.-W.; Hsieh, T.-H. Using Next-Generation Sequencing and Bioinformatic Methods to Predict New Genes That May Be Regulated by CD47 in Oral Squamous Cell Carcinoma. Curr. Issues Mol. Biol. 2022, 44, 2243-2256. https://doi.org/10.3390/cimb44050152
Tseng C-C, Tsou C-H, Huang S-Y, Wu C-W, Hsieh T-H. Using Next-Generation Sequencing and Bioinformatic Methods to Predict New Genes That May Be Regulated by CD47 in Oral Squamous Cell Carcinoma. Current Issues in Molecular Biology. 2022; 44(5):2243-2256. https://doi.org/10.3390/cimb44050152
Chicago/Turabian StyleTseng, Chung-Chih, Chen-Han Tsou, Shi-Ying Huang, Chia-Wei Wu, and Tsung-Hua Hsieh. 2022. "Using Next-Generation Sequencing and Bioinformatic Methods to Predict New Genes That May Be Regulated by CD47 in Oral Squamous Cell Carcinoma" Current Issues in Molecular Biology 44, no. 5: 2243-2256. https://doi.org/10.3390/cimb44050152
APA StyleTseng, C. -C., Tsou, C. -H., Huang, S. -Y., Wu, C. -W., & Hsieh, T. -H. (2022). Using Next-Generation Sequencing and Bioinformatic Methods to Predict New Genes That May Be Regulated by CD47 in Oral Squamous Cell Carcinoma. Current Issues in Molecular Biology, 44(5), 2243-2256. https://doi.org/10.3390/cimb44050152