
Carbonic Anhydrase II Activators in Osteopetrosis Treatment: A Review


Abstract
:1. Introduction
2. Review
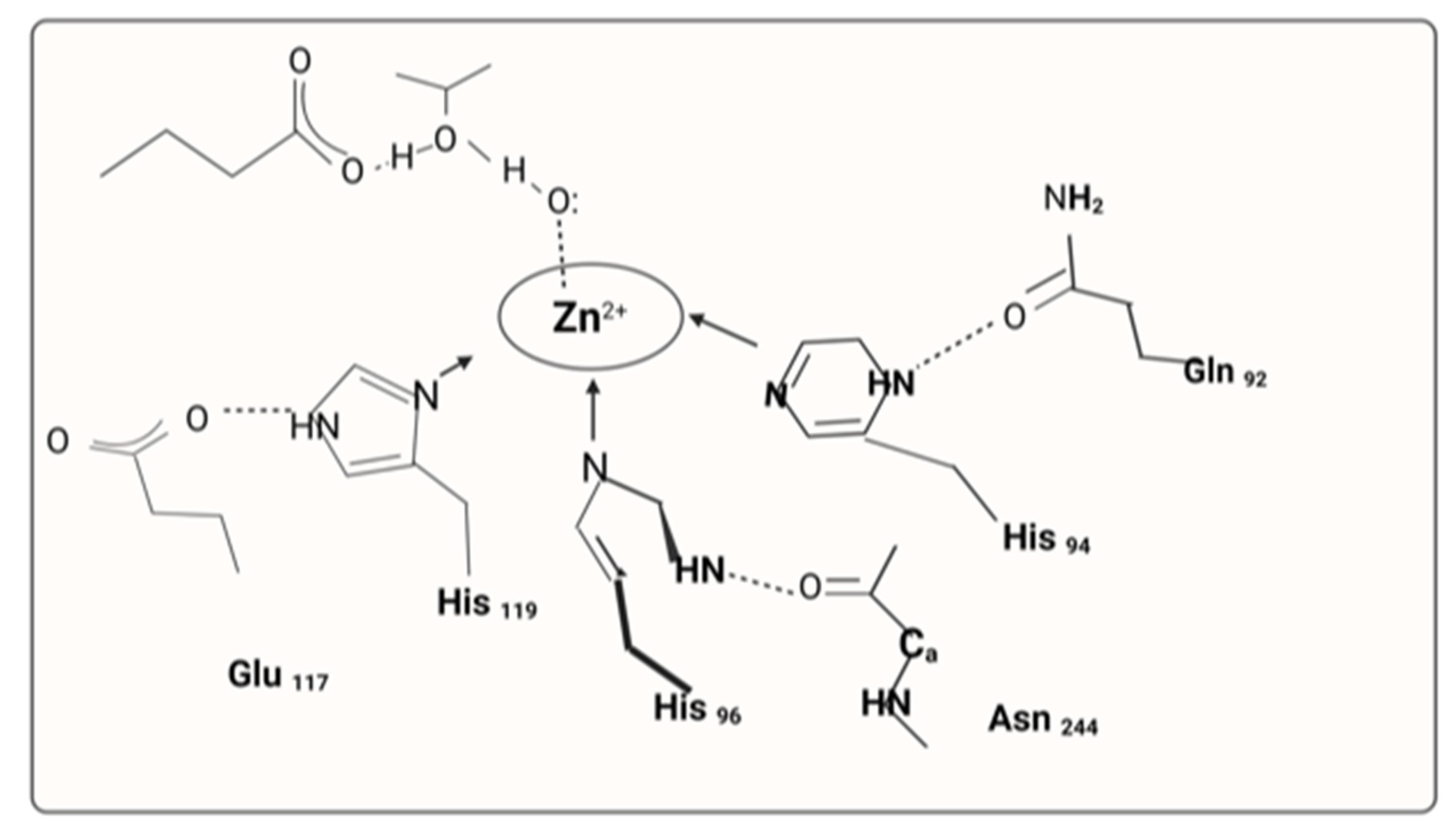
2.1. Carbonic Anhydrase II Activators
2.2. Osteopetrosis
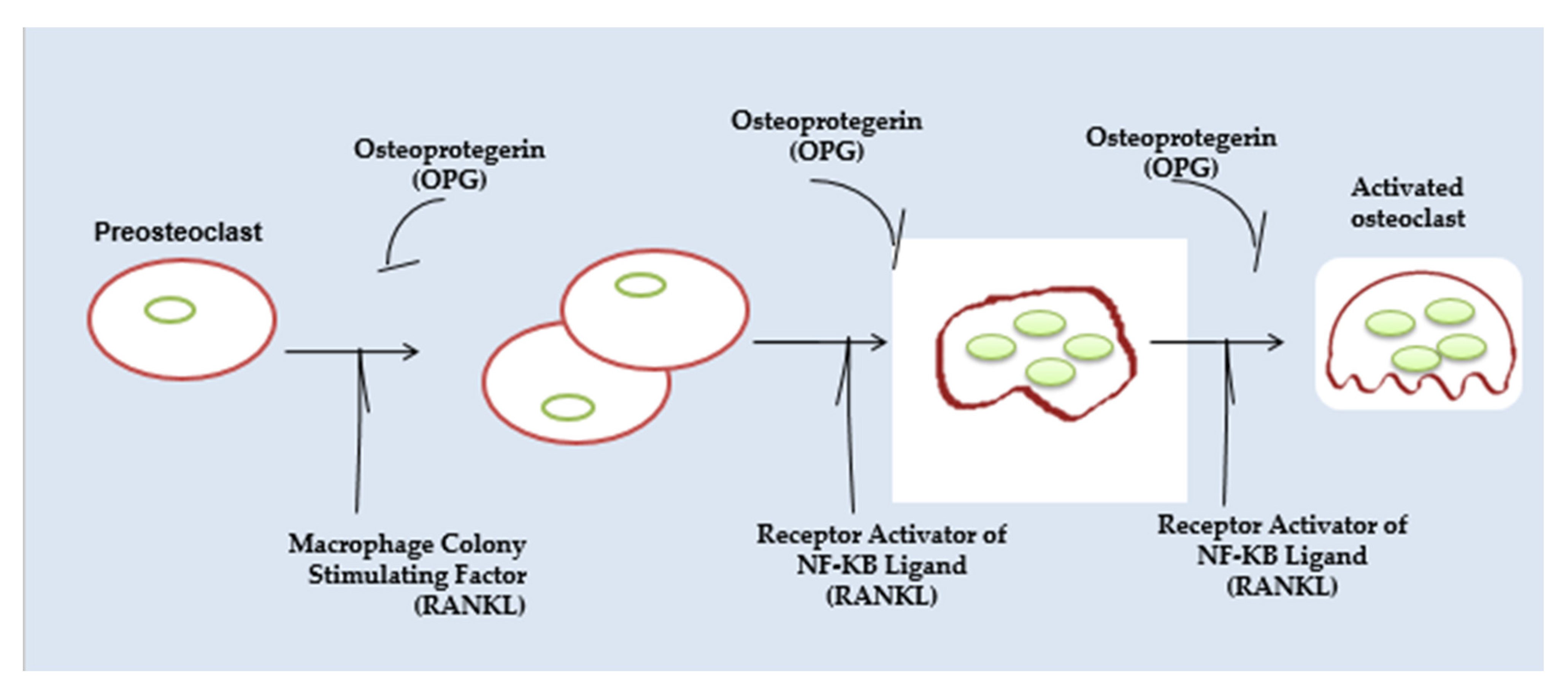
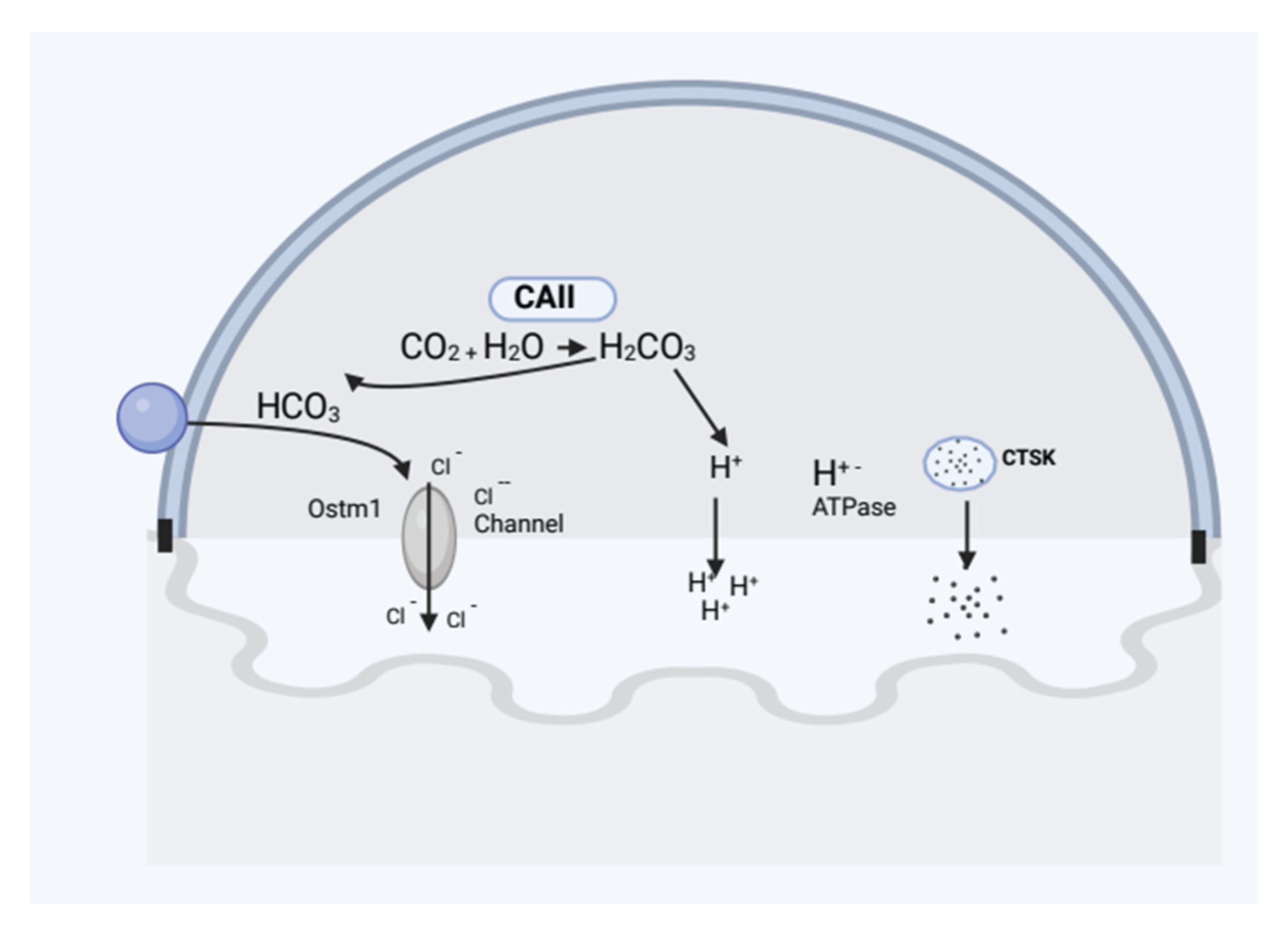
2.2.1. Osteoclast Function
2.2.2. Types of Osteopetrosis
Lack of Carbonic Anhydrase II (CAII) Causes Autosomal Recessive Osteopetrosis
Osteopetrosis with Autosomal Dominance
X-Linked Cases (XLO)
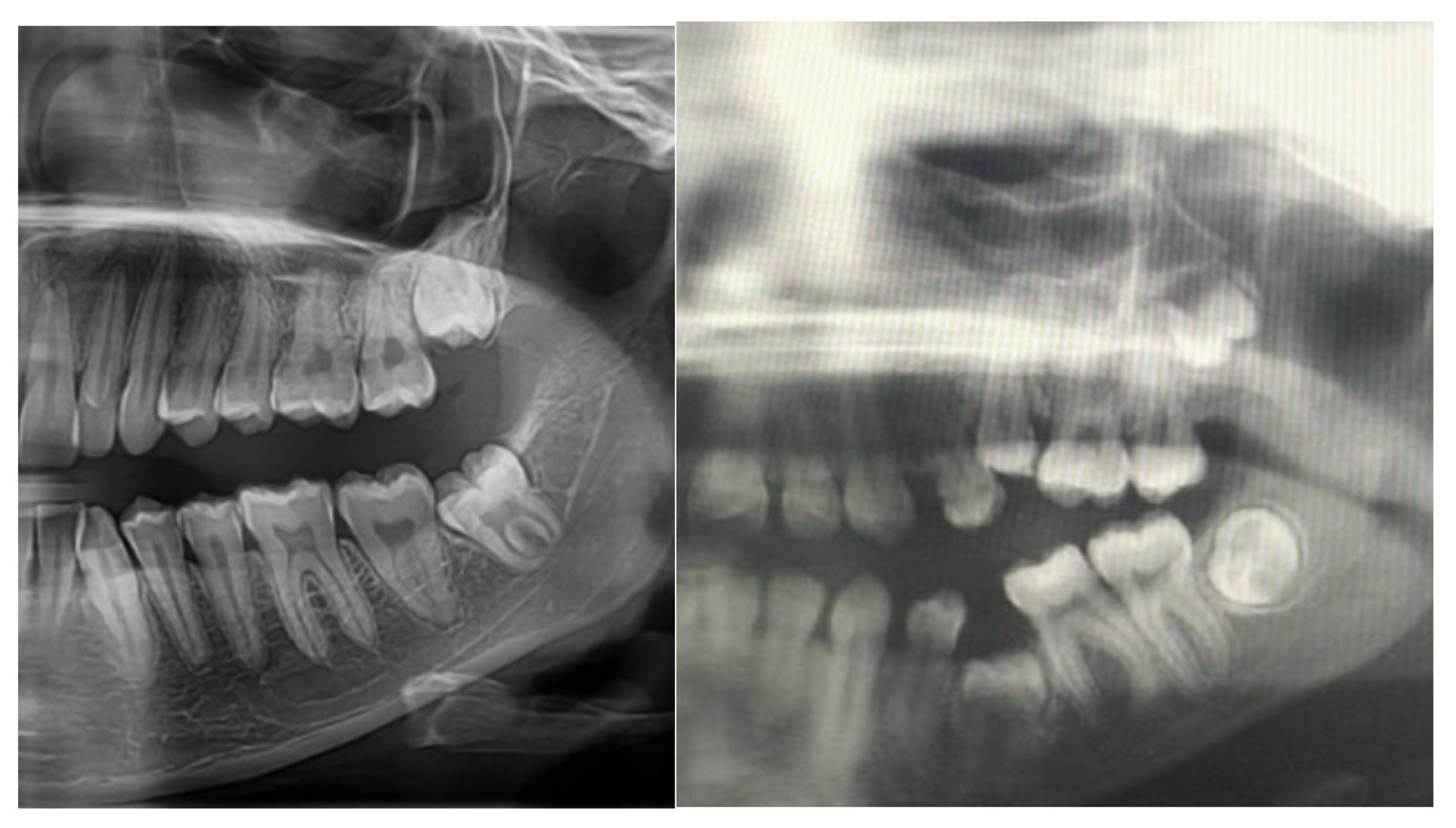
2.3. Pathogenesis of Dental Abnormalities
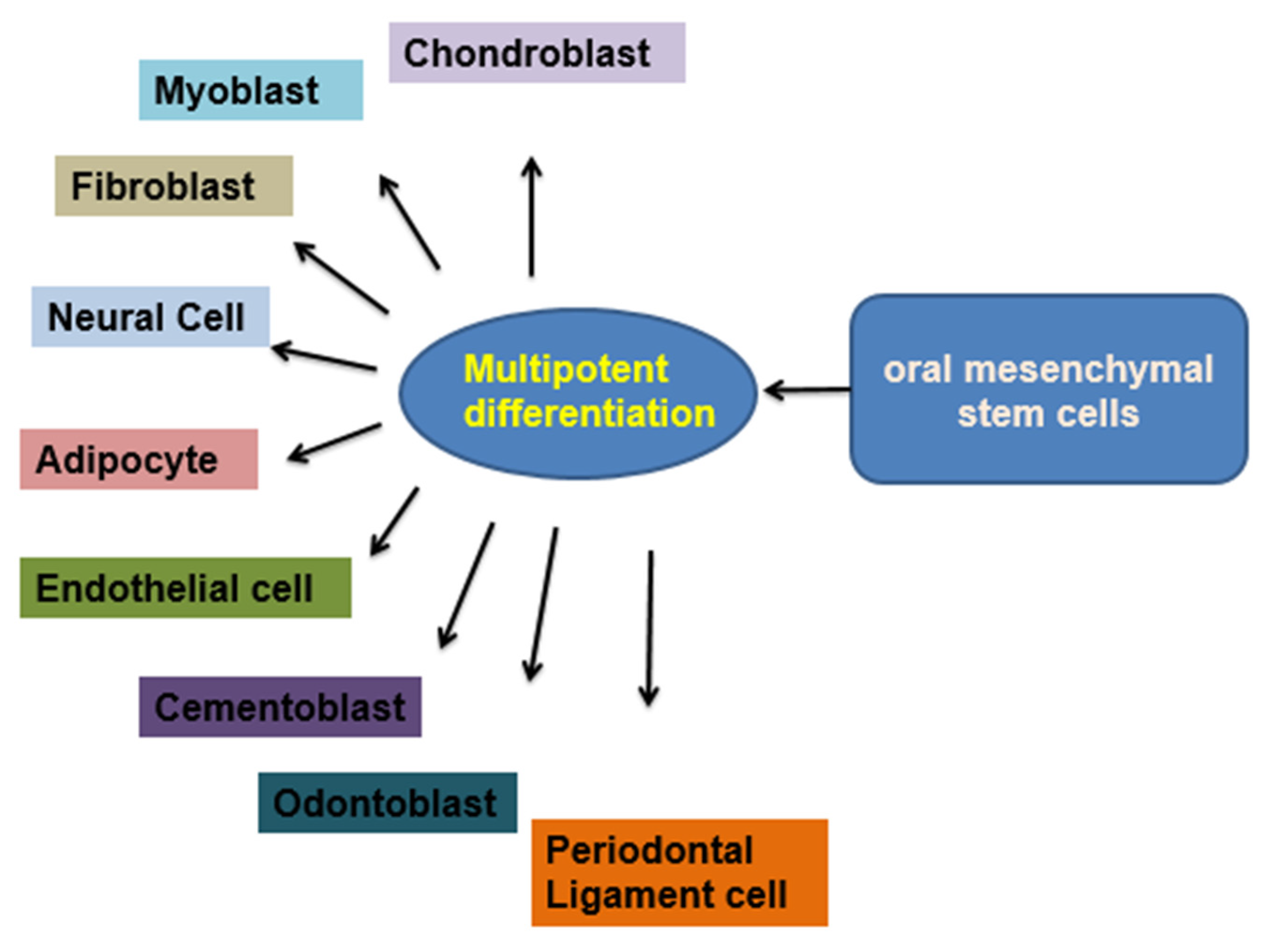
2.4. Dental Pulp Stem Cells (DPSCs)
2.5. Role of Azole and Flucanozole in Osteopetrosis
2.6. Development of Gene Therapy for Osteopetrosis
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hasselby, H.; Danielsson, K.; Sjöström, M. Oral Health in Patients with Osteopetrosis: A Literature Review. Clin. Surg. 2022, 7, 3411. [Google Scholar]
- Alkhayal, Z.; Shinwari, Z.; Gaafar, A.; Alaiya, A. Proteomic Profiling of the First Human Dental Pulp Mesenchymal Stem/Stromal Cells from Carbonic Anhydrase II Deficiency Osteopetrosis Patients. Int. J. Mol. Sci. 2021, 22, 380. [Google Scholar] [CrossRef]
- Allal, S.; Rabuel, V.; Gengler, C.; Douchet, C.; Allal, F.; Zwetyenga, N. Case report of osteomyelitis of the mandible in osteopetrosis and management considerations. Int. J. Surg. Case Rep. 2021, 81, 105813. [Google Scholar] [CrossRef] [PubMed]
- Nocentini, A.; Supuran, C.T. Carbonic anhydrase inhibitors as antitumor/antimetastatic agents: A patent review (2008–2018). Expert Opin. Ther. Pat. 2018, 28, 729–740. [Google Scholar] [CrossRef]
- Kim, J.K.; Kim, J.Y.; Jung, H.D.; Jung, Y.S. Surgical-orthodontic treatment for severe malocclusion in a patient with osteopetrosis and bilateral cleft lip and palate: Case report with a 5-year follow-up. Angle Orthod. 2021, 91, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Ishaque, A.; Farid, E.; Nasir, S.; Qadar, L.T.; Jamal, A. Intermediate Autosomal Recessive Osteopetrosis with an Unusual Absence of Fractures. Ochsner J. 2022, 22, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Ingole, S.N.; Deshpande, M.D.; Kazi, N.; Meshram, D.; Ranadive, P. A Rare Case of Osteoclast-poor Osteopetrosis (RANKL Mutation) with Recurrent Osteomyelitis of Mandible: A Case Report. Int. J. Clin. Pediatr. Dent. 2020, 13, 717. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, E.; López-Arcas, J.M.; Navarro, I.; Pingarrón, L.; Cebrián, J.L. Maxillomandibular osteomyelitis in osteopetrosis. Report of a case and review of the literature. Oral. Maxillofac. Surg. 2009, 13, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Schweikl, C.; Maier-Wohlfart, S.; Schneider, H.; Park, J. Ectodysplasin A1 Deficiency Leads to Osteopetrosis-like Changes in Bones of the Skull Associated with Diminished Osteoclastic Activity. Int. J. Mol. Sci. 2022, 23, 12189. [Google Scholar] [CrossRef]
- Yang, Y.; Tang, N.; Zhu, Y.; Zhang, L.; Cao, X.; Liu, L.; Xia, W.; Li, P.; Yang, Y. A novel homozygous nonsense mutation in the CA2 gene (c. 368G>A, p.W123X) linked to carbonic anhydrase II deficiency syndrome in a Chinese family. Metab. Brain Dis. 2021, 36, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Sallies, M.; Titinchi, F.; Morkel, J.A. Osteopetrosis complicated by osteomyelitis of the maxilla: A rare case report and review of the literature. Dent. Med. Probl. 2020, 57, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Satapathy, A.K.; Pandey, S.; Chaudhary, M.R.; Bagga, A.; Kabra, M.; Uwe, K.; Gupta, N. Report of another mutation proven case of carbonic anhydrase II deficiency. J. Pediatr. Genet. 2019, 8, 91–94. [Google Scholar] [CrossRef]
- Di Palmo, E.; Gallucci, M.; Tronconi, E.; Bergamaschi, R.; Cazzato, S.; La Scola, C.; Ricci, G.; Pession, A. Carbonic anhydrase II deficiency: A rare case of severe obstructive sleep apnea. Front. Pediatr. 2018, 6, 213. [Google Scholar] [CrossRef] [PubMed]
- Al-Shammari, N.R.; Temsah, M.H.; Al Shuaibi, W. Early Diagnosis of Osteopetrosis Type 3 with Neonatal onset with Life-Threatening Complications: A case Report with Review of the Literature. J. Middle East North Afr. Sci. 2020, 6, 18–20. [Google Scholar]
- Chávez-Güitrón, L.E.; Cerón-Torres, T.; Sobacchi, C.; Ochoa-Ruiz, E.; Villegas-Huesca, S. Autosomal recessive osteopetrosis type I: Description of pathogenic variant of TCIRG1 gene. Bol. Médico Hosp. Infant. México 2018, 75, 255–259. [Google Scholar]
- Alsemari, A.; Alsuhaibani, M.; Alhathlool, R.; Ali, B.M. Potential oligogenic disease of mental retardation, short stature, spastic paraparesis, and osteopetrosis. Appl. Clin. Genet. 2018, 11, 129. [Google Scholar] [CrossRef] [PubMed]
- Aspatwar, A.; Tolvanen, M.E.; Barker, H.; Syrjänen, L.; Valanne, S.; Purmonen, S.; Waheed, A.; Sly, W.S.; Parkkila, S. Carbonic anhydrases in metazoan model organisms: Molecules, mechanisms, and physiology. Physiol. Rev. 2022, 102, 1327–1383. [Google Scholar] [CrossRef]
- Athanasiadou, E.; Vlachou, C.; Theocharidou, A.; Tilaveridis, I.; Vargiami, E.; Antoniadis, K.; Arapostathis, K. When a pedodontic examination leads to the diagnosis of osteopetrosis: A case report. Spec. Care Dent. 2020, 40, 113–120. [Google Scholar] [CrossRef]
- Akleyin, E.; Polat, Y.; Yavuz, Y. Three-Year Dentition Follow-up of a Paediatric Case with Malignant Infantile Osteopetrosis: A Review of the Literature. J. Clin. Trials Exp. Investig. 2022, 1, 41–48. [Google Scholar]
- Peraza-Delgado, A.; Sánchez-Gómez, M.B.; Gómez-Salgado, J.; Romero-Martín, M.; Novo-Muñoz, M.; Duarte-Clíments, G. Non-Pharmacological Interventions towards Preventing the Triad Osteoporosis-Falls Risk-Hip Fracture, in Population Older than 65. Scoping Review. J. Clin. Med. 2020, 9, 2329. [Google Scholar] [CrossRef]
- Omi, M.; Mishina, Y. Role of osteoclasts in oral homeostasis and jawbone diseases. Oral Sci. Int. 2021, 18, 14–27. [Google Scholar] [CrossRef]
- Raghunath Reddy, M.H.; Vivekananda, M.R. Case report Albers schonberg disease (osteopetrosis) in a young adult—A rare case report. RGUHS J. Dent. Sci. 2018, 10, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Penna, S.; Capo, V.; Palagano, E.; Sobacchi, C.; Villa, A. One disease, many genes: Implications for the treatment of osteopetroses. nFront. Endocrinol. 2019, 10, 85. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhang, P.; Zhang, X.; Lv, L.; Zhou, Y. Advances in mesenchymal stem cell transplantation for the treatment of osteoporosis. Cell Prolif. 2021, 54, e12956. [Google Scholar] [CrossRef] [PubMed]
- Jimi, E.; Katagiri, T. Critical Roles of NF-κB Signaling Molecules in Bone Metabolism Revealed by Genetic Mutations in Osteopetrosis. Int. J. Mol. Sci. 2022, 23, 7995. [Google Scholar] [CrossRef]
- Huybrechts, Y.; Van Hul, W. Osteopetrosis associated with PLEKHM1 and SNX10 genes, both involved in osteoclast vesicular trafficking. Bone 2022, 164, 116520. [Google Scholar] [CrossRef] [PubMed]
- Beishuizen, R.; Caliskan, N.; Gröne, A.; Boroffka, S.A.; Tryfonidou, M.A.; Meij, B.P. Osteopetrosis in a Domestic Shorthair Cat. VCOT Open 2021, 4, e79–e85. [Google Scholar] [CrossRef]
- Tanaka, Y. Managing osteoporosis and joint damage in patients with rheumatoid arthritis: An overview. J. Clin. Med. 2021, 10, 1241. [Google Scholar] [CrossRef]
- Fitz, E.; Cohen, B.D. The Role of FSH in Osteoporosis and Subsequent Oral Disease. FASEB J. 2020, 34, 1. [Google Scholar] [CrossRef]
- Shanks, G.; Sharma, D.; Mishra, V. Prevention and treatment of osteoporosis in women. Obstet. Gynaecol. Reprod. Med. 2019, 29, 201–206. [Google Scholar] [CrossRef]
- Huang, X.L.; Huang, L.Y.; Cheng, Y.T.; Li, F.; Zhou, Q.; Wu, C.; Shi, Q.H.; Guan, Z.Z.; Liao, J.; Hong, W. Zoledronic acid inhibits osteoclast differentiation and function through the regulation of NF-κB and JNK signalling pathways. Int. J. Mol. Med. 2019, 44, 582–592. [Google Scholar] [CrossRef]
- Katz, J.; Rotstein, I. Prevalence of periapical lesions in patients with osteoporosis. J. Endod. 2021, 47, 234–238. [Google Scholar] [CrossRef]
- Dunphy, L.; Warfield, A.; Williams, R. Osteomyelitis of the mandible secondary to malignant infantile osteopetrosis in an adult. BMJ Case Rep. CP 2019, 12, e224452. [Google Scholar] [CrossRef]
- Selvakumar, R.; Chandran, A.; Patil, A.; Harini, T.C.; Dandekeri, S.; Verma, D.; Babu, J.S.; Swarnalatha, C.; Nayyar, A.S. Osteoporosis risk group: Screening for osteoporosis in dental clinics using panoramic radiographs. J. Educ. Health Promot. 2022, 11, 271. [Google Scholar] [PubMed]
- Peng, C.H.; Lin, W.Y.; Yeh, K.T.; Chen, H.; Wu, W.T.; Lin, M.D. The molecular etiology and treatment of glucocorticoid-induced osteoporosis. Tzu-Chi Med. J. 2021, 33, 212. [Google Scholar]
- Föger-Samwald, U.; Dovjak, P.; Azizi-Semrad, U.; Kerschan-Schindl, K.; Pietschmann, P. Osteoporosis: Pathophysiology and therapeutic options. EXCLI J. 2020, 19, 1017. [Google Scholar] [PubMed]
- Minamoto, C.; Miyazawa, K.; Tabuchi, M.; Hirano, M.; Mizuno, M.; Yoshizako, M.; Torii, Y.; Asano, Y.; Sato, T.; Kawatani, M.; et al. Alteration of tooth movement by reveromycin A in osteoprotegerin-deficient mice. Am. J. Orthod. Dentofac. Orthop. 2020, 157, 680–689. [Google Scholar] [CrossRef] [PubMed]
- Xin, Y.; Liu, Y.; Liu, D.; Li, J.; Zhang, C.; Wang, Y.; Zheng, S. New function of runx2 in regulating osteoclast differentiation via the akt/nfatc1/ctsk axis. Calcif. Tissue Int. 2020, 106, 553–566. [Google Scholar] [CrossRef]
- McDonald, M.M.; Kim, A.S.; Mulholland, B.S.; Rauner, M. New insights into osteoclast biology. JBMR Plus 2021, 5, e10539. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yamauchi, K.; Mitsunaga, T. A review on osteoclast diseases and osteoclastogenesis inhibitors recently developed from natural resources. Fitoterapia 2020, 142, 104482. [Google Scholar] [CrossRef] [PubMed]
- Baroncelli, G.I.; Angiolini, M.; Ninni, E.; Galli, V.; Saggese, R.; Giuca, M.R. Prevalence and pathogenesis of dental and periodontal lesions in children with X-linked hypophosphatemic rickets. Eur. J. Paediatr. Dent. 2006, 7, 61–66. [Google Scholar] [PubMed]
- Heboyan, A.; Avetisyan, A.; Karobari, M.I.; Marya, A.; Khurshid, Z.; Rokaya, D.; Zafar, M.S.; Fernandes, G.V.D.O. Tooth root resorption: A review. Sci. Prog. 2006, 105, 00368504221109217. [Google Scholar] [CrossRef] [PubMed]
- Alqahtani, K.A.; Shaheen, E.; Morgan, N.; Shujaat, S.; Politis, C.; Jacobs, R. Impact of orthognathic surgery on root resorption: A systematic review. J. Stomatol. Oral Maxillofac. Surg. 2022, 123, e260–e267. [Google Scholar] [CrossRef] [PubMed]
- Unnam, D.; Singaraju, G.S.; Mandava, P.; Reddy, G.V.; Mallineni, S.K.; Nuuvula, S. Accelerated orthodontics—An overview. J. Dent. Craniofac. Res. 2018, 3, 4. [Google Scholar] [CrossRef]
- Lee, S.; Kim, D.; Jeong, H.G. Detecting 17 fine-grained dental anomalies from panoramic dental radiography using artificial intelligence. Sci. Rep. 2022, 12, 5172. [Google Scholar] [CrossRef] [PubMed]
- Bijle, M.N.; Mallineni, S.K.; Tsoi, J. Qualitative and Quantitative Micro-CT Analysis of Natal and Neonatal Teeth. Children 2022, 9, 560. [Google Scholar] [CrossRef]
- Marzouk, T.; Alves, I.L.; Wong, C.L.; DeLucia, L.; McKinney, C.M.; Pendleton, C.; Howe, B.J.; Marazita, M.L.; Peter, T.K.; Kopycka-Kedzierawski, D.T.; et al. Association between dental anomalies and orofacial clefts: A meta-analysis. JDR Clin. Transl. Res. 2021, 6, 368–381. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.; Shi, X.; Xiao, F.; Yang, Y.; Zhang, X.; Wang, L.S.; Wu, C.T.; Wang, H. Transplantation of hepatocyte growth factor–modified dental pulp stem cells prevents bone loss in the early phase of ovariectomy-induced osteoporosis. Hum. Gene Ther. 2018, 29, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Rolph, D.N.; Deb, M.; Kanji, S.; Greene, C.J.; Das, M.; Joseph, M.; Aggarwal, R.; Leblebicioglu, B.; Das, H. Ferutinin directs dental pulp-derived stem cells towards the osteogenic lineage by epigenetically regulating canonical Wnt signaling. Biochim. Biophys. Acta BBA—Mol. Basis Dis. 2020, 1866, 165314. [Google Scholar] [CrossRef]
- Okawa, R.; Yokota, Y.; Morita, Y.; Mikasa, Y.; Nakano, K. Osteomyelitis Possibly Caused by Exfoliation of Primary Teeth in a Patient with Osteopetrosis. Children 2022, 9, 1894. [Google Scholar] [CrossRef]
- Gofur, A.R.P.; Kusumo, A.D. A review of human dental pulp stem cells inhibit maxillary alveolar ridge reduction in osteoporosis senile patient. Biochem. Cell. Arch. 2020, 20, 3107–3111. [Google Scholar]
- Li, P.L.; Wang, Y.X.; Zhao, Z.D.; Li, Z.L.; Liang, J.W.; Wang, Q.; Yin, B.F.; Hao, R.C.; Han, M.Y.; Ding, L.; et al. Clinical-grade human dental pulp stem cells suppressed the activation of osteoarthritic macrophages and attenuated cartilaginous damage in a rabbit osteoarthritis model. Stem Cell Res. Ther. 2021, 12, 260. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.; Kim, J.E.; Park, S.O.; Kim, E.Y.; Oh, S.; Choi, H.; Yoon, D.; Min, H.J.; Kim, H.R.; Chang, E.J. Sphingosine-1-phosphate hinders the osteogenic differentiation of dental pulp stem cells in association with AKT signaling pathways. Int. J. Oral Sci. 2022, 14, 21. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Tang, C.; Chen, J.; Tang, H.; Zhang, L.; Tang, G. Changes in Bone Marrow Fatty Acids Early after Ovariectomy-Induced Osteoporosis in Rats and Potential Functions. Metabolites 2023, 13, 36. [Google Scholar] [CrossRef]
- Permaul, P.; Narla, A.; Hornick, J.L.; Pai, S.Y. Allogeneic hematopoietic stem cell transplantation for X-linked ectodermal dysplasia and immunodeficiency: Case report and review of outcomes. Immunol. Res. 2009, 44, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Even-Or, E.; NaserEddin, A.; Dinur Schejter, Y.; Shadur, B.; Zaidman, I.; Stepensky, P. Haploidentical stem cell transplantation with post-transplant cyclophosphamide for osteopetrosis and other nonmalignant diseases. Bone Marrow Transplant. 2021, 56, 434–441. [Google Scholar] [CrossRef] [PubMed]
- St-Pierre, J.; Dufresne, P.J.; Carignan, A.; Lévesque, É.; Bernard, F.; Longtin, J.; LeBlanc, L. Case series: Report of the first two human indigenous cases of Cryptococcus gattii infection in eastern Canada. Mycopathologia 2018, 183, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Löfvall, H.; Rothe, M.; Schambach, A.; Henriksen, K.; Richter, J.; Moscatelli, I. Hematopoietic stem cell-targeted neonatal gene therapy with a clinically applicable lentiviral vector corrects osteopetrosis in oc/oc mice. Hum. Gene Ther. 2019, 30, 1395–1404. [Google Scholar] [CrossRef] [PubMed]
- Vacher, J. OSTM1 pleiotropic roles from osteopetrosis to neurodegeneration. Bone 2022, 163, 116505. [Google Scholar] [CrossRef]
- Wu, C.C.; Econs, M.J.; DiMeglio, L.A.; Insogna, K.L.; Levine, M.A.; Orchard, P.J.; Miller, W.P.; Petryk, A.; Rush, E.T.; Shoback, D.M.; et al. Diagnosis and Management of Osteopetrosis: Consensus Guidelines From the Osteopetrosis Working Group. J. Clin. Endocrinol. Metab. 2017, 102, 3111–3123. [Google Scholar] [CrossRef]
- Bhandi, S.; Alkahtani, A.; Reda, R.; Mashyakhy, M.; Boreak, N.; Maganur, P.C.; Vishwanathaiah, S.; Mehta, D.; Vyas, N.; Patil, V.; et al. Parathyroid Hormone Secretion and Receptor Expression Determine the Age-Related Degree of Osteogenic Differentiation in Dental Pulp Stem Cells. J. Pers. Med. 2021, 11, 349. [Google Scholar] [CrossRef]
- Penna, S.; Villa, A.; Capo, V. Autosomal recessive osteopetrosis: Mechanisms and treatments. Dis. Model. Mech. 2021, 14, dmm048940. [Google Scholar] [CrossRef] [PubMed]
- Kenneth, S.; Booth, M.; Bramer, M. Managing Osteopetrosis in the Complex Polytrauma Orthopedic Patient. Cureus 2022, 14, e21886. [Google Scholar]
- Charoenngam, N.; Nasr, A.; Shirvani, A.; Holick, M.F. Hereditary Metabolic Bone Diseases: A Review of Pathogenesis, Diagnosis and Management. Genes 2022, 13, 1880. [Google Scholar] [CrossRef]
- Reda, R.; Zanza, A.; Cicconetti, A.; Bhandi, S.; Miccoli, G.; Gambarini, G.; Di Nardo, D. Ultrasound Imaging in Dentistry: A Literature Overview. J. Imaging 2021, 7, 238. [Google Scholar] [CrossRef]
- Bénichou, O.; Cleiren, E.; Gram, J.; Bollerslev, J.; de Vernejoul, M.C.; Van Hul, W. Mapping of autosomal dominant osteopetrosis type II (Albers-Schönberg disease) to chromosome 16p13.3. Am. J. Hum. Genet. 2001, 69, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Coudert, A.E.; de Vernejoul, M.C.; Muraca, M.; Del Fattore, A. Osteopetrosis and its relevance for the discovery of new functions associated with the skeleton. Int. J. Endocrinol. 2015, 2015, 372156. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Human Osteopetrosis | Osteoclasts (OC) Features | Protein | Protein Function |
|---|---|---|---|---|
| Differentiation defects | ||||
| CSF1 | No | The number of OC is less | M-CSF | Cytokine (M-CSF) |
| CSFIR | No | The number of OC is less | C-FMS | MSC-F receptor |
| FOS | No | No OC | c-fos | Transcription factor |
| SPI1 | NO | No OC | PU.1 | Transcription factor |
| TNFRSF11A | No | No OC | RANK | RANKL receptor |
| TNFSF11 | ARO | No OC | RANKL | Cytokine |
| Fusion and polarization defects | ||||
| MITF | No | The number of OC is normal and no RB | MITF | Transcription factor |
| NFKB1/NFKB2 | No | The number of OC is less | NF-kB1/2 | Transcription factor |
| TRAF6 | No | The number of OC is normal and no RB | TRAF6 | Adaptor protein |
| TYROBP | No | The number of OC is less | DAP12 | Adaptor protein |
| Functional defects | ||||
| ACP5 | No | The number of OC is high and no RB | TRACP | Acid phosphatase |
| CLCN7 | ARO ADO | OC is enlarged with less developed RB | CLCN7 | Chloride channel |
| CTSK | Pycnodystosis | The number of OC is normal with irregular RB | Cathepsin K | Cathepsin |
| ITGBI | No | The number of OC is high and no RB | Itg | Integrin |
| OSTM1 | ARO | The number of OC is high with underdeveloped RB, and deformed cytoskeleton | OSTM1 | Subunit for CLCN7? |
| SRC | No | The number of OC is high and no RB | c-src | Tyrosine kinase |
| TCIRGI | ARO | The number of OC is high and no RB | TCIRGI | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alkhayal, Z.; Shinwari, Z.; Gaafar, A.; Alaiya, A. Carbonic Anhydrase II Activators in Osteopetrosis Treatment: A Review. Curr. Issues Mol. Biol. 2023, 45, 1373-1386. https://doi.org/10.3390/cimb45020089
Alkhayal Z, Shinwari Z, Gaafar A, Alaiya A. Carbonic Anhydrase II Activators in Osteopetrosis Treatment: A Review. Current Issues in Molecular Biology. 2023; 45(2):1373-1386. https://doi.org/10.3390/cimb45020089
Chicago/Turabian StyleAlkhayal, Zikra, Zakia Shinwari, Ameera Gaafar, and Ayodele Alaiya. 2023. "Carbonic Anhydrase II Activators in Osteopetrosis Treatment: A Review" Current Issues in Molecular Biology 45, no. 2: 1373-1386. https://doi.org/10.3390/cimb45020089
APA StyleAlkhayal, Z., Shinwari, Z., Gaafar, A., & Alaiya, A. (2023). Carbonic Anhydrase II Activators in Osteopetrosis Treatment: A Review. Current Issues in Molecular Biology, 45(2), 1373-1386. https://doi.org/10.3390/cimb45020089