
NOTCH Signaling in Osteosarcoma
Abstract
:1. Introduction
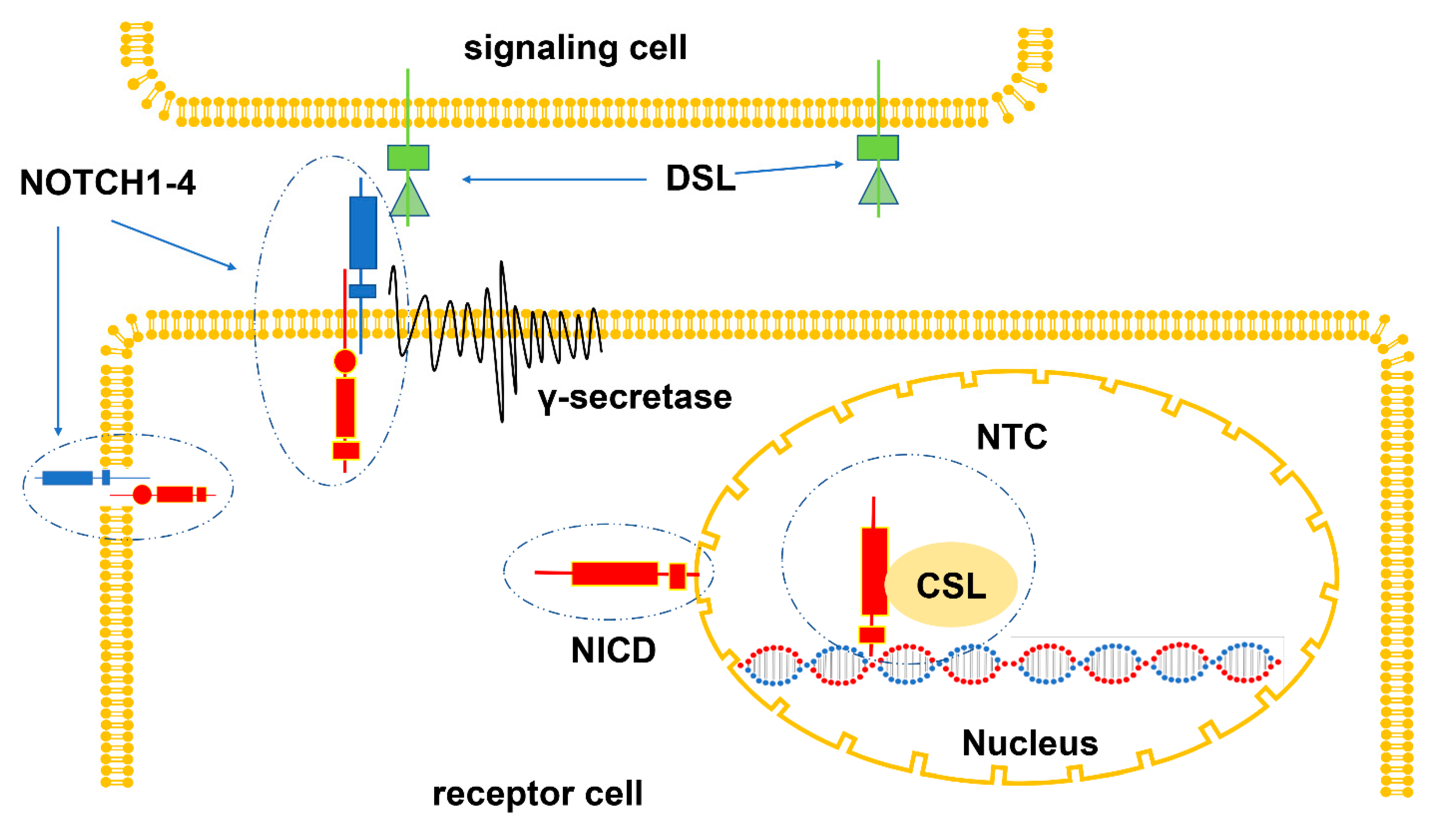
2. Composition of the NOTCH Signaling Pathway
3. Biological Functions of the NOTCH Signaling Pathway
4. Clinical Significance of Dysfunction of the NOTCH Signaling Pathway in Osteosarcoma Signaling
5. Effect of the NOTCH Signaling Pathway on Osteosarcoma
5.1. The NOTCH Signaling Pathway Regulates Osteogenic Differentiation in Osteosarcoma
5.2. The NOTCH Signaling Pathway Maintains the Self-Renewal Ability of Cancer Stem Cells in Osteosarcoma
5.3. The NOTCH Signaling Pathway Promotes Proliferation and Inhibits Apoptosis in Osteosarcoma

{kind=link}
{kind=link}
| Component | Gene | Study/Reference | Functional Status or Expression Level | Clinical Outcome | Cases | Detection Methods |
|---|---|---|---|---|---|---|
| Ligands | Jag1 | [30] | Upregulated | No report | 10 | RT-PCR |
| [16] | Upregulated | Increased metastasis rate and recurrence rate | 68 | IHC | ||
| [54] | Upregulated | No report | 10 | RT-PCR | ||
| Dll1 | [54] | Downregulated | No report | 10 | RT-PCR | |
| Receptors | NOTCH1 | [33] | Upregulated | Reduced cisplatin sensitivity; lower overall survival | 12 | IHC |
| [30] | Upregulated | No report | 10 | RT-PCR | ||
| [54] | Downregulated | No report | 10 | RT-PCR | ||
| [31] | High heterogeneity | Positively correlated with cisplatin sensitivity | 8 | IHC | ||
| NOTCH2 | [54] | Upregulated | No report | 10 | RT-PCR | |
| NOTCH3 | [32] | Upregulated | Lower survival rates; increased metastasis rates | 70 | IHC | |
| Downstream targets | Hes1 | [33] | Upregulated | Reduced cisplatin sensitivity; lower overall survival | 12 | IHC |
| [17] | Upregulated | Decreased survival rates | 16 | RT-PCR | ||
| [30] | Upregulated | No report | 10 | RT-PCR | ||
| Hey1 | [54] | Upregulated | No report | 10 | RT-PCR | |
| Hey2 | [30] | Upregulated | No report | 10 | RT-PCR | |
| [54] | Upregulated | No report | 10 | RT-PCR |
5.4. The NOTCH Signaling Pathway Promotes Tumor Metastasis and Invasion
5.5. The NOTCH Signaling Pathway Promotes Tumor Angiogenesis
5.6. The NOTCH Signaling Pathway Induces Chemoresistance
5.7. The NOTCH Signaling Pathway Regulates Immune Infiltration in Osteosarcoma Environment
6. NOTCH Signaling in Animal Models of Osteosarcoma
6.1. Animal Model of Spontaneous Osteosarcoma
6.2. Animal Transplantation Model of Human Osteosarcoma
6.3. Animal Transplantation Model of Allogeneic Osteosarcoma
6.4. Genetically Engineered Animal Osteosarcoma Model
7. Osteosarcoma Treatment Strategy Based on NOTCH Signaling
7.1. The Biological Agents Targeting the NOTCH Signaling Pathway
7.1.1. Monoclonal Antibodies
7.1.2. Blocking Peptides
7.2. γ-Secretase Inhibitors
7.3. Natural Products
| Drug | Target | Study Types | Study/Reference |
|---|---|---|---|
| Demcizumab | Dll4 | Clinical trial phase I | [101] |
| OMP-5948 | NOTCH2 and NOTCH3 | Clinical trial phase I | [98] |
| DAPT | γ-secretase | Studies performed in in vitro cells and pre-clinical animal models | [106,107,108] |
| RO4929097 | γ-secretase | Trial performed in pre-clinical animal models | [110] |
| Clinical trial phase I | [111] | ||
| Diallyl trisulfide | A natural product (non-selective inhibitors) | Studies performed in invitro cells | [77] |
| Oleanolic acid | A natural product (non-selective inhibitors) | Studies performed in invitro cells | [57] |
| Curcumin | A natural product (non-selective inhibitors) | Studies performed in invitro cells | [122] |
| Cinobufacin | A natural product (non-selective inhibitors) | Studies performed in invitro cells and pre-clinical animal models | [56] |
8. Discussion and Future Directions
| Component | Gene | Study/Reference | Major Effect | Cell Line | Animal Mode |
|---|---|---|---|---|---|
| Ligands | Jag1 | [16] | Inducing proliferation, migration, and invasion; | F5M2 | No |
| [55] | Inducing angiogenesis; promoting tumor growth and metastasis; | K7M2 | Orthotopic allograft mouse model and lung metastatic allograft mouse model | ||
| [91] | Inducing proliferation, migration, and invasion | 143B, SJSA1, SAOS2, U2OS, MG63 | Orthotopic xenograft mouse model | ||
| Dll1 | [17] | Inducing proliferation, migration, and invasion; promoting tumor growth and metastasis | OS 187, COL, LM7, SAOS2 | Orthotopic xenograft mouse model | |
| Receptors | NOTCH1 | [39] | Inducing osteoblast differentiation | MG63 | No |
| [40] | Inducing osteoblast carcinogenesis; inducing proliferation, migration, and invasion; inducing genomic instability | Primary osteosarcoma cells isolated form mice | Genetically engineered mouse model | ||
| [46] | Inducing the activity of cancer stem cell | 143B, MG63 | Subcutaneous xenograft mouse model | ||
| [47] | Promoting tumor recurrence and metastasis | hFOB, SAOS2, MG63, MNNG/HOS, LM5, HuO9, LM132 | Subcutaneous xenograft mouse model | ||
| [48] | Inducing the activity of cancer stem cell | U2OS, 143B | No | ||
| [49] | Inducing the activity of cancer stem cells; promoting tumor proliferation and recurrence | 143B, U2OS, MG63 | Subcutaneous xenograft mouse model | ||
| [55] | Inducing angiogenesis; promoting tumor proliferation and metastasis | K7M2 | Orthotopic allograft mouse model and lung metastatic allograft mouse model | ||
| [33] | Enhancing chemoresistance; inducing the activity of cancer stem cells; inducing proliferation, migration, and invasion; promoting tumor growth, recurrence, and metastasis | U2OS, MG63, 143B | Subcutaneous xenograft mouse model | ||
| [56] | Inhibiting apoptosis | U2OS, MG-63, 143B | Subcutaneous xenograft mouse model | ||
| [57] | Inhibiting apoptosis | SAOS2, MG63 | No | ||
| [58] | Inhibiting apoptosis | MG63 | No | ||
| [77] | Inducing angiogenesis | U2OS, SAOS2, MG63 | No | ||
| [31] | Enhancing chemosensitivity; inhibiting apoptosis | SAOS2, MG63 | No | ||
| [79] | Enhancing chemoresistance | 143B | No | ||
| [84] | Inhibiting the polarization of TAMs to the M2 phenotype | S180, mouse macrophages differentiated by primary bone marrow cells | Subcutaneous xenograft mouse model | ||
| [91] | Inducing proliferation, migration, and invasion | 143B, SJSA1, SAOS2, U2OS, MG63 | Orthotopic xenograft mouse model | ||
| [17] | Inducing proliferation, migration, and invasion; promoting tumor growth and metastasis | OS 187, COL, LM7, SAOS2 | Orthotopic xenograft mouse model | ||
| [30] | Inducing the activity of cancer stem cells; inducing proliferation, migration, and invasion; promoting tumor growth | SJSA1, SaOs2, CRL1423 | Subcutaneous xenograft mouse model | ||
| NOTCH2 | [39] | Inducing osteoblast differentiation | MG63 | No | |
| [59] | Inducing proliferation | 143B, U2OS, MG63, HOS, hFOB | No | ||
| [65] | Attenuating EMT; inducing proliferation, migration, and invasion; promoting tumor growth and metastasis | 143B | Subcutaneous xenograft mouse model | ||
| [17] | Inducing proliferation, migration, and invasion; promoting tumor growth and metastasis | OS 187, COL, LM7, SAOS2 | Orthotopic xenograft mouse model | ||
| NOTCH3 | [39] | Inducing osteoblast differentiation | MG63 | No | |
| [32] | Inducing proliferation, migration, and invasion; promoting tumor growth and metastasis | U2OS, hFOB1.19, MTH | Lung metastatic allograft mouse model | ||
| NOTCH4 | [39] | Inducing osteoblast differentiation | MG63 | No | |
| Downstream targets | Hey1 | [39] | Inducing osteoblast differentiation; | MG63 | No |
| [48] | Increasing the activity of cancer stem cell | U2OS, 143B | No | ||
| [49] | Increasing the activity of cancer stem cells; inducing proliferation; promoting tumor growth and recurrence | 143B, U2OS, MG63 | Subcutaneous xenograft mouse model | ||
| [65] | Attenuating EMT; inducing proliferation, migration, and invasion; promoting tumor growth and metastasis | 143B | Subcutaneous xenograft mouse model | ||
| [79] | Enhancing chemoresistance | 143B | No | ||
| Hey2 | [91] | Inducing proliferation, migration, and invasion | 143B, SJSA1, SAOS2, U2OS, MG63 | Orthotopic xenograft mouse model | |
| Hes1 | [46] | Inducing the activity of cancer stem cell | 143B, MG63 | Subcutaneous xenograft mouse model | |
| [47] | Promoting tumor growth, recurrence, and metastasis | hFOB, SAOS2, MG63, MNNG/HOS, LM5, HuO9, LM132 | Subcutaneous xenograft mouse model | ||
| [48] | Inducing the activity of cancer stem cell | U2OS, 143B | No | ||
| [49] | Inducing the activity of cancer stem cells; promoting tumor growth and recurrence | 143B, U2OS, MG63 | Subcutaneous xenograft mouse model | ||
| [54] | Inducing proliferation; promoting tumor growth | HOS, 143B, SAOS2, U2OS | Subcutaneous xenograft mouse model | ||
| [33] | Enhancing chemoresistance; inducing the activity of cancer stem cells; inducing proliferation, migration, and invasion; promoting tumor growth, recurrence, and metastasis | U2OS, MG63, 143B | Subcutaneous xenograft mouse model | ||
| [17] | Inducing proliferation, migration, and invasion; promoting tumor growth and metastasis | OS 187, COL, LM7, SAOS2 | Orthotopic xenograft mouse model | ||
| [32] | Inducing proliferation, migration, and invasion; promoting tumor growth and metastasis | U2OS, hFOB1.19, MTH | Lung metastatic allograft mouse model | ||
| [79] | Enhancing chemoresistance | 143B | No | ||
| [30] | Inducing the activity of cancer stem cells; inducing proliferation, migration, and invasion; promoting tumor growth | SJSA1, SaOs2, CRL1423 | Subcutaneous xenograft mouse model |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Yui, Y.; Itoh, K.; Yoshioka, K.; Naka, N.; Watanabe, M.; Hiraumi, Y.; Matsubara, H.; Watanabe, K.-I.; Sano, K.; Nakahata, T.; et al. Mesenchymal mode of migration participates in pulmonary metastasis of mouse osteosarcoma LM8. Clin. Exp. Metastasis 2010, 27, 619–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lallier, M.; Marchandet, L.; Moukengue, B.; Charrier, C.; Baud’Huin, M.; Verrecchia, F.; Ory, B.; Lamoureux, F. Molecular Chaperones in Osteosarcoma: Diagnosis and Therapeutic Issues. Cells 2021, 10, 754. [Google Scholar] [CrossRef] [PubMed]
- Bacci, G.; Longhi, A.; Versari, M.; Mercuri, M.; Briccoli, A.; Picci, P. Prognostic factors for osteosarcoma of the extremity treated with neoadjuvant chemotherapy: 15-year experience in 789 patients treated at a single institution. Cancer Interdiscip. Int. J. Am. Cancer Soc. 2006, 106, 1154–1161. [Google Scholar] [CrossRef]
- Chawla, S.P.; Goel, S.; Chow, W.; Braiteh, F.; Singh, A.S.; Olson, J.E.G.; Osada, A.; Bobe, I.; Riedel, R.F. A Phase 1b Dose Escalation Trial of NC-6300 (Nanoparticle Epirubicin) in Patients with Advanced Solid Tumors or Advanced, Metastatic, or Unresectable Soft-tissue Sarcoma. Clin. Cancer Res. 2020, 26, 4225–4232. [Google Scholar] [CrossRef]
- Davis, L.E.; Bolejack, V.; Ryan, C.W.; Ganjoo, K.N.; Loggers, E.T.; Chawla, S.; Agulnik, M.; Livingston, M.B.; Reed, D.; Keedy, V.; et al. Randomized Double-Blind Phase II Study of Regorafenib in Patients with Metastatic Osteosarcoma. J. Clin. Oncol. 2019, 37, 1424–1431. [Google Scholar] [CrossRef]
- Duffaud, F.; Mir, O.; Boudou-Rouquette, P.; Piperno-Neumann, S.; Penel, N.; Bompas, E.; Delcambre, C.; Kalbacher, E.; Italiano, A.; Collard, O.; et al. Efficacy and safety of regorafenib in adult patients with metastatic osteosarcoma: A non-comparative, randomised, double-blind, placebo-controlled, phase 2 study. Lancet Oncol. 2018, 20, 120–133. [Google Scholar] [CrossRef]
- Blay, J.-Y.; Duffaud, F.; George, S.; Maki, R.G.; Penel, N. Regorafenib for the Treatment of Sarcoma. Curr. Treat. Options Oncol. 2022, 23, 1477–1502. [Google Scholar] [CrossRef]
- Tawbi, H.A.; Burgess, M.; Bolejack, V.; Van Tine, B.A.; Schuetze, S.M.; Hu, J.; D’Angelo, S.; Attia, S.; Riedel, R.F.; Priebat, D.A.; et al. Pembrolizumab in advanced soft-tissue sarcoma and bone sarcoma (SARC028): A multicentre, two-cohort, single-arm, open-label, phase 2 trial. Lancet Oncol. 2017, 18, 1493–1501. [Google Scholar] [CrossRef]
- Grignani, G.; Palmerini, E.; Ferraresi, V.; D’Ambrosio, L.; Bertulli, R.; Asaftei, S.D.; Tamburini, A.; Pignochino, Y.; Sangiolo, D.; Marchesi, E.; et al. Sorafenib and everolimus for patients with unresectable high-grade osteosarcoma progressing after standard treatment: A non-randomised phase 2 clinical trial. Lancet Oncol. 2015, 16, 98–107. [Google Scholar] [CrossRef]
- Tang, N.; Song, W.-X.; Luo, J.; Haydon, R.C.; He, T.-C. Osteosarcoma Development and Stem Cell Differentiation. Clin. Orthop. Relat. Res. 2008, 466, 2114–2130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haydon, R.C.; Luu, H.H.; He, T.C. Osteosarcoma and osteoblastic differentiation: A new perspective on oncogenesis. Clin. Orthop. Relat. Res. 2007, 454, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Zieba, J.T.; Chen, Y.-T.; Lee, B.H.; Bae, Y. Notch Signaling in Skeletal Development, Homeostasis and Pathogenesis. Biomolecules 2020, 10, 332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aster, J.C.; Pear, W.S.; Blacklow, S.C. The Varied Roles of Notch in Cancer. Annu. Rev. Pathol. Mech. Dis. 2017, 12, 245–275. [Google Scholar] [CrossRef] [Green Version]
- McManus, M.M.; Weiss, K.R.; Hughes, D.P.M. Understanding the Role of Notch in Osteosarcoma. Curr. Adv. Osteosarcoma 2014, 804, 67–92. [Google Scholar] [CrossRef]
- Zhang, J.; Na Li, N.; Lu, S.; Chen, Y.; Shan, L.; Zhao, X.; Xu, Y. The role of Notch ligand Jagged1 in osteosarcoma proliferation, metastasis, and recurrence. J. Orthop. Surg. Res. 2021, 16, 226. [Google Scholar] [CrossRef]
- Hughes, D.P.M. How the NOTCH Pathway Contributes to the Ability of Osteosarcoma Cells to Metastasize. Pediatr. Adolesc. Osteosarcoma 2009, 152, 479–496. [Google Scholar] [CrossRef]
- Kopan, R.; Ilagan, M.X.G. The Canonical Notch Signaling Pathway: Unfolding the Activation Mechanism. Cell 2009, 137, 216–233. [Google Scholar] [CrossRef] [Green Version]
- Sprinzak, D.; Blacklow, S.C. Biophysics of Notch Signaling. Annu. Rev. Biophys. 2021, 50, 157–189. [Google Scholar] [CrossRef]
- Gama-Norton, L.; Ferrando, E.; Ruiz-Herguido, C.; Liu, Z.; Guiu, J.; Islam, A.B.M.M.K.; Lee, S.-U.; Yan, M.; Guidos, C.J.; Lopez-Bigas, N.; et al. Notch signal strength controls cell fate in the haemogenic endothelium. Nat. Commun. 2015, 6, 8510. [Google Scholar] [CrossRef] [Green Version]
- Nandagopal, N.; Santat, L.A.; LeBon, L.; Sprinzak, D.; Bronner, M.E.; Elowitz, M.B. Dynamic Ligand Discrimination in the Notch Signaling Pathway. Cell 2018, 172, 869–880.e19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van de Walle, I.; Waegemans, E.; De Medts, J.; De Smet, G.; De Smedt, M.; Snauwaert, S.; Vandekerckhove, B.; Kerre, T.; Leclercq, G.; Plum, J.; et al. Specific Notch receptor–ligand interactions control human TCR-αβ/γδ development by inducing differential Notch signal strength. J. Exp. Med. 2013, 210, 683–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artavanis-Tsakonas, S.; Muskavitch, M.A. Notch: The past, the present, and the future. Curr. Top. Dev. Biol. 2010, 92, 1–29. [Google Scholar] [PubMed]
- Zweidler-McKay, P.A.; Pear, W.S. Notch and T cell malignancy. Semin. Cancer Biol. 2004, 14, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, Y.; Maitra, A.; Ghosh, B.; Zechner, U.; Argani, P.; Iacobuzio-Donahue, C.A.; Sriuranpong, V.; Iso, T.; Meszoely, I.M.; Wolfe, M.S.; et al. Notch mediates TGFα-induced changes in epithelial differentiation during pancreatic tumorigenesis. Cancer Cell 2003, 3, 565–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Es, J.H.; Van Gijn, M.E.; Riccio, O.; Van Den Born, M.; Vooijs, M.; Begthel, H.; Cozijnsen, M.; Robine, S.; Winton, D.J.; Radtke, F.; et al. Notch/γ-secretase inhibition turns proliferative cells in intestinal crypts and adenomas into goblet cells. Nature 2005, 435, 959–963. [Google Scholar] [CrossRef]
- Zweidler-McKay, P.; He, Y.; Xu, L.; Rodriguez, C.G.; Karnell, F.G.; Carpenter, A.C.; Aster, J.C.; Allman, D.; Pear, W.S. Notch signaling is a potent inducer of growth arrest and apoptosis in a wide range of B-cell malignancies. Blood 2005, 106, 3898–3906. [Google Scholar] [CrossRef]
- Proweller, A.; Tu, L.; Lepore, J.J.; Cheng, L.; Lu, M.M.; Seykora, J.; Millar, S.E.; Pear, W.S.; Parmacek, M.S. Impaired Notch Signaling Promotes De novo Squamous Cell Carcinoma Formation. Cancer Res. 2006, 66, 7438–7444. [Google Scholar] [CrossRef] [Green Version]
- Zage, P.E.; Nolo, R.; Fang, W.; Stewart, J.; Garcia-Manero, G.; Zweidler-McKay, P.A. Notch pathway activation induces neuroblastoma tumor cell growth arrest. Pediatr. Blood Cancer 2012, 58, 682–689. [Google Scholar] [CrossRef] [Green Version]
- Engin, F.; Bertin, T.; Ma, O.; Jiang, M.M.; Wang, L.; Sutton, R.E.; Donehower, L.A.; Lee, B. Notch signaling contributes to the pathogenesis of human osteosarcomas. Hum. Mol. Genet. 2009, 18, 1464–1470. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Jin, F.; Qin, A.; Hao, Y.; Dong, Y.; Ge, S.; Dai, K. Targeting Notch1 signaling pathway positively affects the sensitivity of osteosarcoma to cisplatin by regulating the expression and/or activity of Caspase family. Mol. Cancer 2014, 13, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.-F.; Cao, Y.; Peng, D.-B.; Zhao, G.-S.; Zeng, Y.; Gao, Z.-R.; Lv, Y.-F.; Guo, Q.-N. Overexpression of Notch3 is associated with metastasis and poor prognosis in osteosarcoma patients. Cancer Manag. Res. 2019, 11, 547–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Xia, K.; Gao, T.; Chen, J.; Zhang, Z.; Sun, X.; Simões, B.M.; Eyre, R.; Fan, Z.; Guo, W.; et al. The Notch Pathway Promotes Osteosarcoma Progression through Activation of Ephrin Reverse Signaling. Mol. Cancer Res. 2019, 17, 2383–2394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haydon, R.C.; Zhou, L.; Feng, T.; Breyer, B.; Cheng, H.; Jiang, W.; Ishikawa, A.; Peabody, T.; Montag, A.; Simon, M.A.; et al. Nuclear receptor agonists as potential differentiation therapy agents for human osteosarcoma. Clin. Cancer Res. 2002, 8, 1288–1294. [Google Scholar] [PubMed]
- Manara, M.C.; Baldini, N.; Serra, M.; Lollini, P.L.; De Giovanni, C.; Vaccari, M.; Argnani, A.; Benini, S.; Maurici, D.; Picci, P.; et al. Reversal of malignant phenotype in human osteosarcoma cells transduced with the alkaline phosphatase gene. Bone 2000, 26, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Andela, V.B.; Sheu, T.-J.; Puzas, E.J.; Schwarz, E.M.; O’Keefe, R.J.; Rosier, R.N. Malignant reversion of a human osteosarcoma cell line, Saos-2, by inhibition of NFκB. Biochem. Biophys. Res. Commun. 2002, 297, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Sharff, K.A.; Song, W.-X.; Luo, X.; Tang, N.; Luo, J.; Chen, J.; Bi, Y.; He, B.-C.; Huang, J.; Li, X.; et al. Hey1 Basic Helix-Loop-Helix Protein Plays an Important Role in Mediating BMP9-induced Osteogenic Differentiation of Mesenchymal Progenitor Cells. J. Biol. Chem. 2009, 284, 649–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilton, M.J.; Tu, X.; Wu, X.; Bai, S.; Zhao, H.; Kobayashi, T.; Kronenberg, H.M.; Teitelbaum, S.L.; Ross, F.P.; Kopan, R.; et al. Notch signaling maintains bone marrow mesenchymal progenitors by suppressing osteoblast differentiation. Nat. Med. 2008, 14, 306–314. [Google Scholar] [CrossRef] [Green Version]
- Ongaro, A.; Pellati, A.; Bagheri, L.; Rizzo, P.; Caliceti, C.; Massari, L.; De Mattei, M. Characterization of Notch Signaling During Osteogenic Differentiation in Human Osteosarcoma Cell Line MG63. J. Cell. Physiol. 2016, 231, 2652–2663. [Google Scholar] [CrossRef]
- Tao, J.; Jiang, M.-M.; Jiang, L.; Salvo, J.S.; Zeng, H.-C.; Dawson, B.; Bertin, T.K.; Rao, P.H.; Chen, R.; Donehower, L.A.; et al. Notch Activation as a Driver of Osteogenic Sarcoma. Cancer Cell 2014, 26, 390–401. [Google Scholar] [CrossRef] [Green Version]
- Visvader, J.E.; Lindeman, G.J. Cancer stem cells in solid tumours: Accumulating evidence and unresolved questions. Nat. Rev. Cancer 2008, 8, 755–768. [Google Scholar] [CrossRef] [PubMed]
- Lytle, N.K.; Barber, A.G.; Reya, T. Stem cell fate in cancer growth, progression and therapy resistance. Nat. Rev. Cancer 2018, 18, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Ni, J.; Beretov, J.; Graham, P.; Li, Y. Cancer stem cell in breast cancer therapeutic resistance. Cancer Treat. Rev. 2018, 69, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Hoey, T.; Yen, W.-C.; Axelrod, F.; Basi, J.; Donigian, L.; Dylla, S.; Fitch-Bruhns, M.; Lazetic, S.; Park, I.-K.; Sato, A.; et al. DLL4 Blockade Inhibits Tumor Growth and Reduces Tumor-Initiating Cell Frequency. Cell Stem Cell 2009, 5, 168–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, A.; Wyatt, D.; Bocchetta, M.; Li, J.; Filipovic, A.; Green, A.; Peiffer, D.S.; Fuqua, S.; Miele, L.A.-O.; Albain, K.S.; et al. Notch-1-PTEN-ERK1/2 signaling axis promotes HER2+ breast cancer cell proliferation and stem cell survival. Oncogene 2018, 37, 4489–4504. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, B.; Wang, X.; Shen, Y.; Li, J.; Li, S.; Li, Y.; Wang, R. Interleukin-24 inhibits the phenotype and tumorigenicity of cancer stem cell in osteosarcoma via downregulation Notch and Wnt/β-catenin signaling. J. Bone Oncol. 2021, 31, 100403. [Google Scholar] [CrossRef]
- Jin, H.; Luo, S.; Wang, Y.; Liu, C.; Piao, Z.; Xu, M.; Guan, W.; Li, Q.; Zou, H.; Tan, Q.-Y.; et al. miR-135b Stimulates Osteosarcoma Recurrence and Lung Metastasis via Notch and Wnt/β-Catenin Signaling. Mol. Ther. Nucleic Acids 2017, 8, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Hu, Y.; Wang, L.; Sun, X.; Yu, L.; Guo, W. Human umbilical vein endothelial cells derived-exosomes promote osteosarcoma cell stemness by activating Notch signaling pathway. Bioengineered 2021, 12, 11007–11017. [Google Scholar] [CrossRef]
- Yu, L.; Fan, Z.; Fang, S.; Yang, J.; Gao, T.; Simões, B.M.; Eyre, R.; Guo, W.; Clarke, R.B. Cisplatin selects for stem-like cells in osteosarcoma by activating Notch signaling. Oncotarget 2016, 7, 33055–33068. [Google Scholar] [CrossRef]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- Evan, G.I.; Vousden, K.H. Proliferation, cell cycle and apoptosis in cancer. Nature 2001, 411, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Cohen, B.J.; Shimizu, M.; Izrailit, J.; Ng, N.F.L.; Buchman, Y.; Pan, J.G.; Dering, J.; Reedijk, M. Cyclin D1 is a direct target of JAG1-mediated Notch signaling in breast cancer. Breast Cancer Res. Treat. 2009, 123, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Wilson, G.; Huang, B.; Peng, M.; Teng, G.; Zhang, D.; Zhang, R.; Ebert, M.P.A.; Chen, J.; Wong, B.C.Y.; et al. Silencing of Jagged1 inhibits cell growth and invasion in colorectal cancer. Cell Death Dis. 2014, 5, e1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, M.; Setoguchi, T.; Hirotsu, M.; Gao, H.; Sasaki, H.; Matsunoshita, Y.; Komiya, S. Inhibition of Notch pathway prevents osteosarcoma growth by cell cycle regulation. Br. J. Cancer 2009, 100, 1957–1965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, J.; Wang, R.; Zhao, C.; Wen, J.; Dong, H.; Wang, S.; Li, Y.; Zhao, Y.; Li, J.; Yang, Y.; et al. Notch signaling regulates osteosarcoma proliferation and migration through Erk phosphorylation. Tissue Cell 2019, 59, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Yu, L.; Dai, G.; Zhang, S.; Zhang, Z.; Gao, T.; Guo, W. Cinobufagin induces apoptosis of osteosarcoma cells through inactivation of Notch signaling. Eur. J. Pharmacol. 2017, 794, 77–84. [Google Scholar] [CrossRef]
- Xu, Y.; Shu, B.; Tian, Y.; Wang, G.; Wang, Y.; Wang, J.; Dong, Y. Oleanolic acid induces osteosarcoma cell apoptosis by inhibition of Notch signaling. Mol. Carcinog. 2018, 57, 896–902. [Google Scholar] [CrossRef]
- Chen, L.; Wang, J.; Li, J.-W.; Zhao, X.-W.; Tian, L.-F. LncRNA MEG3 inhibits proliferation and promotes apoptosis of osteosarcoma cells through regulating Notch signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 581–590. [Google Scholar]
- Zhou, S.; Yu, L.; Xiong, M.; Dai, G. LncRNA SNHG12 promotes tumorigenesis and metastasis in osteosarcoma by upregulating Notch2 by sponging miR-195-5p. Biochem. Biophys. Res. Commun. 2018, 495, 1822–1832. [Google Scholar] [CrossRef]
- Chambers, A.F.; Groom, A.C.; MacDonald, I.C. Dissemination and growth of cancer cells in metastatic sites. Nat. Rev. Cancer 2002, 2, 563–572. [Google Scholar] [CrossRef]
- Sahlgren, C.; Gustafsson, M.V.; Jin, S.; Poellinger, L.; Lendahl, U. Notch signaling mediates hypoxia-induced tumor cell migration and invasion. Proc. Natl. Acad. Sci. USA 2008, 105, 6392–6397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, M.; Zhang, L.; He, C.-S.; Xu, F.; Liu, J.-L.; Hu, Z.-H.; Zhao, L.-P.; Tian, Y. Activation of Notch-1 enhances epithelial-mesenchymal transition in gefitinib-acquired resistant lung cancer cells. J. Cell. Biochem. 2011, 113, 1501–1513. [Google Scholar] [CrossRef]
- Peinado, H.; Portillo, F.; Cano, A. Switching On-Off Snail: LOXL2 Versus GSK3? Cell Cycle 2005, 4, 1749–1752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Zhao, F.; Lu, J.; Li, T.; Yang, H.; Wu, C.; Liu, Y. Notch-1 signaling promotes the malignant features of human breast cancer through NF-κB activation. PLoS ONE 2014, 9, e95912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, G.; Liu, G.; Zheng, D.; Song, Q. Inhibition of the Notch signaling pathway attenuates progression of cell motility, metastasis, and epithelial-to-mesenchymal transition-like phenomena induced by low concentrations of cisplatin in osteosarcoma. Eur. J. Pharmacol. 2021, 899, 174058. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.-W.; Wu, T.-Y.; Yi, X.; Ren, W.-P.; Zhou, Z.-B.; Sun, Y.-Q.; Zhang, C.-Q. Prognostic significance of VEGF expression in osteosarcoma: A meta-analysis. Tumor Biol. 2013, 35, 155–160. [Google Scholar] [CrossRef]
- Lammli, J.; Fan, M.; Rosenthal, H.G.; Patni, M.; Rinehart, E.; Vergara, G.; Ablah, E.; Wooley, P.H.; Lucas, G.; Yang, S.-Y. Expression of Vascular Endothelial Growth Factor correlates with the advance of clinical osteosarcoma. Int. Orthop. 2012, 36, 2307–2313. [Google Scholar] [CrossRef] [Green Version]
- Ferrara, N.; Hillan, K.J.; Gerber, H.-P.; Novotny, W. Discovery and development of bevacizumab, an anti-VEGF antibody for treating cancer. Nat. Rev. Drug Discov. 2004, 3, 391–400. [Google Scholar] [CrossRef]
- Riccardi, C.A.-O.; Napolitano, E.A.-O.; Platella, C.; Musumeci, D.; Melone, M.A.B.; Montesarchio, D.A.-O. Anti-VEGF DNA-based aptamers in cancer therapeutics and diagnostics. Med. Res. Rev. 2021, 4, 464–506. [Google Scholar] [CrossRef]
- Ramasamy, S.K.; Kusumbe, A.P.; Wang, L.; Adams, R.H. Endothelial Notch activity promotes angiogenesis and osteogenesis in bone. Nature 2014, 507, 376–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siekmann, A.F.; Lawson, N. Notch signalling limits angiogenic cell behaviour in developing zebrafish arteries. Nature 2007, 445, 781–784. [Google Scholar] [CrossRef] [PubMed]
- Dufraine, J.; Funahashi, Y.; Kitajewski, J. Notch signaling regulates tumor angiogenesis by diverse mechanisms. Oncogene 2008, 27, 5132–5137. [Google Scholar] [CrossRef] [Green Version]
- Potente, M.; Gerhardt, H.; Carmeliet, P. Basic and Therapeutic Aspects of Angiogenesis. Cell 2011, 146, 873–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goel, H.L.; Mercurio, A.M. VEGF targets the tumour cell. Nat. Rev. Cancer 2013, 13, 871–882. [Google Scholar] [CrossRef]
- Hellström, M.; Phng, L.-K.; Hofmann, J.J.; Wallgard, E.; Coultas, L.; Lindblom, P.; Alva, J.; Nilsson, A.-K.; Karlsson, L.; Gaiano, N.; et al. Dll4 signalling through Notch1 regulates formation of tip cells during angiogenesis. Nature 2007, 445, 776–780. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, J.; Zhang, L.; Si, M.; Yin, H.; Li, J. Diallyl trisulfide inhibits proliferation, invasion and angiogenesis of osteosarcoma cells by switching on suppressor microRNAs and inactivating of Notch-1 signaling. Carcinogenesis 2013, 34, 1601–1610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, S.; Teoh, S.L. Notch signalling pathways and their importance in the treatment of cancers. Curr. Drug Targets 2018, 19, 128–143. [Google Scholar] [CrossRef]
- Mei, H.; Yu, L.; Ji, P.; Yang, J.; Fang, S.; Guo, W.; Liu, Y.; Chen, X. Doxorubicin activates the Notch signaling pathway in osteosarcoma. Oncol. Lett. 2015, 9, 2905–2909. [Google Scholar] [CrossRef] [Green Version]
- Meurette, O.; Mehlen, P. Notch Signaling in the Tumor Microenvironment. Cancer Cell 2018, 34, 536–548. [Google Scholar] [CrossRef] [Green Version]
- Dumars, C.; Ngyuen, J.-M.; Gaultier, A.; Lanel, R.; Corradini, N.; Gouin, F.; Heymann, D.; Heymann, M.-F. Dysregulation of macrophage polarization is associated with the metastatic process in osteosarcoma. Oncotarget 2016, 7, 78343–78354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ségaliny, A.I.; Mohamadi, A.; Dizier, B.; Lokajczyk, A.; Brion, R.; Lanel, R.; Amiaud, J.; Charrier, C.; Boisson-Vidal, C.; Heymann, D. Interleukin-34 promotes tumor progression and metastatic process in osteosarcoma through induction of angiogenesis and macrophage recruitment. Int. J. Cancer 2015, 137, 73–85. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Xian, M.; Xiang, S.; Xiang, D.; Shao, X.; Wang, J.; Cao, J.; Yang, X.; Yang, B.; Ying, M.; et al. All-Trans Retinoic Acid Prevents Osteosarcoma Metastasis by Inhibiting M2 Polarization of Tumor-Associated Macrophages. Cancer Immunol. Res. 2017, 5, 547–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, S.; Zhang, X.; Hu, Y.; Wu, J.; Ju, Y.; Sun, X.; Liu, Y.; Shan, B. Blocking the Notch signal transduction pathway promotes tumor growth in osteosarcoma by affecting polarization of TAM to M2 phenotype. Ann. Transl. Med. 2020, 8, 1057. [Google Scholar] [CrossRef] [PubMed]
- Mendes, N.; Carvalho, P.D.; Martins, F.; Mendonça, S.; Malheiro, A.R.; Ribeiro, A.; Carvalho, J.; Velho, S. Animal Models to Study Cancer and Its Microenvironment. Tumor Microenviron. Main Driv. Metab. Adapt. 2020, 1219, 389–401. [Google Scholar] [CrossRef]
- Guijarro, M.V.; Ghivizzani, S.C.; Gibbs, C.P. Animal Models in Osteosarcoma. Front. Oncol. 2014, 4, 189. [Google Scholar] [CrossRef] [Green Version]
- Maynard, L.H.; Humbert, O.; Peterson, C.W.; Kiem, H.-P. Genome editing in large animal models. Mol. Ther. 2021, 29, 3140–3152. [Google Scholar] [CrossRef]
- Yu, X.; Yustein, J.T.; Xu, J. Research models and mesenchymal/epithelial plasticity of osteosarcoma. Cell Biosci. 2021, 11, 94. [Google Scholar] [CrossRef]
- Wilk, S.; Zabielska-Koczywąs, K. Molecular Mechanisms of Canine Osteosarcoma Metastasis. Int. J. Mol. Sci. 2021, 22, 3639. [Google Scholar] [CrossRef]
- Dailey, D.D.; Anfinsen, K.P.; Pfaff, L.E.; Ehrhart, E.; Charles, J.B.; Bønsdorff, T.B.; Thamm, D.H.; Powers, B.E.; Jonasdottir, T.J.; Duval, D.L. HES1, a target of Notch signaling, is elevated in canine osteosarcoma, but reduced in the most aggressive tumors. BMC Vet. Res. 2013, 9, 130. [Google Scholar] [CrossRef] [Green Version]
- Bae, Y.; Zeng, H.; Chen, Y.; Ketkar, S.; Munivez, E.; Yu, Z.; Gannon, F.H.; Lee, B.H. miRNA-34c Suppresses Osteosarcoma Progression In Vivo by Targeting Notch and E2F. JBMR Plus 2022, 6, e10623. [Google Scholar] [CrossRef] [PubMed]
- Walkley, C.R.; Qudsi, R.; Sankaran, V.G.; Perry, J.A.; Gostissa, M.; Roth, S.I.; Rodda, S.J.; Snay, E.; Dunning, P.; Fahey, F.H.; et al. Conditional mouse osteosarcoma, dependent on p53 loss and potentiated by loss of Rb, mimics the human disease. Genes Dev. 2008, 22, 1662–1676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berman, S.D.; Calo, E.; Landman, A.S.; Danielian, P.S.; Miller, E.S.; West, J.C.; Fonhoue, B.D.; Caron, A.; Bronson, R.; Bouxsein, M.L.; et al. Metastatic osteosarcoma induced by inactivation of Rb and p53 in the osteoblast lineage. Proc. Natl. Acad. Sci. USA 2008, 105, 11851–11856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shih Ie, M.; Wang, T.L. Notch signaling, gamma-secretase inhibitors, and cancer therapy. Cancer Res. 2007, 67, 1879–1882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Previs, R.A.; Coleman, R.L.; Harris, A.L.; Sood, A.K. Molecular Pathways: Translational and Therapeutic Implications of the Notch Signaling Pathway in Cancer. Clin. Cancer Res. 2015, 21, 955–961. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Cain-Hom, C.; Choy, L.; Hagenbeek, T.J.; De Leon, G.P.; Chen, Y.; Finkle, D.; Venook, R.; Wu, X.; Ridgway, J.; et al. Therapeutic antibody targeting of individual Notch receptors. Nature 2010, 464, 1052–1057. [Google Scholar] [CrossRef] [Green Version]
- Aste-Amézaga, M.; Zhang, N.; Lineberger, J.E.; Arnold, B.A.; Toner, T.J.; Gu, M.; Huang, L.; Vitelli, S.; Vo, K.T.; Haytko, P.; et al. Characterization of Notch1 Antibodies That Inhibit Signaling of Both Normal and Mutated Notch1 Receptors. PLoS ONE 2010, 5, e9094. [Google Scholar] [CrossRef]
- Smith, D.C.; Chugh, R.; Patnaik, A.; Papadopoulos, K.P.; Wang, M.; Kapoun, A.M.; Xu, L.; Dupont, J.; Stagg, R.J.; Tolcher, A. A phase 1 dose escalation and expansion study of Tarextumab (OMP-59R5) in patients with solid tumors. Investig. New Drugs 2018, 37, 722–730. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.I.; Bendell, J.C.; Bullock, A.; LoConte, N.K.; Hatoum, H.; Ritch, P.; Hool, H.; Leach, J.W.; Sanchez, J.; Sohal, D.P.S.; et al. A randomized phase II trial of nab-paclitaxel and gemcitabine with tarextumab or placebo in patients with untreated metastatic pancreatic cancer. Cancer Med. 2019, 8, 5148–5157. [Google Scholar] [CrossRef] [Green Version]
- Espinoza, I.; Miele, L. Notch inhibitors for cancer treatment. Pharmacol. Ther. 2013, 139, 95–110. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.C.; Eisenberg, P.D.; Manikhas, G.; Chugh, R.; Gubens, M.A.; Stagg, R.J.; Kapoun, A.M.; Xu, L.; Dupont, J.; Sikic, B. A Phase I Dose Escalation and Expansion Study of the Anticancer Stem Cell Agent Demcizumab (Anti-DLL4) in Patients with Previously Treated Solid Tumors. Clin. Cancer Res. 2014, 20, 6295–6303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- KleinJan, A.; Tindemans, I.; Montgomery, J.E.; Lukkes, M.; de Bruijn, M.J.; van Nimwegen, M.; Bergen, I.; Moellering, R.E.; Hoogsteden, H.C.; Boon, L.; et al. The Notch pathway inhibitor stapled α-helical peptide derived from mastermind-like 1 (SAHM1) abrogates the hallmarks of allergic asthma. J. Allergy Clin. Immunol. 2017, 142, 76–85.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Micchelli, C.A.; Esler, W.P.; Kimberly, W.T.; Jack, C.; Berezovska, O.; Kornilova, A.; Hyman, B.T.; Perrimon, N.; Wolfe, M.S. γ-Secretase/presenilin inhibitors for Alzheimer’s disease phenocopy Notch mutations in Drosophila. FASEB J. 2003, 17, 79–81. [Google Scholar] [CrossRef] [Green Version]
- Li, J.Y.; Li, R.J.; Wang, H.D. γ-secretase inhibitor DAPT sensitizes t-AUCB-induced apoptosis of human glioblastoma cells in vitro via blocking the p38 MAPK/MAPKAPK2/Hsp27 pathway. Acta Pharmacol. Sin. 2014, 35, 825–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnamurthy, N.; Kurzrock, R. Targeting the Wnt/beta-catenin pathway in cancer: Update on effectors and inhibitors. Cancer Treat. Rev. 2017, 62, 50–60. [Google Scholar] [CrossRef]
- Li, J.; Wang, R.; Qin, J.; Zeng, H.; Wang, K.; He, Q.; Wang, D.; Wang, S. Confocal Raman Spectral Imaging Study of DAPT, a γ-secretase Inhibitor, Induced Physiological and Biochemical Reponses in Osteosarcoma Cells. Int. J. Med. Sci. 2020, 17, 577–590. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Li, J.; Wang, H.; Qin, J.; Zeng, H.; Wang, K.; Wang, S. Unveiling osteosarcoma responses to DAPT combined with cisplatin by using confocal Raman microscopy. Biomed. Opt. Express 2021, 12, 5514–5528. [Google Scholar] [CrossRef]
- Dai, G.; Deng, S.; Guo, W.; Yu, L.; Yang, J.; Zhou, S.; Gao, T. Notch pathway inhibition using DAPT, a γ-secretase inhibitor (GSI), enhances the antitumor effect of cisplatin in resistant osteosarcoma. Mol. Carcinog. 2018, 58, 3–18. [Google Scholar] [CrossRef]
- Huang, T.; Zhao, C.; Zhao, Y.; Zhou, Y.; Wang, L.; Hang, D. RO4929097 regulates RANKL-induced osteoclast formation and LPS-mediated bone resorption. Aging 2021, 13, 12526–12536. [Google Scholar] [CrossRef]
- Kolb, E.A.; Gorlick, R.; Keir, S.T.; Maris, J.M.; Lock, R.; Carol, H.; Kurmasheva, R.T.; Reynolds, C.P.; Kang, M.H.; Wu, J.; et al. Initial testing (stage 1) by the pediatric preclinical testing program of RO4929097, a γ-secretase inhibitor targeting notch signaling. Pediatr. Blood Cancer 2011, 58, 815–818. [Google Scholar] [CrossRef] [Green Version]
- Tolcher, A.W.; Messersmith, W.A.; Mikulski, S.M.; Papadopoulos, K.P.; Kwak, E.L.; Gibbon, D.G.; Patnaik, A.; Falchook, G.S.; Dasari, A.; Shapiro, G.I.; et al. Phase I Study of RO4929097, a Gamma Secretase Inhibitor of Notch Signaling, in Patients with Refractory Metastatic or Locally Advanced Solid Tumors. J. Clin. Oncol. 2012, 30, 2348–2353. [Google Scholar] [CrossRef] [PubMed]
- Mikaili, P.; Maadirad, S.; Moloudizargari, M.; Aghajanshakeri, S.; Sarahroodi, S. Therapeutic Uses and Pharmacological Properties of Garlic, Shallot, and Their Biologically Active Compounds. Iran. J. Basic Med. Sci. 2013, 16, 1031–1048. [Google Scholar] [CrossRef] [PubMed]
- Antony, M.L.; Singh, S.V. Molecular mechanisms and targets of cancer chemoprevention by garlic-derived bioactive compound diallyl trisulfide. Experiment 2011, 49, 805–816. [Google Scholar]
- Seki, T.; Hosono, T.; Hosono-Fukao, T.; Inada, K.; Tanaka, R.; Ogihara, J.; Ariga, T. Anticancer effects of diallyl trisulfide derived from garlic. Asia Pac. J. Clin. Nutr. 2008, 17, 249–252. [Google Scholar] [PubMed]
- He, P.; Wang, Z.; Sheng, B.; Xu, Y.; Feng, S.; Huang, Y.; Gong, F.; Tang, L.; Xie, L. Diallyl trisulfide regulates cell apoptosis and invasion in human Osteosarcoma U2OS cells through regulating PI3K/AKT/GSK3β signaling pathway. Histol. Histopathol. 2020, 35, 1511–1520. [Google Scholar]
- Lisiak, N.M.; Lewicka, I.; Kaczmarek, M.; Kujawski, J.; Bednarczyk-Cwynar, B.; Zaprutko, L.; Rubis, B. Oleanolic Acid’s Semisynthetic Derivatives HIMOXOL and Br-HIMOLID Show Proautophagic Potential and Inhibit Migration of HER2-Positive Breast Cancer Cells In Vitro. Int. J. Mol. Sci. 2021, 22, 11273. [Google Scholar] [CrossRef]
- Du, Z.; Li, G.; Zhou, X.; Zhang, J. Synthesis of MeON-Glycoside Derivatives of Oleanolic Acid by Neoglycosylation and Evaluation of Their Cytotoxicity against Selected Cancer Cell Lines. Molecules 2021, 26, 772. [Google Scholar] [CrossRef]
- Bao, Y.; Zhang, S.; Chen, Z.; Chen, A.T.; Ma, J.; Deng, G.; Xu, W.; Zhou, J.; Yu, Z.-Q.; Yao, G.; et al. Synergistic Chemotherapy for Breast Cancer and Breast Cancer Brain Metastases via Paclitaxel-Loaded Oleanolic Acid Nanoparticles. Mol. Pharm. 2020, 17, 1343–1351. [Google Scholar] [CrossRef]
- Patel, B.B.; Majumdar, A.P.N. Synergistic Role of Curcumin with Current Therapeutics in Colorectal Cancer: Minireview. Nutr. Cancer 2009, 61, 842–846. [Google Scholar] [CrossRef]
- Chakravarti, N.; Kadara, H.; Yoon, D.-J.; Shay, J.W.; Myers, J.N.; Lotan, D.; Sonenberg, N.; Lotan, R. Differential Inhibition of Protein Translation Machinery by Curcumin in Normal, Immortalized, and Malignant Oral Epithelial Cells. Cancer Prev. Res. 2010, 3, 331–338. [Google Scholar] [CrossRef] [Green Version]
- Walters, D.K.; Muff, R.; Langsam, B.; Born, W.; Fuchs, B. Cytotoxic effects of curcumin on osteosarcoma cell lines. Investig. New Drugs 2007, 26, 289–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zhang, J.; Ma, D.; Zhang, L.; Si, M.; Yin, H.; Li, J. Curcumin inhibits proliferation and invasion of osteosarcoma cells through inactivation of Notch-1 signaling. FEBS J. 2012, 279, 2247–2259. [Google Scholar] [CrossRef] [PubMed]
- Emam, H.; Zhao, Q.-L.; Furusawa, Y.; Refaat, A.; Ahmed, K.; Kadowaki, M.; Kondo, T. Apoptotic cell death by the novel natural compound, cinobufotalin. Chem. Interact. 2012, 199, 154–160. [Google Scholar] [CrossRef]
- Kai, S.; Lu, J.-H.; Hui, P.-P.; Zhao, H. Pre-clinical evaluation of cinobufotalin as a potential anti-lung cancer agent. Biochem. Biophys. Res. Commun. 2014, 452, 768–774. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Song, B.; Jin, H.; Pi, J.; Liu, L.; Jiang, J.; Cai, J. Cinobufacini induced MDA-MB-231 cell apoptosis-associated cell cycle arrest and cytoskeleton function. Bioorg. Med. Chem. Lett. 2012, 22, 1459–1463. [Google Scholar] [CrossRef]
- Yin, J.-H.; Zhu, X.-Y.; Shi, W.-D.; Liu, L.-M. Huachansu injection inhibits metastasis of pancreatic cancer in mice model of human tumor xenograft. BMC Complement. Altern. Med. 2014, 14, 483. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.-M.; Chan, W.Y.; Yu, S.; Zhao, J.; Cheng, C.H. Bufalin induces autophagy-mediated cell death in human colon cancer cells through reactive oxygen species generation and JNK activation. Free Radic. Biol. Med. 2011, 51, 1365–1375. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Wu, W.; Shao, Z. NOTCH Signaling in Osteosarcoma. Curr. Issues Mol. Biol. 2023, 45, 2266-2283. https://doi.org/10.3390/cimb45030146
Zhang Z, Wu W, Shao Z. NOTCH Signaling in Osteosarcoma. Current Issues in Molecular Biology. 2023; 45(3):2266-2283. https://doi.org/10.3390/cimb45030146
Chicago/Turabian StyleZhang, Zhenhao, Wei Wu, and Zengwu Shao. 2023. "NOTCH Signaling in Osteosarcoma" Current Issues in Molecular Biology 45, no. 3: 2266-2283. https://doi.org/10.3390/cimb45030146
APA StyleZhang, Z., Wu, W., & Shao, Z. (2023). NOTCH Signaling in Osteosarcoma. Current Issues in Molecular Biology, 45(3), 2266-2283. https://doi.org/10.3390/cimb45030146