
Abietane Diterpenoids Isolated from Torreya nucifera Disrupt Replication of Influenza Virus by Blocking the Phosphatidylinositol-3-Kinase (PI3K)-Akt and ERK Signaling Pathway



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell and Viruses
2.3. Time Course (Time-of-Addition) Antiviral Assays
2.4. Chemiluminescent Neuraminidase Inhibition (NAI) Assay
2.5. Quantitative Real-Time PCR (qRT-PCR)
2.6. Western Blotting Analysis
2.7. Confocal Fluorescence Imaging
2.8. Statistical Analysis
3. Results
3.1. Inhibition of the Influenza Virus Life Cycle by 18-Hydroxyferruginol (1) and 18-Oxoferruginol (2)
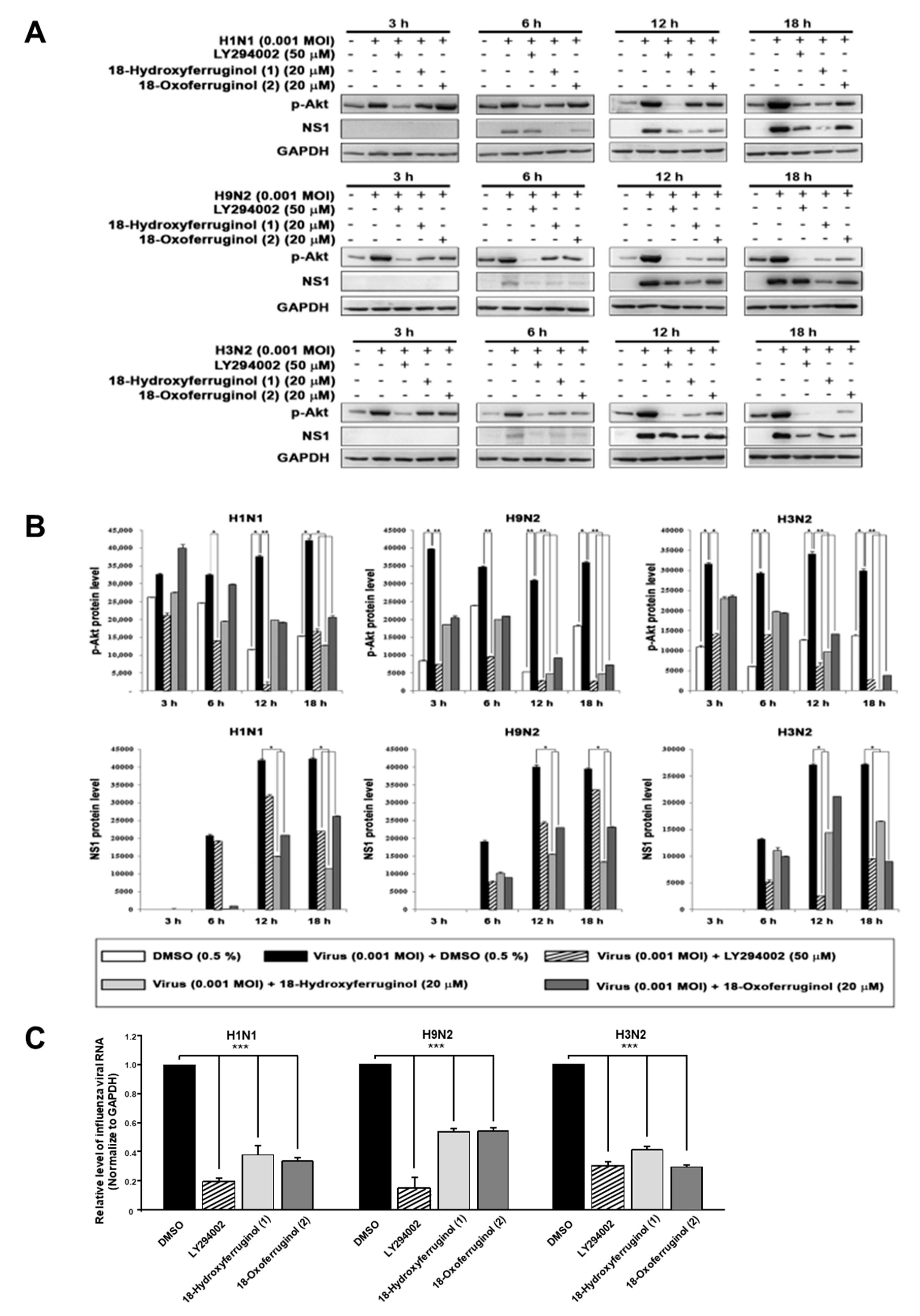
3.2. Inhibition of Influenza-Virus-Induced Activation of the PI3K-Akt Signaling Pathway by 18-Hydroxyferruginol (1) and 18-Oxoferruginol (2)
3.3. Inhibition of Influenza Ribonucleoprotein (RNP) Nuclear Export by 18-Hydroxyferruginol (1) and 18-Oxoferruginol (2)
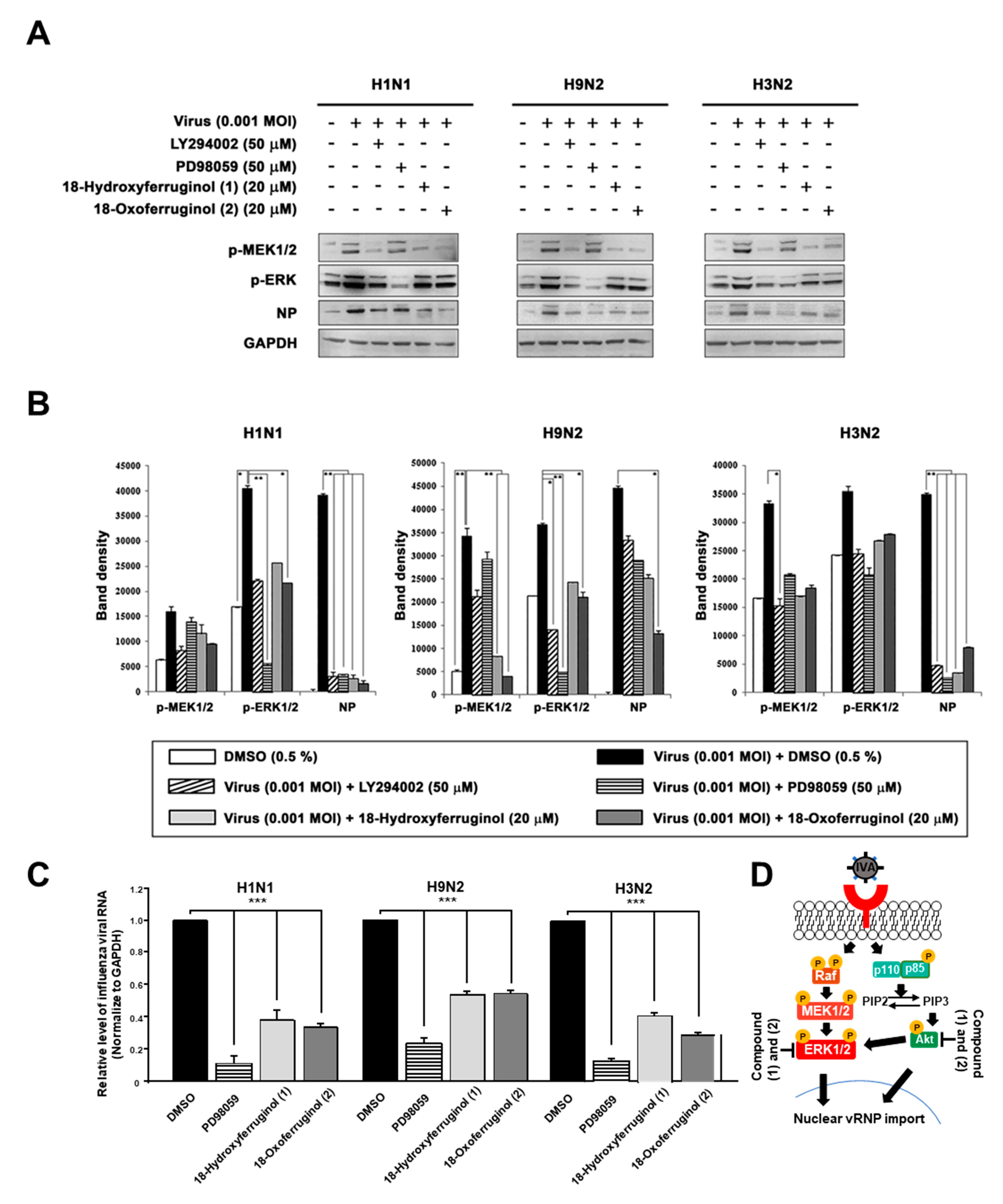
3.4. Downregulation of Influenza-Virus-Induced ERK Phosphorylation by 18-Hydroxyferruginol (1) and 18-Oxoferruginol (2) in MDCK Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shaw, M.L.; Palese, P. Orthomyxoviridae: The viruses and their replication. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 1691–1740. [Google Scholar]
- Palese, P. Influenza: Old and new threats. Nat. Med. 2004, 10, S82–S87. [Google Scholar] [CrossRef]
- Novel Swine-Origin Influenza A (H1N1) Virus Investigation Team; Dawood, F.S.; Jain, S.; Finelli, L.; Shaw, M.W.; Lindstrom, S.; Garten, R.J.; Gubareva, L.V.; Xu, X.; Bridges, C.B.; et al. Emergence of a Novel Swine-Origin Influenza A (H1N1) Virus in Humans. N. Engl. J. Med. 2009, 360, 2605–2615. [Google Scholar] [PubMed] [Green Version]
- Zimmer, S.M.; Burke, D.S. Historical Perspective—Emergence of Influenza A (H1N1) Viruses. N. Engl. J. Med. 2009, 361, 279–285. [Google Scholar] [CrossRef] [PubMed]
- O’Hanlon, R.; Shaw, M.L. Baloxavir marboxil: The new influenza drug on the market. Curr. Opin. Virol. 2019, 35, 14–18. [Google Scholar] [CrossRef]
- Hayden, F.G. Antiviral Resistance in Influenza Viruses—Implications for Management and Pandemic Response. N. Engl. J. Med. 2006, 354, 785–788. [Google Scholar] [CrossRef]
- Layne, S.P.; Monto, A.S.; Taubenberger, J.K. Pandemic influenza: An inconvenient mutation. Science 2009, 323, 1560–1561. [Google Scholar] [CrossRef] [Green Version]
- Cooray, S. The pivotal role of phosphatidylinositol 3-kinase-Akt signal transduction in virus survival. J. Gen. Virol. 2004, 85, 1065–1076. [Google Scholar] [CrossRef]
- Wu, M.S.; Yen, H.R.; Chang, C.W.; Peng, T.Y.; Hsieh, C.F.; Chen, C.J.; Lin, T.Y.; Horng, J.T. Mechanism of action of the suppression of influenza virus replication by Ko-Ken Tang through inhibition of the phosphatidylinositol 3-kinase/Akt signaling pathway and viral RNP nuclear export. J. Ethnopharmacol. 2011, 134, 614–623. [Google Scholar] [CrossRef]
- Cantrell, D.A. Phosphoinositide 3-kinase signalling pathways. J. Cell Sci. 2001, 114, 1439–1445. [Google Scholar] [CrossRef]
- Sarkar, S.N.; Peters, K.L.; Elco, C.P.; Sakamoto, S.; Pal, S.; Sen, G.C. Novel roles of TLR3 tyrosine phosphorylation and PI3 kinase in double-stranded RNA signaling. Nat. Struct. Mol. Biol. 2004, 11, 1060–1067. [Google Scholar] [CrossRef]
- Ehrhardt, C.; Marjuki, H.; Wolff, T.; Nürnberg, B.; Planz, O.; Pleschka, S.; Ludwig, S. Bivalent role of the phosphatidylinositol-3-kinase (PI3K) during influenza virus infection and host cell defence. Cell. Microbiol. 2006, 8, 1336–1348. [Google Scholar] [CrossRef] [PubMed]
- Ehrhardt, C.; Wolff, T.; Ludwig, S. Activation of Phosphatidylinositol 3-Kinase Signaling by the Nonstructural NS1 Protein Is Not Conserved among Type A and B Influenza Viruses. J. Virol. 2007, 81, 12097–12100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Zhang, Y.; Wang, S.; Liu, W.; Hao, C.; Wang, W. Inhibition effects of patchouli alcohol against influenza a virus through targeting cellular PI3K/Akt and ERK/MAPK signaling pathways. Virol. J. 2019, 16, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, R.; Herold, S.; Cakarova, L.; Hoegner, K.; Lohmeyer, J.; Planz, O.; Pleschka, S. Inhibition of influenza virus-induced NF-kappaB and Raf/MEK/ERK activation can reduce both virus titers and cytokine expression simultaneously in vitro and in vivo. Antivir. Res. 2011, 92, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Pleschka, S.; Wolff, T.; Ehrhardt, C.; Hobom, G.; Planz, O.; Rapp, U.R.; Ludwig, S. Influenza virus propagation is impaired by inhibition of the Raf/MEK/ERK signalling cascade. Nat. Cell Biol. 2001, 3, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Bae, K.H. The Medicinal Plants of Korea; Kyo-Hak Publishing Co., Ltd.: Seoul, Republic of Korea, 2000; Volume 45. [Google Scholar]
- Harrison, L.J.; Asakawa, Y. 18-Oxoferruginol from the leaf of Torreya nucifera. Phytochemistry 1987, 26, 1211–1212. [Google Scholar] [CrossRef]
- Orihara, Y.; Yang, J.W.; Komiya, N.; Koge, K.; Yoshikawa, T. Abietane diterpenoids from suspension cultured cells of Torreya nucifera var. radicans. Phytochemistry 2002, 59, 385–389. [Google Scholar] [CrossRef]
- Inatani, R.; Nakatani, N.; Fuwa, H. Antioxidative Effect of the Constituents of Rosemary (Rosmarinus officinalis L.) and Their Derivatives. Agric. Biol. Chem. 1983, 47, 521–528. [Google Scholar] [CrossRef]
- Ryu, Y.B.; Jeong, H.J.; Kim, J.H.; Kim, Y.M.; Park, J.Y.; Kim, D.; Nguyen, T.T.; Park, S.J.; Chang, J.S.; Park, K.H.; et al. Biflavonoids from Torreya nucifera displaying SARS-CoV 3CLpro inhibition. Bioorg. Med. Chem. 2010, 18, 7940–7947. [Google Scholar] [CrossRef]
- de Jesus, M.B.; Zambuzzi, W.F.; de Sousa, R.R.R.; Areche, C.; de Souza, A.C.S.; Aoyama, H.; Schmeda-Hirschmann, G.; Rodríguez, J.A.; de Souza Brito, A.R.M.; Peppelenbosch, M.P.; et al. Ferruginol suppresses survival signaling pathways in androgen-independent human prostate cancer cells. Biochimie 2008, 90, 843–854. [Google Scholar] [CrossRef]
- Ulubelen, A.; Oksuz, S.; Kolak, U.; Birman, H.; Voelter, W. Cardioactive terpenoids and a new rearranged diterpene from Salvia syriaca. Planta Med. 2000, 66, 627–629. [Google Scholar] [CrossRef]
- Kwon, H.J.; Ryu, Y.B.; Kim, Y.M.; Song, N.; Kim, C.Y.; Rho, M.C.; Jeong, J.H.; Cho, K.O.; Lee, W.S.; Park, S.J. In vitro antiviral activity of phlorotannins isolated from Ecklonia cava against porcine epidemic diarrhea coronavirus infection and hemagglutination. Bioorg. Med. Chem. 2013, 21, 4706–4713. [Google Scholar] [CrossRef]
- Chang, S.T.; Chen, P.F.; Wang, S.Y.; Wu, H. Antimite activity of essential oils and their constituents from Taiwania cryptomerioides. J. Med. Entomol. 2001, 38, 455–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujioka, Y.; Tsuda, M.; Hattori, T.; Sasaki, J.; Sasaki, T.; Miyazaki, T.; Ohba, Y. The Ras-PI3K Signaling Pathway Is Involved in Clathrin-Independent Endocytosis and the Internalization of Influenza Viruses. PLoS ONE 2011, 6, e16324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwig, S.; Planz, O.; Pleschka, S.; Wolff, T. Influenza-virus-induced signaling cascades: Targets for antiviral therapy? Trends Mol. Med. 2003, 9, 46–52. [Google Scholar] [CrossRef]
- Ludwig, S.; Pleschka, S.; Planz, O.; Wolff, T. Ringing the alarm bells: Signalling and apoptosis in influenza virus infected cells. Cell. Microbiol. 2006, 8, 375–386. [Google Scholar] [CrossRef]
- Shin, Y.K.; Liu, Q.; Tikoo, S.K.; Babiuk, L.A.; Zhou, Y. Effect of the phosphatidylinositol 3-kinase/Akt pathway on influenza A virus propagation. J. Gen. Virol. 2007, 88, 942–950. [Google Scholar] [CrossRef] [PubMed]
- Carracedo, A.; Pandolfi, P.P. The PTEN-PI3K pathway: Of feedbacks and cross-talks. Oncogene 2008, 27, 5527–5541. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extracts or Compounds | CC50 (μM) a | A/PR/8/34 (H1N1) | A/Chicken/Korea/MS96/96 (H9N2) | A/Hong Kong/8/68 (H3N2) | |||
|---|---|---|---|---|---|---|---|
| IC50 (μM) b | SI c | IC50 (μM) b | SI c | IC50 (μM) b | SI c | ||
| Olseltamivir | >500 | 1.35 ± 1.2 | >370 | <0.7 | >714 | 1.09 ± 0.5 | >458 |
| EtOH extract | 95.7 ± 1.2 μg/mL | 57.9 ± 3.5 μg/mL | 1.65 | - | - | - | - |
| 18-Hydroxyferruginol (1) | 35.5 ± 0.4 | 13.6 ± 1.4 | 2.61 | 12.8 ± 0.1 | 2.77 | - | - |
| 18-Oxoferruginol (2) | 41.2 ± 1.5 | 18.3 ± 6.5 | 2.25 | 10.8 ± 2.2 | 3.81 | 29.2 ± 1.9 | 1.41 |
| Compounds | Neuraminiase IC50 (μM) a | ||
|---|---|---|---|
| A/PR/8/34 (H1N1) | A/Chicken/Korea/MS96/96 (H9N2) | A/Hong Kong/8/68 (H3N2) | |
| Olseltamivir | 0.011 | 0.005 | 0.011 |
| 18-Hydroxyferruginol (1) | >100 | 82.2 | >100 |
| 18-Oxoferruginol (2) | >100 | >100 | >100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bae, J.; Kwon, H.-J.; Park, J.S.; Jung, J.; Ryu, Y.B.; Kim, W.S.; Lee, J.H.; Jeong, J.-H.; Lim, J.S.; Lee, W.S.; et al. Abietane Diterpenoids Isolated from Torreya nucifera Disrupt Replication of Influenza Virus by Blocking the Phosphatidylinositol-3-Kinase (PI3K)-Akt and ERK Signaling Pathway. Curr. Issues Mol. Biol. 2023, 45, 2284-2295. https://doi.org/10.3390/cimb45030147
Bae J, Kwon H-J, Park JS, Jung J, Ryu YB, Kim WS, Lee JH, Jeong J-H, Lim JS, Lee WS, et al. Abietane Diterpenoids Isolated from Torreya nucifera Disrupt Replication of Influenza Virus by Blocking the Phosphatidylinositol-3-Kinase (PI3K)-Akt and ERK Signaling Pathway. Current Issues in Molecular Biology. 2023; 45(3):2284-2295. https://doi.org/10.3390/cimb45030147
Chicago/Turabian StyleBae, Jaehoon, Hyung-Jun Kwon, Ji Sun Park, Jinseok Jung, Young Bae Ryu, Woo Sik Kim, Ju Huck Lee, Jae-Ho Jeong, Jae Sung Lim, Woo Song Lee, and et al. 2023. "Abietane Diterpenoids Isolated from Torreya nucifera Disrupt Replication of Influenza Virus by Blocking the Phosphatidylinositol-3-Kinase (PI3K)-Akt and ERK Signaling Pathway" Current Issues in Molecular Biology 45, no. 3: 2284-2295. https://doi.org/10.3390/cimb45030147
APA StyleBae, J., Kwon, H. -J., Park, J. S., Jung, J., Ryu, Y. B., Kim, W. S., Lee, J. H., Jeong, J. -H., Lim, J. S., Lee, W. S., & Park, S. -J. (2023). Abietane Diterpenoids Isolated from Torreya nucifera Disrupt Replication of Influenza Virus by Blocking the Phosphatidylinositol-3-Kinase (PI3K)-Akt and ERK Signaling Pathway. Current Issues in Molecular Biology, 45(3), 2284-2295. https://doi.org/10.3390/cimb45030147