

Plant Antimicrobial Peptides: Insights into Structure-Function Relationships for Practical Applications


Abstract
:1. Introduction
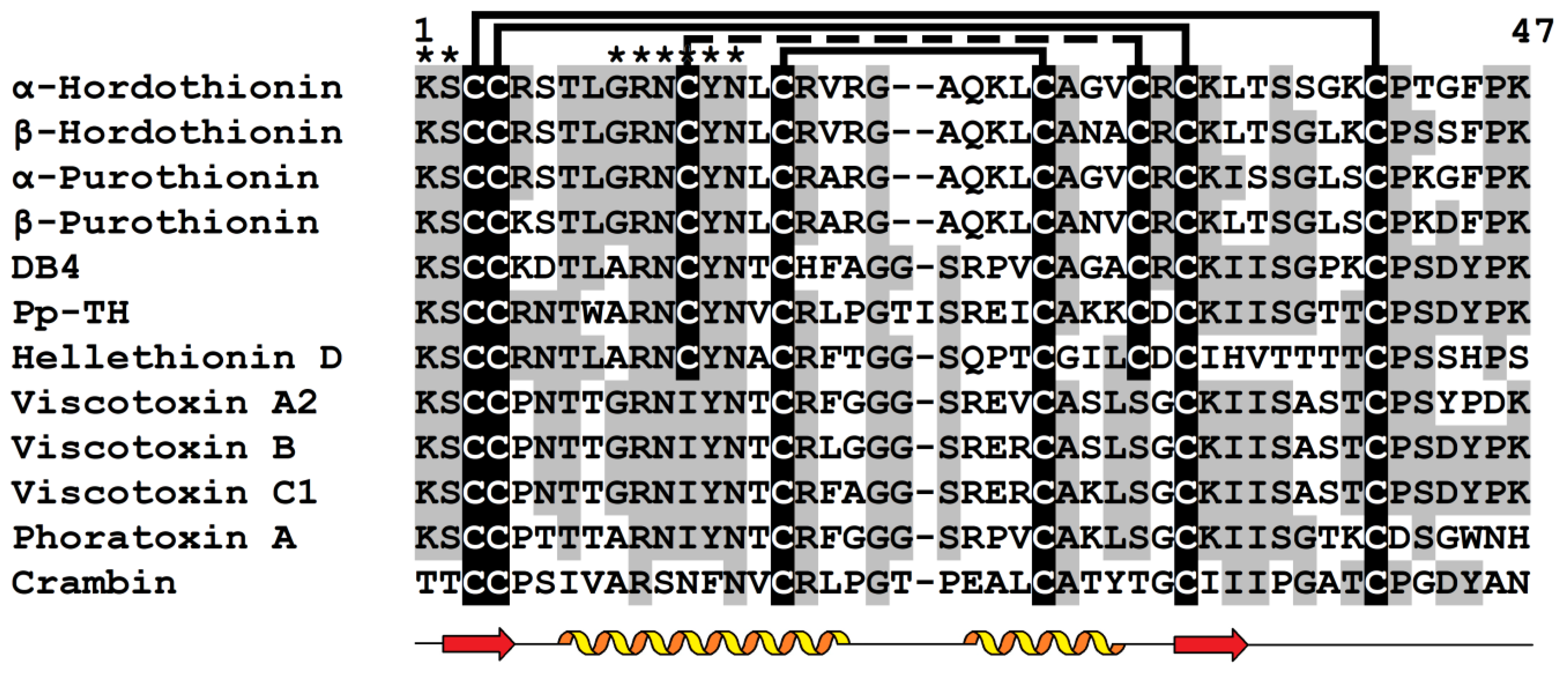
2. Thionins
2.1. General Characteristics
2.2. Biosynthesis
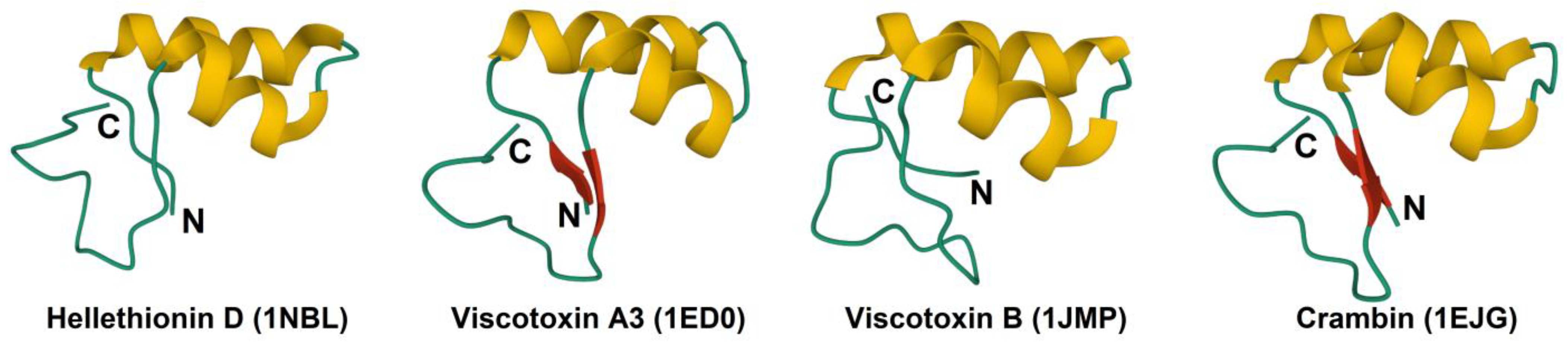
2.3. 3D Structure
2.4. Biological Activity and Phospholipid Binding
2.5. Structure-Function Relationships
2.6. Disulfide Bonds
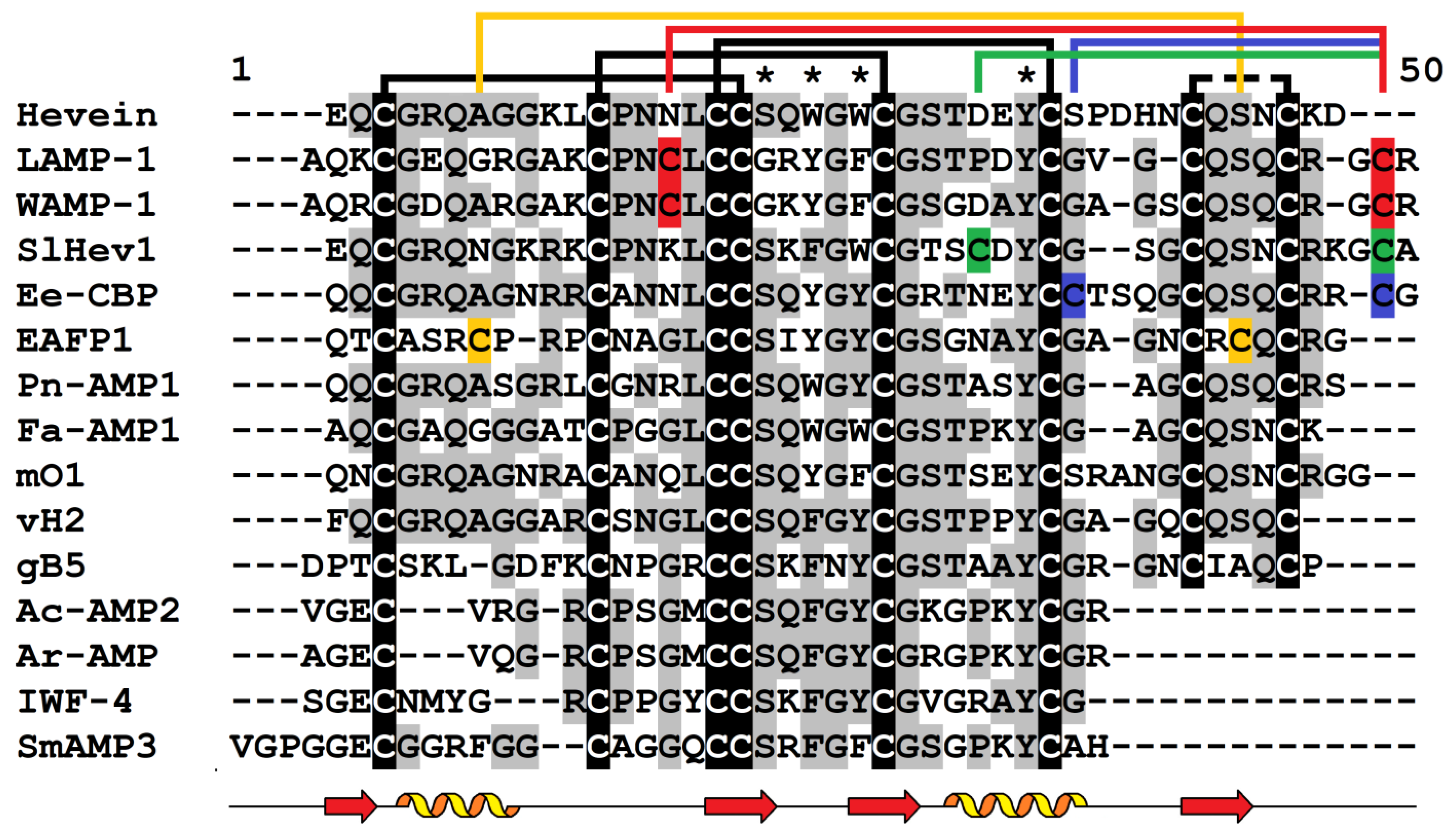
3. Hevein-like Peptides
3.1. General Characteristics
3.2. Biosynthesis
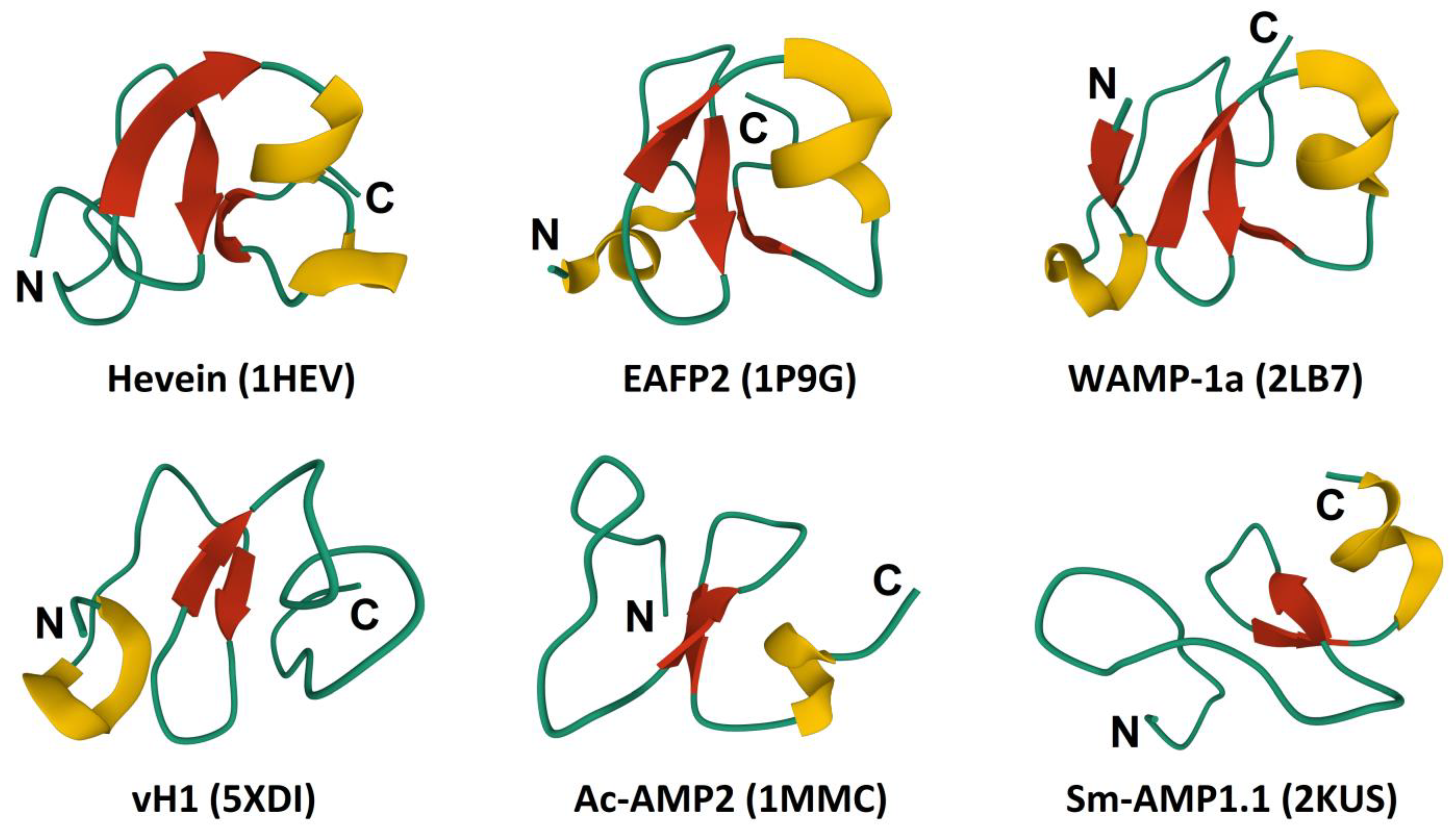
3.3. 3D Structure
3.4. Biological Activity and Chitin-Binding
3.5. Structure-Function Relationships
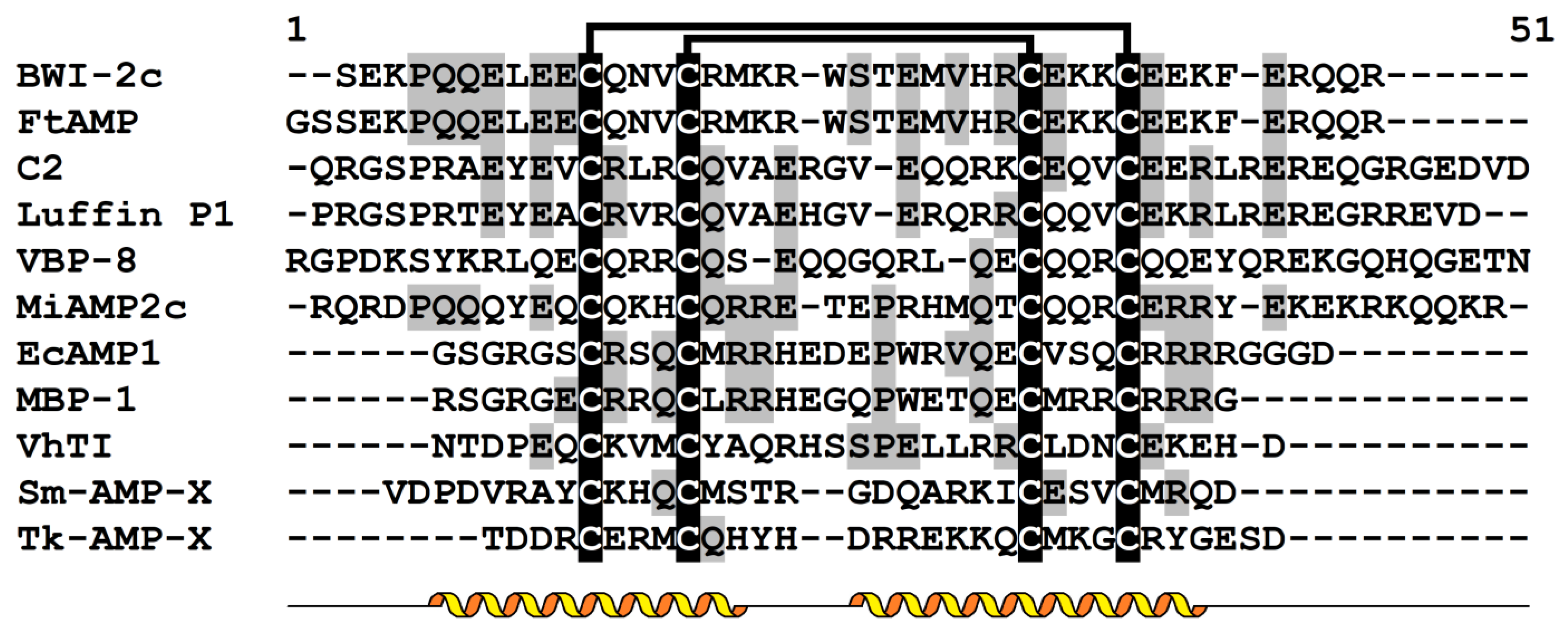
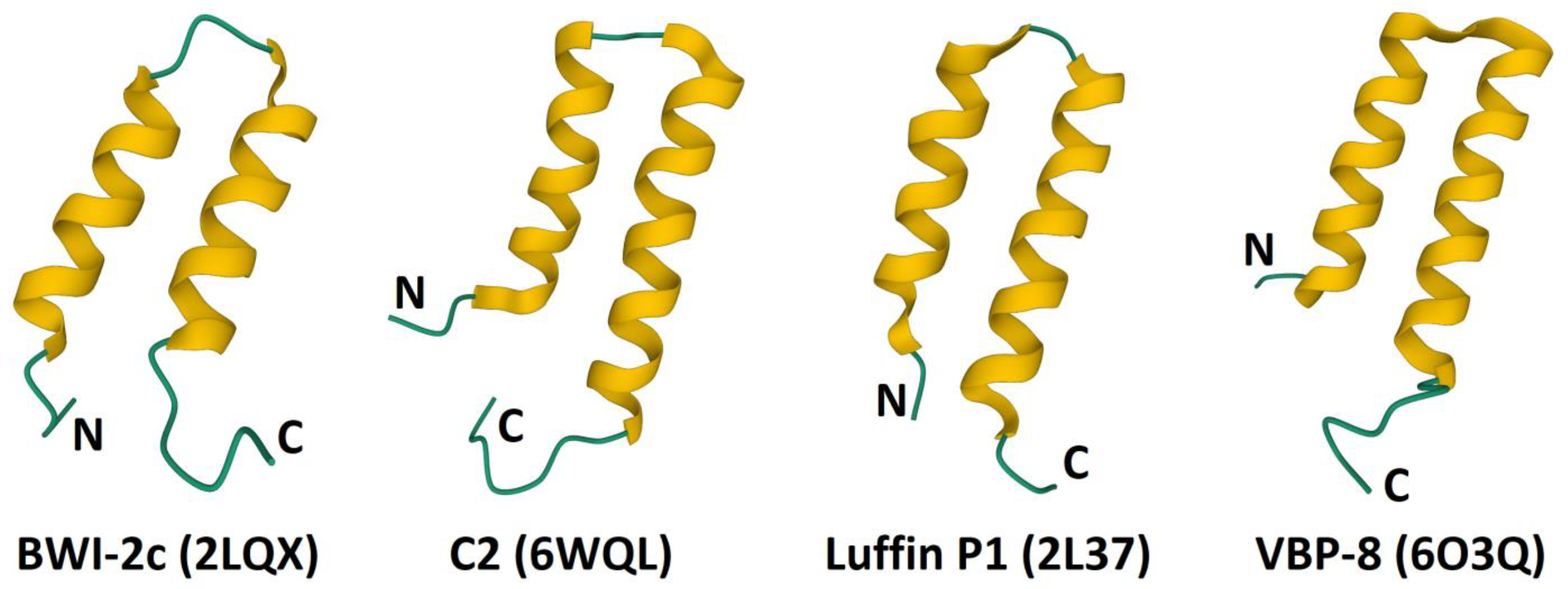
4. α-Hairpinins
4.1. General Characteristics
4.2. Biosynthesis
4.3. Biological Activity
4.4. Structure-Function Relationships
5. Impatiens balsamina Antimicrobial Peptides (Ib-AMPs)
5.1. General Characteristics
5.2. Biosynthesis
5.3. 3D Structure
5.4. Biological Activity
5.5. Structure-Function Relationships
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Bosch, T.C.G.; Zasloff, M. Antimicrobial peptides-or how our ancestors learned to control the microbiome. mBio 2021, 12, e0184721. [Google Scholar] [CrossRef] [PubMed]
- Lazzaro, B.P.; Zasloff, M.; Rolff, J. Antimicrobial peptides: Application informed by evolution. Science 2020, 368, eaau5480. [Google Scholar] [CrossRef] [PubMed]
- Campos, M.L.; de Souza, C.M.; de Oliveira, K.B.S.; Dias, S.C.; Franco, O.L. The role of antimicrobial peptides in plant immunity. J. Exp. Bot. 2018, 69, 4997–5011. [Google Scholar] [CrossRef] [PubMed]
- Tam, J.P.; Wang, S.; Wong, K.H.; Tan, W.L. Antimicrobial peptides from plants. Pharmaceuticals 2015, 8, 711–757. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hu, S.; Jian, W.; Xie, C.; Yang, X. Plant antimicrobial peptides: Structures, functions, and applications. Bot. Stud. 2021, 62, 5. [Google Scholar] [CrossRef] [PubMed]
- Omardien, S.; Brul, S.; Zaat, S.A.J. Activity of cationic antimicrobial peptides against Gram-positives: Current progress made in understanding the mode of action and the response of bacteria. Front. Cell Dev. Biol. 2016, 4, 111. [Google Scholar] [CrossRef]
- Sharma, P.; Kaur, J.; Sharma, G.; Kashyap, P. Plant derived antimicrobial peptides: Mechanism of target, isolation techniques, sources and pharmaceutical applications. J. Food Biochem. 2022, 46, e14348. [Google Scholar] [CrossRef]
- Tailor, R.H.; Acland, D.P.; Attenborough, S.; Cammue, B.P.; Evans, I.J.; Osborn, R.W.; Ray, J.A.; Rees, S.B.; Broekaert, W.F. A novel family of small cysteine-rich antimicrobial peptides from seed of Impatiens balsamina is derived from a single precursor protein. J. Biol. Chem. 1997, 272, 24480–24487. [Google Scholar] [CrossRef]
- Park, C.J.; Park, C.B.; Hong, S.S.; Lee, H.S.; Lee, S.Y.; Kim, S.C. Characterization and cDNA cloning of two glycine- and histidine-rich antimicrobial peptides from the roots of shepherd’s purse, Capsella bursa-pastoris. Plant Mol. Biol. 2000, 44, 187–197. [Google Scholar] [CrossRef]
- León-Buitimea, A.; Garza-Cárdenas, C.R.; Garza-Cervantes, J.A.; Lerma-Escalera, J.A.; Morones-Ramírez, J.R. The demand for new antibiotics: Antimicrobial peptides, nanoparticles, and combinatorial therapies as future strategies in antibacterial agent design. Front. Microbiol. 2020, 11, 1669. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Dou, X.; Song, J.; Lyu, Y.; Zhu, X.; Xu, L.; Li, W.; Shan, A. Antimicrobial peptides: Promising alternatives in the post feeding antibiotic era. Med. Res. Rev. 2019, 39, 831–859. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Dashora, K.; Ameta, K.L.; Singh, N.P.; El-Enshasy, H.A.; Pagano, M.C.; Hesham, A.E.; Sharma, G.D.; Sharma, M.; Bhargava, A. Cysteine-rich antimicrobial peptides from plants: The future of antimicrobial therapy. Phytother. Res. 2021, 35, 256–277. [Google Scholar] [CrossRef]
- Lau, J.L.; Dunn, M.K. Therapeutic peptides: Historical perspectives, current development trends, and future directions. Bioorg. Med. Chem. 2018, 26, 2700–2707. [Google Scholar] [CrossRef]
- Kaspar, A.A.; Reichert, J.M. Future directions for peptide therapeutics development. Drug. Discov. Today 2013, 18, 807–817. [Google Scholar] [CrossRef] [PubMed]
- Yount, N.Y.; Yeaman, M.R. Multidimensional signatures in antimicrobial peptides. Proc. Natl. Acad. Sci. USA 2004, 101, 7363–7368. [Google Scholar] [CrossRef] [PubMed]
- Slezina, M.P.; Istomina, E.A.; Korostyleva, T.V.; Odintsova, T.I. The γ-core motif peptides of plant AMPs as novel antimicrobials for medicine and agriculture. Int. J. Mol. Sci. 2022, 24, 483. [Google Scholar] [CrossRef]
- Stec, B.; Markman, O.; Rao, U.; Heffron, G.; Henderson, S.; Vernon, L.P.; Brumfeld, V.; Teeter, M.M. Proposal for molecular mechanism of thionins deduced from physico-chemical studies of plant toxins. J. Pept. Res. 2004, 64, 210–224. [Google Scholar] [CrossRef]
- Stec, B. Plant thionins—The structural perspective. Cell Mol. Life Sci. 2006, 63, 1370–1385. [Google Scholar] [CrossRef]
- Egorov, T.A.; Odintsova, T.I.; Pukhalsky, V.A.; Grishin, E.V. Diversity of wheat anti-microbial peptides. Peptides 2005, 26, 2064–2073. [Google Scholar] [CrossRef]
- Bohlmann, H.; Apel, K. Isolation and characterization of cDNAs coding leaf-specific thionins closely related to endosperm-specific hordothionin of barley (Hordeum vulgare L.). Mol. Gen. Genet. 1987, 207, 446–454. [Google Scholar] [CrossRef]
- Vernon, L.P. Pyrularia thionin: Physical properties, biological response and comparison to other thionins and cardiotoxin. J. Toxicol. 1992, 11, 169–191. [Google Scholar] [CrossRef]
- Samuelsson, G.; Pettersson, B.M. Separation of viscotoxins from the European mistletoe Viscum album L. (Loranthaceae) by chromatography on sulfoethyl Sephadex. Acta Chem. Scand. 1970, 24, 2751–2756. [Google Scholar] [CrossRef]
- Mellstrand, S.T.; Samuelsson, G. Phoratoxin, a toxic protein from the mistletoe Phoradendron tomentosum subsp. macrophyllum (Loranthaceae). Improvements in the isolation procedure and further studies on the properties. Eur. J. Biochem. 1973, 32, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Vanetten, C.H.; Nielsen, H.C.; Peters, J.E. A crystalline polypeptide from the seed of Crambe abyssinica. Phytochemistry 1965, 4, 467–473. [Google Scholar] [CrossRef]
- Castagnaro, A.; Marana, C.; Carbonero, P.; Garcia-Olmedo, F. cDNA cloning and nucleotide sequences of α1 and α2 thionins from hexaploid wheat endosperm. Plant Physiol. 1994, 106, 1221–1222. [Google Scholar] [CrossRef] [PubMed]
- Odintsova, T.I.; Slezina, M.P.; Istomina, E.A. Plant thionins: Structure, biological functions and potential use in biotechnology. Vavilovskii Zhurnal Genet. Sel. 2018, 22, 667–675. [Google Scholar] [CrossRef]
- Bohlmann, H.; Broekaert, W.F.A. The role of thionins in plant protection. Crit. Rev. Plant Sci. 1994, 13, 1–16. [Google Scholar] [CrossRef]
- Oard, S.; Karki, B.; Enright, F. Is there a difference in metal ion-based inhibition between members of thionin family: Molecular dynamics simulation study. Biophys. Chem. 2007, 130, 65–75. [Google Scholar] [CrossRef]
- Rao, A.G.; Hassan, M.; Hempel, J. Validation of the structure-function properties of α-hordothionin and derivatives through protein modeling. Protein Eng. 1993, 6, 117. [Google Scholar]
- Osório e Castro, V.R.; Vernon, L.P. Stimulation of prothrombinase activity by the nonapeptide Thr-Trp-Ala-Arg-Asn-Ser-Tyr-Asn-Val, a segment of a plant thionin. Peptides 2003, 24, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Wada, K.; Ozaki, Y.; Matsubara, H.; Yoshizumi, H. Studies on purothionin by chemical modifications. J. Biochem. 1982, 91, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Fernandez de Caleya, R.; Gonzalez-Pascual, B.; Garcia-Olmedo, F.; Carbonero, P. Susceptibility of phytopathogenic bacteria to wheat purothionins in vitro. Appl. Microbiol. 1972, 23, 998–1000. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.; Wang, Y.D.; Shaw, K.P.; Vernon, L.P. Cellular responses to Pyrularia thionin are mediated by Ca2+ influx and phospholipase A2 activation and are inhibited by thionin tyrosine iodination. Proc. Natl. Acad. Sci. USA 1989, 86, 5849–5853. [Google Scholar] [CrossRef]
- Vila-Perello, M.; Sanchez-Vallet, A.; Garcia-Olmedo, F.; Molina, A.; Andreu, D. Synthetic and structural studies on Pyrularia pubera thionin: A single-residue mutation enhances activity against Gram-negative bacteria. FEBS Lett. 2003, 536, 215–219. [Google Scholar] [CrossRef]
- Vila-Perello, M.; Sanchez-Vallet, A.; Garcia-Olmedo, F.; Molina, A.; Andreu, D. Structural dissection of a highly knotted peptide reveals minimal motif with antimicrobial activity. J. Biol. Chem. 2005, 280, 1661–1668. [Google Scholar] [CrossRef]
- Vila-Perelló, M.; Tognon, S.; Sánchez-Vallet, A.; García-Olmedo, F.; Molina, A.; Andreu, D. A minimalist design approach to antimicrobial agents based on a thionin template. J. Med. Chem. 2006, 49, 448–451. [Google Scholar] [CrossRef]
- Schaller, G.; Urech, K. Cytotoxicity of different viscotoxins and extracts from the european subspecies of Viscum album L. Phythother. Res. 1996, 10, 473–477. [Google Scholar] [CrossRef]
- Coulon, A.; Mosbah, A.; Lopez, A.; Sautereau, A.M.; Schaller, G.; Urech, K.; Rougé, P.; Darbon, H. Comparative membrane interaction study of viscotoxins A3, A2 and B from mistletoe (Viscum album) and connections with their structures. Biochem. J. 2003, 374, 71–78. [Google Scholar] [CrossRef]
- Fracki, W.S.; Li, D.; Owen, N.; Perry, C.; Naisbitt, G.H.; Vernon, L.P. Role of Tyr and Trp in membrane responses of Pyrularia thionin determined by optical and NMR spectra following Tyr iodination and Trp modification. Toxicon 1992, 30, 1427–1440. [Google Scholar] [CrossRef]
- Guzmán-Rodríguez, J.J.; Ochoa-Zarzosa, A.; López-Gómez, R.; López- Meza, J.E. Plant antimicrobial peptides as potential anticancer agents. Biomed Res. Int. 2015, 2015, 735087. [Google Scholar] [CrossRef] [PubMed]
- Giudici, M.; Poveda, J.A.; Molina, M.L.; de la Canal, L.; González-Ros, J.M.; Pfüller, K.; Pfüller, U.; Villalaín, J. Antifungal effects and mechanism of action of viscotoxin A3. FEBS J. 2006, 273, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Büssing, A.; Stein, G.M.; Wagner, M.; Wagner, B.; Schaller, G.; Pfüller, U.; Schietzel, M. Accidental cell death and generation of reactive oxygen intermediates in human lymphocytes induced by thionins from Viscum album L. Eur. J. Biochem. 1999, 262, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Büssing, A.; Vervecken, W.; Wagner, M.; Wagner, B.; Pfüller, U.; Schietzel, M. Expression of mitochondrial Apo2.7 molecules and caspase-3 activation in human lymphocytes treated with the ribosome-inhibiting mistletoe lectins and the cell membrane permeabilizing viscotoxins. Cytometry 1999, 37, 133–139. [Google Scholar] [CrossRef]
- Coulon, A.; Berkane, E.; Sautereau, A.M.; Urech, K.; Rouge, P.; Lopez, A. Modes of membrane interaction of a natural cysteine-rich peptide: Viscotoxin A3. Biochim. Biophys. Acta 2002, 1559, 145–159. [Google Scholar] [CrossRef]
- Chen, Y.; Guarnieri, M.T.; Vasil, A.I.; Vasil, M.L.; Mant, C.T.; Hodges, R.S. Role of peptide hydrophobicity in the mechanism of action of 𝛼-helical antimicrobial peptides. Antimicrob. Agents Chemother. 2007, 51, 1398–1406. [Google Scholar] [CrossRef]
- Romagnoli, S.; Fogolari, F.; Catalano, M.; Zetta, L.; Schaller, G.; Urech, K.; Giannattasio, M.; Ragona, L.; Molinari, H. NMR solution structure of viscotoxin C1 from Viscum album species Coloratum ohwi: Toward a structure-function analysis of viscotoxins. Biochemistry 2003, 42, 12503–12510. [Google Scholar] [CrossRef]
- Vila-Perello, M.; Andreu, D. Characterization and structural role of disulfide bonds in a highly knotted thionin from Pyrularia pubera. Biopolymers 2005, 80, 697–707. [Google Scholar] [CrossRef]
- Milbradt, A.G.; Kerek, F.; Moroder, L.; Renner, C. Structural characterization of hellethionins from Helleborus purpurascens. Biochemistry 2003, 42, 2404–2411. [Google Scholar] [CrossRef]
- Giudici, M.; Pascual, R.; de la Canal, L.; Pfüller, K.; Pfüller, U.; Villalaín, J. Interaction of viscotoxins A3 and B with membrane model systems: Implications to their mechanism of action. Biophys. J. 2003, 85, 971–981. [Google Scholar] [CrossRef]
- Archer, B.L. The Proteins of Hevea brasiliensis latex. 4. Isolation and characterization of crystalline hevein. Biochem. J. 1960, 75, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Slavokhotova, A.A.; Shelenkov, A.A.; Andreev, Y.A.; Odintsova, T.I. Hevein-like antimicrobial peptides of plants. Biochemistry 2017, 82, 1659–1674. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.H.; Tan, W.L.; Serra, A.; Xiao, T.; Sze, S.K.; Yang, D.; Tam, J.P. Ginkgotides: Proline-rich hevein-like peptides from gymnosperm Ginkgo biloba. Front. Plant Sci. 2016, 7, 1639. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.H.; Tan, W.L.; Kini, S.G.; Xiao, T.; Serra, A.; Sze, S.K.; Tam, J.P. Vaccatides: Antifungal glutamine-rich hevein-like peptides from Vaccaria hispanica. Front. Plant Sci. 2017, 8, 1100. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.K.; Nielsen, J.E.; Madrid, S.M.; Mikkelsen, J.D. Characterization of a new antifungal chitin-binding peptide from sugar beet leaves. Plant Physiol. 1997, 113, 83–91. [Google Scholar] [CrossRef] [PubMed]
- De Bolle, M.F.; Osborn, R.W.; Goderis, I.J.; Noe, L.; Acland, D.; Hart, C.A.; Torrekens, S.; Van Leuven, F.; Broekaert, W.F. Antimicrobial peptides from Mirabilis jalapa and Amaranthus caudatus: Expression, processing, localization and biological activity in transgenic tobacco. Plant Mol. Biol. 1996, 31, 993–1008. [Google Scholar] [CrossRef] [PubMed]
- Andreev, Y.A.; Korostyleva, T.V.; Slavokhotova, A.A.; Rogozhin, E.A.; Utkina, L.L.; Vassilevski, A.A.; Grishin, E.V.; Egorov, T.A.; Odintsova, T.I. Genes encoding hevein-like defense peptides in wheat: Distribution, evolution, and role in stress response. Biochimie 2012, 94, 1009–1016. [Google Scholar] [CrossRef]
- Kini, S.G.; Wong, K.H.; Tan, W.L.; Xiao, T.; Tam, J.P. Morintides: Cargo-free chitin-binding peptides from Moringa oleifera. BMC Plant Biol. 2017, 17, 68. [Google Scholar] [CrossRef]
- Slavokhotova, A.A.; Shelenkov, A.A.; Korostyleva, T.V.; Rogozhin, E.A.; Melnikova, N.V.; Kudryavtseva, A.V.; Odintsova, T.I. Defense peptide repertoire of Stellaria media predicted by high throughput next generation sequencing. Biochimie 2017, 135, 15–27. [Google Scholar] [CrossRef]
- Loo, S.; Tay, S.V.; Kam, A.; Tang, F.; Fan, J.S.; Yang, D.; Tam, J.P. Anti-fungal hevein-like peptides biosynthesized from quinoa cleavable hololectins. Molecules 2021, 26, 5909. [Google Scholar] [CrossRef]
- Loo, S.; Tay, S.V.; Kam, A.; Lee, W.; Tam, J.P. Hololectin interdomain linker determines asparaginyl endopeptidase-mediated maturation of antifungal hevein-like peptides in oats. Front. Plant Sci. 2022, 13, 899740. [Google Scholar] [CrossRef] [PubMed]
- Van den Bergh, K.P.; Rougé, P.; Proost, P.; Coosemans, J.; Krouglova, T.; Engelborghs, Y.; Peumans, W.J.; Van Damme, E.J. Synergistic antifungal activity of two chitin-binding proteins from spindle tree (Euonymus europaeus L.). Planta 2004, 219, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Beintema, J.J. Structural features of plant chitinases and chitin-binding proteins. FEBS Lett. 1994, 350, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, S.; Iida, Y.; Kawasaki, Y.; Minami, Y.; Watanabe, K.; Yagi, F. The chitin-binding capability of Cy-AMP1 from cycad is essential to antifungal activity. J. Pept. Sci. 2009, 15, 492–497. [Google Scholar] [CrossRef]
- Koo, J.C.; Lee, B.; Young, M.E.; Koo, S.C.; Cooper, J.A.; Baek, D.; Lim, C.O.; Lee, S.Y.; Yun, D.J.; Cho, M.J. Pn-AMP1, a plant defense protein, induces actin depolarization in yeasts. Plant Cell Physiol. 2004, 45, 1669–1680. [Google Scholar] [CrossRef]
- Asensio, J.L.; Canada, F.J.; Bruix, M.; Rodriguez-Romero, A.; Jimenez-Barbero, J. The interaction of hevein with N-acetylglucosamine-containing oligosaccharides. Solution structure of hevein complexed to chitobiose. Eur. J. Biochem. 1995, 230, 621–633. [Google Scholar] [CrossRef] [PubMed]
- Asensio, J.L.; Siebert, H.C.; von Der Lieth, C.W.; Laynez, J.; Bruix, M.; Soedjanaamadja, U.M.; Beintema, J.J.; Canada, F.J.; Gabius, H.J.; Jimenez-Barbero, J. NMR investigations of protein-carbohydrate interactions: Studies on the relevance of Trp/Tyr variations in lectin binding sites as deduced from titration microcalorimetry and NMR studies on hevein domains. Determination of the NMR structure of the complex between pseudohevein and N,N’,N”-triacetylchitotriose. Proteins 2000, 40, 218–236. [Google Scholar]
- Muraki, M. The importance of Ch/π interactions to the function of carbohydrate binding proteins. Protein Pept. Lett. 2002, 9, 195–209. [Google Scholar] [CrossRef]
- Aboitiz, N.; Vila-Perelló, M.; Groves, P.; Asensio, J.L.; Andreu, D.; Cañada, F.J.; Jiménez-Barbero, J. NMR and modeling studies of protein-carbohydrate interactions: Synthesis, three-dimensional structure, and recognition properties of a minimum hevein domain with binding affinity for chitooligosaccharides. Chembiochem 2004, 5, 1245–1255. [Google Scholar] [CrossRef]
- Chávez, M.I.; Vila-Perelló, M.; Cañada, F.J.; Andreu, D.; Jiménez-Barbero, J. Effect of a serine-to-aspartate replacement on the recognition of chitin oligosaccharides by truncated hevein. A 3D view by using NMR. Carbohydr. Res. 2010, 345, 1461–1468. [Google Scholar] [CrossRef]
- Muraki, M.; Morii, H.; Harata, K. Chemically prepared hevein domains: Effect of C-terminal truncation and the mutagenesis of aromatic residues on the affinity for chitin. Protein Eng. 2000, 13, 385–389. [Google Scholar] [CrossRef] [PubMed]
- Martins, J.C.; Maes, D.; Loris, R.; Pepermans, H.A.; Wyns, L.; Willem, R.; Verheyden, P. 1H NMR study of the solution structure of Ac-AMP2, a sugar binding antimicrobial protein isolated from Amaranthus caudatus. J. Mol. Biol. 1996, 258, 322–333. [Google Scholar] [CrossRef] [PubMed]
- Van Parijs, J.; Broekart, W.F.; Goldstein, I.J.; Peumans, W.J. Hevein: An antifungal protein from rubber-tree (Hevea brasiliensis) latex. Planta 1991, 183, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Utkina, L.L.; Zhabon, E.O.; Slavokhotova, A.A.; Rogozhin, E.A.; Shiyan, A.N.; Grishin, E.V.; Egorov, T.A.; Odintsova, T.I.; Pukhal’skiy, V.A. Heterologous expression of a synthetic gene encoding a novel hevein-type antimicrobial peptide of Leymus arenarius in Escherichia coli cells. Russ. J. Genet. 2010, 46, 1449–1454. [Google Scholar] [CrossRef]
- Odintsova, T.I.; Vassilevski, A.A.; Slavokhotova, A.A.; Musolyamov, A.K.; Finkina, E.I.; Khadeeva, N.V.; Rogozhin, E.A.; Korostyleva, T.V.; Pukhalsky, V.A.; Grishin, E.V.; et al. A novel antifungal hevein-type peptide from Triticum kiharae seeds with a unique 10-cysteine motif. FEBS J. 2009, 276, 4266–4275. [Google Scholar] [CrossRef]
- Slezina, M.P.; Korostyleva, T.V.; Slavokhotova, A.A.; Istomina, E.A.; Shcherbakova, L.A.; Pukhalskij, V.A.; Odintsova, T.I. Genes encoding hevein-like antimicrobial peptides from Elytrigia repens (L.) Desv. ex Nevski. Russ. J. Genet. 2018, 54, 1152–1159. [Google Scholar] [CrossRef]
- Slavokhotova, A.A.; Naumann, T.A.; Price, N.P.; Rogozhin, E.A.; Andreev, Y.A.; Vassilevski, A.A.; Odintsova, T.I. Novel mode of action of plant defense peptides − hevein-like antimicrobial peptides from wheat inhibit fungal metalloproteases. FEBS J. 2014, 281, 4754–4764. [Google Scholar] [CrossRef]
- Odintsova, T.; Shcherbakova, L.; Slezina, M.; Pasechnik, T.; Kartabaeva, B.; Istomina, E.; Dzhavakhiya, V. Hevein-like antimicrobial peptides WAMPs: Structure–function relationship in antifungal activity and sensitization of plant pathogenic fungi to tebuconazole by WAMP-2-derived peptides. Int. J. Mol. Sci. 2020, 21, 7912. [Google Scholar] [CrossRef]
- Van den Bergh, K.P.; Proost, P.; Van Damme, J.; Coosemans, J.; Van Damme, E.J.; Peumans, W.J. Five disulfide bridges stabilize a hevein-type antimicrobial peptide from the bark of spindle tree (Euonymus europaeus L.). FEBS Lett. 2002, 530, 181–185. [Google Scholar] [CrossRef]
- Huang, R.H.; Xiang, Y.; Liu, X.Z.; Zhang, Y.; Hu, Z.; Wang, D.C. Two novel antifungal peptides distinct with a five-disulfide motif from the bark of Eucommia ulmoides Oliv. FEBS Lett. 2002, 521, 87–90. [Google Scholar] [CrossRef]
- Koo, J.C.; Lee, S.Y.; Chun, H.J.; Cheong, Y.H.; Choi, J.S.; Kawabata, S.; Miyagi, M.; Tsunasawa, S.; Ha, K.S.; Bae, D.W.; et al. Two hevein homologs isolated from the seed of Pharbitis nil L. exhibit potent anti-fungal activity. Biochim. Biophys. Acta 1998, 1382, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Fujimura, M.; Minami, Y.; Watanabe, K.; Tadera, K. Purification, characterization, and sequencing of a novel type of antimicrobial peptides, Fa-AMP1 and Fa-AMP2, from seeds of buckwheat (Fagopyrum esculentum Moench.). Biosci. Biotechnol. Biochem. 2003, 67, 1636–1642. [Google Scholar] [CrossRef] [PubMed]
- Broekaert, W.F.; Mariën, W.; Terras, F.R.; De Bolle, M.F.; Proost, P.; Van Damme, J.; Dillen, L.; Claeys, M.; Rees, S.B.; Vanderleyden, J.; et al. Antimicrobial peptides from Amaranthus caudatus seeds with sequence homology to the cysteine/glycine-rich domain of chitin-binding proteins. Biochemistry 1992, 31, 4308–4314. [Google Scholar] [CrossRef] [PubMed]
- Lipkin, A.; Anisimova, V.; Nikonorova, A.; Babakov, A.; Krause, E.; Bienert, M.; Grishin, E.; Egorov, T. An antimicrobial peptide Ar-AMP from amaranth (Amaranthus retroflexus L.) seeds. Phytochemistry 2005, 66, 2426–2431. [Google Scholar] [CrossRef]
- Rogozhin, E.A.; Slezina, M.P.; Slavokhotova, A.A.; Istomina, E.A.; Korostyleva, T.V.; Smirnov, A.N.; Grishin, E.V.; Egorov, T.A.; Odintsova, T.I. A novel antifungal peptide from leaves of the weed Stellaria media L. Biochimie 2015, 116, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Huang, R.H.; Liu, X.Z.; Zhang, Y.; Wang, D.C. Crystal structure of a novel antifungal protein distinct with five disulfide bridges from Eucommia ulmoides Oliver at an atomic resolution. J. Struct. Biol. 2004, 148, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.H.; Xiang, Y.; Tu, G.Z.; Zhang, Y.; Wang, D.C. Solution structure of Eucommia antifungal peptide: A novel structural model distinct with a five-disulfide motif. Biochemistry 2004, 43, 6005–6012. [Google Scholar] [CrossRef]
- Dubovskii, P.V.; Vassilevski, A.A.; Slavokhotova, A.A.; Odintsova, T.I.; Grishin, E.V.; Egorov, T.A.; Arseniev, A.S. Solution structure of a defense peptide from wheat with a 10-cysteine motif. Biochem. Biophys. Res. Commun. 2011, 411, 14–18. [Google Scholar] [CrossRef]
- Istomina, E.A.; Slavokhotova, A.A.; Korostyleva, T.V.; Semina, Y.V.; Shcherbakova, L.A.; Pukhalskij, V.A.; Odintsova, T.I. Genes encoding hevein-like antimicrobial peptides WAMPs in the species of the genus Aegilops L. Russ. J. Genet. 2017, 53, 1320–1327. [Google Scholar] [CrossRef]
- Conners, R.; Konarev, A.V.; Forsyth, J.; Lovegrove, A.; Marsh, J.; Joseph-Horne, T.; Shewry, P.; Brady, R.L. An unusual helix-turn-helix protease inhibitory motif in a novel trypsin inhibitor from seeds of veronica (Veronica hederifolia L.). J. Biol. Chem. 2007, 282, 27760–27768. [Google Scholar] [CrossRef]
- Nolde, S.B.; Vassilevski, A.A.; Rogozhin, E.A.; Barinov, N.A.; Balashova, T.A.; Samsonova, O.V.; Baranov, Y.V.; Feofanov, A.V.; Egorov, T.A.; Arseniev, A.S.; et al. Disulfide-stabilized helical hairpin structure and activity of a novel antifungal peptide EcAMP1 from seeds of barnyard grass (Echinochloa crus-galli). J. Biol. Chem. 2011, 286, 25145–25153. [Google Scholar] [CrossRef]
- Ng, Y.M.; Yang, Y.; Sze, K.H.; Zhang, X.; Zheng, Y.T.; Shaw, P.C. Structural characterization and anti-HIV-1 activities of arginine/glutamate-rich polypeptide Luffin P1 from the seeds of sponge gourd (Luffa cylindrica). J. Struct. Biol. 2011, 174, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Oparin, P.B.; Mineev, K.S.; Dunaevsky, Y.E.; Arseniev, A.S.; Belozersky, M.A.; Grishin, E.V.; Egorov, T.A.; Vassilevski, A.A. Buckwheat trypsin inhibitor with helical hairpin structure belongs to a new family of plant defense peptides. Biochem. J. 2012, 446, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Du, J.; Li, J.; Wang, Z. Inhibitory site of α-hairpinin peptide from tartary buckwheat has no effect on its antimicrobial activities. Acta Biochim. Biophys. Sin. 2018, 50, 408–416. [Google Scholar] [CrossRef]
- Marcus, J.P.; Green, J.L.; Goulter, K.C.; Manners, J.M. A family of antimicrobial peptides is produced by processing of a 7S globulin protein in Macadamia integrifolia kernels. Plant J. 1999, 19, 699–710. [Google Scholar] [CrossRef]
- Utkina, L.L.; Andreev, Y.A.; Rogozhin, E.A.; Korostyleva, T.V.; Slavokhotova, A.A.; Oparin, P.B.; Vassilevski, A.A.; Grishin, E.V.; Egorov, T.A.; Odintsova, T.I. Genes encoding 4-Cys antimicrobial peptides in wheat Triticum kiharae Dorof. et Migush.: Multimodular structural organization, instraspecific variability, distribution and role in defence. FEBS J. 2013, 280, 3594–3608. [Google Scholar] [CrossRef]
- Berkut, A.A.; Usmanova, D.R.; Peigneur, S.; Oparin, P.B.; Mineev, K.S.; Odintsova, T.I.; Tytgat, J.; Arseniev, A.S.; Grishin, E.V.; Vassilevski, A.A. Structural similarity between defense peptide from wheat and scorpion neurotoxin permits rational functional design. J. Biol. Chem. 2014, 289, 14331–14340. [Google Scholar] [CrossRef]
- Marcus, J.P.; Goulter, K.C.; Manners, J.M. Peptide fragments from plant vicilins expressed in Escherichia coli exhibit antimicrobial activity in vitro. Plant Mol. Biol. Rep. 2008, 26, 75–87. [Google Scholar] [CrossRef]
- Slavokhotova, A.A.; Rogozhin, E.A.; Musolyamov, A.K.; Andreev, Y.A.; Oparin, P.B.; Berkut, A.A.; Vassilevski, A.A.; Egorov, T.A.; Grishin, E.V.; Odintsova, T.I. Novel antifungal alpha-hairpinin peptide from Stellaria media seeds: Structure, biosynthesis, gene structure and evolution. Plant Mol. Biol. 2014, 84, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Sousa, D.A.; Porto, W.F.; Silva, M.Z.; Da Silva, T.R.; Franco, O.L. Influence of cysteine and tryptophan substitution on DNA-binding activity on maize α-hairpinin antimicrobial peptide. Molecules 2016, 21, 1062. [Google Scholar] [CrossRef]
- Duvick, J.P.; Rood, T.; Rao, A.G.; Marshak, D.R. Purification and characterization of a novel antimicrobial peptide from maize (Zea mays L.) kernels. J. Biol. Chem. 1992, 267, 18814–18820. [Google Scholar] [CrossRef] [PubMed]
- Rogozhin, E.; Zalevsky, A.; Mikov, A.; Smirnov, A.; Egorov, T. Characterization of hydroxyproline-containing hairpin-like antimicrobial peptide EcAMP1-Hyp from barnyard grass (Echinochloa crusgalli L.) seeds: Structural identification and comparative analysis of antifungal activity. Int. J. Mol. Sci. 2018, 19, 3449. [Google Scholar] [CrossRef] [PubMed]
- Barashkova, A.S.; Ryazantsev, D.Y.; Rogozhin, E.A. Rational design of plant hairpin-like peptide EcAMP1: Structural–functional correlations to reveal antibacterial and antifungal activity. Molecules 2022, 27, 3554. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, K.N.; Sivaraja, V.; Huys, I.; Sasaki, T.; Cheng, B.; Kumar, T.K.; Sato, K.; Tytgat, J.; Yu, C.; San, B.C.; et al. κ-Hefutoxin1, a novel toxin from the scorpion Heterometrus fulvipes with unique structure and function. Importance of the functional diad in potassium channel selectivity. J. Biol. Chem. 2002, 277, 30040–30047. [Google Scholar] [CrossRef]
- Patel, S.U.; Osborn, R.; Rees, S.; Thornton, J.M. Structural studies of Impatiens balsamina antimicrobial protein (Ib-AMP1). Biochemistry 1998, 37, 983–990. [Google Scholar] [CrossRef]
- Lee, D.G.; Shin, S.Y.; Kim, D.-H.; Seo, M.Y.; Kang, J.H.; Lee, Y.; Kim, K.L.; Hahm, K.S. Antifungal mechanism of a cysteine-rich antimicrobial peptide, Ib-AMP1, from Impatiens balsamina against Candida albicans. Biotechnol. Lett. 1999, 21, 1047–1050. [Google Scholar] [CrossRef]
- Wang, P.; Bang, J.K.; Kim, H.J.; Kim, J.K.; Kim, Y.; Shin, S.Y. Antimicrobial specificity and mechanism of action of disulfide-removed linear analogs of the plant-derived Cys-rich antimicrobial peptide Ib-AMP1. Peptides 2009, 30, 2144–2149. [Google Scholar] [CrossRef]
- Fan, X.; Schäfer, H.; Reichling, J.; Wink, M. Bactericidal properties of the antimicrobial peptide Ib-AMP4 from Impatiens balsamina produced as a recombinant fusion-protein in Escherichia coli. Biotechnol. J. 2013, 8, 1213–1220. [Google Scholar] [CrossRef]
- Fan, X.; Reichling, J.; Wink, M. Antibacterial activity of the recombinant antimicrobial peptide Ib-AMP4 from Impatiens balsamina and its synergy with other antimicrobial agents against drug resistant bacteria. Pharmazie 2013, 68, 628–630. [Google Scholar]
- Wu, W.H.; Di, R.; Matthews, K.R. Antibacterial mode of action of Ib-AMP1 against Escherichia coli O157:H7. Probiotics Antimicrob. Proteins 2013, 5, 131–141. [Google Scholar] [CrossRef]
- Thevissen, K.; François, I.E.; Sijtsma, L.; van Amerongen, A.; Schaaper, W.M.; Meloen, R.; Posthuma-Trumpie, T.; Broekaert, W.F.; Cammue, B.P. Antifungal activity of synthetic peptides derived from Impatiens balsamina antimicrobial peptides Ib-AMP1 and Ib-AMP4. Peptides 2005, 26, 1113–1119. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Amino Acid Sequence | Net Charge at pH 7.0 |
|---|---|---|
| Antimicrobial Activity and Toxicity | Reference | |
| α-Purothionin | KSCCRSTLGRNCYNLCRARG--AQKLCAGVCRCKISSGLSCPKGFPK | +10 |
| MIC (minimal inhibition concentration) = 3 μg/mL and MBC (minimal bactericidal concentration) = 6 μg/mL against Pseudomonas solanacearum; MIC = 6 μg/mL and MBC = 12 μg/mL against Xanthomonas phaseoli. | [33] | |
| β-Purothionin | KSCCKSTLGRNCYNLCRARG--AQKLCANVCRCKLTSGLSCPKDFPK | +9 |
| Complete growth suppression of yeasts at a concentration of 4.7 μg/mL. Lethal to mice within 40 min when administered at 0.1 mg. | [32] | |
| MIC = 1.5 μg/mL and MBC = 1.5 μg/mL against P. solanacearum; MIC = 12 μg/mL and MBC = 25 μg/mL against X. phaseoli. | [33] | |
| Acetyl derivative of β-purothionin | KSCCKSTLGRNCYNLCRARG--AQKLCANVCRCKLTSGLSCPKDFPK | nd |
| No toxity towards yeast cells at a concentration of 11.0 μg/mL. | [32] | |
| Succinyl derivative of β-purothionin | KSCCKSTLGRNCYNLCRARG--AQKLCANVCRCKLTSGLSCPKDFPK | nd |
| No toxity towards yeast cells at a concentration of 10.0 μg/mL. | [32] | |
| Iodinated derivative of β-purothionin | KSCCKSTLGRNCYNLCRARG--AQKLCANVCRCKLTSGLSCPKDFPK | nd |
| No toxity towards yeast cells at a concentration of 10.0 μg/mL. Scarcely lethal to mice when administered at 10.0 mg. | [32] | |
| Nitrated derivative of β-purothionin | KSCCKSTLGRNCYNLCRARG--AQKLCANVCRCKLTSGLSCPKDFPK | nd |
| Weakly toxic toward yeast cells at a concentration of 12.6 μg/mL. Toward mice, the efficiency is less than 30% compared to that of native purothinin. | [32] | |
| Pp-TH | KSCCRNTWARNCYNVCRLPGTISREICAKKCDCKIISGTTCPSDYPK | +6 |
| For natural Pp-TH: | ||
| 50% hemolysis at 20 μg/mL. IC50 (concentration required for 50% growth inhibition) = 5.0 μg/mL against mouse B16 melanoma cells; IC50 = 14 μg/mL against HeLa cells; 100% lethal to mice within 4.5 min when administered at 100 μg. | [34] | |
| EC50 (Effective concentration for 50% inhibition) > 20 μM (>20 μM) against Rhizobium meliloti; EC50 = 3.1 μM (3.67 μM) against Xanthamonas campestris pv. campestris; EC50 = 1.7 μM against X. campestris pv. translucens; EC50 = 0.48 (0.23 μM) μM against Clavibacter michiganensis subsp. sepedonicus C5; EC50 = 0.43 μM (0.38 μM) against Fusarium oxysporum f. sp. conglutinans; EC50 = 0.7 μM against Plectosphaerella cucumerina; EC50 = 2.2 μM against Botrytis cinerea; | [35], the figures in parentheses [36] | |
| For synthetic Pp-TH: EC50 > 20 μM against R. meliloti; EC50 = 2.9 μM against X. campestris pv. campestris; EC50 = 1.4 μM against X. campestris pv. translucens; EC50 = 0.48 μM against C. michiganensis; EC50 = 0.33 μM against F. oxysporum f. sp. conglutinans; EC50 = 0.8 μM against P. cucumerina; | [35] | |
| EC50 > 20 μM against R. meliloti; EC50 = 3.65 μM against X. campestris; EC50 = 0.30 μM against C. michiganensis; EC50 = 0.36 μM against P. cucumerina; EC50 = 0.32 μM against B. cinerea. | [37] | |
| Iodinated derivative of Pp-TH | KSCCRNTWARNCYNVCRLPGTISREICAKKCDCKIISGTTCPSDYPK | nd |
| 0% hemolysis at 20 μg/mL. No activity against mouse B16 melanoma and HeLa cells at 100 μg/mL; Non-lethal to mice when administered at 100 μg. | [34] | |
| Pp-TH(D32R) or PpTHR or TH32R | KSCCRNTWARNCYNVCRLPGTISREICAKKCRCKIISGTTCPSDYPK | +8 |
| EC50 = 3.3 μM (3.8 μM) against R. meliloti; EC50 = 1.2 μM (0.38 μM) against X. campestris pv. campestris; EC50 = 0.8 μM against X. campestris pv. translucens; EC50 = 0.38 μM (0.23 μM) against C. michiganensis subsp. sepedonicus C5; EC50 = 0.73 μM (1.52 μM) against F. oxysporum f. sp. conglutinans; EC50 = 1.25 μM against P. cucumerina; EC50 = 2.5 μM against B. cinerea; | [35], the figures in parentheses [36] | |
| EC50 = 0.8 μM against R. meliloti; EC50 = 0.3 μM against X. campestris; EC50 = 0.37 μM against C. michiganensis; EC50 = 0.36 μM against P. cucumerina; EC50 = 0.80 μM against B. cinerea. | [37] | |
| PpTH(3-41) | --CCRNTWARNCYNVCRLPGTISREICAKKCDCKIISGTTC | +5 |
| EC50 > 20 μM against R. meliloti; EC50 = 7.5 μM against X. campestris pv. campestris; EC50 = 0.18 μM against C. michiganensis subsp. sepedonicus C5; EC50 = 1.73 μM against F. oxysporum f. sp. conglutinans. | [36] | |
| PpTHR(3-41) | --CCRNTWARNCYNVCRLPGTISREICAKKCRCKIISGTTC | +7 |
| EC50 = 5.3 μM against R. meliloti; EC50 = 3.27 μM against X. campestris pv. campestris; EC50 = 0.21 μM against C. michiganensis subsp. sepedonicus C5; EC50 = 2.5 μM against F. oxysporum f. sp. conglutinans. | [36] | |
| PpTH(7-32) | ------TWARNCYNVCRLPGTISREICAKKCD | +3 |
| EC50 > 20 μM against R. meliloti; EC50 > 20 μM against X. campestris pv. campestris; EC50 = 0.63 μM against C. michiganensis subsp. sepedonicus C5; EC50 = 1.8 μM against F. oxysporum f. sp. conglutinans. | [36] | |
| PpTH(7-32)b (disulfide bridges C1-C2 and C3-C4) | ------TWARNCYNVCRLPGTISREICAKKCD | +3 |
| EC50 > 20 μM against R. meliloti; EC50 > 20 μM against X. campestris pv. campestris; EC50 = 1.8 μM against C. michiganensis subsp. sepedonicus C5; EC50 > 20 μM against F. oxysporum f. sp. conglutinans. | [36] | |
| PpTHR(7-32) or TH(7-32R) | ------TWARNCYNVCRLPGTISREICAKKCR | +5 |
| EC50 = 4 μM against R. meliloti; EC50 = 7.2 μM against X. campestris pv. campestris; EC50 = 1.58 μM against C. michiganensis subsp. sepedonicus C5; EC50 = 0.5 μM against F. oxysporum f. sp. conglutinans; | [36] | |
| EC50 = 4 μM against R. meliloti; EC50 = 4.6 μM against X. campestris; EC50 = 0.80 μM against C. michiganensis; EC50 = 7.5 μM against P. cucumerina; EC50 = 0.80 μM against B. cinerea. | [37] | |
| PpTH(24-32) | -----------------------REICAKKCD | +1 |
| EC50 > 20 μM against R. meliloti; EC50 > 20 μM against X. campestris pv. campestris; EC50 > 20 μM against C. michiganensis subsp. sepedonicus C5; EC50 > 20 μM against F. oxysporum f. sp. conglutinans. | [36] | |
| PpTHR(24-32) or TH(24-32R) | -----------------------REICAKKCR | +3 |
| EC50 > 20 μM against R. meliloti; EC50 > 20 μM against X. campestris pv. campestris; EC50 = 16 μM against C. michiganensis subsp. sepedonicus C5; EC50 > 20 μM against F. oxysporum f. sp. conglutinans; | [36] | |
| EC50 > 50 μM against R. meliloti; EC50 > 50 μM against X. campestris; EC50 = 21 μM against C. michiganensis; EC50 = 20 μM against P. cucumerina; EC50 = 3.50 μM against B. cinerea. | [37] | |
| TH(24-32R)Abu | -----------------------REIBAKKBR | nd |
| EC50 > 50 μM against R. meliloti; EC50 > 50 μM against X. campestris; EC50 = 5.13 μM against C. michiganensis; EC50 = 15.67 μM against P. cucumerina; EC50 = 4.10 μM against B. cinerea. | [37] | |
| TH(24-32R)cC | -----------------------REIcAKKCR | nd |
| EC50 > 50 μM against R. meliloti; EC50 > 50 μM against X. campestris; EC50 = 0.90 μM against C. michiganensis; EC50 = 0.80 μM against P. cucumerina; EC50 = 0.55 μM against B. cinerea. | [37] | |
| TH(24-32R)cH | -----------------------REIcAKKXR | nd |
| EC50 > 50 μM against R. meliloti; EC50 > 50 μM against X. campestris; EC50 = 0.90 μM against C. michiganensis; EC50 = 0.78 μM against P. cucumerina; EC50 = 0.40 μM against B. cinerea. | [37] | |
| PpTH(7-19) | ------TWARNCYNVCRLP | +2 |
| EC50 > 20 μM against R. meliloti; EC50 > 20 μM against X. campestris pv. campestris; EC50 = 15 μM against C. michiganensis subsp. sepedonicus C5; EC50 > 20 μM against F. oxysporum f. sp. conglutinans; | [36] | |
| EC50 > 50 μM against R. meliloti; EC50 > 50 μM against X. campestris; EC50 > 50 μM against C. michiganensis; EC50 = 29 μM against P. cucumerina; EC50 = 5.50 μM against B. cinerea. | [37] | |
| PpTH(7-19)Abu | ------TWARNBYNVBRLP | nd |
| EC50 > 50 μM against R. meliloti; EC50 > 50 μM against X. campestris; EC50 = 3.27 μM against C. michiganensis; EC50 = 3.97 μM against P. cucumerina; EC50 = 1.60 μM against B. cinerea. | [37] | |
| PpTH(7-19)cC | ------TWARNcYNVCRLP | nd |
| EC50 > 50 μM against R. meliloti; EC50 = 29.33 μM against X. campestris; EC50 = 0.51 μM against C. michiganensis; EC50 = 2.77 μM against P. cucumerina; EC50 = 0.20 μM against B. cinerea. | [37] | |
| PpTH(7-19)cH | ------TWARNcYNVXRLP | nd |
| EC50 > 50 μM against R. meliloti; EC50 = 18.33 μM against X. campestris; EC50 = 0.74 μM against C. michiganensis; EC50 = 3.50 μM against P. cucumerina; EC50 = 0.19 μM against B. cinerea. | [37] | |
| PpTH(15−28) | --------------VCRLPGTISREICA | +1 |
| EC50 > 20 μM against R. meliloti; EC50 > 20 μM against X. campestris pv. campestris; EC50 > 20 μM against C. michiganensis subsp. sepedonicus C5; EC50 > 20 μM against F. oxysporum f. sp. conglutinans. | [36] | |
| TH(7-19)(24-32R) | ------TWARNCYNVCRLP-+ -REICAKKCR | +2 and +3 |
| EC50 = 2.07 μM against R. meliloti; EC50 = 1.03 μM against X. campestris; EC50 = 0.03 μM against C. michiganensis; EC50 = 0.16 μM against P. cucumerina; EC50 = 0.08 μM against B. cinerea. | [37] | |
| Viscotoxin A3 | KSCCPNTTGRNIYNACRLTGA-PRPTCAKLSGCKIISGSTCPSDYPK | +6 |
| ED50 (concentration of substance inhibiting 3H-thymidine incorporation 50%) = 0.31 μg/mL against Yoshida sarcoma cells; | [38] | |
| Can penetrate into the model monolayer membrane (critical surface pressure πc ≤ 39.6 μN/m). | [39] | |
| Viscotoxin A2 | KSCCPNTTGRNIYNTCRFGGG-SREVCASLSGCKIISASTCPSYPDK | +4 |
| ED50 = 1.06 μg/mL against Yoshida sarcoma cells; | [38] | |
| Can penetrate into the model monolayer membrane (critical surface pressure πc ≤ 32.3 μN/m). | [39] | |
| Viscotoxin A1 | KSCCPSTTGRNIYNTCRLTGS-SRETCAKLSGCKIISASTCPSNYPK | +6 |
| ED50 = 0.87 μg/mL against Yoshida cells. | [38] | |
| Viscotoxin B | KSCCPNTTGRNIYNTCRLGGG-SRERCASLSGCKIISASTCPSDYPK | +5 |
| ED50 = 4.58 μg/mL against Yoshida sarcoma cells; | [38] | |
| Penetration into the model monolayer membrane is unlikely (critical surface pressure πc ≤ 27.0 μN/m). | [39] |
| Peptide | Amino Acid Sequence | Net Charge at pH 7.0 |
|---|---|---|
| Antimicrobial Activity and Toxicity | Reference | |
| Hevein | ----EQCGRQAGGKLCPNNLCCSQWGWCGSTDEYCSPDHNCQSNCKD--- | −2 |
| IC50 = 500 μg/mL against B. cinerea (MUCL 30158); IC50 = 600 μg/mL against F. culmorum (IMI 180420); IC50 = 1250 μg/mL against F. oxysporum (IMI 236441); IC50 = 300 μg/mL against Phycomyces blakesleeanus strain K1 (ATCC 5633); IC50 = 350 μg/mL against Pyrenophora tritici-repentis strain 45101; IC50 = 500 μg/mL against Pyricularia oryzae (MUCL 30166); IC50 = 500 μg/mL against Septoria nodorum (MUCL 30111); IC50 = 90 μg/mL against Trichoderma hamatum strain 10401. | [73] | |
| LAMP-1a | ---AQKCGEQGRGAKCPNCLCCGRYGFCGSTPDYCGV-G-CQSQCR-GC- | +3 |
| IC50 = 2.7 μM (24 h after inoculation) and 5.6 μM (48 h after inoculation) against Bipolaris sorokiniana strain 6/10; IC50 = 4.1 μM (24 h after inoculation) and 6.0 μM (48 h after inoculation) against F. oxysporum strain 16/10. | [74] | |
| WAMP-1a | ---AQRCGDQARGAKCPNCLCCGKYGFCGSGDAYCGA-GSCQSQCR-GC- | +3 |
| IC50 = 5 μg/mL against B. sorokiniana strain 6/10; IC50 = 20 μg/mL against B. cinerea VKM F-85; IC50 = 5 μg/mL against F. oxysporum TSA-4; IC50 = 5 μg/mL against Fusarium solani VKM F-142; IC50 = 30 μg/mL against Fusarium verticillioides VKM F-670; IC50 = 10 μg/mL against Neurospora crassa VKM F-184; Growth inhibition of Phytophtora infestans strains Pril 2 and OSV 12 at 5 μM; Growth inhibition of Pseudomonas syringae VKM B-1546, Clavibacter michiganensis subsp. michiganensis VKM Ac-1144 and Erwinia carotovora subsp. carotovora VKM B-1247 at 2.5 μg/50μL; | [75] | |
| IC50 = 2.1 μM (24 h after inoculation) and 6.2 μM (48 h after inoculation) against B. sorokiniana strain 6/10; IC50 = 2.9 μM (24 h after inoculation) and 5.9 μM (48 h after inoculation) against F. oxysporum strain 16/10. | [74] | |
| WAMP-1b | ---AQRCGDQARGAKCPNCLCCGKYGFCGSGDAYCGA-GSCQSQCR-GCR | +4 |
| IC50 = 4.9 μM against B. sorokiniana; IC50 = 16.0 μM against F. oxysporum F-137; IC50 = 18.0 μM against Alternaria alternata; No inhibition of Cladosporium cucumerinum C5 at 150 μg/mL; | [76] | |
| IC50 = 2.7 μM against F. verticillioides VKM F-670. | [77] | |
| WAMP-2 | ---AQRCGDQARGAKCPNCLCCGKYGFCGSGDAYCGK-GSCQSQCR-GCR | +5 |
| IC50 = 6.6 μM against B. sorokiniana KrD-81; IC50 = 8.8 μM against F. oxysporum F37; IC50 = 52.9 μM against F. culmorum OR-02-37; IC50 = 23.0 μM against A. alternata MRD-12; IC50 = 8.0 μM against C. cucumerinum C5; | [78] | |
| IC50 = 2.2 μM against F. verticillioides VKM F-670. | [77] | |
| WAMP-3.1 | ---AQRCGDQARGAKCPNCLCCGKYGFCGSGDAYCGE-GSCQSQCR-GCR | +3 |
| IC50 = 4.8 μM against B. sorokiniana KrD-81; IC50 = 6.8 μM against F. oxysporum F37; IC50 = 17.8 μM against A. alternata MRD-12; No inhibition of C. cucumerinum C5 at 150 μg/mL; | [78] | |
| IC50 = 3.5 μM against F. verticillioides VKM F-670. | [77] | |
| WAMP-4 | ---AQRCGDQARGAKCPNCLCCGKYGFCGSGDAYCGN-GSCQSQCR-GCR | +4 |
| IC50 = 5.2 μM against B. sorokiniana KrD-81; IC50 = 11.2 μM against F. oxysporum F37; IC50 = 23.2 μM against A. alternata MRD-12; IC50 = 12.5 μM against C. cucumerinum C5; | [78] | |
| IC50 = 3.0 μM against F. verticillioides VKM F-670. | [77] | |
| WAMP-5 | ---AQRCGDQARGAKCPNCLCCGKYGFCGSGDAYCGV-GSCQSQCR-GCR | +4 |
| IC50 = 5.4 μM against B. sorokiniana; IC50 = 12.1 μM against F. oxysporum F-137; IC50 = 13.8 μM against A. alternata; IC50 = 11.6 μM against C. cucumerinum. | [76] | |
| WAMP-N | ---AQRCGDQARGAKC | +2 |
| IC50 = 53.5 μM against B. sorokiniana KrD-81; IC50 = 174.6 μM against F. oxysporum F37; IC50 = 243.5 μM against F. culmorum OR-02-37; IC50 > 500 μM against Fusarium avenaceum Br-04-60; IC50 = 75.3 μM against A. alternata MRD-12; IC50 = 205.5 μM against C. cucumerinum C5; IC50 = 161.5 μM against Parastagonospora nodorum B-9/47. | [78] | |
| WAMP-G1 | -------------------LCCGKYGFCGSG | +1 |
| IC50 = 228.7 μM against B. sorokiniana KrD-81; IC50 > 500 μM against F. oxysporum F37; IC50 > 500 μM against F. culmorum OR-02-37; No activity against F. avenaceum Br-04-60 at 400 μg/mL; IC50 > 500 μM against A. alternata MRD-12; IC50 > 500 μM against C. cucumerinum C5; IC50 > 500 μM against P. nodorum B-9/47. | [78] | |
| WAMP-G2 | --------------------CCGKYGFCGSGDAYC | 0 |
| IC50 = 127.3 μM against B. sorokiniana KrD-81; IC50 = 255.1 μM against F. oxysporum F37; IC50 > 500 μM against F. culmorum OR-02-37; IC50 = 393.1 μM against F. avenaceum Br-04-60; IC50 = 94.9 μM against A. alternata MRD-12; IC50 = 267.4 μM against C. cucumerinum C5; IC50 = 276.5 μM against P. nodorum B-9/47. | [78] | |
| WAMP-C | -----------------------------------GK-GSCQSQCR-GCR | +3 |
| IC50 = 313.6 μM against B. sorokiniana KrD-81; IC50 > 500 μM against F. oxysporum F37; IC50 > 500 μM against F. culmorum OR-02-37; No activity against F. avenaceum Br-04-60 at 400 μg/mL; IC50 = 401.9 μM against A. alternata MRD-12; IC50 = 3.9 μM against C. cucumerinum C5; IC50 = 240.7 μM against P. nodorum B-9/47. | [78] | |
| Ee-CBP | ----QQCGRQAGNRRCANNLCCSQYGYCGRTNEYCCTSQGCQSQCRR-CG | +5 |
| IC50 = 3 μg/mL (0.6 μM) against Alternaria brassicicola MUCL 20297; IC50 = 1 μg/mL (0.2 μM) against B. cinerea MUCL 6492; IC50 = 3 μg/mL (0.6 μM) against F. culmorum IMI 180420; IC50 = 15 μg/mL (3 μM) against F. oxysporum f. sp. cubense; IC50 = 5 μg/mL (1 μM) against F. oxysporum f. sp. matthiolae CBS 247.61; IC50 = 6 μg/mL (1.2 μM) against Mycosphaerella eumusae; IC50 = 2 μg/mL (0.4 μM) against N. crassa FGSC 2489; IC50 = 33 μg/mL (6.6 μM) against Phoma exigua CBS 431.74; IC50 = 25 μg/mL (5 μM) against Phytophthora cryptogea CBS 418.71; IC50 = 33 μg/mL (6.6 μM) against Pythium ultimum MUCL 30159; IC50 = 25 μg/mL (5 μM) against Rhizoctonia solani CBS 207.84; IC50 = 100 μg/mL (20 μM) against Trichoderma hamatum ATCC 20765; IC50 = 2 μg/mL against Bacillus megaterium ATCC 13632; IC50 = 7 μg/mL against Sarcina lutea ATCC 9341. | [79], the figures in parentheses [62] | |
| EAFP1 | ----QTCASRCP-RPCNAGLCCSIYGYCGSGNAYCGA-GNCRCQCRG--- | +4 |
| IC50 = 155 μg/mL against Aculops lycopersici; IC50 = 56 μg/mL against Fusarium moniliforme; IC50 = 46 μg/mL against F. oxysporum; IC50 = 35 μg/mL against Colletotrichum gossypii; No effect on the growth of Bacillus megaterium and Pseudomonas syringae. | [80] | |
| EAFP2 | ----QTCASRCP-RPCNAGLCCSIYGYCGSGAAYCGA-GNCRCQCRG--- | +4 |
| IC50 = 109 μg/mL against A. lycopersici; IC50 = 18 μg/mL against F. moniliforme; IC50 = 94 μg/mL against F. oxysporum; IC50 = 56 μg/mL against C. gossypii; No effect on the growth of B. megaterium and P. syringae. | [80] | |
| Pn-AMP1 | ----QQCGRQASGRLCGNRLCCSQWGYCGSTASYCG--AGCQSQCRS--- | +4 |
| IC50 = 16 μg/mL against B. cinerea; IC50 = 10 μg/mL against Colletotrichum langenarium; IC50 = 11 μg/mL against Sclerotinia sclerotiorum; IC50 = 10 μg/mL against F. oxysporum; IC50 = 26 μg/mL against R. solani; IC50 = 5 μg/mL against Phytophthora capsici; IC50 = 3 μg/mL against Phytophthora parasitica cv. nicotianae; IC50 = 14 μg/mL against Saccharomyces cerevisiae EGY48; No activity against Escherichia coli, Agrobacterium tumefaciens and cultured cells Spodoptera frugiperda 9 and MA104 at concentrations up to 200 μg/mL; IC50 = 38 μg/mL against Bacillus subtilis. | [81] | |
| Pn-AMP2 | ----QQCGRQASGRLCGNRLCCSQWGYCGSTASYCG--AGCQSQCR---- | +4 |
| IC50 = 2 μg/mL against B. cinerea; IC50 = 4 μg/mL against C. langenarium; IC50 = 3 μg/mL against S. sclerotiorum; IC50 = 2.5 μg/mL against F. oxysporum; IC50 = 75 μg/mL against R. solani; IC50 = 0.6 μg/mL against P. capsici; IC50 = 2 μg/mL against P. parasitica cv. nicotianae; IC50 = 8 μg/mL against S. cerevisiae EGY48; IC50 = 2.5 μg/mL against Pythium spp.; No activity against Gram-negative E. coli, A. tumefaciens and cultured cells S. frugiperda 9 and MA104 at concentrations up to 200 μg/mL; IC50 = 20 μg/mL against B. subtilis. | [81] | |
| Fa-AMP1 | ----AQCGAQGGGATCPGGLCCSQWGWCGSTPKYCG--AGCQSNCK---- | +2 |
| IC50 = 19 μg/mL against F. oxysporum IFO 6384; IC50 = 36 μg/mL against Geotrichum candidum; IC50 = 11 μg/mL against Erwinia carotovora subsp. carotovora MAFF 106567; IC50 = 24 μg/mL against Agrobacterium radiobacter MAFF 520028; IC50 = 20 μg/mL against Agrobacterium rhizogenes MAFF 210265; IC50 = 14 μg/mL against C. michiganensis subsp. michiganensis MAFF 301044; IC50 = 13 μg/mL against Curtobacterium flaccumfaciens pv. oorti MAFF 301203. | [82] | |
| Fa-AMP2 | ----AQCGAQGGGATCPGGLCCSQWGWCGSTPKYCG--AGCQSNCR---- | +2 |
| IC50 = 29 μg/mL against F. oxysporum IFO 6384; IC50 = 25 μg/mL against G. candidum; IC50 = 15 μg/mL against E. carotovora subsp. carotovora MAFF 106567; IC50 = 17 μg/mL against A. radiobacter MAFF 520028; IC50 = 24 μg/mL against A. rhizogenes MAFF 210265; IC50 = 17 μg/mL against C. michiganensis subsp. michiganensis MAFF 301044; IC50 = 15 μg/mL against C. flaccumfaciens pv. oorti MAFF 301203. | [82] | |
| mO1 | ----QNCGRQAGNRACANQLCCSQYGFCGSTSEYCSRANGCQSNCRGG-- | +3 |
| IC50 = 25.51 μg/mL against A. alternata CICC 2465; IC50 = 60.43 μg/mL against A. brassicicola CICC 2646; No activity against Curvularia lunata CICC 40301, F. oxysporum CICC 2532, Aspergillus niger CICC 2089, Verticillium dahilae CICC 2534, R. solani CICC 40259 at a concentration of 70 μg/mL; No significant cytotoxic effect on Vero cells with concentrations up to 100 μM. | [58] | |
| vH2 | ----FQCGRQAGGARCSNGLCCSQFGYCGSTPPYCGA-GQCQSQC----- | +2 |
| IC50 = 21.87 μg/mL against A. alternata CICC 40292; IC50 = 16.10 μg/mL against C. lunata CICC 40301; IC50 = 5.05 μg/mL against F. oxysporum CICC 2532; IC50 = 1.77 μg/mL against R. solani CICC 40259. | [54] | |
| gB5 | ---DPTCSKL-GDFKCNPGRCCSKFNYCGSTAAYCGR-GNCIAQCP---- | +3 |
| IC50 = 6.8 μg/mL against A. niger; IC50 = 10.0 μg/mL against C. lunata CICC 40301; IC50 = 69.2 μg/mL against F. oxysporum CICC 2532; IC50 = 20.0 μg/mL against R. solani CICC 40259. | [53] | |
| Ac-AMP1 | ---VGEC---VRG-RCPSGMCCSQFGYCGKGPKYCG-------------- | +3 |
| IC50 = 7 μg/mL against A. brassicola; IC50 = 8 μg/mL against A. pisi; IC50 = 10 μg/mL against B. cinerea; IC50 = 8 μg/mL against C. lindemuthianum; IC50 = 2 μg/mL against F. culmorum; IC50 = 7 μg/mL against T. hamatum; IC50 = 6 μg/mL against V. dahlia; IC50 = 40 μg/mL against B. megaterium; IC50 = 250 μg/mL against S. lutea. | [83] | |
| Ac-AMP2 | ---VGEC---VRG-RCPSGMCCSQFGYCGKGPKYCGR------------- | +4 |
| IC50 = 4 μg/mL against A. brassicola; IC50 = 8 μg/mL against A. pisi; IC50 = 8 μg/mL against B. cinerea; IC50 = 8 μg/mL against C. lindemuthianum; IC50 = 2 μg/mL against F. culmorum; IC50 = 3 μg/mL against T. hamatum; IC50 = 8 μg/mL against V. dahliae; IC50 = 10 μg/mL against B. megaterium; IC50 = 40 μg/mL against S. lutea; | [83] | |
| IC50 = 50 μg/mL against A. brassicicola MUCL 20297; IC50 = 2 μg/mL against B. cinerea MUCL 6492; IC50 = 6 μg/mL against F. culmorum IMI 180420; IC50 = 100 μg/mL against F. oxysporum f. sp. cubense; IC50 = 30 μg/mL against F. oxysporum f. sp. matthiolae CBS 247.61; IC50 = 8 μg/mL against Mycosphaerella eumusae; IC50 = 3 μg/mL against N. crassa FGSC 2489; IC50 = 30 μg/mL against Phoma exigua CBS 431.74; IC50 = 50 μg/mL against Phytophthora cryptogea CBS 418.71; IC50 = 95 μg/mL against Pythium ultimum MUCL 30159; IC50 = 100 μg/mL against R. solani CBS 207.84; IC50 = 100 μg/mL against T. hamatum ATCC 20765; IC50 = 7 μg/mL against B. megaterium ATCC 13632; IC50 = 20 μg/mL against S. lutea ATCC 9341. | [79] | |
| Ar-AMP | ---AGEC---VQG-RCPSGMCCSQFGYCGRGPKYCGR------------- | +3 |
| Growth inhibition of F. culmorum at 3.5 μM; Growth inhibition of Helminthosporium sativum and B. cinerea at 10.6 μM; Growth inhibition of Alternaria consortiale at 31.8 μM; No growth inhibition of R. solani at 286.0 μM. | [84] | |
| IWF4 | ---SGECNMYG---RCPPGYCCSKFGYCGVGRAYCG-------------- | +2 |
| IC50 = 0.7 μM against Cercospora beticola isolate FC573 (oxidized and nonoxidized IWF4 showed the same activity level). | [55] | |
| SmAMP3 | VGPGGECGGRFGG--CAGGQCCSRFGFCGSGPKYCAH------------- | +2 |
| IC50 = 5.4 μM against A. niger; IC50 = 2.0 μM against B. sorokiniana; IC50 = 1.6 μM against B. cinerea; IC50 = 3.7 μM against F. solani; IC50 = 5.0 μM against A. alternata. | [85] | |
| SmAMP1.1 | SGPNGQCGPGWGG--CRGGLCCSQYGYCGSGPKYCAH------------- | +2 |
| IC50 = 2.4 μM against B. cinerea; IC50 = 3.5 μM against F. solani; IC50 = 2.6 μM against A. alternata. | [85] |
| Peptide | Amino Acid Sequence | Net Charge at pH 7.0 |
|---|---|---|
| Biological Activity | Reference | |
| MBP-1 | ------RSGRGECRRQCLRRHEGQPWETQECMRRCRRRG---- | +7 |
| 99.9% growth inhibition of E. coli DH5 at a concentration of 3 μg/mL; 81% growth inhibition of C. michiganense ssp. nebraskense at a concentration of 30 μg/mL; Almost complete growth inhibition of F. graminearum, Sclerotina sclerotiorum, and Alternaria longipes at 60 μg/mL; Almost complete growth inhibition of Sclerotina trifoliorum at 30 μg/mL; Growth inhibition of F. moniliforme at 60 μg/mL; Weak growth inhibition of Aspergillus flavus at 60 μg/mL; | [101] | |
| MIC = 50 μM against E. coli DH5-α. | [100] | |
| Var 1 (W20A) | ------RSGRGECRRQCLRRHEGQPAETQECMRRCRRRG---- | +7 |
| MIC > 400 μM against E. coli DH5-α. | [100] | |
| Var 2 | ------RSGRGEARRQALRRHEGQPWETQEAMRRARRRG---- | +7 |
| MIC > 400 μM against E. coli DH5-α. | [100] | |
| VhTI | ------NTDPEQCKVMCYAQRHSSPELLRRCLDNCEKEHD--- | −2 |
| Trypsin inhibitor. | [90] | |
| VhTI (5-31) | ----------EQCKVMCYAQRHSSPELLRRCLDNCEK | +1 |
| Trypsin inhibitor. | [90] | |
| FtAMP | GSSEKPQQELEECQNVCRMKRWSTEMVHRCEKKCEEKFERQQR | +1 |
| Ki (trypsin) = 1.90 × 10−9 M; No inhibitory activity against elastase or α-chymotrypsin. MIC > 128 μM against E. coli BNCC 337271; MIC = 128 μM against B. subtilis BNCC 124990; MIC = 128 μM against S. aureus BNCC 186335; MIC = 16 μM against F. oxysporum BNCC 164775; MIC = 8 μM against Rhizopus sp. BNCC 147803; MIC = 8 μM against Trichoderma koningii BNCC 189731. | [94] | |
| FtAMP-R21A | GSSEKPQQELEECQNVCRMKAWSTEMVHRCEKKCEEKFERQQR | 0 |
| 17.3% tripsin-inhibitory activity of FtAMP; Ki (elastase) = 2.47 × 10−9 M; MIC > 128 μM against E. coli BNCC 337271; MIC = 128 μM against B. subtilis BNCC 124990; MIC = 128 μM against S. aureus BNCC 186335; MIC = 16 μM against F. oxysporum BNCC 164775; MIC = 8 μM against Rhizopus sp. BNCC 147803; MIC = 8 μM against T. koningii BNCC 189731. | [94] | |
| FtAMP-R21F | GSSEKPQQELEECQNVCRMKFWSTEMVHRCEKKCEEKFERQQR | 0 |
| 14.7% tripsin-inhibitory activity of FtAMP; Ki (α-chymotrypsin) = 2.73 × 10−9 M; MIC > 128 μM against E. coli BNCC 337271; MIC = 128 μM against B. subtilis BNCC 124990; MIC = 128 μM against S. aureus BNCC 186335; MIC = 16 μM against F. oxysporum BNCC 164775; MIC = 8 μM against Rhizopus sp. BNCC 147803; MIC = 8 μM against T. koningii BNCC 189731. | [94] | |
| Sm-AMP-X | ----VDPDVRAYCKHQCMSTRGDQARKICESVCMRQD------ | +1 |
| IC50 = 12.5 μM against A. alternata strain DVZ; IC50 = 4.0 μM against A. niger VKM F-33; IC50 > 32.0 μM against B. sorokiniana 6/10; IC50 = 16.2 μM against B. cinerea SGR-1; IC50 = 5.4 μM against F. oxysporum 16/10; IC50 = 7.2 μM against F. solani IVK; IC50 > 32.0 μM against P. infestans OSV 12; IC50 > 32.0 μM against P. ultimum F-1506; No activity against C. michiganensis subsp. michiganensis VKM Ac-1144, E. carotovora subsp. carotovora VKM B-1247, E. coli XL1-blue, and P. syringae VKM B-1546 at concentrations up to 40 μM. | [99] | |
| Sm-AMP-L | ----VDPDVRAYCKHQCLSTRGDQARKICESVCLRQD------ | +1 |
| IC50 = 12.9 μM against A. alternata strain DVZ; IC50 = 3.6 μM against A. niger VKM F-33; IC50 > 32.0 μM against B. sorokiniana 6/10; IC50 = 18.0 μM against B. cinerea SGR-1; IC50 = 6.1 μM against F. oxysporum 16/10; IC50 = 7.2 μM against F. solani IVK; IC50 > 32.0 μM against P. infestans OSV 12; IC50 > 32.0 μM against P. ultimum F-1506. | [99] | |
| Sm-AMP-X1 | ------------CKHQCMSTRGDQARKICESVCM | +2 |
| IC50 > 32.0 μM against A. alternata strain DVZ; IC50 = 8.0 μM against A. niger VKM F-33; IC50 > 32.0 μM against B. sorokiniana 6/10; IC50 > 32.0 μM against B. cinerea SGR-1; IC50 = 24.4 μM against F. oxysporum 16/10; IC50 = 19.0 μM against F. solani IVK; IC50 > 32.0 μM against P. infestans OSV 12; IC50 > 32.0 μM against P. ultimum F-1506. | [99] | |
| Sm-AMP-X2 | ----------------CMSTRGDQARKICE | +1 |
| IC50 > 32.0 μM against A. alternata strain DVZ; IC50 = 16.9 μM against A. niger VKM F-33; IC50 > 32.0 μM against B. sorokiniana 6/10; IC50 > 32.0 μM against B. cinerea SGR-1; IC50 = 25.0 μM against F. oxysporum 16/10; IC50 = 22.5 μM against F. solani IVK; IC50 > 32.0 μM against P. infestans OSV 12; IC50 > 32.0 μM against P. ultimum F-1506. | [99] | |
| Tk-AMP-X1 | --------TDDRCERMCQHYHDRREKKQCMKGCRYGESD--- | +1 |
| IC50 = 7.5 μg/mL against F. graminearum; IC50 = 15.0 μg/mL against F. verticillioides; IC50 = 30 μg/mL against Diplodia maydis; IC50 > 30 μg/mL against Colletotrichum graminicola. | [96] | |
| Tk-AMP-X2 | --------ADDRCERMCQRYHDRREKKQCMKGCRYG------ | +4 |
| IC50 = 7.5 μg/mL against F. graminearum; IC50 = 10.0 μg/mL against F. verticillioides; IC50 = 17 μg/mL against D. maydis; IC50 > 30 μg/mL against C. graminicola; | [96] | |
| No activity against the tested voltage-gated potassium channels (members of the Shaker (Kv1.1–Kv1.6 and Shaker IR), Shab (Kv2.1), Shaw (Kv3.1), and erg (hERG) families) even at concentrations up to 250 μM. | [97] | |
| Tk-hefu | --------ADDRCYRMCQRYHDRREKKQCKEGCRYG------ | +4 |
| Selectively targets members of the Shaker family: at 40 μM, it inhibited the potassium currents through Kv1.2, Kv1.3, and Kv1.6 channels by 8.3, 58.4, and 7.3%, respectively. No activity on other channels. It blocked Kv1.3 channels with similar potency (IC50 34.0 μM) to κ-hefutoxin 1 (IC50~40.0 μM). | [97] | |
| EcAMP1 | ------GSGRGSCRSQCMRRHEDEPWRVQECVSQCRRRRGGGD | +4 |
| EC50 = 16.0 μM against A. alternata; EC50 = 14.0 μM against A. solani; EC50 > 32 μM against A. niger; EC50 = 18.2 μM against B. sorokiniana; EC50 > 10 μM against C.graminicola; EC50 > 10 μM against D. maydis; EC50 = 4.5 μM against F. graminearum; EC50 = 8.5 μM against F. oxysporum; EC50 = 4.0 μM against F. solani; EC50 = 8.1 μM against F. verticillioides; EC50 = 6.0 μM against Phoma betae; EC50 = 16.3 μM against P. infestans; EC50 = 12.0 μM against Pythium debaryanum; EC50 = 14.4 μM against P. ultimum; EC50 > 32 μM against Trichoderma album; | [91] | |
| IC50 = 3.8 μM against F. solani; | [102] | |
| IC50 = 5 μM against S. aureus; No activity against E. coli and P. aeruginosa at 80 μM; IC50 = 0.625 μM (MIC99 = 1.25 μM) against Candida albicans; IC50 = 6.8 (5.0)* μM against F. graminearum VKM F-1668; IC50 = 12.9 (9.4)* μM against F. oxysporum TSKHA-4; IC50 = 5.4 (5.6)* μM against F. solani; IC50 > 32 μM against A. niger VKM F-33; IC50 = 25.7 μM against B. sorokiniana VKM F-1446; IC50 = 18.4 μM against A. alternata. | [103] | |
| EcAMP1-X1 | ------------CRSQCMRRHEDEPWRVQECVSQC | 0 |
| IC50 = 9.0 μM against F. graminearum VKM F-1668; IC50 = 15.4 μM against F. oxysporum TSKHA-4; IC50 = 6.9 μM against F. solani; IC50 > 32 μM against A. niger VKM F-33; IC50 > 32.0 μM against B. sorokiniana VKM F-1446; IC50 = 21.1 μM against A. alternata; No activity against S. aureus, E. coli, P. aeruginosa, and C. albicans at 80 μM. | [103] | |
| EcAMP1-X2 | ----------------CMRRHEDEPWRVQEC | −1 |
| IC50 = 18.1 μM against F. graminearum VKM F-1668; IC50 = 23.2 μM against F. oxysporum TSKHA-4; IC50 = 11.0 μM against F. solani; IC50 > 32 μM against A. niger VKM F-33; IC50 > 32.0 μM against B. sorokiniana VKM F-1446; IC50 > 32.0 μM against A. alternata; No activity against S. aureus, E. coli, P. aeruginosa, and C. albicans at 80 μM. | [103] | |
| EcAMP1-X3 | ------GSGRGSCRSQCMRRHEDEPARVQECVSQCRRRRGGGD | +4 |
| IC50 = 9.9 μM against F. graminearum VKM F-1668; IC50 = 15.0 μM against F. oxysporum TSKHA-4; IC50 = 8.6 μM against F. solani. | [103] | |
| EcAMP1-X4 | ------GSGRGSCRSQCMRRHEDEPWRVQECVSQCRR | +3 |
| IC50 = 8.5 μM against F. graminearum VKM F-1668; IC50 = 15.8 μM against F. oxysporum TSKHA-4; IC50 = 7.8 μM against F. solani. | [103] | |
| EcAMP1-Hyp | ------GSGRGSCRSQCMRRHEDEOWRVQECVSQCRRRRGGGD | nd |
| IC50 = 5.4 μM against F. solani. | [102] |
| Peptide | Amino Acid Sequence | Net Charge at pH 7.0 |
|---|---|---|
| Antimicrobial Activity and Toxicity | Reference | |
| Ib-AMP2 | QYGRRCCNWGPGRRYCKRWC | +6 |
| IC50 = 12 μg/mL against Alternaria longipes CBS62083; IC50 = 25 μg/mL against B. cinerea K1147; IC50 = 6 μg/mL against Cladosporium sphaerospermum K0791; IC50 = 6 μg/mL against F. culmorum K0311; IC50 = 6 μg/mL against Penicillium digitatum K0879; IC50 = 12 μg/mL against Trichoderma viride K1127; IC50 = 12 μg/mL against Verticillium alboatrum K0937; No activity on erythrocytes and cultured fibroblasts at a concentration of 200 μg/mL. | [9] | |
| Ib-AMP3 | QYRHRCCAWGPGRKYCKRWC | +6 |
| IC50 = 6 μg/mL against A. longipes CBS62083; IC50 = 6 μg/mL against B. cinerea K1147; IC50 = 3 μg/mL against C. sphaerospermum K0791; IC50 = 6 μg/mL against F. culmorum K0311; IC50 = 3 μg/mL against P. digitatum K0879; IC50 = 12 μg/mL against T. viride K1127; IC50 = 6 μg/mL against V. alboatrum K0937. | [9] | |
| Ib-AMP4 | QWGRRCCGWGPGRRYCRRWC | +6 |
| IC50 = 3 μg/mL against A. longipes CBS62083; IC50 = 6 μg/mL against B. cinerea K1147; IC50 = 1 μg/mL against C. sphaerospermum K0791; IC50 = 1 μg/mL against F. culmorum K0311; IC50 = 3 μg/mL against P. digitatum K0879; IC50 = 6 μg/mL against T. viride K1127; IC50 = 6 μg/mL against V. alboatrum K0937; IC50 = 5 μg/mL against Bacillus subtilis JHCC 55331; IC50 = 5 μg/mL against Micrococcus luteus ATCC 9341; IC50 = 20 μg/mL against Staphylococcus aureus ATCC 25923; IC50 = 5 μg/mL against Streptococcus faecalis ATCC 29212; IC50 > 500 μg/mL against E. coli HB101; IC50 > 500 μg/mL against Proteus vulgaris JHCC 558711; IC50 > 100 μg/mL against Pseudomonas solanacearum R48/a; IC50 > 100 μg/mL against Erwinia amylovora CFBP1430; IC50 = 6 μg/mL against X. campestris INRA 10342; IC50 = 15 μg/mL against X. oryzae ETH 698; No activity on erythrocytes and cultured fibroblasts at a concentration of 200 μg/mL. | [9] | |
| Ib-AMP1 | QWGRRCCGWGPGRRYCVRWC | +5 |
| IC50 = 3 μg/mL against A. longipes CBS62083; IC50 = 12 μg/mL against B. cinerea K1147; IC50 = 1 μg/mL against C. sphaerospermum K0791; IC50 = 1 μg/mL against F. culmorum K0311; IC50 = 3 μg/mL against P. digitatum K0879; IC50 = 6 μg/mL against T. viride K1127; IC50 = 3 μg/mL against V. alboatrum K0937; IC50 = 10 μg/mL against B. subtilis JHCC 55331; IC50 = 10 μg/mL against M. luteus ATCC 9341; IC50 = 30 μg/mL against S. aureus ATCC 25923; IC50 = 6 μg/mL against S. faecalis ATCC 29212; IC50 > 500 μg/mL against E. coli HB101; IC50 > 500 μg/mL against P. vulgaris JHCC 558711; IC50 > 500 μg/mL against P. solanacearum R48/a; | [9] | |
| MIC = 2.5 μM against Aspergillus flavus KCTC 1375; MIC = 5 μM against Candida albicans. | [106] | |
| Ib-AMP1 (reduced form) | QWGRRCCGWGPGRRYCVRWC | +5 |
| MIC = 10 μM against A. flavus KCTC 1375; MIC = 20 μM against C. albicans. | [106] | |
| Ib-AMP4 [111] | EWGRRCCGWGPGRRYCRRWC | +5 |
| IC50 = 3 (2.5)* μM against B. cinerea JHCC 8973; IC50 = 1.0 (2.5)* μM against F. culmorum IMI 180420; IC50 = 1.2 μM against Neurospora crassa FGSC 2489; IC50 = 13 μM against Saccharomyces cerevisiae BY4741; IC50 = 5 μM against Pichia pastoris GS115. | [111] | |
| Ib-AMP1 [111] | EWGRRCCGWGPGRRYCVRWC | +4 |
| IC50 = 1.5 μM against B. cinerea JHCC 8973; IC50 = 1.4 μM against F. culmorum IMI 180420; IC50 = 0.5 μM against N. crassa FGSC 2489; IC50 = 15 μM against S. cerevisiae BY4741; IC50 = 16 μM against P. pastoris GS115; | [111] | |
| MIC = 16 μM against E. coli KCTC 1682; MIC > 32 μM against Pseudomonas aeruginosa KCTC 1637; MIC > 32 μM against P. aeruginosa (MDRPA) CCARM 2095; MIC > 32 μM against Salmonella typhimurium KCTC 1926; MIC = 16 μM against B. subtilis KCTC 3068; MIC = 16 μM against Staphylococcus epidermidis KCTC 1917; MIC = 16 μM against S. aureus KCTC 1621; MIC = 16 μM against S. aureus (MRSA) CCARM 3543. | [107] | |
| Analog 1 | -WGRR--GWGPGRRY-VRW-NH2 | nd |
| MIC = 16 μM against E. coli KCTC 1682; MIC = 16 μM against P. aeruginosa KCTC 1637; MIC = 16 μM against P. aeruginosa (MDRPA) CCARM 2095; MIC = 4 μM against S. typhimurium KCTC 1926; MIC = 8 μM against B. subtilis KCTC 3068; MIC = 8 μM against S. epidermidis KCTC 1917; MIC = 4 μM against S. aureus KCTC 1621; MIC = 2 μM against S. aureus (MRSA) CCARM 3543. | [107] | |
| Analog 2 | -WGRR--GWGpGRRY-VRW-NH2 | nd |
| MIC = 8 μM against E. coli KCTC 1682; MIC = 8 μM against P. aeruginosa KCTC 1637; MIC = 32 μM against P. aeruginosa (MDRPA) CCARM 2095; MIC = 4 μM against S. typhimurium KCTC 1926; MIC = 4 μM against B. subtilis KCTC 3068; MIC = 8 μM against S. epidermidis KCTC 1917; MIC = 4 μM against S. aureus KCTC 1621; MIC = 2 μM against S. aureus (MRSA) CCARM 3543. | [107] | |
| Analog 3 | --WGRR--GWGaGRRY-VRW-NH2 | nd |
| MIC = 16 μM against E. coli KCTC 1682; MIC = 16 μM against P. aeruginosa KCTC 1637; MIC = 16 μM against P. aeruginosa (MDRPA) CCARM 2095; MIC = 4 μM against S. typhimurium KCTC 1926; MIC = 4 μM against B. subtilis KCTC 3068; MIC = 8 μM against S. epidermidis KCTC 1917; MIC = 4 μM against S. aureus KCTC 1621; MIC = 2 μM against S. aureus (MRSA) CCARM 3543. | [107] | |
| Analog 4 | --WGRR--GWGkGRRY-VRW-NH2 | nd |
| MIC = 8 μM against E. coli KCTC 1682; MIC = 8 μM against P. aeruginosa KCTC 1637; MIC = 16 μM against P. aeruginosa (MDRPA) CCARM 2095; MIC = 4 μM against S. typhimurium KCTC 1926; MIC = 4 μM against B. subtilis KCTC 3068; MIC = 8 μM against S. epidermidis KCTC 1917; MIC = 4 μM against S. aureus KCTC 1621; MIC = 2 μM against S. aureus (MRSA) CCARM 3543. | [107] | |
| MCE01 | <EWGRRBBGWGPGRRYBVRWB | nd |
| IC50 = 1.5 μM against B. cinerea JHCC 8973; IC50 = 3 μM against N. crassa FGSC 2489; IC50 = 5 μM against F. culmorum IMI 180420; IC50 = 20 μM against S. cerevisiae BY4741; IC50 = 6 μM against P. pastoris GS115. | [111] | |
| MCE02 | <EWGRRBBGWGPGRRYBRRWB | nd |
| IC50 = 2 (4.5)* μM against B. cinerea JHCC 8973; IC50 = 3 μM against N. crassa FGSC 2489; IC50 = 5 (4.5)* μM against F. culmorum IMI 180420; IC50 = 20 μM against S. cerevisiae BY4741; IC50 = 4 μM against P. pastoris GS115. | [111] | |
| MCD26 | <ewgrrbbgwgpgrrybvrwb | nd |
| IC50 = 0.5 μM against B. cinerea JHCC 8973; IC50 = 0.8 μM against N. crassa FGSC 2489; IC50 = 1.4 μM against F. culmorum IMI 180420; IC50 = 2 μM against S. cerevisiae BY4741; IC50 = 2 μM against P. pastoris GS115. | [111] | |
| MCD30 | <ewgrrbbgwgpgrrybrrwb | nd |
| IC50 = 1 μM against B. cinerea JHCC 8973; IC50 = 1.5 μM against N. crassa FGSC 2489; IC50 = 0.5 μM against F. culmorum IMI 180420; IC50 = 7.5 μM against S. cerevisiae BY4741; IC50 = 1 μM against P. pastoris GS115. | [111] | |
| MCC02 | -RWGRRBBGWGPGRRYBRRWB | nd |
| IC50 = 3.5 μM against B. cinerea JHCC 8973. | [111] | |
| MCC03 | <ERGRRBBGWGPGRRYBRRWB | nd |
| IC50 = 3.5 μM against B. cinerea JHCC 8973. | [111] | |
| MCC04 | <EWRRRBBGWGPGRRYBRRWB | nd |
| IC50 = 3.0 μM against B. cinerea JHCC 8973. | [111] | |
| MCC05 | <EWGRRRBGWGPGRRYBRRWB | nd |
| IC50 = 4.5 μM against B. cinerea JHCC 8973. | [111] | |
| MCC06 | <EWGRRBRGWGPGRRYBRRWB | nd |
| IC50 = 3.0 μM against B. cinerea JHCC 8973. | [111] | |
| MCC07 | <EWGRRBBRWGPGRRYBRRWB | nd |
| IC50 = 3.0 μM against B. cinerea JHCC 8973. | [111] | |
| MCC08 | <EWGRRBBGRGPGRRYBRRWB | nd |
| IC50 = 2.5 μM against B. cinerea JHCC 8973. | [111] | |
| MCC09 | <EWGRRBBGWRPGRRYBRRWB | nd |
| IC50 = 2.5 μM against B. cinerea JHCC 8973. | [111] | |
| MCC10 | <EWGRRBBGWGRGRRYBRRWB | nd |
| IC50 = 2.5 μM against B. cinerea JHCC 8973. | [111] | |
| MCC11 | <EWGRRBBGWGPRRRYBRRWB | nd |
| IC50 = 3.0 μM against B. cinerea JHCC 8973. | [111] | |
| MCC12 | <EWGRRBBGWGPGRRRBRRWB | nd |
| IC50 = 3.0 μM against B. cinerea JHCC 8973. | [111] | |
| MCC13 | <EWGRRBBGWGPGRRYRRRWB | nd |
| IC50 = 3.0 μM against B. cinerea JHCC 8973. | [111] | |
| MCC14 | <EWGRRBBGWGPGRRYBRRRB | nd |
| IC50 = 4.0 μM against B. cinerea JHCC 8973. | [111] | |
| MCC15 | <EWGRRBBGWGPGRRYBRRWR | nd |
| IC50 = 4.5 μM against B. cinerea JHCC 8973. | [111] | |
| MCC16 | -WWGRRBBGWGPGRRYBRRWB | nd |
| IC50 = 6.5 μM against F. culmorum IMI 180420. | [111] | |
| MCC17 | <EWWRRBBGWGPGRRYBRRWB | nd |
| IC50 = 3.0 μM against F. culmorum IMI 180420. | [111] | |
| MCC18 | <EWGWRBBGWGPGRRYBRRWB | nd |
| IC50 = 3.0 μM against F. culmorum IMI 180420. | [111] | |
| MCC19 | <EWGRWBBGWGPGRRYBRRWB | nd |
| IC50 = 3.0 μM against F. culmorum IMI 180420. | [111] | |
| MCC21 | <EWGRRWBGWGPGRRYBRRWB | nd |
| IC50 = 4.5 μM against F. culmorum IMI 180420. | [111] | |
| MCC22 | <EWWRRBWGWGPGRRYBRRWB | nd |
| IC50 = 2.5 μM against F. culmorum IMI 180420. | [111] | |
| MCC23 | <EWWRRBBWWGPGRRYBRRWB | nd |
| IC50 = 3.0 μM against F. culmorum IMI 180420. | [111] | |
| MCC24 | <EWWRRBBGWWPGRRYBRRWB | nd |
| IC50 = 3.0 μM against F. culmorum IMI 180420. | [111] | |
| MCC25 | <EWWRRBBGWGWGRRYBRRWB | nd |
| IC50 = 3.0 μM against F. culmorum IMI 180420. | [111] | |
| MCD17 | <EWWRRBBGWGPWRRYBRRWB | nd |
| IC50 = 3.5 μM against F. culmorum IMI 180420. | [111] | |
| MCD18 | <EWWRRBBGWGPGWRYBRRWB | nd |
| IC50 = 3.5 μM against F. culmorum IMI 180420. | [111] | |
| MCD19 | <EWWRRBBGWGPGRWYBRRWB | nd |
| IC50 = 3.5 μM against F. culmorum IMI 180420. | [111] | |
| MCD21 | <EWWRRBBGWGPGRRWBRRWB | nd |
| IC50 = 3.0 μM against F. culmorum IMI 180420. | [111] | |
| MCD22 | <EWWRRBBGWGPGRRYWRRWB | nd |
| IC50 = 3.0 μM against F. culmorum IMI 180420. | [111] | |
| MCD23 | <EWWRRBBGWGPGRRYBWRWB | nd |
| IC50 = 3.0 μM against F. culmorum IMI 180420. | [111] | |
| MCD24 | <EWWRRBBGWGPGRRYBRWWB | nd |
| IC50 = 2.0 μM against F. culmorum IMI 180420. | [111] | |
| MCD25 | <EWWRRBBGWGPGRRYBRRWW | nd |
| IC50 = 2.0 μM against F. culmorum IMI 180420. | [111] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Slezina, M.P.; Odintsova, T.I. Plant Antimicrobial Peptides: Insights into Structure-Function Relationships for Practical Applications. Curr. Issues Mol. Biol. 2023, 45, 3674-3704. https://doi.org/10.3390/cimb45040239
Slezina MP, Odintsova TI. Plant Antimicrobial Peptides: Insights into Structure-Function Relationships for Practical Applications. Current Issues in Molecular Biology. 2023; 45(4):3674-3704. https://doi.org/10.3390/cimb45040239
Chicago/Turabian StyleSlezina, Marina P., and Tatyana I. Odintsova. 2023. "Plant Antimicrobial Peptides: Insights into Structure-Function Relationships for Practical Applications" Current Issues in Molecular Biology 45, no. 4: 3674-3704. https://doi.org/10.3390/cimb45040239
APA StyleSlezina, M. P., & Odintsova, T. I. (2023). Plant Antimicrobial Peptides: Insights into Structure-Function Relationships for Practical Applications. Current Issues in Molecular Biology, 45(4), 3674-3704. https://doi.org/10.3390/cimb45040239