

Phenotypic Characterization and Comparative Genomic Analyses of Mycobacteriophage WIVsmall as A New Member Assigned to F1 Subcluster

Abstract
:1. Introduction
2. Materials and Methods
2.1. Phage Isolation and Preparation
2.2. DNA Extraction
2.3. Sequence Data and Phylogenetic Analysis
2.4. Optimal Multiplicity of Infection (MOI)
2.5. Single-Step Growth Experiment
2.6. Stability of the Phage under Various Conditions
2.7. Genomic Bioinformatics Analysis
2.8. Identification of Phage Structural Proteins
3. Results
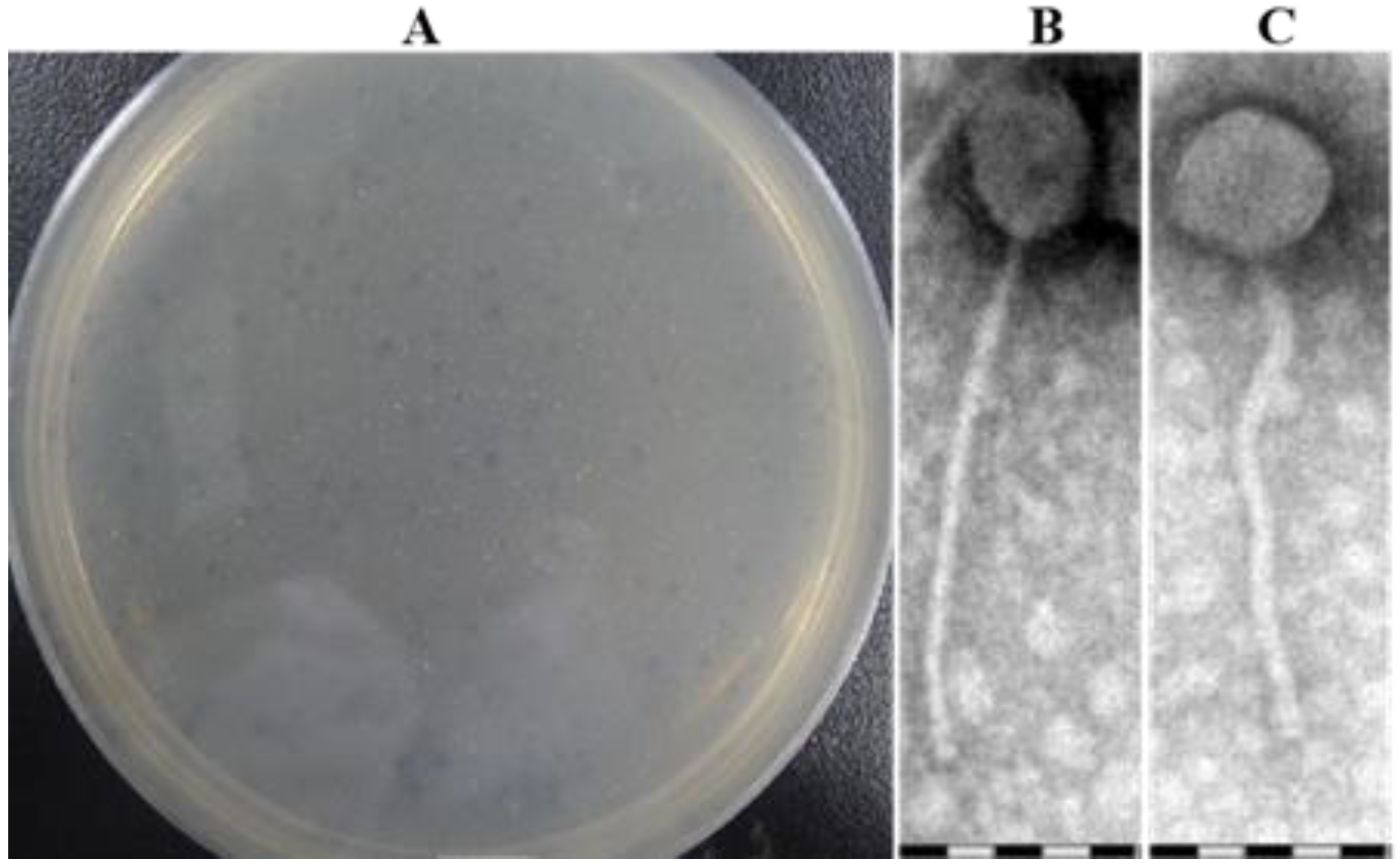
3.1. Phage Isolation and Morphology Analysis of Plaque
3.2. Optimal Multiplicity of Infection (MOI) and One-Step Growth Curve
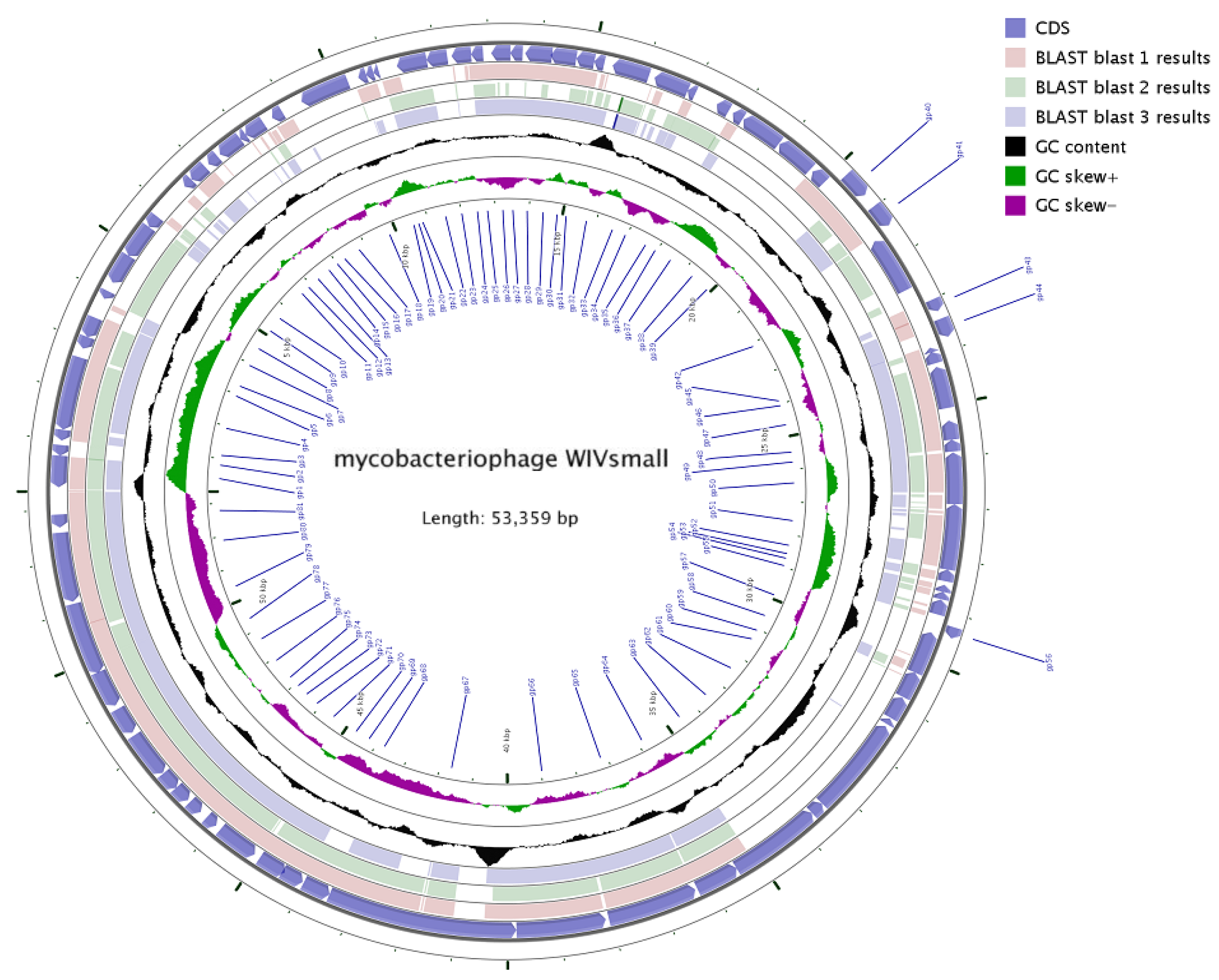
3.3. General Genome Analysis
3.4. Putative Functions of the Predicted ORFs
3.5. Phylogenetic Relationships
3.6. Comparative Genomics Analysis
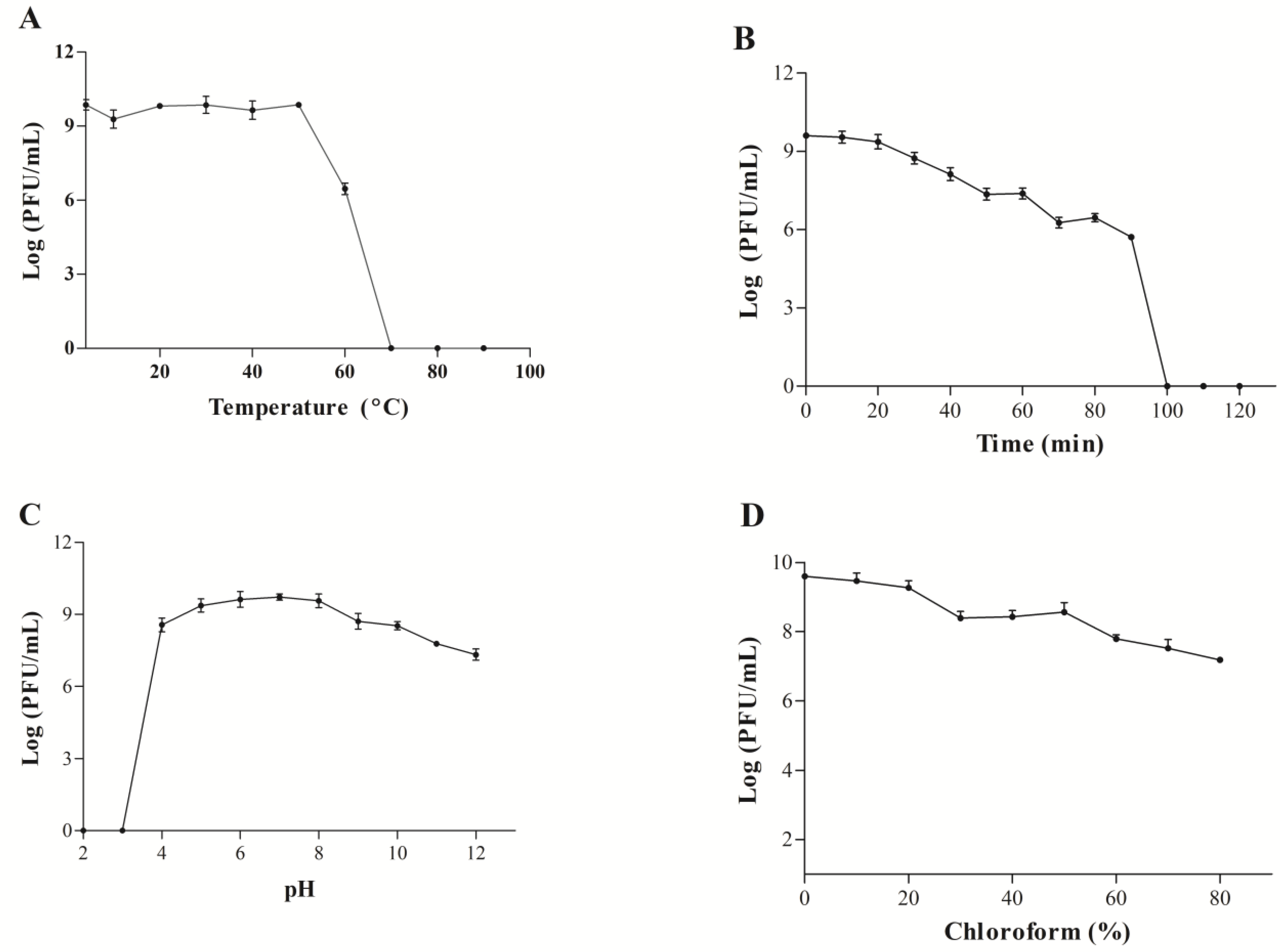
3.7. Biological Characteristics of the WIVsmall Phage
3.8. Mass-Spectrometric Identification of Phage Proteins
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Darby, E.M.; Trampari, E. Molecular mechanisms of antibiotic resistance revisited. Nat. Rev. Microbiol. 2023, 21, 280–295. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.H.; Hsieh, Y.H.; Powers, Z.M.; Kao, C.Y. Defeating Antibiotic-Resistant Bacteria: Exploring Alternative Therapies for a Post-Antibiotic Era. Int. J. Mol. Sci. 2020, 21, 1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrigy, N.B.; Larsen, S.E.; Reese, V.; Pecor, T.; Harrison, M.; Kuehl, P.J.; Hatfull, G.F.; Sauvageau, D.; Baldwin, S.L.; Finlay, W.H.; et al. Prophylaxis of Mycobacterium tuberculosis H37Rv Infection in a Preclinical Mouse Model via Inhalation of Nebulized Bacteriophage D29. Antimicrob. Agents Chemother. 2019, 63, e00871. [Google Scholar] [CrossRef]
- Li, X.; He, Y.; Wang, Z.; Wei, J.; Hu, T.; Si, J.; Tao, G.; Zhang, L.; Xie, L.; Abdalla, A.E.; et al. A combination therapy of Phages and Antibiotics: Two is better than one. Int. J. Biol. Sci. 2021, 17, 3573–3582. [Google Scholar] [CrossRef]
- Jariah, R.O.A.; Hakim, M.S. Interaction of phages, bacteria, and the human immune system: Evolutionary changes in phage therapy. Rev. Med. Virol. 2019, 29, e2055. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Wu, N.; Zeng, Y.; Chen, L.; Li, L.; Yang, L.; Zhang, Y.; Guo, M.; Li, L.; Li, J.; et al. Non-active antibiotic and bacteriophage synergism to successfully treat recurrent urinary tract infection caused by extensively drug-resistant Klebsiella pneumoniae. Emerg. Microbes Infect. 2020, 9, 771–774. [Google Scholar] [CrossRef] [Green Version]
- Nick, J.A.; Dedrick, R.M.; Gray, A.L.; Vladar, E.K.; Smith, B.E.; Freeman, K.G.; Malcolm, K.C.; Epperson, L.E.; Hasan, N.A.; Hendrix, J.; et al. Host and pathogen response to bacteriophage engineered against Mycobacterium abscessus lung infection. Cell 2022, 185, 1860–1874.e12. [Google Scholar] [CrossRef]
- Dedrick, R.M.; Freeman, K.G.; Nguyen, J.A.; Bahadirli-Talbott, A.; Cardin, M.E.; Cristinziano, M.; Smith, B.E.; Jeong, S.; Ignatius, E.H.; Lin, C.T.; et al. Nebulized Bacteriophage in a Patient With Refractory Mycobacterium abscessus Lung Disease. Open Forum Infect. Dis. 2022, 9, ofac194. [Google Scholar] [CrossRef]
- Tarín-Pelló, A.; Suay-García, B.; Pérez-Gracia, M.T. Antibiotic resistant bacteria: Current situation and treatment options to accelerate the development of a new antimicrobial arsenal. Expert Rev. Anti-Infect. Ther. 2022, 20, 1095–1108. [Google Scholar] [CrossRef]
- Chakaya, J.; Petersen, E.; Nantanda, R.; Mungai, B.N.; Migliori, G.B.; Amanullah, F.; Lungu, P.; Ntoumi, F.; Kumarasamy, N.; Maeurer, M.; et al. The WHO Global Tuberculosis 2021 Report—Not so good news and turning the tide back to End TB. Int. J. Infect. Dis. 2022, 124 (Suppl. 1), S26–S29. [Google Scholar] [CrossRef]
- Traoré, A.N.; Rikhotso, M.C. Effectiveness of the Novel Anti-TB Bedaquiline against Drug-Resistant TB in Africa: A Systematic Review of the Literature. Pathogens 2022, 11, 636. [Google Scholar] [CrossRef] [PubMed]
- Walker, T.M.; Miotto, P.; Köser, C.U.; Fowler, P.W.; Knaggs, J.; Iqbal, Z.; Hunt, M.; Chindelevitch, L.; Farhat, M.; Cirillo, D.M.; et al. The 2021 WHO catalogue of Mycobacterium tuberculosis complex mutations associated with drug resistance: A genotypic analysis. Lancet Microbe 2022, 3, e265–e273. [Google Scholar] [CrossRef] [PubMed]
- Hosseiniporgham, S.; Sechi, L.A. A Review on Mycobacteriophages: From Classification to Applications. Pathogens 2022, 11, 777. [Google Scholar] [CrossRef]
- Dedrick, R.M.; Smith, B.E.; Cristinziano, M.; Freeman, K.G.; Jacobs-Sera, D.; Belessis, Y.; Whitney Brown, A.; Cohen, K.A.; Davidson, R.M.; van Duin, D.; et al. Phage Therapy of Mycobacterium Infections: Compassionate Use of Phages in 20 Patients With Drug-Resistant Mycobacterial Disease. Clin. Infect. Dis. 2023, 76, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Little, J.S.; Dedrick, R.M.; Freeman, K.G.; Cristinziano, M.; Smith, B.E.; Benson, C.A.; Jhaveri, T.A.; Baden, L.R.; Solomon, D.A.; Hatfull, G.F. Bacteriophage treatment of disseminated cutaneous Mycobacterium chelonae infection. Nat. Commun. 2022, 13, 2313. [Google Scholar] [CrossRef] [PubMed]
- Rondón, L.; Urdániz, E.; Latini, C.; Payaslian, F.; Matteo, M.; Sosa, E.J.; Do Porto, D.F.; Turjanski, A.G.; Nemirovsky, S.; Hatfull, G.F.; et al. Fluoromycobacteriophages Can Detect Viable Mycobacterium tuberculosis and Determine Phenotypic Rifampicin Resistance in 3-5 Days From Sputum Collection. Front. Microbiol. 2018, 9, 1471. [Google Scholar] [CrossRef]
- Li, X.; Hu, T.; Wei, J.; He, Y.; Abdalla, A.E.; Wang, G.; Li, Y.; Teng, T. Characterization of a Novel Bacteriophage Henu2 and Evaluation of the Synergistic Antibacterial Activity of Phage-Antibiotics. Antibiotics 2021, 10, 174. [Google Scholar] [CrossRef]
- Teng, T.; Li, Q.; Liu, Z.; Li, X.; Liu, Z.; Liu, H.; Liu, F.; Xie, L.; Wang, H.; Zhang, L.; et al. Characterization and genome analysis of novel Klebsiella phage Henu1 with lytic activity against clinical strains of Klebsiella pneumoniae. Arch. Virol. 2019, 164, 2389–2393. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Teng, T.; Zhang, G.; Fan, X.; Zhang, Z.; Zhang, L.; Wu, D.; Chen, S.; Li, Y.; Jin, J. Complete genome sequence analysis of PS2, a novel T4-like bacteriophage that infects Serratia marcescens clinical isolates. Arch. Virol. 2018, 163, 1997–2000. [Google Scholar] [CrossRef]
- Wdowiak, M.; Paczesny, J.; Raza, S. Enhancing the Stability of Bacteriophages Using Physical, Chemical, and Nano-Based Approaches: A Review. Pharmaceutics 2022, 14, 1936. [Google Scholar] [CrossRef] [PubMed]
- Besemer, J.; Borodovsky, M. GeneMark: Web software for gene finding in prokaryotes, eukaryotes and viruses. Nucleic Acids Res. 2005, 33, W451–W454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Chan, P.P.; Lin, B.Y.; Mak, A.J.; Lowe, T.M. tRNAscan-SE 2.0: Improved detection and functional classification of transfer RNA genes. Nucleic Acids Res. 2021, 49, 9077–9096. [Google Scholar] [CrossRef]
- Laslett, D.; Canback, B. ARAGORN, a program to detect tRNA genes and tmRNA genes in nucleotide sequences. Nucleic Acids Res. 2004, 32, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Stothard, P.; Grant, J.R.; Van Domselaar, G. Visualizing and comparing circular genomes using the CGView family of tools. Brief. Bioinform. 2019, 20, 1576–1582. [Google Scholar] [CrossRef] [Green Version]
- Carver, T.; Thomson, N.; Bleasby, A.; Berriman, M.; Parkhill, J. DNAPlotter: Circular and linear interactive genome visualization. Bioinformatics 2009, 25, 119–120. [Google Scholar] [CrossRef] [Green Version]
- Gudlavalleti, B.S.; Phung, T.; Barton, C.L.; Becker, A.; Graul, B.L.; Griffin, J.T.; Hays, C.J.; Horn, B.; Liang, D.R.; Rutledge, L.M.; et al. Whole genome sequencing identifies an allele responsible for clear vs. turbid plaque morphology in a Mycobacteriophage. BMC Microbiol. 2020, 20, 148. [Google Scholar] [CrossRef]
- Kumar, V.; Loganathan, P.; Sivaramakrishnan, G.; Kriakov, J.; Dusthakeer, A.; Subramanyam, B.; Chan, J.; Jacobs, W.R., Jr.; Paranji Rama, N. Characterization of temperate phage Che12 and construction of a new tool for diagnosis of tuberculosis. Tuberculosis 2008, 88, 616–623. [Google Scholar] [CrossRef] [Green Version]
- Lo, C.Y.; Gao, Y. DNA Helicase-Polymerase Coupling in Bacteriophage DNA Replication. Viruses 2021, 13, 1739. [Google Scholar] [CrossRef]
- Abouhmad, A.; Korany, A.H.; Hatti-Kaul, R.; Grey, C.; Dishisha, T. Exploring the Enzymatic and Antibacterial Activities of Novel Mycobacteriophage Lysin B Enzymes. Int. J. Mol. Sci. 2020, 21, 3176. [Google Scholar] [CrossRef] [PubMed]
- Catalão, M.J.; Pimentel, M. Mycobacteriophage Lysis Enzymes: Targeting the Mycobacterial Cell Envelope. Viruses 2018, 10, 428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bavda, V.R.; Yadav, A.; Jain, V. Decoding the molecular properties of mycobacteriophage D29 Holin provides insights into Holin engineering. J. Virol. 2021, 95, e02173-20. [Google Scholar] [CrossRef]
- Ghai, R.; Mehrshad, M.; Mizuno, C.M.; Rodriguez-Valera, F. Metagenomic recovery of phage genomes of uncultured freshwater actinobacteria. ISME J. 2017, 11, 304–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatfull, G.F. Mycobacteriophages: Genes and genomes. Annu. Rev. Microbiol. 2010, 64, 331–356. [Google Scholar] [CrossRef] [PubMed]
- Hatfull, G.F.; Jacobs-Sera, D.; Lawrence, J.G.; Pope, W.H.; Russell, D.A.; Ko, C.C.; Weber, R.J.; Patel, M.C.; Germane, K.L.; Edgar, R.H.; et al. Comparative genomic analysis of 60 Mycobacteriophage genomes: Genome clustering, gene acquisition, and gene size. J. Mol. Biol. 2010, 397, 119–143. [Google Scholar] [CrossRef] [Green Version]
- Gissendanner, C.R.; Wiedemeier, A.M.; Wiedemeier, P.D.; Minton, R.L.; Bhuiyan, S.; Harmson, J.S.; Findley, A.M. A web-based restriction endonuclease tool for mycobacteriophage cluster prediction. J. Basic Microbiol. 2014, 54, 1140–1145. [Google Scholar] [CrossRef] [PubMed]
- Paton, N.I.; Cousins, C.; Suresh, C.; Burhan, E.; Chew, K.L.; Dalay, V.B.; Lu, Q.; Kusmiati, T.; Balanag, V.M.; Lee, S.L.; et al. Treatment Strategy for Rifampin-Susceptible Tuberculosis. N. Engl. J. Med. 2023, 388, 873–887. [Google Scholar] [CrossRef]
- Hatfull, G.F. Mycobacteriophages: From Petri dish to patient. PLoS Pathog. 2022, 18, e1010602. [Google Scholar] [CrossRef]
- Peng, H.; Borg, R.E.; Dow, L.P.; Pruitt, B.L.; Chen, I.A. Controlled phage therapy by photothermal ablation of specific bacterial species using gold nanorods targeted by chimeric phages. Proc. Natl. Acad. Sci. USA 2020, 117, 1951–1961. [Google Scholar] [CrossRef] [Green Version]
- Yehl, K.; Lemire, S.; Yang, A.C.; Ando, H.; Mimee, M.; Torres, M.T.; de la Fuente-Nunez, C.; Lu, T.K. Engineering Phage Host-Range and Suppressing Bacterial Resistance through Phage Tail Fiber Mutagenesis. Cell 2019, 179, 459–469.e9. [Google Scholar] [CrossRef] [PubMed]
- Kot, W.; Hammer, K.; Neve, H.; Vogensen, F.K. Identification of the receptor-binding protein in lytic Leuconostoc pseudomesenteroides bacteriophages. Appl. Environ. Microbiol. 2013, 79, 3311–3314. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ORF Number | Start and Stop Position | Strand | Length (bp) | MW (kDa) | Molecular Function | E-Value | Coverage (%) | Identity (%) |
|---|---|---|---|---|---|---|---|---|
| 1 | 83–697 | − | 615 | 22.83 | Glucosyltransferase | 2E-142 | 100 | 95.59 |
| 2 | 694–918 | − | 225 | 8.08 | Hypothetical protein | 2E-43 | 100 | 95.59 |
| 3 | 989–1201 | − | 213 | 7.78 | Hypothetical protein | 2E-42 | 100 | 97.14 |
| 4 | 1204–2823 | − | 1620 | 61.18 | Glycosyltransferase | 0.00 | 100 | 93.70 |
| 5 | 2820–3029 | − | 210 | 7.46 | Hypothetical protein | 7E-43 | 100 | 100 |
| 6 | 3059–3460 | − | 402 | 14.36 | Hypothetical protein | 4E-89 | 100 | 99.25 |
| 6.1 | 3457–3984 | − | 528 | 20.44 | Hypothetical protein | 3E-69 | 71 | 86.40 |
| 6.2 | 3702–3839 | − | 138 | 4.86 | Hypothetical protein | 1E-24 | 100 | 100 |
| 7 | 3866–4030 | − | 165 | 6.07 | GIY-YIG nuclease | 8E-25 | 90 | 95.92 |
| 7.1 | 3936–4181 | + | 246 | 8.75 | Hypothetical protein | 1E-27 | 77 | 95 |
| 8 | 4217–4819 | − | 603 | 22.39 | HtrLYibB protein | 2E-143 | 100 | 98.50 |
| 9 | 4816–5496 | − | 681 | 25.77 | Hypothetical protein | 2E-162 | 100 | 100 |
| 10 | 5499–5795 | − | 297 | 11.10 | Hypothetical protein | 3E-53 | 84 | 100 |
| 11 | 5832–6020 | + | 189 | 6.66 | Hypothetical protein | 2E-38 | 100 | 100 |
| 11.1 | 6167–6469 | − | 303 | 10.65 | Hypothetical protein | 5E-21 | 67 | 80.36 |
| 12 | 6531–6716 | − | 186 | 7.12 | Hypothetical protein | 9E-36 | 100 | 98.36 |
| 13 | 6719–7075 | − | 357 | 13.68 | Mobile element MPME | 2E-79 | 96 | 99.12 |
| 14 | 7158–7385 | − | 228 | 8.38 | Hypothetical protein | 9E-43 | 97 | 95.89 |
| 15 | 7444–7932 | − | 489 | 17.83 | Hypothetical protein | 2E-113 | 100 | 99.38 |
| 15.1 | 7929–8066 | − | 138 | 4.71 | Hypothetical protein | 8E-22 | 100 | 97.78 |
| 16 | 8063–8470 | − | 408 | 14.38 | Hypothetical protein | 1E-37 | 95 | 68.09 |
| 16.1 | 8393–8683 | + | 291 | 10.59 | Hypothetical protein | 3E-40 | 85 | 91 |
| 16.2 | 8467–8661 | − | 195 | 7.07 | Hypothetical protein | 1E-38 | 100 | 98.44 |
| 17 | 8661–8885 | − | 225 | 8.37 | Hypothetical protein | 3E-40 | 100 | 93.24 |
| 17.1 | 9041–9304 | − | 264 | 9.31 | Hypothetical protein | 1E-52 | 100 | 95.40 |
| 18 | 9301–10,263 | − | 963 | 34.65 | Replication initiation protein | 0.0 | 100 | 98.44 |
| 18.1 | 10,474–10,716 | − | 243 | 9.25 | Hypothetical protein | 4E-42 | 100 | 86.25 |
| 19 | 10,632–10,778 | − | 147 | 5.40 | Hypothetical protein | 5E-27 | 100 | 100 |
| 20 | 10,778–10,882 | − | 105 | 3.79 | Hypothetical protein | 2E-14 | 100 | 97.06 |
| 20.1 | 10,879–11,241 | − | 363 | 13.72 | HNH endonuclease | 6E-83 | 100 | 98.33 |
| 21 | 11,238–11,816 | − | 579 | 20.60 | Single strand annealing protein | 3E-135 | 100 | 99.48 |
| 22 | 11,813–12,295 | − | 483 | 18.13 | HNH endonuclease | 5E-113 | 100 | 97.50 |
| 23 | 12,292–12,666 | − | 375 | 13.94 | DNA methyltransferase | 2E-67 | 100 | 89.68 |
| 24 | 12,663–12,893 | − | 231 | 8.26 | Hypothetical protein | 3E-43 | 100 | 97.37 |
| 25 | 13,048–13,413 | − | 366 | 13.39 | DNA binding protein | 3E-83 | 100 | 99.17 |
| 26 | 13,410–13,643 | − | 234 | 8.57 | WhiB transcriptional factor | 3E-44 | 94 | 98.63 |
| 27 | 13,692–14,129 | − | 438 | 16.57 | Hypothetical protein | 2E-98 | 100 | 100 |
| 28 | 14,174–14,668 | − | 495 | 18.60 | WhiB transcriptional factor | 6E-117 | 100 | 98.78 |
| 29 | 14,668–15,015 | − | 348 | 12.35 | Hypothetical protein | 3E-62 | 99 | 79.82 |
| 30 | 15,012–15,290 | − | 279 | 10.67 | Hypothetical protein | 7E-56 | 100 | 93.48 |
| 31 | 15,369–16,169 | − | 801 | 27.87 | Hypothetical protein | 2E-109 | 100 | 76.69 |
| 32 | 15,379–16,041 | + | 663 | 22.30 | Hypothetical protein | 1E-139 | 100 | 100 |
| 33 | 16,199–16,765 | − | 567 | 20.32 | Hypothetical protein | 5E-28 | 100 | 56.12 |
| 33.1 | 16,873–17,031 | − | 159 | 6.10 | Hypothetical protein | 2E-27 | 100 | 100 |
| 33.2 | 17,058–17,489 | − | 432 | 16.18 | HNH endonuclease | 9E-91 | 100 | 78.57 |
| 34 | 17,489–17,746 | − | 258 | 9.82 | Excisionase | 1E-51 | 100 | 96.51 |
| 34.1 | 17,727–18,215 | − | 99 | 5.87 | Hypothetical protein | 2E-11 | 100 | 96.88 |
| 34.2 | 17,743–17,841 | − | 324 | 11.63 | Hypothetical protein | 1E-42 | 66 | 98.59 |
| 35 | 17,838–18,161 | − | 324 | 11.63 | Hypothetical protein | 1E-42 | 66 | 98.59 |
| 36 | 18,076–18,891 | − | 816 | 30.12 | Hypothetical protein | 2E-26 | 95 | 32 |
| 37 | 18,907–19,506 | − | 600 | 21.83 | Hypothetical protein | 2E-85 | 98 | 67.30 |
| 38 | 19,729–20,034 | − | 306 | 11.50 | Transcriptional repressor | 2E-67 | 100 | 99.01 |
| 39 | 20,218–20,721 | + | 504 | 18.73 | Transcriptional repressor | 3E-112 | 100 | 97.60 |
| 40 | 20,984–21,430 | + | 447 | 15.55 | Pin protein | 2E-83 | 100 | 91.33 |
| 41 | 21,522–22,640 | − | 1119 | 41.52 | Integrase | 0.0 | 100 | 99.73 |
| 42 | 21,960–22,634 | + | 675 | 22.65 | Hypothetical protein | 2E-148 | 100 | 100 |
| 43 | 23,014–23,250 | + | 237 | 8.49 | Hypothetical protein | 5E-22 | 94 | 62.67 |
| 44 | 23,250–23,417 | + | 168 | 8.49 | Hypothetical protein | 6E-31 | 100 | 98.18 |
| 45 | 23,399–23,758 | − | 360 | 13.37 | Hypothetical protein | 2E-77 | 94 | 98.23 |
| 46 | 23,928–24,161 | + | 234 | 8.39 | Hypothetical protein | 1E-46 | 100 | 98.70 |
| 47 | 24,234–25,055 | − | 822 | 31.03 | DNA polymerase exonuclease subunit | 0.0 | 100 | 99.63 |
| 47.1 | 25,042–25,302 | − | 261 | 9.92 | Hypothetical protein | 4E-56 | 100 | 100 |
| 48 | 25,224–25,931 | − | 708 | 25.33 | Hypothetical protein | 8E-169 | 100 | 100 |
| 49 | 25,299–25,673 | − | 375 | 14.21 | Minor tail protein | 2E-80 | 100 | 99.19 |
| 49.1 | 25,670–25,903 | − | 234 | 7.78 | Holin | 7E-43 | 100 | 98.70 |
| 50 | 25,920–26,921 | − | 1002 | 36.63 | Endolysin | 0.0 | 100 | 98.80 |
| 51 | 26,921–28,198 | − | 1278 | 47.42 | Endolysin | 0.0 | 100 | 95.29 |
| 52 | 28,195–28,428 | − | 234 | 8.29 | Hypothetical protein | 2E-41 | 100 | 98.70 |
| 53 | 28,499–28,639 | − | 141 | 4.94 | Hypothetical protein | 1E-22 | 100 | 95.65 |
| 54 | 28,636–28,785 | − | 150 | 5.38 | Hypothetical protein | 7E-21 | 95 | 85.11 |
| 55 | 28,792–29,088 | − | 297 | 10.31 | Hypothetical protein | 4E-41 | 100 | 84.69 |
| 55.1 | 29,232–29,444 | + | 213 | 8.16 | Hypothetical protein | 4E-36 | 100 | 92.86 |
| 56 | 29,452–30,291 | − | 840 | 26.26 | Minor tail protein | 2E-98 | 100 | 90.32 |
| 57 | 30,288–30,815 | − | 528 | 17.50 | Minor tail protein | 2E-50 | 84 | 61.49 |
| 58 | 30,830–31,282 | − | 453 | 15.63 | Hypothetical protein | 7E-46 | 100 | 62.00 |
| 59 | 31,299–31,745 | + | 447 | 15.54 | Hypothetical protein | 7E-08 | 32 | 56.00 |
| 60 | 31,468–33,264 | − | 1797 | 59.21 | Hypothetical protein | Minor tail protein | 7E-50 | 38 |
| 61 | 32,291–33,682 | − | 1392 | 45.56 | Minor tail protein | 7E-07 | 67 | 25 |
| 61.1 | 33,395–33,610 | − | 216 | 8.00 | Hypothetical protein | 6E-16 | 100 | 52.44 |
| 62 | 33,660–35,447 | − | 1788 | 63.70 | Minor tail protein | 6E-94 | 63 | 49.00 |
| 63 | 35,444–36,289 | − | 846 | 28.88 | Minor tail protein | 0.0 | 99 | 98.21 |
| 64 | 36,332–38,065 | + | 1734 | 64.58 | Minor tail protein | 0.0 | 98 | 98.77 |
| 65 | 38,124–39,833 | − | 1710 | 63.44 | Putative structural protein | 0.0 | 100 | 98.95 |
| 66 | 39,819–42,185 | + | 2367 | 88.37 | Hypothetical protein | 0.0 | 100 | 100 |
| 67 | 39,834–43,472 | + | 3639 | 122.34 | Taillength tape measure protein | 0.0 | 100 | 98.51 |
| 68 | 42,040–42,633 | − | 594 | 20.41 | Hypothetical protein | 7E-136 | 100 | 100 |
| 69 | 42,931–43,464 | − | 534 | 18.59 | Hypothetical protein | 1E-113 | 100 | 100 |
| 70 | 43,472–44,011 | − | 540 | 20.12 | HNH endonuclease | 1E-129 | 100 | 98.88 |
| 71 | 44,035–44,538 | − | 504 | 19.21 | Tail assembly chaperone | 4E-114 | 100 | 98.20 |
| 72 | 44,420–44,971 | − | 552 | 20.50 | Tail assembly chaperone | 2E-129 | 100 | 99.45 |
| 73 | 45,090–45,899 | − | 810 | 29.78 | Major tail protein | 0.0 | 100 | 99.26 |
| 74 | 46,008–46,412 | − | 405 | 14.70 | Head–tail adaptor | 6E-89 | 100 | 99.25 |
| 75 | 46,402–46,626 | − | 225 | 7.83 | Head–tail connector protein | 8E-46 | 100 | 98.65 |
| 75.1 | 46,735–47,064 | − | 330 | 11.74 | Head–tail adaptor | 3E-70 | 100 | 98.17 |
| 76 | 47,061–47,432 | − | 372 | 13.21 | Head–tail adaptor Ad1 | 5E-81 | 100 | 99.19 |
| 77 | 47,445–48,353 | − | 909 | 31.71 | Major capsid subunit | 5E-162 | 99 | 74.67 |
| 78 | 48,416–48,952 | − | 537 | 19.55 | Head scaffolding protein | 8E-122 | 100 | 99.44 |
| 79 | 49,031–49,582 | − | 552 | 20.14 | Head maturation protease | 1E-131 | 100 | 98.91 |
| 80 | 49,740–51,161 | − | 1422 | 51.43 | Portal protein | 0.0 | 100 | 99.15 |
| 81 | 51,202–52,689 | − | 1488 | 53.90 | Terminase | 0.0 | 100 | 99.60 |
| 82 | 52,661–52,915 | − | 255 | 9.36 | Terminase small subunit | 4E-51 | 100 | 98.81 |
| 83 | 52,988–53,320 | − | 333 | 12.13 | HNH endonuclease | 3E-73 | 100 | 99.09 |
| 83.1 | 53,146–53,358 | − | 213 | 7.59 | Hypothetical protein | 9E-36 | 95 | 98.51 |
| Phage Name | Query Cover | Identity | Accession Number | Genome Size (bp) |
|---|---|---|---|---|
| SG4 | 64% | 95.74% | NC_026593.1 | 59,419 |
| Ramsey | 63% | 96% | NC_011289.1 | 58,578 |
| Coco12 | 62% | 95.34% | NC_051644.1 | 57,693 |
| Job42 | 62% | 97.47% | NC_021538.1 | 59,626 |
| BuzzLyseyear | 56% | 88.73% | NC_023699.1 | 59,016 |
| ShiLan | 48% | 93.36% | NC_041988.1 | 59,794 |
| Pacc40 | 47% | 93.53% | NC_011287.1 | 58,554 |
| Squirty | 37% | 95% | NC_026588.1 | 60,285 |
| Jabbawokkie | 22% | 95.03% | NC_022069.1 | 55,213 |
| Yoshi | 18% | 80.90% | NC_042030.1 | 58,714 |
| Protein | Strand | Location | AA Length | Best Match | |||||
|---|---|---|---|---|---|---|---|---|---|
| Phage_Gene | Host | Phage Cluster | Predicted Function | Cov. %/ Ident. % | E Value | ||||
| Gp58 | - | 30,830– 31,282 | 453 | TPA4_27 | Tsukamurella paurometabola CON55 | Singleton | Hypothetical protein | 100/74 | 7E-62 |
| GMA1_27 | Gordonia malaquae G239 | Singleton | Hypothetical protein | 100/62 | 7E-46 | ||||
| REQ3_58 | Rhodococcus equi Requ28 | Singleton | Hypothetical protein | 100/58 | 2E-45 | ||||
| PatrickStar_38 | Gordonia terrae 3612 | CX | Hypothetical protein | 96/58 | 3E-43 | ||||
| Keelan_56 | Gordonia terrae 3612 | DP | Hypothetical protein | 100/55 | 2E-39 | ||||
| Gp59 | - | 31,299– 31,745 | 447 | IdentityCrisis_23 | Mycobacterium smegmatis mc2155 | Singleton | Hypothetical protein | 32/56 | 7E-08 |
| Shweta_22 | Mycobacterium smegmatis mc2155 | N | Hypothetical protein | 34/49 | 8E-07 | ||||
| Ruthiejr_30 | Mycobacterium smegmatis mc2155 | K4 | Hypothetical protein | 34/49 | 8E-07 | ||||
| Willsammy_23 | Mycobacterium smegmatis mc2155 | P1 | Hypothetical protein | 34/46 | 7E-06 | ||||
| Taquito_28 | Mycobacterium smegmatis mc2155 | K4 | Hypothetical protein | 34/46 | 7E-06 | ||||
| Gp61 | - | 32,291– 33,682 | 1392 | Moosehead_28 | Gordonia terrae 3612 | CZ6 | Hypothetical protein | 67/25 | 7E-07 |
| Wilder_24 | Mycobacterium smegmatis mc2155 | L2 | Minor tail protein | 40/23 | 1E-06 | ||||
| MkaliMitinis3_24 | Mycobacterium smegmatis mc2155 | L2 | Minor tail protein | 50/23 | 1E-06 | ||||
| LilDestine_24 | Mycobacterium smegmatis mc2155 | L2 | Minor tail protein | 50/23 | 1E-06 | ||||
| Lewan_24 | Mycobacterium smegmatis mc2155 | Singleton | Minor tail protein | 50/23 | 1E-06 | ||||
| No. | Detected Proteins | Predicated Function | Molecular Mass |
|---|---|---|---|
| 1 | Gp60 | Phage minor tail protein | 59.30 kDa |
| 2 | Gp62 | Phage minor tail protein | 63.75 kDa |
| 3 | Gp64 | Phage minor tail protein | 64.63 kDa |
| 4 | Gp65 | Putative structural protein | 63.52 kDa |
| 5 | Gp67 | Tail-length tape measure protein | 122.46 kDa |
| 6 | Gp73 | Major tail protein | 29.82 kDa |
| 7 | Gp77 | Major capsid subunit | 31.73 kDa |
| 8 | Gp80 | Putative portal protein | 51.45 kDa |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, X.; Zhang, J.; Wang, Y.; Zhou, F.; Li, Q.; Teng, T. Phenotypic Characterization and Comparative Genomic Analyses of Mycobacteriophage WIVsmall as A New Member Assigned to F1 Subcluster. Curr. Issues Mol. Biol. 2023, 45, 6432-6448. https://doi.org/10.3390/cimb45080406
Guo X, Zhang J, Wang Y, Zhou F, Li Q, Teng T. Phenotypic Characterization and Comparative Genomic Analyses of Mycobacteriophage WIVsmall as A New Member Assigned to F1 Subcluster. Current Issues in Molecular Biology. 2023; 45(8):6432-6448. https://doi.org/10.3390/cimb45080406
Chicago/Turabian StyleGuo, Xinge, Jing Zhang, Yuhan Wang, Fang Zhou, Qiming Li, and Tieshan Teng. 2023. "Phenotypic Characterization and Comparative Genomic Analyses of Mycobacteriophage WIVsmall as A New Member Assigned to F1 Subcluster" Current Issues in Molecular Biology 45, no. 8: 6432-6448. https://doi.org/10.3390/cimb45080406
APA StyleGuo, X., Zhang, J., Wang, Y., Zhou, F., Li, Q., & Teng, T. (2023). Phenotypic Characterization and Comparative Genomic Analyses of Mycobacteriophage WIVsmall as A New Member Assigned to F1 Subcluster. Current Issues in Molecular Biology, 45(8), 6432-6448. https://doi.org/10.3390/cimb45080406