
Escin Activates Canonical Wnt/β-Catenin Signaling Pathway by Facilitating the Proteasomal Degradation of Glycogen Synthase Kinase-3β in Cultured Human Dermal Papilla Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. TOPFlash Reporter Assay
2.4. Western Blotting
2.5. Quantitative Real-Time PCR
2.6. Immunocytochemistry
2.7. Data and Statistical Analysis
3. Results
3.1. Escin Activated Wnt/β-Catenin Signaling in WRHEK293A Cells
3.2. Escin Activated Wnt/β-Catenin Signaling in Cultured hDPCs
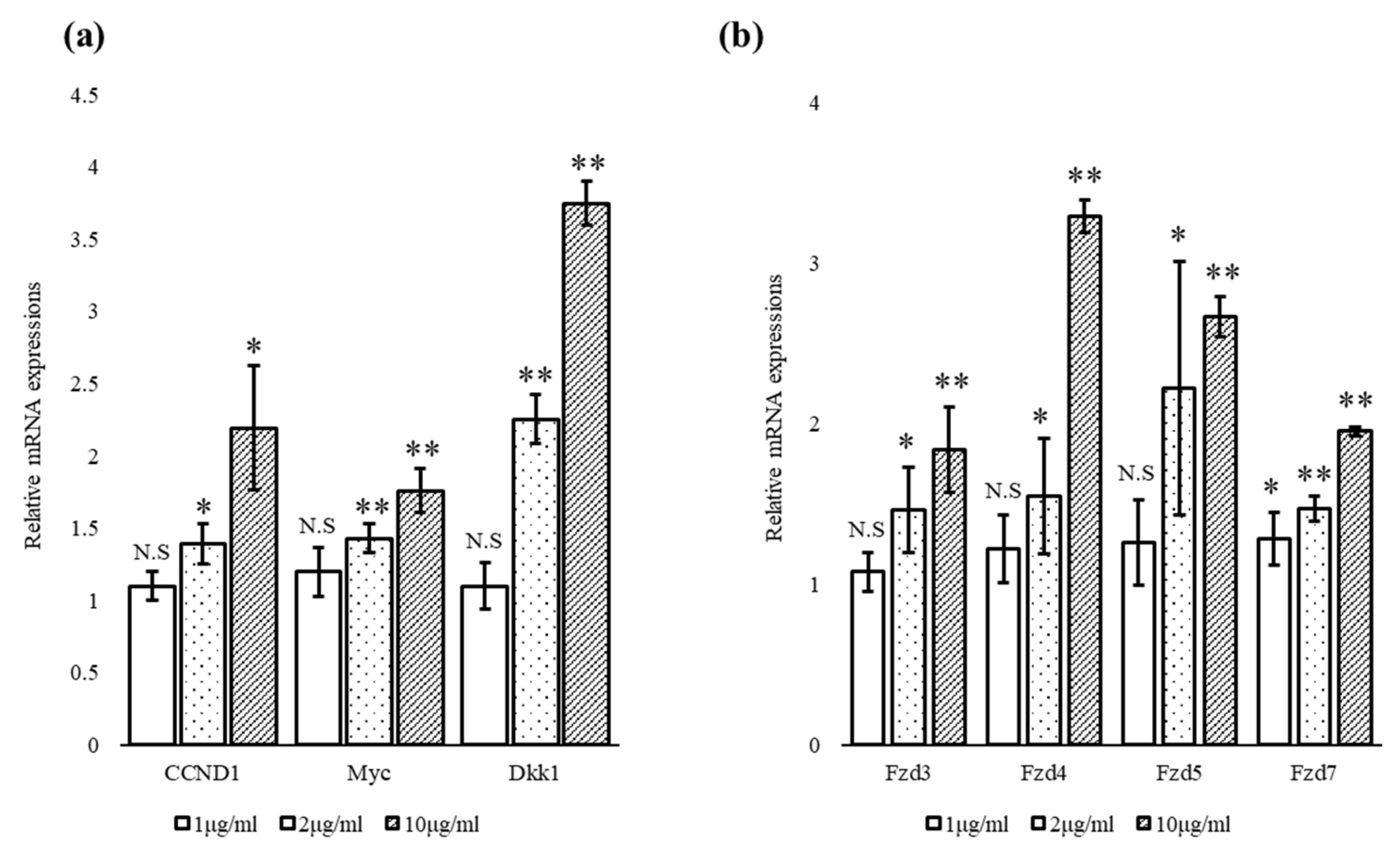
3.3. Escin Stimulated the Expression of Wnt Target Genes and Wnt Receptors
3.4. Escin Facilitated the Degradation of GSK-3β via 26S Proteasome
3.5. Escin-Induced TOPFlash Activities Were Abrogated by sFRP1 and sFRP2 Treatment
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Clevers, H.; Nusse, R. Wnt/beta-catenin signaling and disease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacDonald, B.T.; Tamai, K.; He, X. Wnt/beta-catenin signaling: Components, mechanisms, and diseases. Dev. Cell 2009, 17, 9–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamos, J.L.; Weis, W.I. The beta-catenin destruction complex. Cold Spring Harb. Perspect. Biol. 2013, 5, a007898. [Google Scholar] [CrossRef] [PubMed]
- Aberle, H.; Bauer, A.; Stappert, J.; Kispert, A.; Kemler, R. beta-catenin is a target for the ubiquitin-proteasome pathway. EMBO J. 1997, 16, 3797–3804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubinfeld, B.; Albert, I.; Porfiri, E.; Fiol, C.; Munemitsu, S.; Polakis, P. Binding of GSK3beta to the APC-beta-catenin complex and regulation of complex assembly. Science 1996, 272, 1023–1026. [Google Scholar] [CrossRef]
- Wang, X.; Dong, L.; Cheng, J.; Verdine, G.L.; Lin, A.; Chu, Q. Targeted beta-catenin ubiquitination and degradation by multifunctional stapled peptides. J. Pept. Sci. 2022, 28, e3389. [Google Scholar] [CrossRef]
- Clevers, H. Wnt/beta-catenin signaling in development and disease. Cell 2006, 127, 469–480. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.; Kim, A.; Li, B.; Ransick, A.; Bugacov, H.; Chen, X.; Lindstrom, N.; Brown, A.; Oxburgh, L.; Ren, B.; et al. A beta-catenin-driven switch in TCF/LEF transcription factor binding to DNA target sites promotes commitment of mammalian nephron progenitor cells. Elife 2021, 10, e64444. [Google Scholar] [CrossRef]
- Kretzschmar, K.; Clevers, H. Wnt/beta-catenin signaling in adult mammalian epithelial stem cells. Dev. Biol. 2017, 428, 273–282. [Google Scholar] [CrossRef]
- Rabbani, P.; Takeo, M.; Chou, W.; Myung, P.; Bosenberg, M.; Chin, L.; Taketo, M.M.; Ito, M. Coordinated activation of Wnt in epithelial and melanocyte stem cells initiates pigmented hair regeneration. Cell 2011, 145, 941–955. [Google Scholar] [CrossRef] [Green Version]
- Sato, N.; Meijer, L.; Skaltsounis, L.; Greengard, P.; Brivanlou, A.H. Maintenance of pluripotency in human and mouse embryonic stem cells through activation of Wnt signaling by a pharmacological GSK-3-specific inhibitor. Nat. Med. 2004, 10, 55–63. [Google Scholar] [CrossRef]
- Ng, L.F.; Kaur, P.; Bunnag, N.; Suresh, J.; Sung, I.C.H.; Tan, Q.H.; Gruber, J.; Tolwinski, N.S. WNT Signaling in Disease. Cells 2019, 8, 826. [Google Scholar] [CrossRef] [Green Version]
- Lien, W.H.; Polak, L.; Lin, M.; Lay, K.; Zheng, D.; Fuchs, E. In vivo transcriptional governance of hair follicle stem cells by canonical Wnt regulators. Nat. Cell Biol. 2014, 16, 179–190. [Google Scholar] [CrossRef] [Green Version]
- Myung, P.S.; Takeo, M.; Ito, M.; Atit, R.P. Epithelial Wnt ligand secretion is required for adult hair follicle growth and regeneration. J. Investig. Dermatol. 2013, 133, 31–41. [Google Scholar] [CrossRef] [Green Version]
- Regazzetti, C.; Joly, F.; Marty, C.; Rivier, M.; Mehul, B.; Reiniche, P.; Mounier, C.; Rival, Y.; Piwnica, D.; Cavalie, M.; et al. Transcriptional Analysis of Vitiligo Skin Reveals the Alteration of WNT Pathway: A Promising Target for Repigmenting Vitiligo Patients. J. Investig. Dermatol. 2015, 135, 3105–3114. [Google Scholar] [CrossRef] [Green Version]
- Uluckan, O.; Jimenez, M.; Karbach, S.; Jeschke, A.; Grana, O.; Keller, J.; Busse, B.; Croxford, A.L.; Finzel, S.; Koenders, M.; et al. Chronic skin inflammation leads to bone loss by IL-17-mediated inhibition of Wnt signaling in osteoblasts. Sci. Transl. Med. 2016, 8, 330ra37. [Google Scholar] [CrossRef]
- Takeo, M.; Lee, W.; Rabbani, P.; Sun, Q.; Hu, H.; Lim, C.H.; Manga, P.; Ito, M. EdnrB Governs Regenerative Response of Melanocyte Stem Cells by Crosstalk with Wnt Signaling. Cell Rep. 2016, 15, 1291–1302. [Google Scholar] [CrossRef] [Green Version]
- Huang, P.; Yan, R.; Zhang, X.; Wang, L.; Ke, X.; Qu, Y. Activating Wnt/beta-catenin signaling pathway for disease therapy: Challenges and opportunities. Pharmacol. Ther. 2019, 196, 79–90. [Google Scholar] [CrossRef]
- Sun, Z.; Xu, S.; Cai, Q.; Zhou, W.; Jiao, X.; Bao, M.; Yu, X. Wnt/beta-catenin agonist BIO alleviates cisplatin-induced nephrotoxicity without compromising its efficacy of anti-proliferation in ovarian cancer. Life Sci. 2020, 263, 118672. [Google Scholar] [CrossRef]
- Bilir, B.; Kucuk, O.; Moreno, C.S. Wnt signaling blockage inhibits cell proliferation and migration, and induces apoptosis in triple-negative breast cancer cells. J. Transl. Med. 2013, 11, 280. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.C.; Kim, O.H.; Lee, S.K.; Kim, S.J. IWR-1 inhibits epithelial-mesenchymal transition of colorectal cancer cells through suppressing Wnt/beta-catenin signaling as well as survivin expression. Oncotarget 2015, 6, 27146–27159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins-Neves, S.R.; Paiva-Oliveira, D.I.; Fontes-Ribeiro, C.; Bovee, J.; Cleton-Jansen, A.M.; Gomes, C.M.F. IWR-1, a tankyrase inhibitor, attenuates Wnt/beta-catenin signaling in cancer stem-like cells and inhibits in vivo the growth of a subcutaneous human osteosarcoma xenograft. Cancer Lett. 2018, 414, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Luo, F.; Li, J.; Zhong, X.; Liu, K. Tankyrase 1 inhibitior XAV939 increases chemosensitivity in colon cancer cell lines via inhibition of the Wnt signaling pathway. Int. J. Oncol. 2016, 48, 1333–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wang, X. Targeting the Wnt/beta-catenin signaling pathway in cancer. J. Hematol. Oncol. 2020, 13, 165. [Google Scholar] [CrossRef]
- Guo, H.; Xing, Y.; Liu, Y.; Luo, Y.; Deng, F.; Yang, T.; Yang, K.; Li, Y. Wnt/beta-catenin signaling pathway activates melanocyte stem cells in vitro and in vivo. J. Dermatol. Sci. 2016, 83, 45–51. [Google Scholar] [CrossRef]
- Heilmann, S.; Kiefer, A.K.; Fricker, N.; Drichel, D.; Hillmer, A.M.; Herold, C.; Tung, J.Y.; Eriksson, N.; Redler, S.; Betz, R.C.; et al. Androgenetic alopecia: Identification of four genetic risk loci and evidence for the contribution of WNT signaling to its etiology. J. Investig. Dermatol. 2013, 133, 1489–1496. [Google Scholar] [CrossRef] [Green Version]
- Leiros, G.J.; Ceruti, J.M.; Castellanos, M.L.; Kusinsky, A.G.; Balana, M.E. Androgens modify Wnt agonists/antagonists expression balance in dermal papilla cells preventing hair follicle stem cell differentiation in androgenetic alopecia. Mol. Cell Endocrinol. 2017, 439, 26–34. [Google Scholar] [CrossRef]
- Zhao, S.J.; Jia, H.; Xu, X.L.; Bu, W.B.; Zhang, Q.; Chen, X.; Ji, J.; Sun, J.F. Identification of the Role of Wnt/beta-Catenin Pathway Through Integrated Analyses and in vivo Experiments in Vitiligo. Clin. Cosmet. Investig. Dermatol. 2021, 14, 1089–1103. [Google Scholar] [CrossRef]
- Choi, B.Y. Targeting Wnt/beta-Catenin Pathway for Developing Therapies for Hair Loss. Int. J. Mol. Sci. 2020, 21, 4915. [Google Scholar] [CrossRef]
- Niida, A.; Hiroko, T.; Kasai, M.; Furukawa, Y.; Nakamura, Y.; Suzuki, Y.; Sugano, S.; Akiyama, T. DKK1, a negative regulator of Wnt signaling, is a target of the beta-catenin/TCF pathway. Oncogene 2004, 23, 8520–8526. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, D.J.; Ho, J.W.; Mammoto, T.; Turbe-Doan, A.; O’Connell, J.T.; Haseley, P.S.; Koo, S.; Kamiya, N.; Ingber, D.E.; Park, P.J.; et al. A Wnt-bmp feedback circuit controls intertissue signaling dynamics in tooth organogenesis. Sci. Signal. 2012, 5, ra4. [Google Scholar] [CrossRef]
- Rohrs, S.; Kutzner, N.; Vlad, A.; Grunwald, T.; Ziegler, S.; Muller, O. Chronological expression of Wnt target genes Ccnd1, Myc, Cdkn1a, Tfrc, Plf1 and Ramp3. Cell Biol. Int. 2009, 33, 501–508. [Google Scholar] [CrossRef]
- Voorneveld, P.W.; Kodach, L.L.; Jacobs, R.J.; van Noesel, C.J.; Peppelenbosch, M.P.; Korkmaz, K.S.; Molendijk, I.; Dekker, E.; Morreau, H.; van Pelt, G.W.; et al. The BMP pathway either enhances or inhibits the Wnt pathway depending on the SMAD4 and p53 status in CRC. Br. J. Cancer 2015, 112, 122–130. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, H.; Kishida, S.; Kishida, M.; Ikeda, S.; Takada, S.; Kikuchi, A. Phosphorylation of axin, a Wnt signal negative regulator, by glycogen synthase kinase-3beta regulates its stability. J. Biol. Chem. 1999, 274, 10681–10684. [Google Scholar] [CrossRef] [Green Version]
- Nong, J.; Kang, K.; Shi, Q.; Zhu, X.; Tao, Q.; Chen, Y.G. Phase separation of Axin organizes the beta-catenin destruction complex. J. Cell Biol. 2021, 220, e202012112. [Google Scholar] [CrossRef]
- Wu, D.; Pan, W. GSK3: A multifaceted kinase in Wnt signaling. Trends Biochem. Sci. 2010, 35, 161–168. [Google Scholar] [CrossRef] [Green Version]
- Griebel, G.; Stemmelin, J.; Lopez-Grancha, M.; Boulay, D.; Boquet, G.; Slowinski, F.; Pichat, P.; Beeske, S.; Tanaka, S.; Mori, A.; et al. The selective GSK3 inhibitor, SAR502250, displays neuroprotective activity and attenuates behavioral impairments in models of neuropsychiatric symptoms of Alzheimer’s disease in rodents. Sci. Rep. 2019, 9, 18045. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.M.; Mishina, Y.M.; Liu, S.; Cheung, A.; Stegmeier, F.; Michaud, G.A.; Charlat, O.; Wiellette, E.; Zhang, Y.; Wiessner, S.; et al. Tankyrase inhibition stabilizes axin and antagonizes Wnt signalling. Nature 2009, 461, 614–620. [Google Scholar] [CrossRef]
- Mao, J.; Wang, J.; Liu, B.; Pan, W.; Farr, G.H., 3rd; Flynn, C.; Yuan, H.; Takada, S.; Kimelman, D.; Li, L.; et al. Low-density lipoprotein receptor-related protein-5 binds to Axin and regulates the canonical Wnt signaling pathway. Mol. Cell 2001, 7, 801–809. [Google Scholar] [CrossRef]
- Ranes, M.; Zaleska, M.; Sakalas, S.; Knight, R.; Guettler, S. Reconstitution of the destruction complex defines roles of AXIN polymers and APC in beta-catenin capture, phosphorylation, and ubiquitylation. Mol. Cell 2021, 81, 3246–3261.e11. [Google Scholar] [CrossRef]
- Thorvaldsen, T.E. Targeting Tankyrase to Fight WNT-dependent Tumours. Basic Clin. Pharmacol. Toxicol. 2017, 121, 81–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariotti, L.; Pollock, K.; Guettler, S. Regulation of Wnt/beta-catenin signalling by tankyrase-dependent poly(ADP-ribosyl)ation and scaffolding. Br. J. Pharmacol. 2017, 174, 4611–4636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, Q.; Chen, J.; Liu, Y. LRP5 and LRP6 in Wnt Signaling: Similarity and Divergence. Front. Cell Dev. Biol. 2021, 9, 670960. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Ringholm, M.; Bast, R.; Ruud, K.; Thorvaldsen, A.J.; Jaszunski, M. Analytic density functional theory calculations of pure vibrational hyperpolarizabilities: The first dipole hyperpolarizability of retinal and related molecules. J. Phys. Chem. A 2014, 118, 748–756. [Google Scholar] [CrossRef] [Green Version]
- Sucre, J.M.; Vijayaraj, P.; Aros, C.J.; Wilkinson, D.; Paul, M.; Dunn, B.; Guttentag, S.H.; Gomperts, B.N. Posttranslational modification of beta-catenin is associated with pathogenic fibroblastic changes in bronchopulmonary dysplasia. Am. J. Physiol. Lung Cell Mol. Physiol. 2017, 312, L186–L195. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, J.Y.; Kim, J.; Choi, Y.-H.; Lee, S.; Kang, N.-G. Escin Activates Canonical Wnt/β-Catenin Signaling Pathway by Facilitating the Proteasomal Degradation of Glycogen Synthase Kinase-3β in Cultured Human Dermal Papilla Cells. Curr. Issues Mol. Biol. 2023, 45, 5902-5913. https://doi.org/10.3390/cimb45070373
Shin JY, Kim J, Choi Y-H, Lee S, Kang N-G. Escin Activates Canonical Wnt/β-Catenin Signaling Pathway by Facilitating the Proteasomal Degradation of Glycogen Synthase Kinase-3β in Cultured Human Dermal Papilla Cells. Current Issues in Molecular Biology. 2023; 45(7):5902-5913. https://doi.org/10.3390/cimb45070373
Chicago/Turabian StyleShin, Jae Young, Jaeyoon Kim, Yun-Ho Choi, Sanghwa Lee, and Nae-Gyu Kang. 2023. "Escin Activates Canonical Wnt/β-Catenin Signaling Pathway by Facilitating the Proteasomal Degradation of Glycogen Synthase Kinase-3β in Cultured Human Dermal Papilla Cells" Current Issues in Molecular Biology 45, no. 7: 5902-5913. https://doi.org/10.3390/cimb45070373
APA StyleShin, J. Y., Kim, J., Choi, Y. -H., Lee, S., & Kang, N. -G. (2023). Escin Activates Canonical Wnt/β-Catenin Signaling Pathway by Facilitating the Proteasomal Degradation of Glycogen Synthase Kinase-3β in Cultured Human Dermal Papilla Cells. Current Issues in Molecular Biology, 45(7), 5902-5913. https://doi.org/10.3390/cimb45070373