
Anti-Inflammatory Effects of Serotonin Receptor and Transient Receptor Potential Channel Ligands in Human Small Intestinal Epithelial Cells

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Maintenance
2.2. Methyl Thiazole Tetrazolium (MTT) Cytotoxicity Assay
2.3. Protein Extraction and Quantification
2.4. Western Immunoblotting
2.5. Statistical Analysis
3. Results
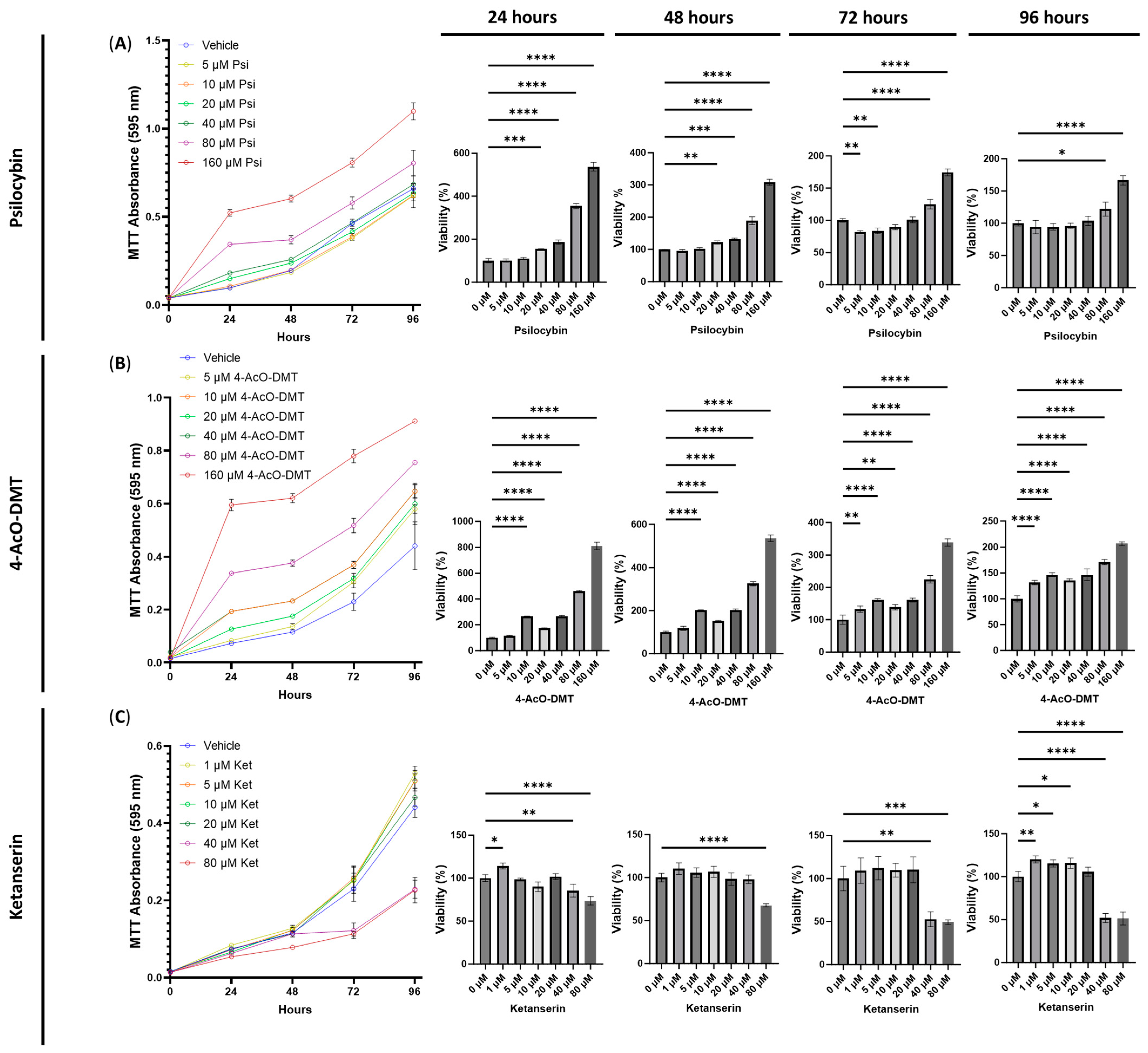
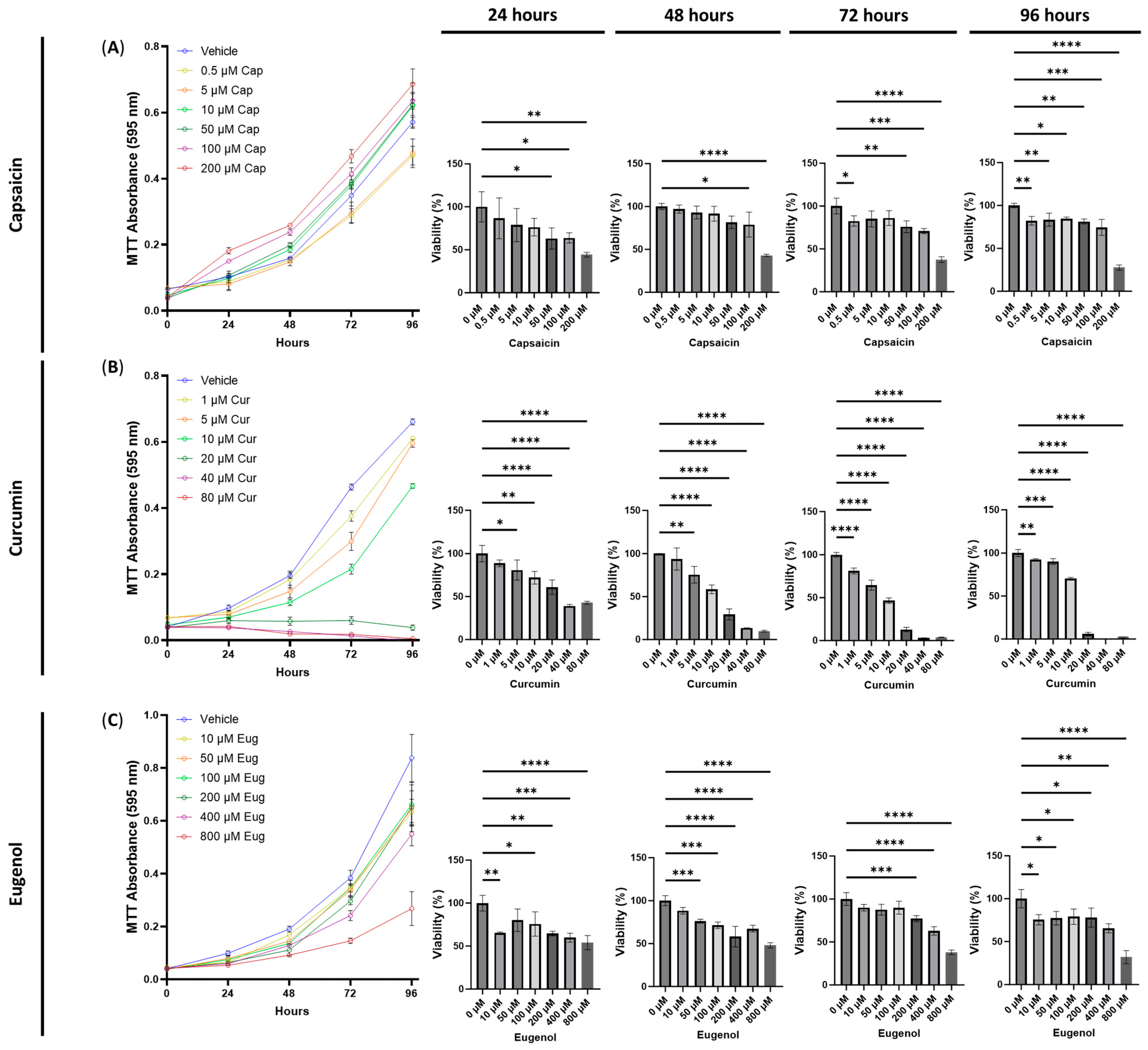
3.1. The Cytotoxic Effects of Serotonin Receptor and Transient Receptor Potential Channel Ligands
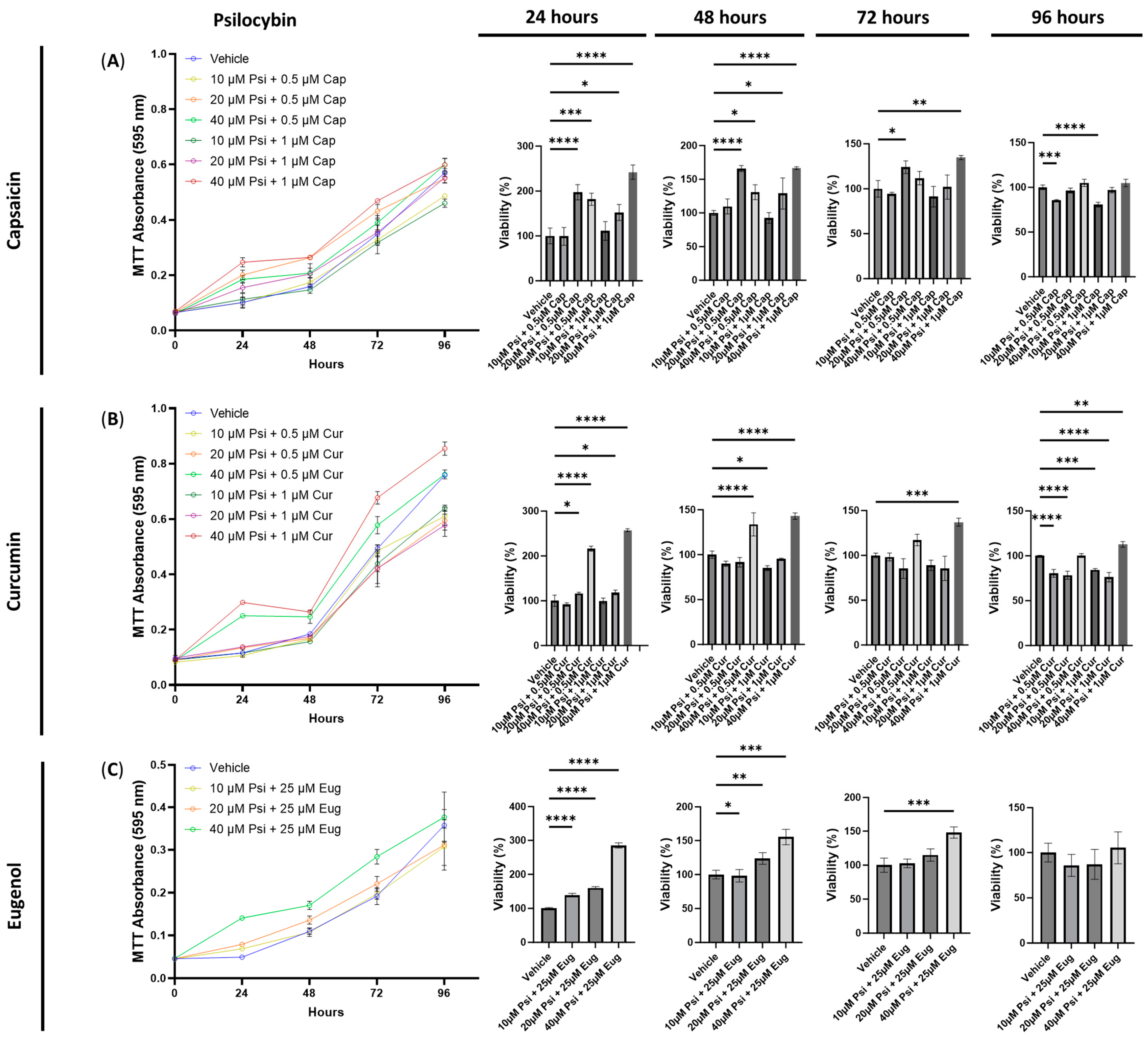
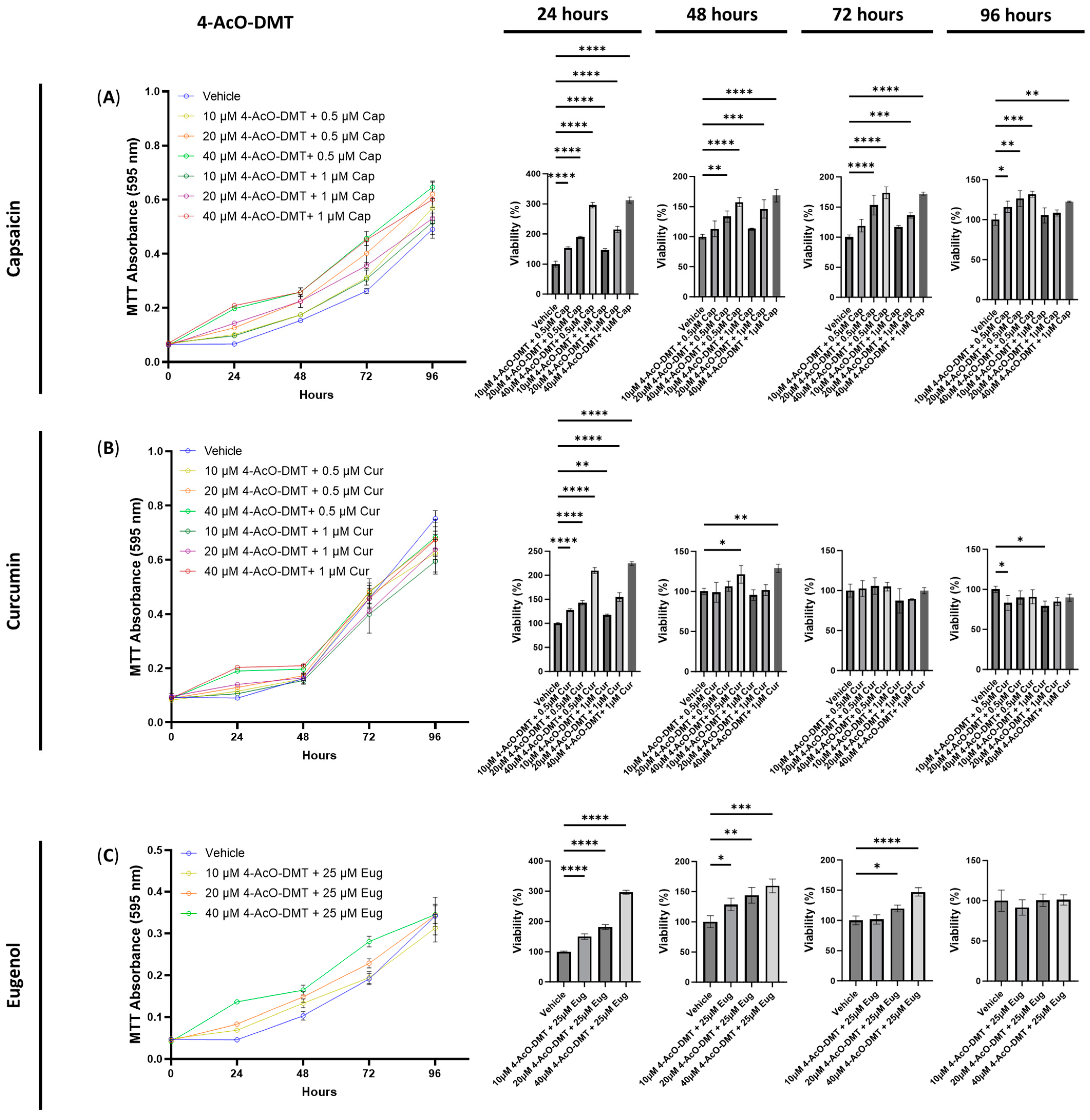
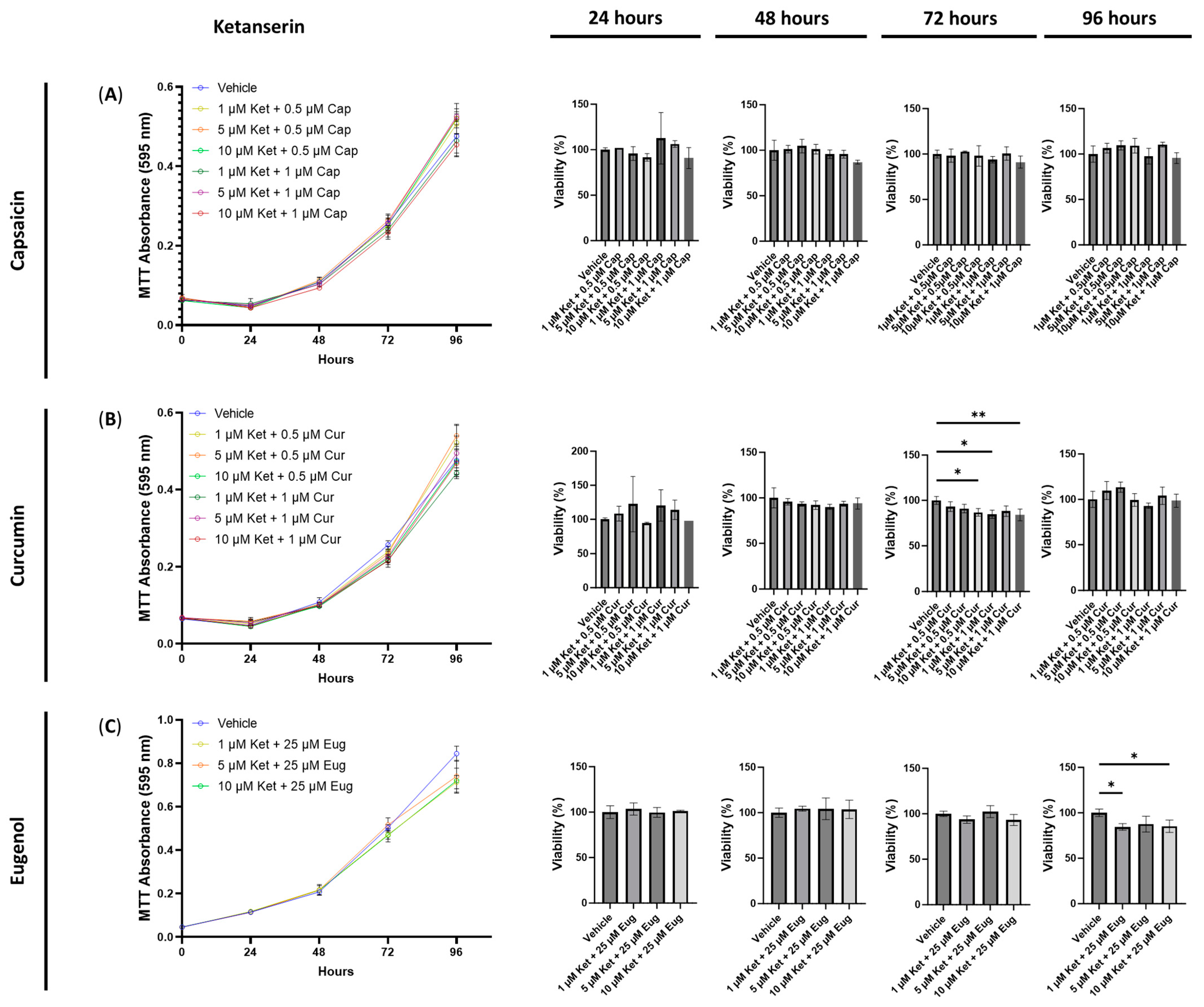
3.2. The Cytotoxic Effects of Combined Treatments of Serotonin Receptor and Transient Receptor Potential Channel Ligands
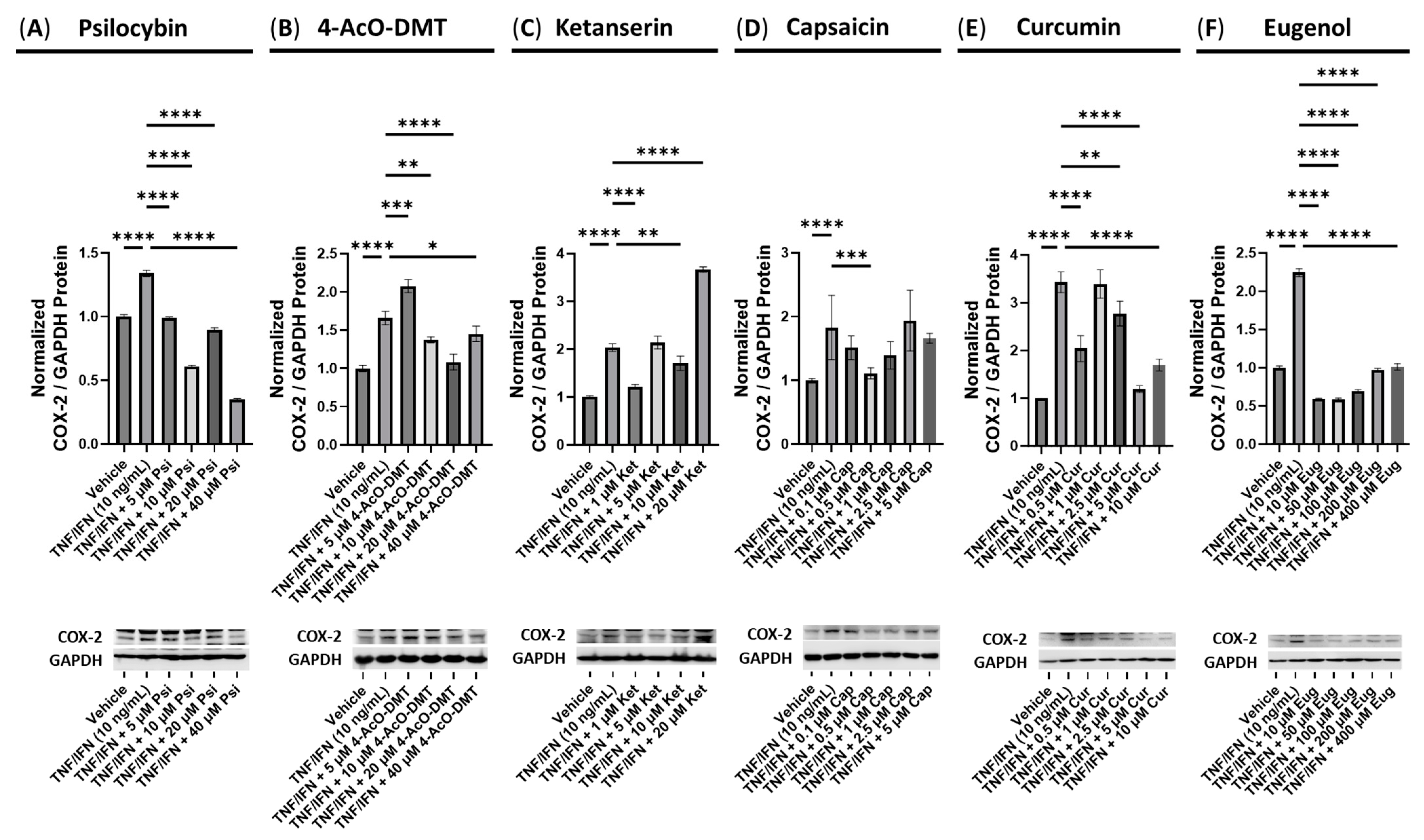
3.3. The Anti-Inflammatory Effects of the Serotonin Receptor or Transient Receptor Potential Channel Ligands
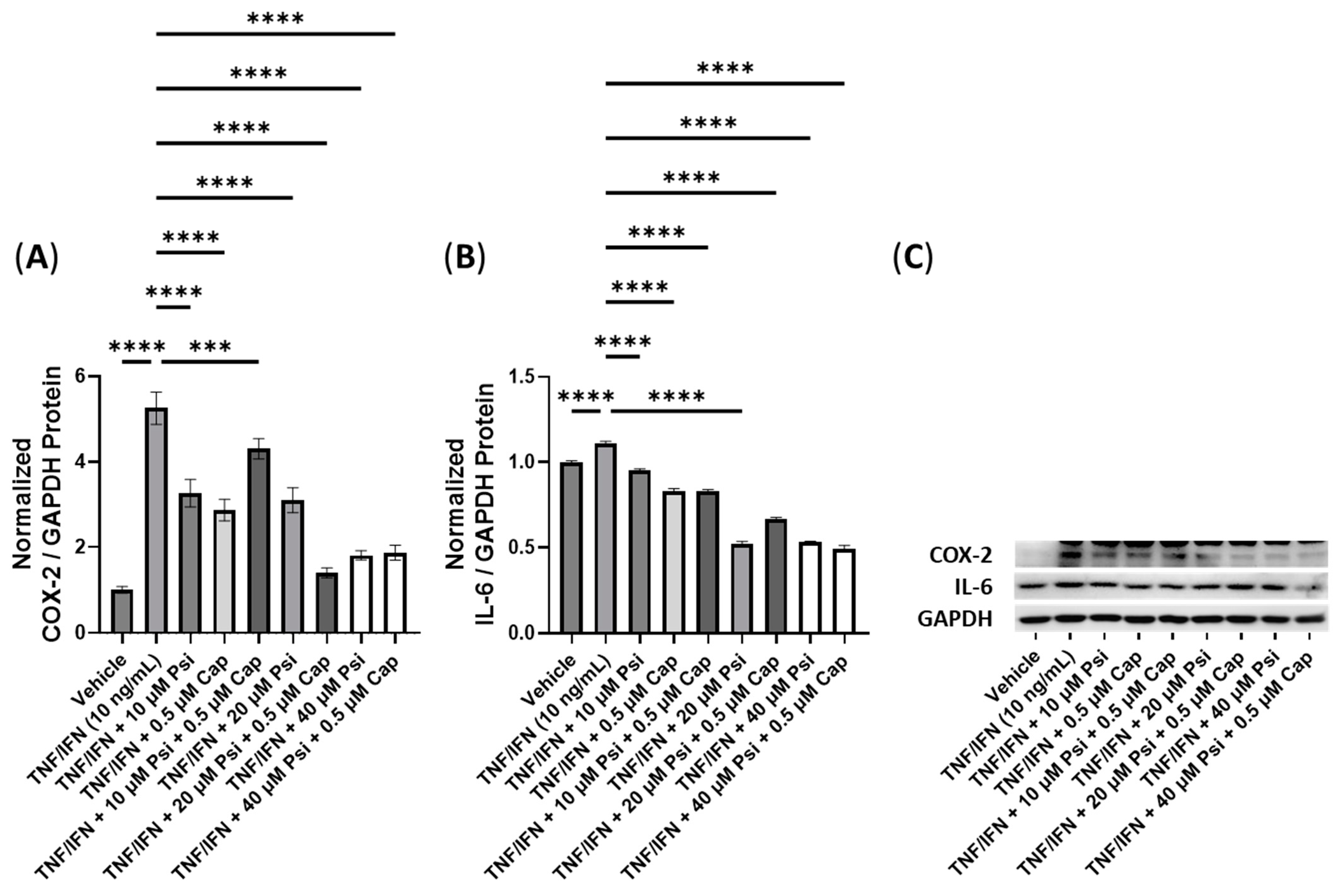
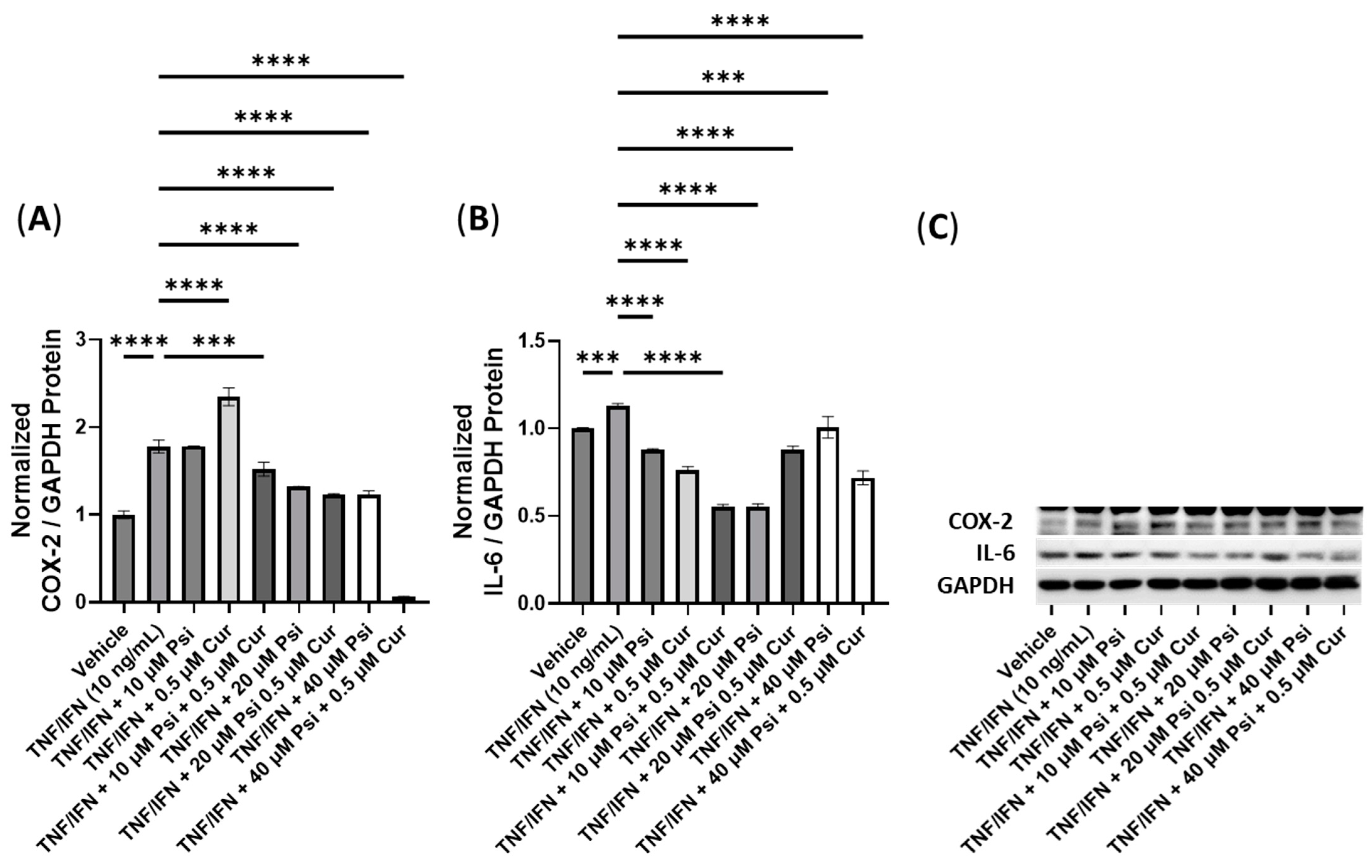
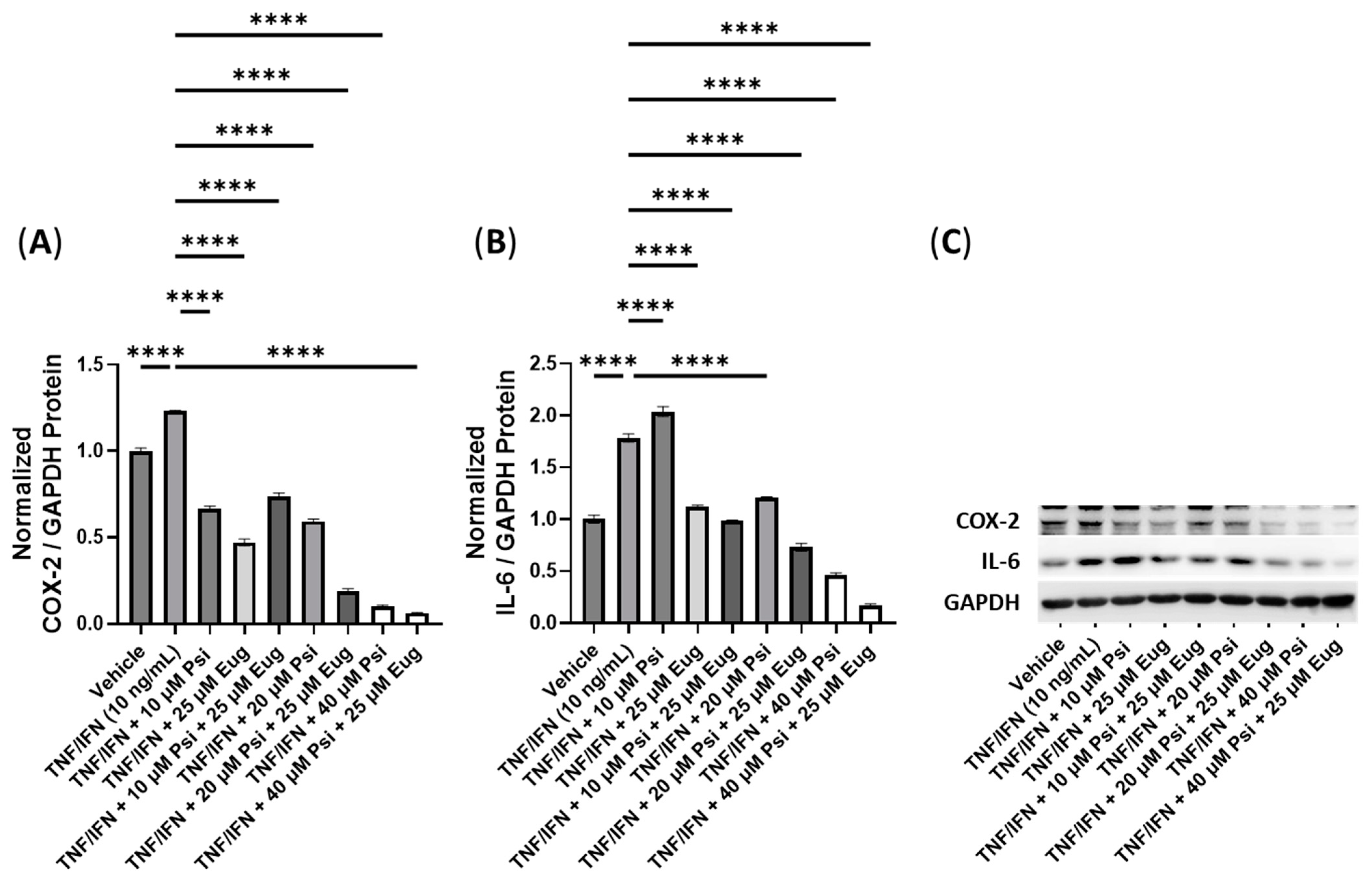
3.4. The Anti-Inflammatory Effects of Psilocybin and Transient Receptor Potential Channel Ligands
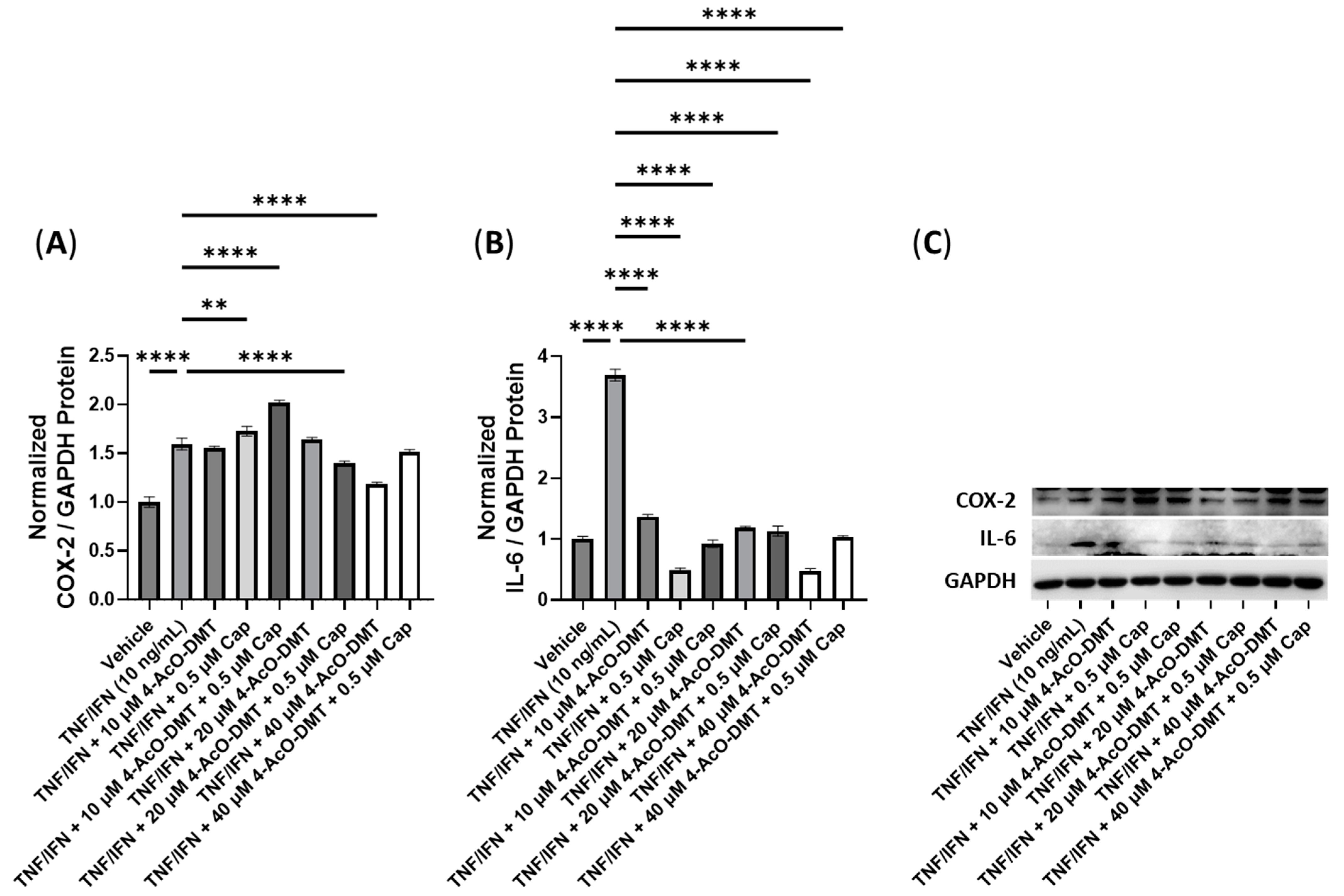
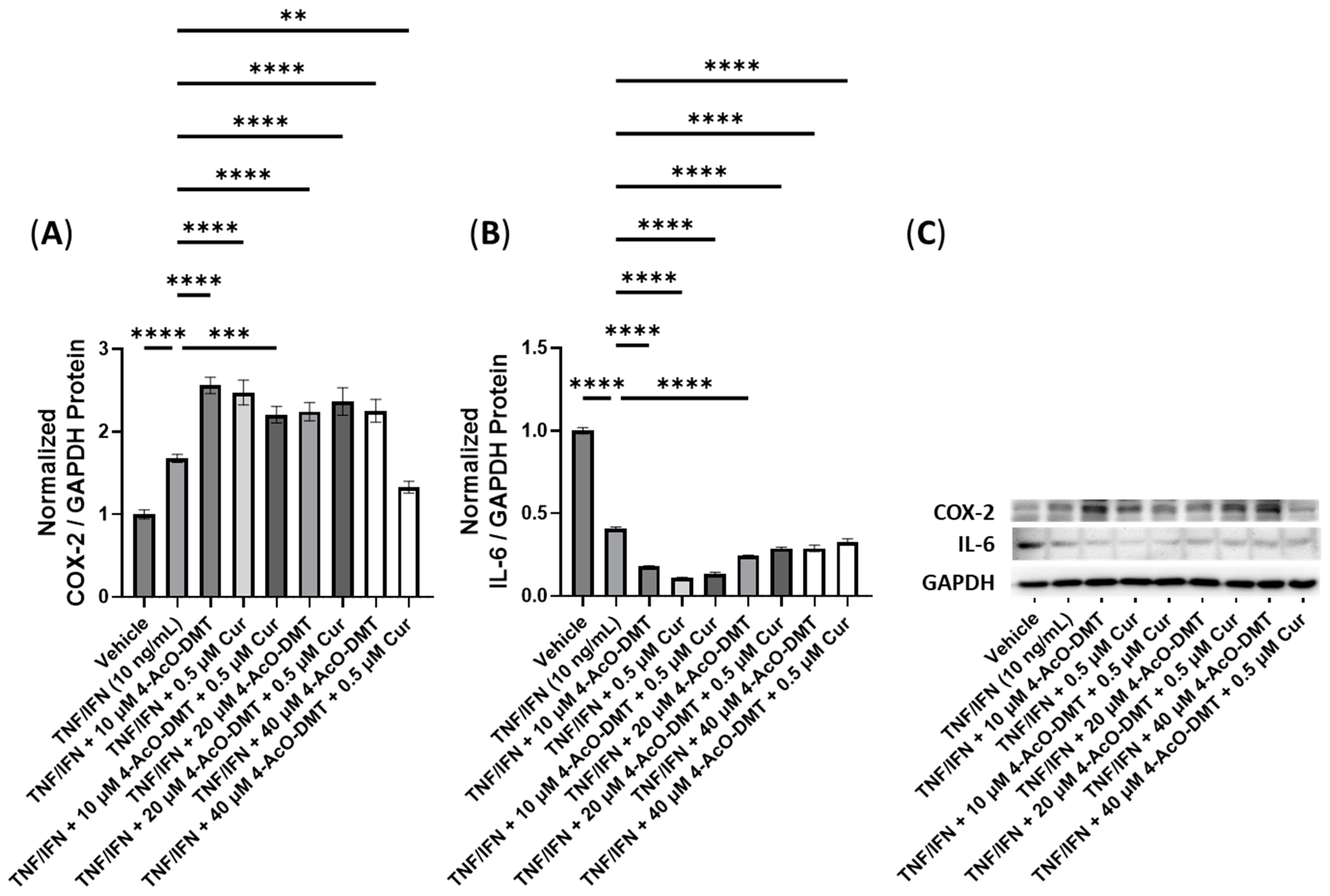
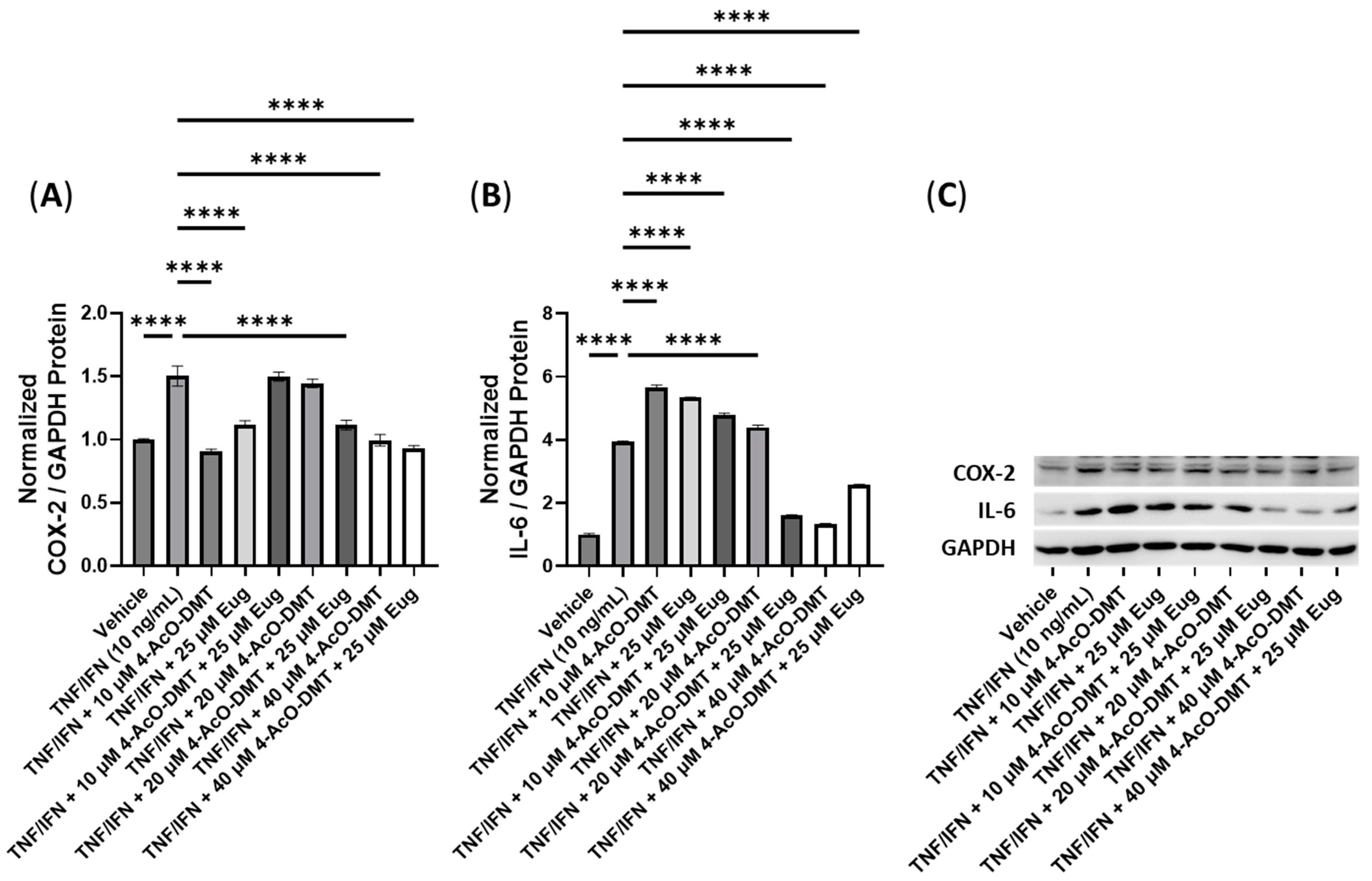
3.5. Anti-Inflammatory Effects of 4-AcO-DMT and Transient Receptor Potential Channel Ligands
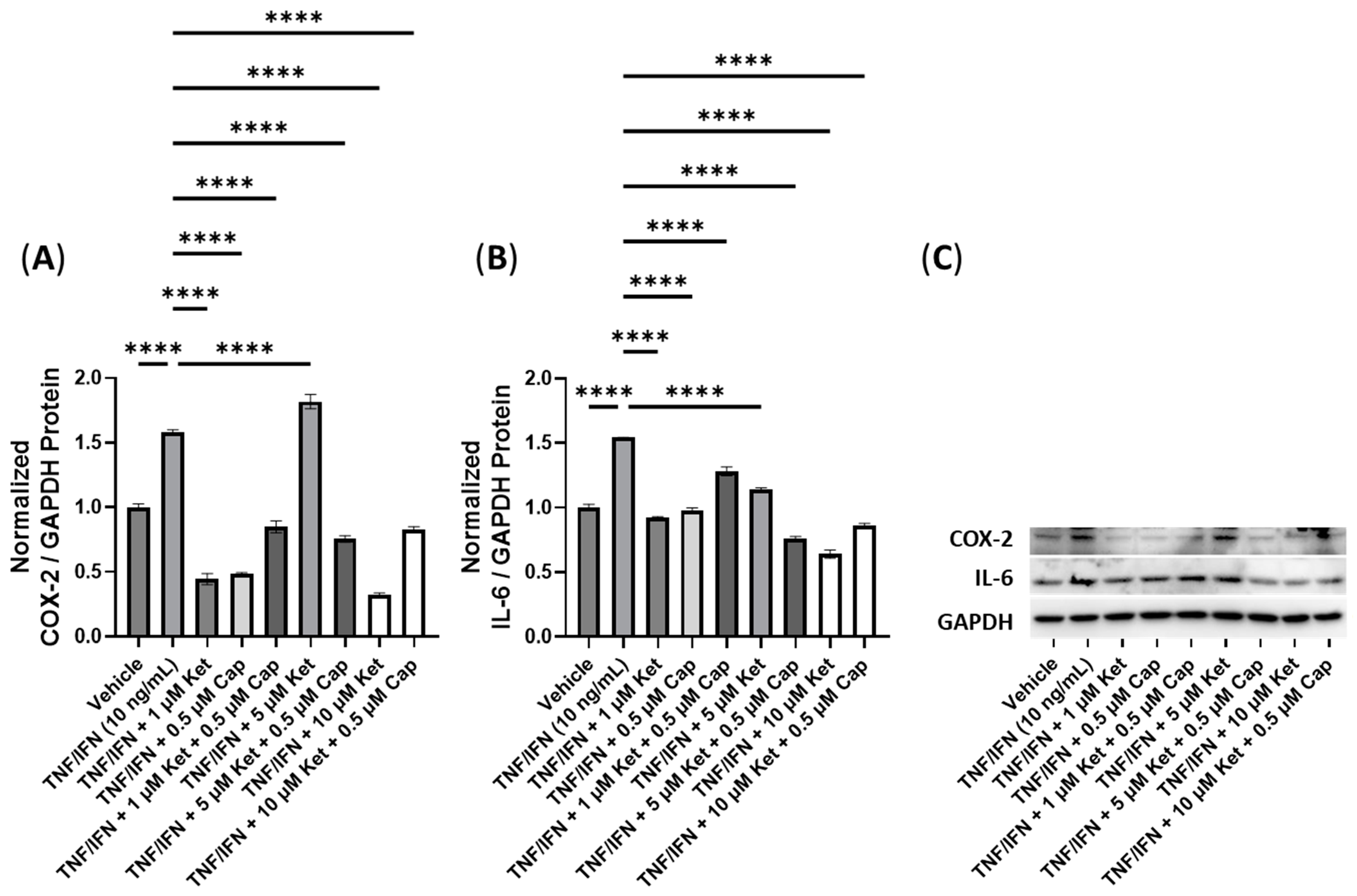
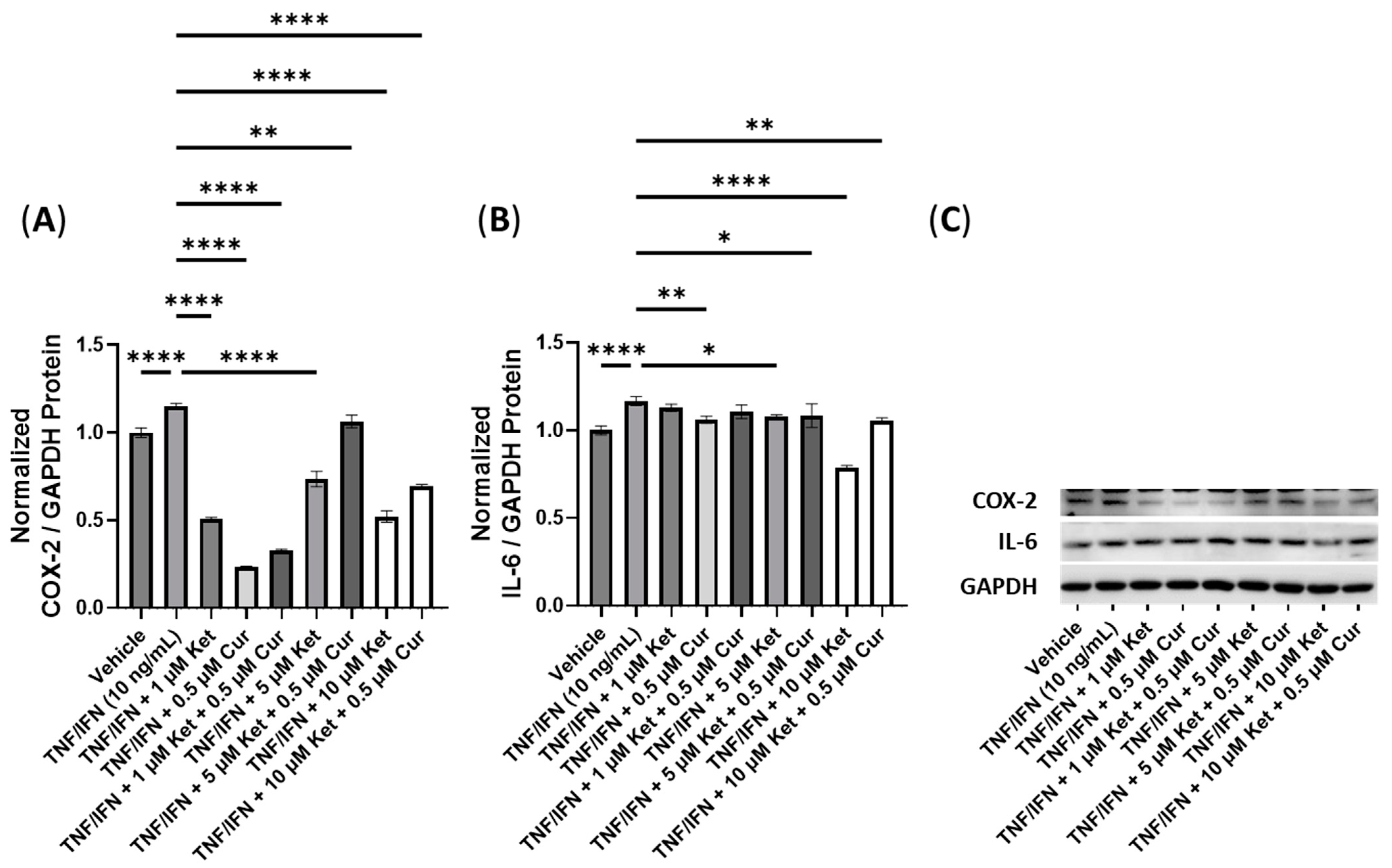
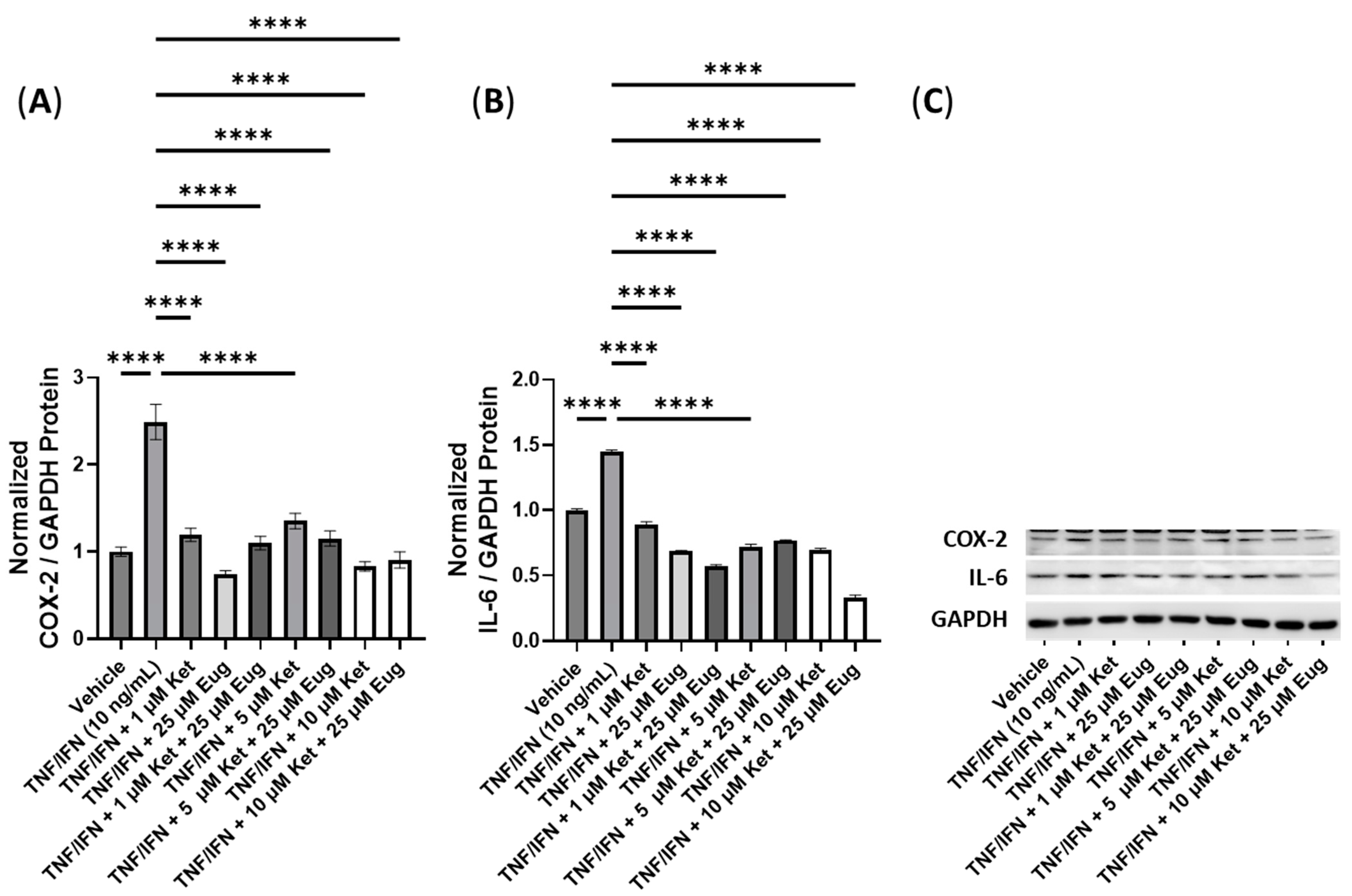
3.6. Anti-Inflammatory Effects of Ketanserin and Transient Receptor Potential Channel Ligands
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abdulla, M.; Mohammed, N. A Review on Inflammatory Bowel Diseases: Recent Molecular Pathophysiology Advances. Biologics 2022, 16, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Li, Z.; Liu, S.; Zhang, D. Global, regional and national burden of inflammatory bowel disease in 204 countries and territories from 1990 to 2019: A systematic analysis based on the Global Burden of Disease Study 2019. BMJ Open 2023, 13, e065186. [Google Scholar] [CrossRef] [PubMed]
- Mak, W.Y.; Zhao, M.; Ng, S.C.; Burisch, J. The epidemiology of inflammatory bowel disease: East meets west. J. Gastroenterol. Hepatol. 2020, 35, 380–389. [Google Scholar] [CrossRef]
- Cohen, N.A.; Rubin, D.T. New Targets in Inflammatory Bowel Disease Therapy: 2021. Curr. Opin. Gastroenterol. 2021, 37, 357–363. [Google Scholar] [CrossRef]
- Flanagan, T.W.; Nichols, C.D. Psychedelics as anti-inflammatory agents. Int. Rev. Psychiatry 2018, 30, 363–375. [Google Scholar] [CrossRef] [PubMed]
- McKenna, D.J.; Repke, D.B.; Lo, L.; Peroutka, S.J. Differential interactions of indolealkylamines with 5-hydroxytryptamine receptor subtypes. Neuropharmacology 1990, 29, 193–198. [Google Scholar] [CrossRef]
- Nichols, D.; Johnson, M.; Nichols, C. Psychedelics as Medicines: An Emerging New Paradigm. Clin. Pharmacol. Ther. 2017, 101, 209–219. [Google Scholar] [CrossRef]
- Kalkman, H.O. Inhibition of Microglial GSK3β Activity Is Common to Different Kinds of Antidepressants: A Proposal for an In Vitro Screen to Detect Novel Antidepressant Principles. Biomedicines 2023, 11, 806. [Google Scholar] [CrossRef]
- Zanikov, T.; Gerasymchuk, M.; Ghasemi Gojani, E.; Robinson, G.I.; Aghari, S.; Groves, A.; Haselhorst, A.; Nandakumar, S.; Stahl, C.; Cameron, M.; et al. The Effect of Combined Treatment of Psilocybin and Eugenol on Lipopolysaccharide-Induced Brain Inflammation in Mice. Molecules 2023, 28, 2624. [Google Scholar] [CrossRef]
- Klein, A.K.; Chatha, M.; Laskowski, L.J.; Anderson, E.I.; Brandt, S.D.; Chapman, S.J.; McCorvy, J.D.; Halberstadt, A.L. Investigation of the Structure-Activity Relationships of Psilocybin Analogues. ACS Pharmacol. Transl. Sci. 2021, 4, 533–542. [Google Scholar] [CrossRef]
- Rapalli, A.; Bertoni, S.; Arcaro, V.; Saccani, F.; Grandi, A.; Vivo, V.; Cartoni, A.M.; Barocelli, E. Dual Role of Endogenous Serotonin in 2,4,6-Trinitrobenzene Sulfonic Acid-Induced Colitis. Front. Pharmacol. 2016, 7, 68. [Google Scholar] [CrossRef]
- Xiao, J.; Shao, L.; Shen, J.; Jiang, W.; Feng, Y.; Zheng, P.; Liu, F. Effects of ketanserin on experimental colitis in mice and macrophage function. Int. J. Mol. Med. 2016, 37, 659–668. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zheng, Y.; Luo, Y.; Du, Y.; Zhang, X.; Fu, J. Curcumin inhibits LPS-induced neuroinflammation by promoting microglial M2 polarization via TREM2/TLR4/NF-κB pathways in BV2 cells. Mol. Immunol. 2019, 116, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, A.; Young, K.N.; Moniruzzaman, M.; Beyenne, A.M.; Do, K.; Kalaiselvi, S.; Min, T. Curcumin and Its Modified Formulations on Inflammatory Bowel Disease (IBD): The Story So Far and Future Outlook. Pharmaceutics 2021, 13, 484. [Google Scholar] [CrossRef] [PubMed]
- Pituch-Zdanowska, A.; Dembiński, Ł.; Banaszkiewicz, A. Old but Fancy: Curcumin in Ulcerative Colitis—Current Overview. Nutrients 2022, 14, 5249. [Google Scholar] [CrossRef]
- EFSA Panel on Food Additives and Nutrient Sources added to Food (ANS). Scientific Opinion on the re-evaluation of curcumin (E 100) as a food additive. EFSA J. 2010, 8, 1679. [Google Scholar] [CrossRef]
- Hui, Q.; Ammeter, E.; Liu, S.; Yang, R.; Lu, P.; Lahaye, L.; Yang, C. Eugenol attenuates inflammatory response and enhances barrier function during lipopolysaccharide-induced inflammation in the porcine intestinal epithelial cells. J. Anim. Sci. 2020, 98, skaa245. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Tang, Y.; Wu, X.; Liang, H.; Chen, D.; Yu, B.; He, H.; Mao, X.; Huang, Z.; Yan, H.; et al. Eugenol Attenuates Transmissible Gastroenteritis Virus-Induced Oxidative Stress and Apoptosis Via ROS-NRF2-ARE Signaling. Antioxidants 2022, 11, 1838. [Google Scholar] [CrossRef]
- Truzzi, F.; Whittaker, A.; D’Amen, E.; Valerii, M.C.; Abduazizova, V.; Spisni, E.; Dinelli, G. Spermidine-Eugenol Supplement Preserved Inflammation-Challenged Intestinal Cells by Stimulating Autophagy. Int. J. Mol. Sci. 2023, 24, 4131. [Google Scholar] [CrossRef]
- Rosca, A.E.; Iesanu, M.I.; Zahiu, C.D.M.; Voiculescu, S.E.; Paslaru, A.C.; Zagrean, A.M. Capsaicin and Gut Microbiota in Health and Disease. Molecules 2020, 25, 5681. [Google Scholar] [CrossRef]
- Zhang, Q.; Luo, P.; Xia, F.; Tang, H.; Chen, J.; Zhang, J.; Liu, D.; Zhu, Y.; Liu, Y.; Gu, L.; et al. Capsaicin ameliorates inflammation in a TRPV1-independent mechanism by inhibiting PKM2-LDHA-mediated Warburg effect in sepsis. Cell Chem. Biol. 2022, 29, 1248–1259.e6. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, E.A.; Alvarez-Leite, J.I. Capsaicin: A Potential Therapy Adjuvant for Intestinal Bowel Disease. JDDD 2019, 2, 8–16. [Google Scholar] [CrossRef]
- Kang, C.; Wang, B.; Kaliannan, K. Gut Microbiota Mediates the Protective Effects of Dietary Capsaicin against Chronic Low-Grade Inflammation and Associated Obesity Induced by High-Fat Diet. mBio 2017, 8, e00470-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berg, K.A.; Maayani, S.; Goldfarb, J.; Clarke, W.P. Pleiotropic behavior of 5-HT2A and 5-HT2C receptor agonists. Ann. N. Y. Acad. Sci. 1998, 861, 104–110. [Google Scholar] [CrossRef]
- Kovalchuk, A.; Wang, B.; Li, D.; Rodriguez-Juarez, R.; Ilnytskyy, S.; Kovalchuk, I.; Kovalchuk, O. Fighting the storm: Could novel anti-TNFα and anti-IL-6 C. sativa cultivars tame cytokine storm in COVID-19? Aging 2021, 13, 1571–1590. [Google Scholar] [CrossRef]
- Woznicki, J.A.; Saini, N.; Flood, P.; Rajaram, S.; Lee, C.M.; Stamou, P.; Skowyra, A.; Bustamante-Garrido, M.; Regazzoni, K.; Crawford, N.; et al. TNF-α synergises with IFN-γ to induce caspase-8-JAK1/2-STAT1-dependent death of intestinal epithelial cells. Cell Death Dis. 2021, 12, 864. [Google Scholar] [CrossRef] [PubMed]
- Chen, C. COX-2’s new role in inflammation. Nat. Chem. Biol. 2010, 6, 401–402. [Google Scholar] [CrossRef]
- Ly, C.; Greb, A.C.; Cameron, L.P.; Wong, J.M.; Barragan, E.V.; Wilson, P.C.; Burbach, K.F.; Zarandi, S.S.; Sood, A.; Paddy, M.R.; et al. Psychedelics Promote Structural and Functional Neural Plasticity. Cell Rep. 2018, 23, 3170–3182. [Google Scholar] [CrossRef]
- Ly, C.; Greb, A.C.; Vargas, M.V.; Duim, W.C.; Grodzki, A.C.G.; Lein, P.J.; Olsen, D.E. Transient Stimulation with Psychoplastogens Is Sufficient to Initiate Neuronal Growth. ACS Pharmacol. Transl. Sci. 2020, 4, 452–460. [Google Scholar] [CrossRef]
- Lima da Cruz, R.V.; Moulin, T.C.; Petiz, L.L.; Leão, R.N. A Single Dose of 5-MeO-DMT Stimulates Cell Proliferation, Neuronal Survivability, Morphological and Functional Changes in Adult Mice Ventral Dentate Gyrus. Front. Mol. Neurosci. 2018, 11, 312. [Google Scholar] [CrossRef]
- Rodriguiz, R.M.; Nadkarni, V.; Means, C.R.; Pogorelov, V.M.; Chiu, Y.T.; Roth, B.L.; Wetsel, W. LSD-stimulated behaviors in mice require β-arrestin 2 but not β-arrestin 1. Sci. Rep. 2021, 11, 17690. [Google Scholar] [CrossRef]
- Sharma, D.; Parameswaran, N. Multifaceted role of β-arrestins in inflammation and disease. Genes Immun. 2015, 16, 499–513. [Google Scholar] [CrossRef] [Green Version]
- Cheshmehkani, A.; Senatorov, I.S.; Dhuguru, J.; Ghoneim, O.; Moniri, N.H. Free-fatty acid receptor-4 (FFA4) modulates ROS generation and COX-2 expression via the C-terminal β-arrestin phosphosensor in Raw 264.7 macrophages. Biochem. Pharmacol. 2017, 146, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, K.; Okamoto, S.; Mukaida, N.; Murakami, S.; Mai, M.; Matsushima, K. Tumor necrosis factor alpha and interferon gamma synergistically induce interleukin 8 production in a human gastric cancer cell line through acting concurrently on AP-1 and NF-kB-like binding sites of the interleukin 8 gene. J. Biol. Chem. 1992, 267, 22506–22511. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Krishan, P. Serotonin 5HT2A receptor antagonism mediated anti-inflammatory and anti-fibrotic effect in adriamycin-induced CKD in rats. Naunyn Schmiedebergs Arch. Pharmacol. 2020, 393, 1269–1279. [Google Scholar] [CrossRef]
- Nkadimeng, S.M.; Nabatanzi, A.; Steinmann, C.M.L.; Eloff, J.N. Phytochemical, Cytotoxicity, Antioxidant and Anti-Inflammatory Effects of Psilocybe Natalensis Magic Mushroom. Plants 2020, 9, 1127. [Google Scholar] [CrossRef] [PubMed]
- Nkadimeng, S.M.; Steinmann, C.M.L.; Eloff, J.N. Anti-Inflammatory Effects of Four Psilocybin-Containing Magic Mushroom Water Extracts in vitro on 15-Lipoxygenase Activity and on Lipopolysaccharide-Induced Cyclooxygenase-2 and Inflammatory Cytokines in Human U937 Macrophage Cells. J. Inflamm. Res. 2021, 14, 3729–3738. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wu, X.; Tang, S.; Yin, J.; Song, Z.; He, X.; Yin, Y. Eugenol Alleviates Dextran Sulfate Sodium-Induced Colitis Independent of Intestinal Microbiota in Mice. J. Agric. Food Chem. 2021, 69, 10506–10514. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Chen, X.; Chen, D.; Yu, B.; He, J.; Yan, H.; Luo, Y.; Zheng, P.; Chen, H.; Huang, Z. Eugenol promotes appetite through TRP channels mediated-CaMKK2/AMPK signaling pathway. Phytother. Res. 2023, 37, 2759–2770. [Google Scholar] [CrossRef]
- Racioppi, L.; Nelson, E.R.; Huang, W.; Mukherjee, D.; Lawrence, S.A.; Lento, W.; Masci, A.M.; Jiao, Y.; Park, S.; York, B.; et al. CaMKK2 in myeloid cells is a key regulator of the immune-suppressive microenvironment in breast cancer. Nat. Commun. 2019, 10, 2450. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Ao, M.; Dong, B.; Jiang, Y.; Yu, L.; Chen, Z.; Hu, C.; Xu, R. Anti-Inflammatory Effects of Curcumin in the Inflammatory Diseases: Status, Limitations and Countermeasures. Drug Des. Dev. Ther. 2021, 15, 4503–4525. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Mu, J.; Zhu, M.; Mukherjee, A.; Zhang, H. Transient Receptor Potential Channels and Inflammatory Bowel Disease. Front. Immunol. 2020, 11, 180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azimirad, M.; Noori, M.; Azimirad, F.; Gholami, F.; Naseri, K.; Yadegar, A.; Aghdaei, H.A.; Zali, M.R. Curcumin and capsaicin regulate apoptosis and alleviate intestinal inflammation induced by Clostridioides difficile in vitro. Ann. Clin. Microbiol. Antimicrob. 2022, 21, 41. [Google Scholar] [CrossRef]
- Zhao, X.; Dong, B.; Friesen, M.; Liu, S.; Zhu, C.; Yang, C. Capsaicin Attenuates Lipopolysaccharide-Induced Inflammation and Barrier Dysfunction in Intestinal Porcine Epithelial Cell Line-J2. Front. Physiol. 2021, 12, 715469. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M.S.; Ghosh, S. Regulation of NF-κB by TNF Family Cytokines. Semin. Immunol. 2014, 26, 253–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kihara, N.; de la Fuente, S.G.; Fujino, K.; Takahashi, T.; Pappas, T.N.; Mantyh, C.R. Vanilloid receptor-1 containing primary sensory neurones mediate dextran sulphate sodium induced colitis in rats. Gut 2003, 52, 713–719. [Google Scholar] [CrossRef]
- Chen, K.; Shen, S.; Chen, Y.; Jiang, M.; Hu, K.; Zou, Y.; Li, L.; Zeng, Z.; Ma, C.; Dang, Y.; et al. A proteomic and RNA-seq transcriptomic dataset of capsaicin-aggravated mouse chronic colitis model. Sci. Data 2022, 9, 549. [Google Scholar] [CrossRef]
- Panpetch, W.; Visitchanakun, P.; Saisorn, W.; Sawatpanich, A.; Chatthanathon, P.; Somboonna, N.; Tumwasorn, S.; Leelahavanichkul, A. Lactobacillus rhamnosus attenuates Thai chili extracts induced gut inflammation and dysbiosis despite capsaicin bactericidal effect against the probiotics, a possible toxicity of high dose capsaicin. PLoS ONE 2021, 16, e0261189. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, B.; Wang, T.; Gao, L.; Yang, Z.J.; Wang, F.F.; Shang, H.W.; Hua, R.H.; Xu, J.D. Biological characteristics of IL-6 and related intestinal diseases. Int. J. Biol. Sci. 2021, 17, 204–219. [Google Scholar] [CrossRef]
- Aliyu, M.; Zohora, F.T.; Anka, A.U.; Ali, K.; Maleknia, S.; Saffarioun, M.; Azizi, G. Interleukin-6 cytokine: An overview of the immune regulation, immune dysregulation, and therapeutic approach. Int. Immunopharmacol. 2022, 111, 109130. [Google Scholar] [CrossRef]
- Rose-John, S.; Jenkins, B.J.; Garbers, C.; Moll, J.M.; Scheller, J. Targeting IL-6 trans-signalling: Past, present and future prospects. Nat. Rev. Immunol. 2023, 1–16. [Google Scholar] [CrossRef]
- Cao, D.; Yu, J.; Wang, H.; Luo, Z.; Liu, X.; He, L.; Qi, J.; Fan, L.; Tang, L.; Chen, Z.; et al. Structure-based discovery of nonhallucinogenic psychedelic analogs. Science 2022, 375, 403–411. [Google Scholar] [CrossRef]
- Osman, S. A mind-changing approach to the therapeutic use of psychedelics. Nat. Struct. Mol. Biol. 2022, 29, 189. [Google Scholar] [CrossRef]
- Liu, C.; Zhang, X.; Zhou, J.X.; Wei, W.; Liu, D.H.; Ke, P.; Zhang, G.F.; Cai, G.J.; Su, D.F. The protective action of ketanserin against lipopolysaccharide-induced shock in mice is mediated by inhibiting inducible NO synthase expression via the MEK/ERK pathway. Free Radic. Biol. Med. 2013, 65, 658–666. [Google Scholar] [CrossRef]
- Pang, Q.; Jin, H.; Ke, X.; Man, Z.; Wang, Y.; Tan, Y.; Lu, Z.; Liu, H. The Role of Serotonin in Concanavalin A-Induced Liver Injury in Mice. Oxid. Med. Cell. Longev. 2020, 2020, 7504521. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, T.K.; Talbot, E.S.; Scherle, P.A.; Ivashkiv, L.B. Rapid inhibition of interleukin-6 signaling and Stat3 activation mediated by mitogen-activated protein kinases. Proc. Natl. Acad. Sci. USA 1998, 95, 11107–11112. [Google Scholar] [CrossRef] [PubMed]
- Seidel, M.F.; Fiebich, B.L.; Ulrich-Merzenich, G.; Candelario-Jalil, E.; Koch, F.W.; Vetter, H. Serotonin mediates PGE2 overexpression through 5-HT2A and 5-HT3 receptor subtypes in serum-free tissue culture of macrophage-like synovial cells. Rheumatol. Int. 2008, 28, 1017–1022. [Google Scholar] [CrossRef] [PubMed]
- Goppelt-Struebe, M.; Hahn, A.; Stroebel, M.; Reiser, C.O. Independent regulation of cyclo-oxygenase 2 expression by p42/44 mitogen-activated protein kinases and Ca2+/calmodulin-dependent kinase. Biochem. J. 1999, 339 Pt 2, 329–334. [Google Scholar] [CrossRef]
- Larsson-Callerfelt, A.K.; Dahlén, S.E.; Kühl, A.R.; Lex, D.; Uhlig, S.; Martin, C. Modulation of antigen-induced responses by serotonin and prostaglandin E2 via EP1 and EP4 receptors in the peripheral rat lung. Eur. J. Pharmacol. 2013, 699, 141–149. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Sun, S.; Wang, P.; Sun, Y.; Hu, Q.; Wang, X. The Mechanism of Secretion and Metabolism of Gut-Derived 5-Hydroxytryptamine. Int. J. Mol. Sci. 2021, 22, 7931. [Google Scholar] [CrossRef]
- Bischoff, S.C.; Barbara, G.; Buurman, W.; Ockhuizen, T.; Schulzke, J.D.; Serino, M.; Tilg, H.; Watson, A.; Wells, J.M. Intestinal permeability--a new target for disease prevention and therapy. BMC Gastroenterol. 2014, 14, 189. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, K.; Lo, M.W.; Hosoya, T.; Tashima, K.; Takayama, H.; Murayama, T.; Horie, S. Experimental colitis alters expression of 5-HT receptors and transient receptor potential vanilloid 1 leading to visceral hypersensitivity in mice. Lab. Investig. 2012, 92, 769–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koopman, N.; Katsavelis, D.; ten Hove, A.S.; Brul, S.; de Jonge, W.J.; Seppen, J. The Multifaceted Role of Serotonin in Intestinal Homeostasis. Int. J. Mol. Sci. 2021, 22, 9487. [Google Scholar] [CrossRef]
- Wang, B.; Sun, S.; Liu, M.; Chen, H.; Liu, N.; Wu, Z.; Wu, G.; Dai, Z. Dietary L-Tryptophan Regulates Colonic Serotonin Homeostasis in Mice with Dextran Sodium Sulfate-Induced Colitis. J. Nutr. 2020, 150, 1966–1976. [Google Scholar] [CrossRef]
- Nau, F.; Yu, B.; Martin, D.; Nichols, C.D. Serotonin 5-HT2A Receptor Activation Blocks TNF-α Mediated Inflammation In Vivo. PLoS ONE 2013, 8, e75426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, L.; Xin, F.; Ren, J.; Xu, S.; Huang, H.; Zha, X.; Wen, X.; Gu, G.; Yang, G.; Cheng, Y.; et al. 5-HT2B-mediated serotonin activation in enterocytes suppresses colitis-associated cancer initiation and promotes cancer progression. Theranostics 2022, 12, 3928–3945. [Google Scholar] [CrossRef] [PubMed]
- González-Maeso, J.; Weisstaub, N.V.; Zhou, M.; Chan, P.; Ivic, L.; Ang, R.; Lira, A.; Bradley-Moore, M.; Ge, Y.; Zhou, Q.; et al. Hallucinogens recruit specific cortical 5-HT(2A) receptor-mediated signaling pathways to affect behavior. Neuron 2007, 53, 439–452. [Google Scholar] [CrossRef] [Green Version]
- Zielińska, M.; Jarmuż, A.; Wasilewski, A.; Sałaga, M.; Fichna, J. Role of transient receptor potential channels in intestinal inflammation and visceral pain: Novel targets in inflammatory bowel diseases. Inflamm. Bowel Dis. 2015, 21, 419–427. [Google Scholar] [CrossRef]
- Csekő, K.; Beckers, B.; Keszthelyi, D.; Helyes, Z. Role of TRPV1 and TRPA1 Ion Channels in Inflammatory Bowel Diseases: Potential Therapeutic Targets? Pharmaceuticals 2019, 12, 48. [Google Scholar] [CrossRef] [Green Version]
- Ramachandran, R.; Hyun, E.; Zhao, L.; Lapointe, T.K.; Chapman, K.; Hirota, C.L.; Ghosh, S.; McKemy, D.D.; Vergnolle, N.; Beck, P.L.; et al. TRPM8 activation attenuates inflammatory responses in mouse models of colitis. Proc. Natl. Acad. Sci. USA 2013, 110, 7476–7481. [Google Scholar] [CrossRef]
- de Jong, P.R.; Takahashi, N.; Peiris, M.; Bertin, S.; Lee, J.; Gareau, M.G.; Paniagua, A.; Harris, A.R.; Herdman, D.S.; Corr, M.; et al. TRPM8 on mucosal sensory nerves regulates colitogenic responses by innate immune cells via CGRP. Mucosal. Immunol. 2015, 8, 491–504. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; eun Kwon, J.; Cho, M.L. Immunological pathogenesis of inflammatory bowel disease. Intest. Res. 2018, 16, 26–42. [Google Scholar] [CrossRef] [Green Version]
- Petranker, R.; Anderson, T.; Maier, L.J.; Barratt, M.J.; Ferris, J.A.; Winstock, A.R. Microdosing psychedelics: Subjective benefits and challenges, substance testing behavior, and the relevance of intention. J. Psychopharmacol. 2022, 36, 85–96. [Google Scholar] [CrossRef]
- Zinberg, N.E. Drug, Set, and Setting: The Basis for Controlled Intoxicant Use; Yale University Press: London, UK, 1986. [Google Scholar]
- Irizarry, R.; Winczura, A.; Dimassi, O.; Dhillon, N.; Minhas, A.; Larice, J. Psilocybin as a Treatment for Psychiatric Illness: A Meta-Analysis. Cureus 2022, 14, e31796. [Google Scholar] [CrossRef]
- Ona, G.; Bouso, J.C. Potential safety, benefits, and influence of the placebo effect in microdosing psychedelic drugs: A systematic review. Neurosci. Biobehav. Rev. 2020, 119, 194–203. [Google Scholar] [CrossRef]
- Kuypers, K.P.C. The therapeutic potential of microdosing psychedelics in depression. Ther. Adv. Psychopharmacol. 2020, 10, 2045125320950567. [Google Scholar] [CrossRef]
- Tang, Q.; Li, Z.Q.; Li, W.; Guo, J.; Sun, H.Y.; Zhang, X.H.; Lau, C.P.; Tse, H.F.; Zhang, S.; Li, G.R. The 5-HT2 antagonist ketanserin is an open channel blocker of human cardiac ether-à-go-go-related gene (hERG) potassium channels. Br. J. Pharmacol. 2008, 155, 365–373. [Google Scholar] [CrossRef] [Green Version]
- Aronson, J.K. (Ed.) Ketanserin. In Meyler’s Side Effects of Drugs, 16th ed.; Elsevier: Amsterdam, The Netherlands, 2016; p. 413. [Google Scholar] [CrossRef]
- Pope, J.; Fenlon, D.; Thompson, A.; Shea, B.; Furst, D.; Wells, G.A.; Silman, A. Ketanserin for Raynaud’s phenomenon in progressive systemic sclerosis. Cochrane Database Syst. Rev. 2000, 1998, CD000954. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Short-Term Cytotoxicity | Long-Term Cytotoxicity | Most Effective Dose | Fold Change of COX-2 | Fold Change of IL-6 |
|---|---|---|---|---|---|
| Psi | N/A | N/A | 40 μM | −3.8× | N/A |
| 4-AcO-DMT | N/A | N/A | 20 μM | −1.5× | N/A |
| Ket | ≥40 μM | ≥5 μM | 1 μM | −1.7× | N/A |
| Cap | ≥50 μM | ≥0.5 μM | 0.5 μM | −1.6× | N/A |
| Cur | ≥5 μM | ≥1 μM | 0.5 μM | −3.8× | N/A |
| Eug | ≥10 μM | ≥10 μM | 50 μM | −3.8× | N/A |
| Psi + Cap | N/A | 10 μM Psi, ≥0.5 μM Cap | 20 μM Psi, 0.5 μM Cap | −3.8× | −1.7× |
| Psi + Cur | 10 μM Psi & 1 μM Cur | 10/20 μM Psi, ≥0.5 μM Cur | 40 μM Psi, 0.5 μM Cur | −28× | −1.6× |
| Psi + Eug | N/A | N/A | 40 μM Psi, 25 μM Eug | −19× | −10× |
| 4-AcO-DMT + Cap | N/A | N/A | 20 μM 4-AcO-DMT, 0.5 μM Cap | −1.1× | −3.3× |
| 4-AcO-DMT + Cur | N/A | 10 μM 4-AcO-DMT, ≥0.5 μM Cur | 40 μM 4-AcO-DMT, 0.5 μM Cur | −1.3× | −1.3× |
| 4-AcO-DMT + Eug | N/A | N/A | 40 μM 4-AcO-DMT, 25 μM Eug | −1.6× | −1.5× |
| Ket + Cap | N/A | N/A | 5 μM Ket, 0.5 μM Cap | −2.1× | −2.0× |
| Ket + Cur | N/A | 10 μM Ket, ≥0.5 μM Cur | 1 μM Ket, 0.5 μM Cur | −3.5× | −1.1× |
| Ket + Eug | N/A | ≥1 μM Ket, 25 μM Eug | 10 μM Ket, 25 μM Eug | −2.8× | −3.0× |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robinson, G.I.; Li, D.; Wang, B.; Zahoruiko, Y.; Gerasymchuk, M.; Hudson, D.; Kovalchuk, O.; Kovalchuk, I. Anti-Inflammatory Effects of Serotonin Receptor and Transient Receptor Potential Channel Ligands in Human Small Intestinal Epithelial Cells. Curr. Issues Mol. Biol. 2023, 45, 6743-6774. https://doi.org/10.3390/cimb45080427
Robinson GI, Li D, Wang B, Zahoruiko Y, Gerasymchuk M, Hudson D, Kovalchuk O, Kovalchuk I. Anti-Inflammatory Effects of Serotonin Receptor and Transient Receptor Potential Channel Ligands in Human Small Intestinal Epithelial Cells. Current Issues in Molecular Biology. 2023; 45(8):6743-6774. https://doi.org/10.3390/cimb45080427
Chicago/Turabian StyleRobinson, Gregory Ian, Dongping Li, Bo Wang, Yeva Zahoruiko, Marta Gerasymchuk, Darryl Hudson, Olga Kovalchuk, and Igor Kovalchuk. 2023. "Anti-Inflammatory Effects of Serotonin Receptor and Transient Receptor Potential Channel Ligands in Human Small Intestinal Epithelial Cells" Current Issues in Molecular Biology 45, no. 8: 6743-6774. https://doi.org/10.3390/cimb45080427
APA StyleRobinson, G. I., Li, D., Wang, B., Zahoruiko, Y., Gerasymchuk, M., Hudson, D., Kovalchuk, O., & Kovalchuk, I. (2023). Anti-Inflammatory Effects of Serotonin Receptor and Transient Receptor Potential Channel Ligands in Human Small Intestinal Epithelial Cells. Current Issues in Molecular Biology, 45(8), 6743-6774. https://doi.org/10.3390/cimb45080427