
Quantitative Proteomic Profiling of Tachyplesin I Targets in U251 Gliomaspheres

Abstract
:1. Introduction
2. Results
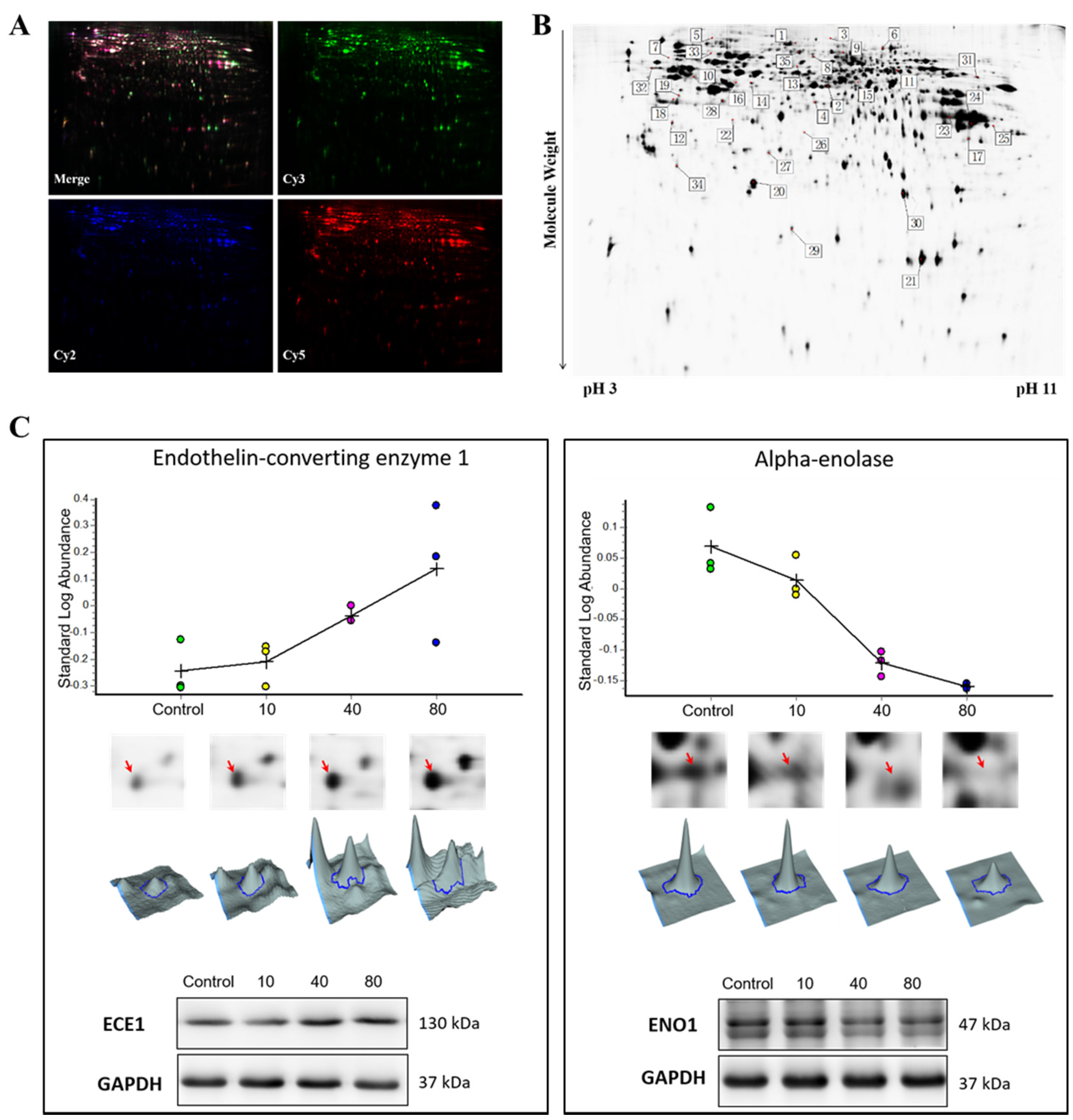
2.1. Protein Expression Profile of Tachyplesin I Treated U251 Gliomaspheres Using 2D-DIGE Analysis
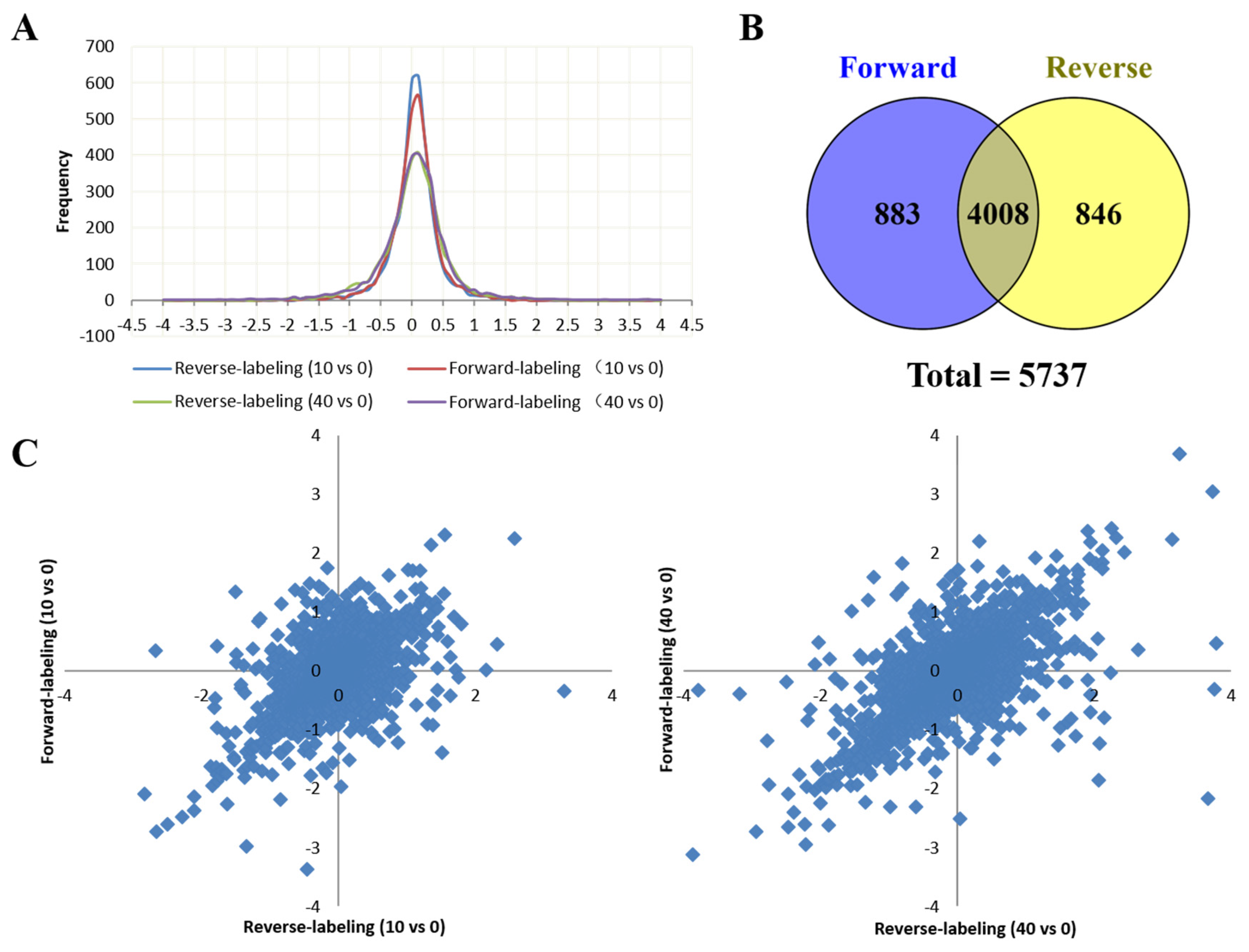
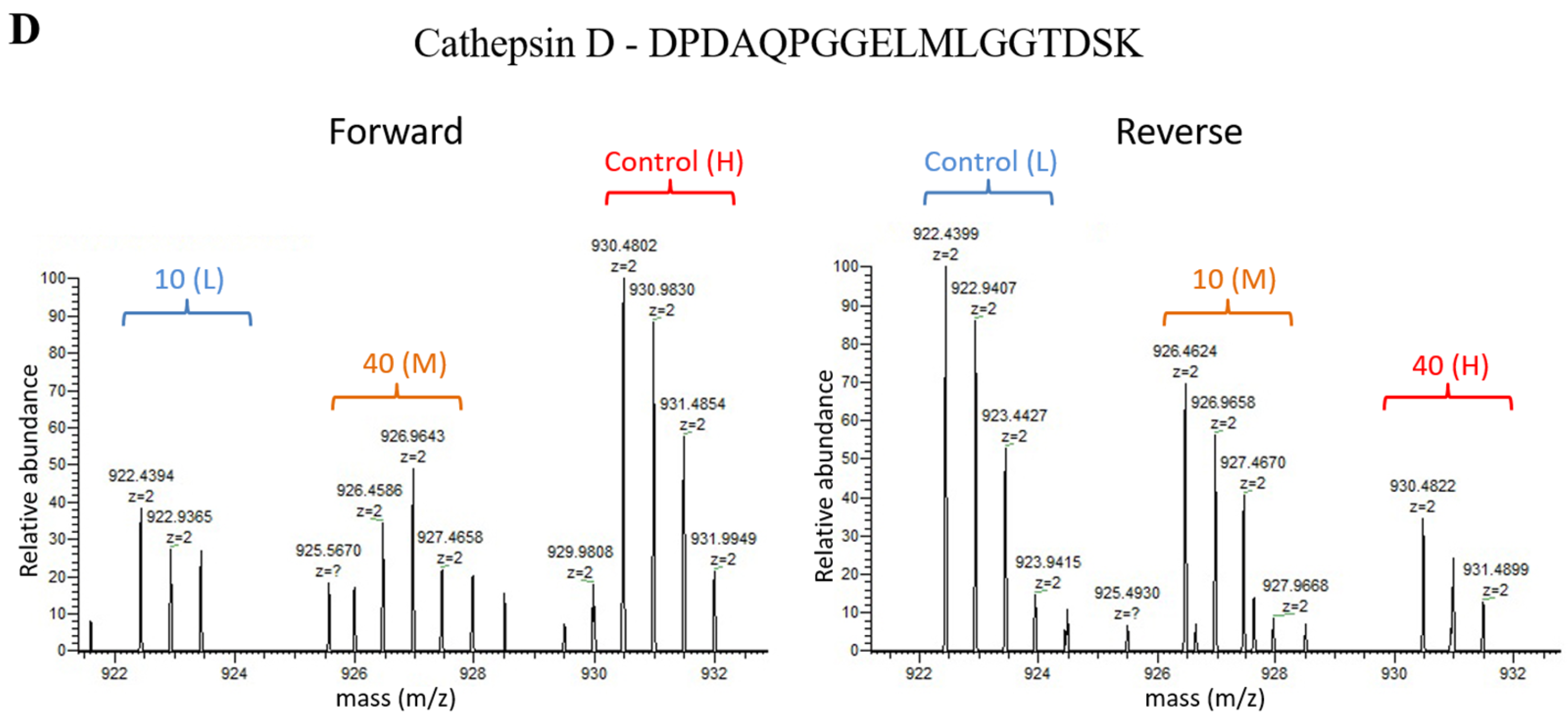
2.2. Relative Quantification Using Dimethyl Labeling Based LC-MS/MS Analysis
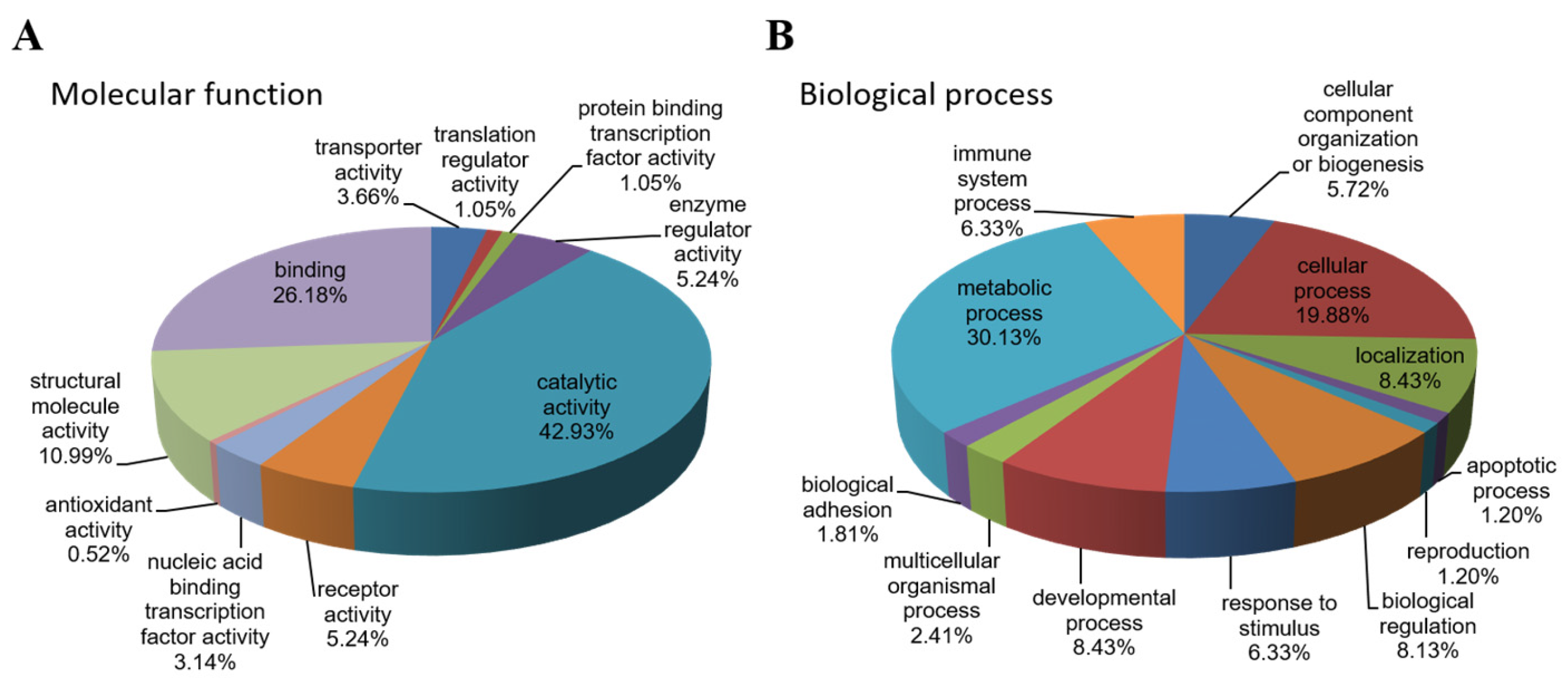
2.3. Cellular Functions of Differentially Expressed Proteins and Associated Pathways
2.4. Tachyplesin I Influences Metabolic Process and Alters the Expressions of Cytoskeleton Proteins
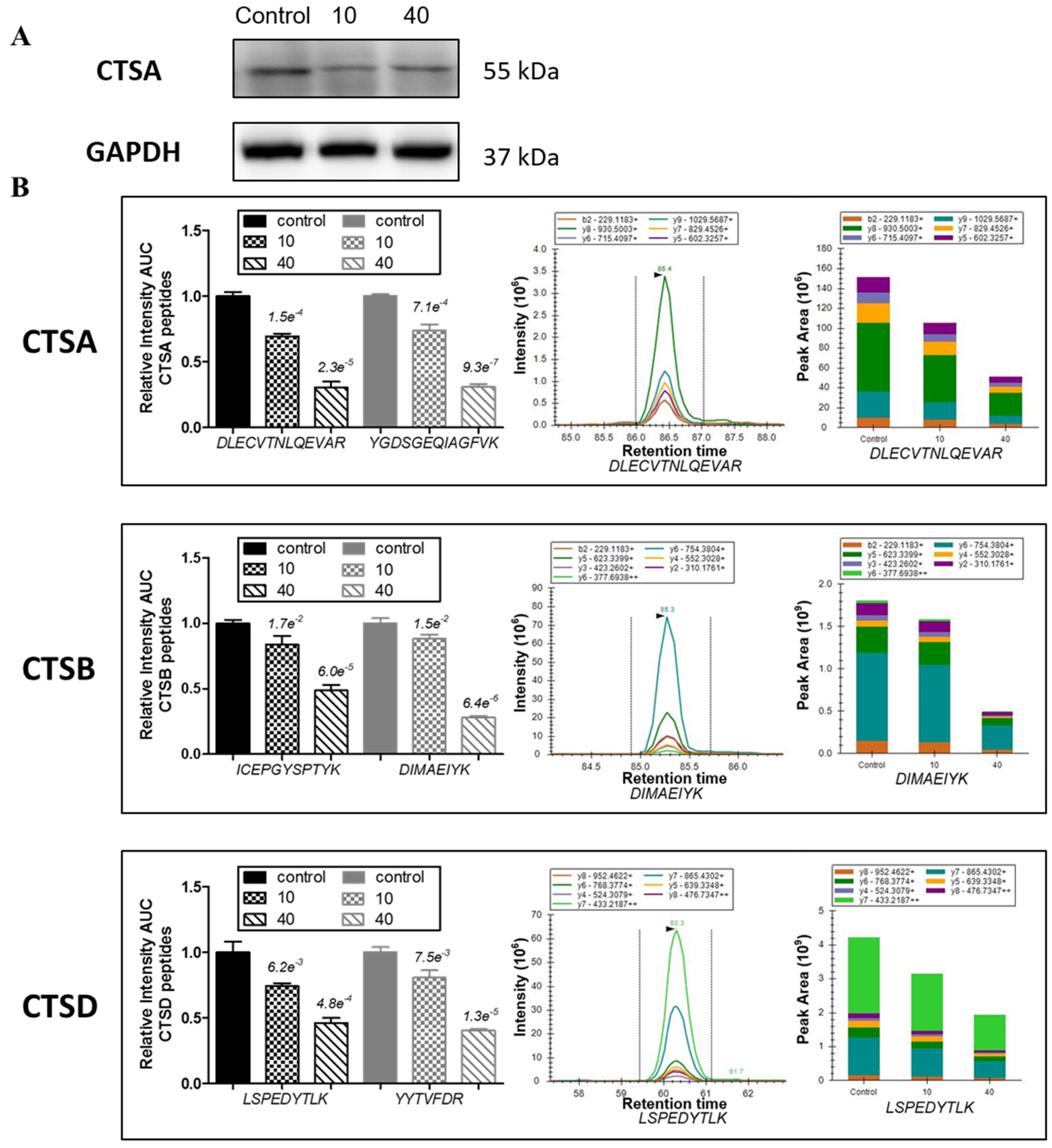
2.5. Tachyplesin I Reduces Expressions of Several Lysosomal Acid Hydrolases
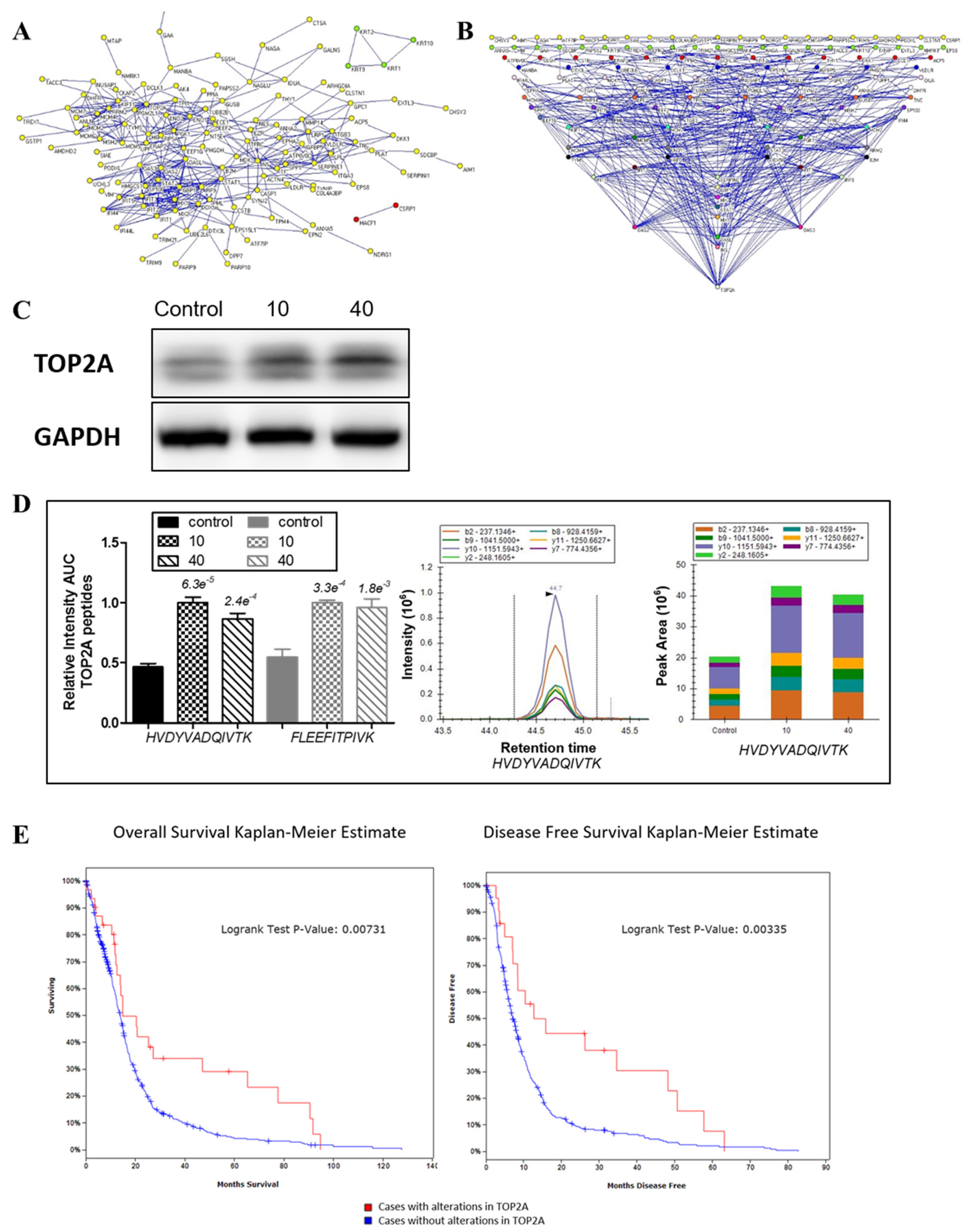
2.6. Protein-Protein Interaction Network of Differentially Expressed Proteins
2.7. Confirmation of the Involvement of TOP2A in the Effects of Tachyplesin I and Correlation with Clinical Prognosis in TCGA Database
3. Discussion
4. Material and Methods
4.1. Cell Culture and Treatment with Tachyplesin I
4.2. CyDye Minimal Labeling of Protein Samples and 2D-DIGE Electrophoresis
4.3. In-Gel Digestion and Protein Identification by MALDI-TOF/TOF
4.4. Dimethyl Labeling of Protein Samples
4.5. High pH Fractionation of Peptides and LC-MS/MS Analysis by Obitrap
4.6. Bioinformatic Analysis
4.7. Parallel Reaction Monitoring (PRM) Mass Spectrometry
4.8. Western Blot Assay
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Alifieris, C.; Trafalis, D.T. Glioblastoma multiforme: Pathogenesis and treatment. Pharmacol. Ther. 2015, 152, 63–82. [Google Scholar] [CrossRef] [PubMed]
- Stopschinski, B.E.; Beier, C.P.; Beier, D. Glioblastoma cancer stem cells—From concept to clinical application. Cancer Lett. 2013, 338, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Hawkins, C.; Clarke, I.D.; Squire, J.A.; Bayani, J.; Hide, T.; Henkelman, R.M.; Cusimano, M.D.; Dirks, P.B. Identification of human brain tumour initiating cells. Nature 2004, 432, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Schonberg, D.L.; Miller, T.E.; Wu, Q.; Flavahan, W.A.; Das, N.K.; Hale, J.S.; Hubert, C.G.; Mack, S.C.; Jarrar, A.M.; Karl, R.T.; et al. Preferential iron trafficking characterizes glioblastoma stem-like cells. Cancer Cell 2015, 28, 441–455. [Google Scholar] [CrossRef] [PubMed]
- Pointer, K.B.; Clark, P.A.; Zorniak, M.; Alrfaei, B.M.; Kuo, J.S. Glioblastoma cancer stem cells: Biomarker and therapeutic advances. Neurochem. Int. 2014, 71, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.; Wu, Q.; Sathornsumetee, S.; Hao, Y.; Li, Z.; Hjelmeland, A.B.; Shi, Q.; McLendon, R.E.; Bigner, D.D.; Rich, J.N. Stem cell-like glioma cells promote tumor angiogenesis through vascular endothelial growth factor. Cancer Res. 2006, 66, 7843–7848. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.B. Three-dimensional tissue culture models in cancer biology. Semin. Cancer Biol. 2005, 15, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Pampaloni, F.; Reynaud, E.G.; Stelzer, E.H. The third dimension bridges the gap between cell culture and live tissue. Nat. Rev. Mol. Cell Biol. 2007, 8, 839–845. [Google Scholar] [CrossRef] [PubMed]
- Kong, B.H.; Park, N.R.; Shim, J.K.; Kim, B.K.; Shin, H.J.; Lee, J.H.; Huh, Y.M.; Lee, S.J.; Kim, S.H.; Kim, E.H.; et al. Isolation of glioma cancer stem cells in relation to histological grades in glioma specimens. Childs Nerv. Syst. 2013, 29, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.B.; Ji, X.Y.; Huang, Q.; Dong, J.; Zhu, Y.D.; Lan, Q. Differentiation profile of brain tumor stem cells: A comparative study with neural stem cells. Cell Res. 2006, 16, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Jin, G.; Zhang, L.; Dai, J.; Dang, J.; Han, Y. Effects of tachyplesin I on human U251 glioma stem cells. Mol. Med. Rep. 2015, 11, 2953–2958. [Google Scholar] [PubMed]
- Nakamura, T.; Furunaka, H.; Miyata, T.; Tokunaga, F.; Muta, T.; Iwanaga, S.; Niwa, M.; Takao, T.; Shimonishi, Y. Tachyplesin, a class of antimicrobial peptide from the hemocytes of the horseshoe crab (Tachypleus tridentatus). Isolation and chemical structure. J. Biol. Chem. 1988, 263, 16709–16713. [Google Scholar] [PubMed]
- Rao, A.G. Conformation and antimicrobial activity of linear derivatives of tachyplesin lacking disulfide bonds. Arch. Biochem. Biophys. 1999, 361, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xu, X.; Hong, S.; Chen, J.; Liu, N.; Underhill, C.B.; Creswell, K.; Zhang, L. RGD-tachyplesin inhibits tumor growth. Cancer Res. 2001, 61, 2434–2438. [Google Scholar] [PubMed]
- Li, Q.F.; Ou-Yang, G.L.; Peng, X.X.; Hong, S.G. Effects of tachyplesin on the regulation of cell cycle in human hepatocarcinoma SMMC-7721 cells. World J. Gastroenterol. 2003, 9, 454–458. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xu, X.M.; Underhill, C.B.; Yang, S.; Wang, L.; Chen, Y.; Hong, S.; Creswell, K.; Zhang, L. Tachyplesin activates the classic complement pathway to kill tumor cells. Cancer Res. 2005, 65, 4614–4622. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, G.L.; Li, Q.F.; Peng, X.X.; Liu, Q.R.; Hong, S.G. Effects of tachyplesin on proliferation and differentiation of human hepatocellular carcinoma SMMC-7721 cells. World J. Gastroenterol. 2002, 8, 1053–1058. [Google Scholar] [CrossRef] [PubMed]
- Hoskin, D.W.; Ramamoorthy, A. Studies on anticancer activities of antimicrobial peptides. Biochim. Biophys. Acta 2008, 1778, 357–375. [Google Scholar] [CrossRef] [PubMed]
- Baggerman, G.; Vierstraete, E.; De Loof, A.; Schoofs, L. Gel-based versus gel-free proteomics: A review. Comb. Chem. High Throughput Screen. 2005, 8, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Ren, X.; Huang, Z.; Yang, X.; Hong, W.; Zhang, Y.; Zhang, H.; Liu, W.; Huang, H.; Huang, X.; et al. Serum proteomic analysis reveals potential serum biomarkers for occupational medicamentosa-like dermatitis caused by trichloroethylene. Toxicol. Lett. 2014, 229, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Peterson, A.C.; Russell, J.D.; Bailey, D.J.; Westphall, M.S.; Coon, J.J. Parallel reaction monitoring for high resolution and high mass accuracy quantitative, targeted proteomics. Mol. Cell. Proteom. 2012, 11, 1475–1488. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.N.; Harlan, R.; Chen, J.; Aiyetan, P.; Liu, Y.; Sokoll, L.J.; Aebersold, R.; Chan, D.W.; Zhang, H. Multiplexed targeted mass spectrometry-based assays for the quantification of N-linked glycosite-containing peptides in serum. Anal. Chem. 2015, 87, 10830–10838. [Google Scholar] [CrossRef] [PubMed]
- Ganapathy-Kanniappan, S.; Geschwind, J.F. Tumor glycolysis as a target for cancer therapy: Progress and prospects. Mol. Cancer 2013, 12, 152. [Google Scholar] [CrossRef] [PubMed]
- Ramao, A.; Gimenez, M.; Laure, H.J.; Izumi, C.; Vida, R.C.; Oba-Shinjo, S.; Marie, S.K.; Rosa, J.C. Changes in the expression of proteins associated with aerobic glycolysis and cell migration are involved in tumorigenic ability of two glioma cell lines. Proteome Sci. 2012, 10, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsetos, C.D.; Reginato, M.J.; Baas, P.W.; D’Agostino, L.; Legido, A.; Tuszyn Ski, J.A.; Draberova, E.; Draber, P. Emerging microtubule targets in glioma therapy. Semin. Pediatr. Neurol. 2015, 22, 49–72. [Google Scholar] [CrossRef] [PubMed]
- Fehrenbacher, N.; Jaattela, M. Lysosomes as targets for cancer therapy. Cancer Res. 2005, 65, 2993–2995. [Google Scholar] [PubMed]
- Kozlowski, L.; Wojtukiewicz, M.Z.; Ostrowska, H. Cathepsin A activity in primary and metastatic human melanocytic tumors. Arch. Dermatol. Res. 2000, 292, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, N.; Sloane, B.F. Cathepsin B: Multiple roles in cancer. Proteom. Clin. Appl. 2014, 8, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Nicotra, G.; Castino, R.; Follo, C.; Peracchio, C.; Valente, G.; Isidoro, C. The dilemma: Does tissue expression of cathepsin D reflect tumor malignancy? The question: Does the assay truly mirror cathepsin D mis-function in the tumor? Cancer Biomark. 2010, 7, 47–64. [Google Scholar] [PubMed]
- Tan, G.J.; Peng, Z.K.; Lu, J.P.; Tang, F.Q. Cathepsins mediate tumor metastasis. World J. Biol. Chem. 2013, 4, 91–101. [Google Scholar] [PubMed]
- Liu, Y.; Zhou, Y.; Zhu, K. Inhibition of glioma cell lysosome exocytosis inhibits glioma invasion. PLoS ONE 2012, 7, e45910. [Google Scholar] [CrossRef] [PubMed]
- Vassetzky, Y.S.; Alghisi, G.C.; Gasser, S.M. DNA topoisomerase II mutations and resistance to anti-tumor drugs. Bioessays 1995, 17, 767–774. [Google Scholar] [CrossRef] [PubMed]
- McPherson, J.P.; Goldenberg, G.J. Induction of apoptosis by deregulated expression of DNA topoisomerase IIalpha. Cancer Res. 1998, 58, 4519–4524. [Google Scholar] [PubMed]
- McPherson, J.P.; Brown, G.A.; Goldenberg, G.J. Characterization of a DNA topoisomerase IIalpha gene rearrangement in adriamycin-resistant P388 leukemia: Expression of a fusion messenger RNA transcript encoding topoisomerase iialpha and the retinoic acid receptor alpha locus. Cancer Res. 1993, 53, 5885–5889. [Google Scholar] [PubMed]
- Withoff, S.; De Jong, S.; De Vries, E.G.; Mulder, N.H. Human DNA topoisomerase II: Biochemistry and role in chemotherapy resistance (review). Anticancer Res. 1996, 16, 1867–1880. [Google Scholar] [PubMed]
- Wessel, D.; Flugge, U.I. A method for the quantitative recovery of protein in dilute solution in the presence of detergents and lipids. Anal. Biochem. 1984, 138, 141–143. [Google Scholar] [CrossRef]
- Boersema, P.J.; Raijmakers, R.; Lemeer, S.; Mohammed, S.; Heck, A.J. Multiplex peptide stable isotope dimethyl labeling for quantitative proteomics. Nat. Protoc. 2009, 4, 484–494. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cbioportal. Sci. Signal. 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacLean, B.; Tomazela, D.M.; Shulman, N.; Chambers, M.; Finney, G.L.; Frewen, B.; Kern, R.; Tabb, D.L.; Liebler, D.C.; MacCoss, M.J. Skyline: An open source document editor for creating and analyzing targeted proteomics experiments. Bioinformatics 2010, 26, 966–968. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, X.; Zhu, Z.; Huang, P.; Zhuang, Z.; Liu, J.; Gao, W.; Liu, Y.; Huang, H. Poly(ADP-ribose) glycohydrolase (PARG) silencing suppresses benzo(a)pyrene induced cell transformation. PLoS ONE 2016, 11, e0151172. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Up-Regulated Proteins of Tachyplesin I Treated U251 Gliomaspheres in the 2D-DIGE Study | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| No. a | Gene Name | Uniprot ID | Protein Name | Mascot Score | Peptides | Protein MW | pI Value | Ratio/p Value b | Ratio/p Value b |
| 10 vs. 0 c | 40 vs. 0 c | ||||||||
| Regulation of cell cycle or apoptosis d | |||||||||
| 2 | PHGDH | O43175 | d-3-phosphoglycerate dehydrogenase | 104 | 3 | 57,356 | 6.3 | 1.54/0.003 | 1.69/0.017 |
| 5 | MSH2 | P43246 | DNA mismatch repair protein Msh2 | 93 | 2 | 104,743 | 5.8 | 1.21/0.009 | 1.77/0.035 |
| 19 | SESN3 | P58005 | Sestrin-3 | 201 | 4 | 57,291 | 6.3 | 1.52/0.036 | 2.01/0.027 |
| 31 | CKAP2 | Q8WWK9 | Cytoskeleton-associated protein 2 | 76 | 7 | 76,987 | 9.4 | ND e | 1.52/0.039 |
| 35 | ECE1 | P42892 | Endothelin-converting enzyme 1 | 46 | 1 | 87,164 | 5.9 | 1.58/0.038 | 3.31/0.026 |
| Cytoskeletal protein d | |||||||||
| 3 | VIM | P08670 | Vimentin | 407 | 12 | 53,676 | 4.9 | ND | 1.64/0.022 |
| 4 | EEF1G | P26641 | Elongation factor 1-gamma | 47 | 2 | 50,429 | 6.3 | ND | 1.51/0.005 |
| 9 | EZR | P15311 | Ezrin | 168 | 3 | 69,484 | 5.9 | 1.03/0.024 | 1.63/0.046 |
| 10 | VIM | P08670 | Vimentin | 524 | 15 | 53,676 | 4.9 | ND | 1.58/0.041 |
| Protein biosynthesis d | |||||||||
| 6 | EEF2 | P13639 | Elongation factor 2 | 60 | 1 | 96,246 | 6.4 | ND | 1.55/0.044 |
| 21 | PPIA | P62937 | Peptidyl-prolyl cis-trans isomerase A | 195 | 16 | 18,229 | 9 | 1.26/0.017 | 1.52/0.028 |
| Transport d | |||||||||
| 23 | SLC25A3 | F8VVM2 | Phosphate carrier protein, mitochondrial | 90 | 5 | 36,161 | 9.3 | ND | 1.72/0.034 |
| 25 | SLC25A3 | F8VVM2 | Phosphate carrier protein, mitochondrial | 183 | 9 | 36,161 | 9.3 | ND | 1.63/0.016 |
| Down-Regulated Proteins of Tachyplesin I Treated U251 Gliomaspheres in the 2D-DIGE Study | |||||||||
| Calcium or iron ion binding protein d | |||||||||
| 7 | EPS15 | P42566 | Epidermal growth factor receptor substrate 15 | 109 | 5 | 98,656 | 5.1 | ND e | −1.88/0.004 |
| 13 | P4HA1 | P13674 | Prolyl 4-hydroxylase subunit alpha-1 | 86 | 5 | 61,296 | 5.6 | −1.51/0.007 | −2.37/0.045 |
| Regulation of cell apoptosis or proliferation d | |||||||||
| 12 | ANXA5 | P08758 | Annexin A5 | 273 | 13 | 35,971 | 4.8 | ND | −1.74/0.033 |
| 20 | GSTP1 | P09211 | Glutathione S-transferase P | 339 | 21 | 23,569 | 5.3 | ND | −1.66/0.001 |
| 33 | COL4A3BP | Q9Y5P4 | Collagen type IV alpha-3-binding protein | 250 | 7 | 70,835 | 5.5 | ND | −1.64/0.033 |
| 34 | ARHGDIA | P52565 | Rho GDP-dissociation inhibitor 1 | 236 | 16 | 23,250 | 4.9 | −1.23/0.037 | −2.64/0.047 |
| Response to stimulus d | |||||||||
| 14 | GNAQ | P50148 | Guanine nucleotide-binding protein G(q) subunit alpha | 193 | 9 | 42,142 | 5.7 | ND | −1.61/0.037 |
| 16 | GNAQ | P50148 | Guanine nucleotide-binding protein G(q) subunit alpha | 294 | 12 | 42,142 | 5.7 | ND | −1.59/0.047 |
| 28 | GNAQ | P50148 | Guanine nucleotide-binding protein G(q) subunit alpha | 182 | 7 | 42,142 | 5.7 | −1.33/0.028 | −1.53/0.036 |
| Glycolysis/Gluconeogenesis d | |||||||||
| 15 | ENO1 | P06733 | Alpha-enolase | 40 | 3 | 47,481 | 7.7 | −1.04/0.005 | −1.92/0.054 |
| 17 | PGK1 | P00558 | Phosphoglycerate kinase 1 | 209 | 7 | 44,985 | 9.2 | −1.68/0.025 | −2.89/0.051 |
| 30 | TPI1 | P60174 | Triosephosphate isomerase | 375 | 15 | 31,057 | 5.6 | −1.17/0.048 | −1.88/0.028 |
| Ribosomal protein d | |||||||||
| 18 | RPSA | P08865 | 40S ribosomal protein SA | 171 | 9 | 32,947 | 4.6 | −1.49/0.036 | −1.89/0.027 |
| The 55 Up-Regulated Proteins Expressed More Than 2 Folds (<1% FDR) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Gene Name | Uniprot ID | Protein Name | Coverage (%) a | Unique Peptides a | 10 vs. 0 Ratio b | 40 vs. 0 Ratio b | Protein Class c | ||
| Forward | Reverse | Forward | Reverse | ||||||
| SPP1 | P10451 | Osteopontin | 33.12 | 5 | 4.961 | 2.943 | 12.876 | 9.484 | cytokine |
| ITGB3 | P05106 | Integrin beta-3 | 5.20 | 3 | 4.754 | 5.953 | 13.279 | 28.714 | receptor, extracellular matrix glycoprotein |
| EPS8 | Q12929 | Epidermal growth factor receptor kinase substrate 8 | 4.01 | 3 | 4.403 | 2.549 | 2.220 | 2.305 | transmembrane receptor regulatory/adaptor protein |
| MCM5 | B1AHB1 | DNA helicase | 5.50 | 3 | 3.304 | 2.022 | 3.801 | 2.194 | DNA helicase |
| DKK1 | O94907 | Dickkopf-related protein 1 | 11.28 | 4 | 3.267 | 2.278 | 3.885 | 2.732 | developmental protein, growth factor activity |
| MCM4 | P33991 | DNA replication licensing factor MCM4 | 7.76 | 4 | 3.248 | 2.119 | 5.189 | 3.723 | DNA binding protein |
| NUSAP1 | Q9BXS6 | Nucleolar and spindle-associated protein 1 | 18.14 | 5 | 2.661 | 2.293 | 3.121 | 3.405 | microtubule-associated protein |
| DHFR | P00374 | Dihydrofolate reductase | 24.06 | 4 | 2.616 | 2.168 | 2.398 | 2.472 | reductase |
| TOP2A | P11388 | DNA topoisomerase 2-alpha | 12.93 | 12 | 2.490 | 2.916 | 3.259 | 2.582 | DNA topoisomerase, enzyme modulator |
| MKI67 | A0A087WV66 | Antigen KI-67 | 12.66 | 24 | 2.396 | 1.870 | 2.688 | 2.047 | regulation of cell proliferation |
| TFRC | P02786 | Transferrin receptor protein 1 | 35.79 | 23 | 2.323 | 2.398 | 2.906 | 2.879 | receptor |
| AIM1 | Q9Y4K1 | Absent in melanoma 1 protein | 21.53 | 25 | 2.298 | 2.329 | 4.036 | 5.403 | carbohydrate binding protein |
| ECE1 | P42892 | Endothelin-converting enzyme 1 | 15.19 | 8 | 2.260 | 2.671 | 3.557 | 4.070 | metalloprotease |
| SYNJ2 | O15056 | Synaptojanin-2 | 8.76 | 11 | 2.239 | 2.501 | 5.349 | 4.740 | phosphatase |
| KIF11 | P52732 | Kinesin-like protein KIF11 | 2.37 | 2 | 2.199 | 2.274 | 2.469 | 2.182 | microtubule binding motor protein |
| DST | Q03001 | Dystonin | 23.49 | 21 | 2.113 | 1.596 | 2.614 | 2.051 | non-motor actin binding protein |
| UPP1 | Q16831 | Uridine phosphorylase 1 | 46.45 | 11 | 2.078 | 2.712 | 2.209 | 3.577 | phosphorylase |
| IGFBP5 | P24593 | Insulin-like growth factor-binding protein 5 | 20.59 | 5 | 2.070 | 2.060 | 4.816 | 4.967 | cell communication |
| RRM2 | P31350 | Ribonucleoside-diphosphate reductase subunit M2 | 34.45 | 11 | 2.030 | 1.981 | 2.286 | 2.423 | reductase |
| CD70 | P32970 | CD70 antigen | 35.23 | 6 | 1.995 | 2.167 | 3.725 | 3.808 | cell communication |
| MDK | E9PPJ5 | Midkine (Fragment) | 27.48 | 2 | 1.935 | 2.538 | 3.569 | 4.336 | cytokine |
| HMGCS1 | Q01581 | Hydroxymethylglutaryl-CoA synthase, cytoplasmic | 42.88 | 17 | 1.929 | 1.939 | 3.322 | 4.339 | transferase, lyase |
| DCLK1 | Q5VZY9 | Serine/threonine-protein kinase DCLK1 | 10.74 | 3 | 1.899 | 3.222 | 4.721 | 8.819 | non-receptor serine/threonine protein kinase |
| MCM7 | P33993 | DNA replication licensing factor MCM7 | 13.21 | 7 | 1.879 | 1.963 | 2.451 | 2.470 | DNA helicase |
| PODXL | O00592 | Podocalyxin | 2.33 | 1 | 1.870 | 2.234 | 2.474 | 2.856 | regulation of adhesion and cell morphology |
| MCM2 | H0Y8E6 | DNA replication licensing factor MCM2 (Fragment) | 8.25 | 6 | 1.865 | 1.513 | 2.167 | 2.664 | DNA helicase |
| LPL | P06858 | Lipoprotein lipase | 34.11 | 12 | 1.848 | 2.078 | 4.136 | 4.351 | storage protein |
| VSNL1 | P62760 | Visinin-like protein 1 | 24.08 | 4 | 1.805 | 2.260 | 3.231 | 2.926 | cell communication |
| MCM6 | Q14566 | DNA replication licensing factor MCM6 | 9.01 | 4 | 1.765 | 2.208 | 2.606 | 3.031 | DNA helicase |
| GPC1 | P35052 | Glypican-1 | 33.69 | 14 | 1.745 | 1.665 | 2.861 | 3.347 | cell division and growth regulation |
| TACC3 | Q9Y6A5 | Transforming acidic coiled-coil-containing protein 3 | 3.22 | 2 | 1.735 | 2.068 | 2.563 | 2.322 | cytoskeleton |
| TNC | P24821 | Tenascin | 40.16 | 5 | 1.730 | 1.803 | 2.327 | 2.112 | signaling molecule |
| PLAT | P00750 | Tissue-type plasminogen activator | 24.73 | 12 | 1.729 | 3.475 | 8.267 | 13.172 | receptor, calmodulin |
| GATM | P50440 | Glycine amidinotransferase, mitochondrial | 28.61 | 9 | 1.704 | 1.793 | 2.591 | 2.740 | catalyze creatine biosynthesis |
| SERPINE1 | P05121 | Plasminogen activator inhibitor 1 | 21.14 | 7 | 1.695 | 1.448 | 4.539 | 3.853 | serine protease inhibitor |
| LMCD1 | Q9NZU5 | LIM and cysteine-rich domains protein 1 | 38.63 | 10 | 1.681 | 1.686 | 2.388 | 2.243 | structural protein |
| TYMS | P04818 | Thymidylate synthase | 19.17 | 4 | 1.675 | 2.752 | 2.151 | 3.228 | methyltransferase |
| ITGA3 | P26006 | Integrin alpha-3 | 21.41 | 19 | 1.671 | 1.729 | 2.847 | 2.944 | receptor, integrin |
| ANLN | Q9NQW6 | Actin-binding protein anillin | 3.91 | 3 | 1.666 | 2.235 | 2.308 | 3.143 | actin binding protein |
| ANXA2 | P07355 | Annexin A2 | 81.42 | 34 | 1.617 | 1.610 | 2.023 | 2.100 | fatty acid metabolic process |
| MACF1 | H3BPE1 | Microtubule-actin cross-linking factor 1, isoforms 1/2/3/5 | 29.64 | 153 | 1.610 | 1.601 | 2.018 | 2.000 | non-motor actin binding protein |
| TPM4 | P67936 | Tropomyosin alpha-4 chain | 48.79 | 8 | 1.534 | 2.126 | 2.878 | 2.893 | actin binding motor protein |
| ACTN4 | K7EJH8 | Alpha-actinin-4 (Fragment) | 68.68 | 1 | 1.526 | 2.411 | 2.251 | 3.153 | non-motor actin binding protein |
| TRIM9 | Q9C026 | E3 ubiquitin-protein ligase TRIM9 | 4.23 | 3 | 1.508 | 1.143 | 2.077 | 2.109 | ubiquitin-protein ligase |
| LDLR | P01130 | Low-density lipoprotein receptor | 6.63 | 5 | 1.498 | 1.202 | 2.266 | 2.096 | receptor, extracellular matrix glycoprotein |
| SDCBP | O00560 | Syntenin-1 | 28.52 | 4 | 1.495 | 1.490 | 2.581 | 3.361 | membrane trafficking regulatory protein |
| TF | P02787 | Serotransferrin | 45.13 | 27 | 1.422 | 1.526 | 2.224 | 2.216 | transfer/carrier protein |
| TENM2 | H7BYZ1 | Teneurin-2 | 13.86 | 24 | 1.422 | 1.735 | 2.046 | 2.501 | receptor, membrane-bound signaling molecule |
| NES | P48681 | Nestin | 58.61 | 78 | 1.396 | 1.363 | 2.140 | 2.167 | structural protein |
| THY1 | E9PIM6 | Thy-1 membrane glycoprotein (Fragment) | 25.66 | 3 | 1.360 | 1.468 | 2.432 | 2.131 | membrane glycoprotein |
| NEFL | P07196 | Neurofilament light polypeptide | 47.88 | 28 | 1.213 | 1.182 | 2.121 | 2.010 | structural protein |
| CLSTN1 | Q5SR54 | Calsyntenin-1 (Fragment) | 4.35 | 3 | 1.181 | 1.201 | 2.338 | 2.552 | cell adhesion molecule, calcium-binding protein |
| ECI2 | A0A0C4DGA2 | Enoyl-CoA delta isomerase 2, mitochondrial | 40.38 | 11 | 1.148 | 1.262 | 2.239 | 2.894 | transfer/carrier protein, enzyme modulator |
| PTPRE | P23469 | Receptor-type tyrosine-protein phosphatase epsilon | 11.29 | 5 | 0.939 | 2.069 | 3.282 | 3.385 | receptor, protein phosphatase |
| LRRC16A | Q5VZK9 | Leucine-rich repeat-containing protein 16A | 1.90 | 2 | ND | 1.408 | 2.281 | 3.321 | transcription cofactor |
| The 111 Down-Regulated Proteins Expressed Less Than 0.5 Folds (<1% FDR) | |||||||||
| OASL | Q15646 | 2′-5′-oligoadenylate synthase-like protein | 12.26 | 4 | 0.127 | 0.393 | 0.215 | ND | nucleotidyltransferase, defense/immunity protein |
| OAS2 | P29728 | 2′-5′-oligoadenylate synthase 2 | 9.74 | 9 | 0.151 | 0.158 | 0.115 | 0.068 | nucleotidyltransferase, defense/immunity protein |
| MX1 | P20591 | Interferon-induced GTP-binding protein Mx1 | 56.50 | 28 | 0.164 | 0.177 | 0.150 | 0.130 | microtubule family cytoskeletal protein |
| IFI44L | Q53G44 | Interferon-induced protein 44-like | 39.60 | 13 | 0.179 | 0.206 | 0.189 | 0.191 | immune response |
| IFI44 | Q8TCB0 | Interferon-induced protein 44 | 33.56 | 13 | 0.193 | 0.231 | 0.158 | 0.181 | immune response |
| CASP1 | G3V169 | Caspase | 19.35 | 4 | 0.209 | 0.325 | 0.235 | 0.180 | regulation of apoptotic process |
| BTN3A2 | E9PRR1 | Butyrophilin subfamily 3 member A2 (Fragment) | 27.55 | 2 | 0.220 | 0.555 | 0.261 | 0.360 | ubiquitin-protein ligase |
| INS | C9JNR5 | Insulin (Fragment) | 7.61 | 1 | 0.228 | 0.231 | 0.634 | 0.769 | growth factor |
| MX2 | P20592 | Interferon-induced GTP-binding protein Mx2 | 16.05 | 5 | 0.235 | 0.140 | 0.129 | 0.215 | microtubule family cytoskeletal protein |
| PARP10 | E9PPE7 | Poly [ADP-ribose] polymerase 10 | 4.71 | 2 | 0.260 | 0.280 | 0.201 | 0.508 | nucleic acid binding |
| ISG15 | A0A096LNZ9 | Ubiquitin-like protein ISG15 (Fragment) | 50.35 | 6 | 0.273 | 0.293 | 0.253 | 0.264 | ribosomal protein |
| TAP1 | Q03518 | Antigen peptide transporter 1 | 29.08 | 15 | 0.287 | 0.385 | 0.288 | 0.288 | ATP-binding cassette (ABC) transporter |
| IFIT3 | O14879 | Interferon-induced protein with tetratricopeptide repeats 3 | 48.57 | 18 | 0.288 | 0.301 | 0.261 | 0.258 | RNA binding |
| IFIT2 | P09913 | Interferon-induced protein with tetratricopeptide repeats 2 | 30.08 | 11 | 0.294 | 0.291 | 0.212 | 0.250 | RNA binding |
| IFIT1 | P09914 | Interferon-induced protein with tetratricopeptide repeats 1 | 45.82 | 16 | 0.300 | 0.321 | 0.296 | 0.301 | RNA binding |
| KRT10 | P13645 | Keratin, type I cytoskeletal 10 | 30.14 | 13 | 0.301 | 0.314 | 0.483 | 0.541 | structural protein |
| DDX58 | O95786 | Probable ATP-dependent RNA helicase DDX58 | 41.73 | 37 | 0.307 | 0.307 | 0.278 | 0.265 | helicase, hydrolase |
| BLOC1S1 | G8JLQ3 | Biogenesis of lysosome-related organelles complex 1 subunit 1 | 50.67 | 3 | 0.308 | 0.461 | 0.296 | 0.417 | transcription factor |
| TRIM21 | P19474 | E3 ubiquitin-protein ligase TRIM21 | 7.79 | 3 | 0.310 | 0.382 | 0.259 | 0.340 | ubiquitin-protein ligase |
| OAS3 | Q9Y6K5 | 2′-5′-oligoadenylate synthase 3 | 30.08 | 29 | 0.316 | 0.306 | 0.246 | 0.237 | nucleotidyltransferase, defense/immunity protein |
| SLC4A4 | Q9Y6R1 | Electrogenic sodium bicarbonate cotransporter 1 | 9.64 | 8 | 0.323 | 0.290 | 0.164 | 0.214 | transporter |
| TAPBP | O15533 | Tapasin | 25.00 | 7 | 0.324 | 0.397 | 0.318 | 0.333 | immunoglobulin receptor superfamily |
| KRT1 | P04264 | Keratin, type II cytoskeletal 1 | 36.49 | 18 | 0.325 | 0.273 | 0.550 | 0.447 | structural protein |
| DTX3L | Q8TDB6 | E3 ubiquitin-protein ligase DTX3L | 25.81 | 12 | 0.326 | 0.426 | 0.375 | 0.377 | ubiquitin-protein ligase |
| TAP2 | Q03519 | Antigen peptide transporter 2 | 22.16 | 10 | 0.350 | 0.350 | 0.280 | 0.270 | ATP-binding cassette (ABC) transporter |
| GBP1 | P32455 | Interferon-induced guanylate-binding protein 1 | 28.38 | 13 | 0.362 | 0.346 | 0.296 | 0.202 | heterotrimeric G-protein |
| KRT9 | P35527 | Keratin, type I cytoskeletal 9 | 35.47 | 15 | 0.363 | 0.409 | 0.632 | 0.759 | structural protein |
| AGTRAP | Q6RW13 | Type-1 angiotensin II receptor-associated protein | 13.84 | 1 | 0.383 | 0.479 | 0.557 | 0.604 | response to hypoxia |
| PARP9 | Q8IXQ6 | Poly [ADP-ribose] polymerase 9 | 16.28 | 12 | 0.387 | 0.392 | 0.395 | 0.359 | nucleic acid binding |
| HLA-B | P30466 | HLA class I histocompatibility antigen, B-18 alpha chain | 57.73 | 1 | 0.396 | 0.414 | 0.307 | 0.344 | immunoglobulin receptor superfamily |
| IRF9 | Q00978 | Interferon regulatory factor 9 | 7.38 | 3 | 0.405 | 0.588 | 0.318 | 0.377 | immune response |
| C19orf66 | Q9NUL5 | UPF0515 protein C19orf66 | 16.15 | 3 | 0.405 | 0.480 | 0.214 | 0.395 | no function identified yet |
| NT5E | P21589 | 5′-nucleotidase | 48.08 | 25 | 0.408 | 0.428 | 0.353 | 0.361 | nucleotide phosphatase |
| STAT1 | P42224 | Signal transducer and activator of transcription 1-alpha/beta | 53.33 | 36 | 0.408 | 0.422 | 0.369 | 0.381 | transcription factor, nucleic acid binding |
| KRT2 | P35908 | Keratin, type II cytoskeletal 2 epidermal | 7.82 | 3 | 0.411 | 0.329 | 0.428 | 0.471 | structural protein |
| SP100 | P23497 | Nuclear autoantigen Sp-100 | 9.56 | 6 | 0.412 | 0.438 | 0.352 | 0.363 | HMG box transcription factor, signaling molecule |
| B2M | P61769 | Beta-2-microglobulin | 37.82 | 4 | 0.419 | 0.413 | 0.369 | 0.332 | major histocompatibility complex antigen |
| ALB | A0A0C4DGB6 | Serum albumin | 16.89 | 9 | 0.427 | 0.455 | 0.668 | 0.654 | transfer/carrier protein |
| BANF1 | O75531 | Barrier-to-autointegration factor | 34.83 | 2 | 0.429 | 0.467 | 0.308 | 0.408 | DNA binding, DNA integration |
| IFIT5 | Q13325 | Interferon-induced protein with tetratricopeptide repeats 5 | 19.71 | 7 | 0.438 | 0.497 | 0.477 | 0.435 | RNA-binding |
| ERAP2 | Q6P179 | Endoplasmic reticulum aminopeptidase 2 | 6.25 | 5 | 0.461 | 0.527 | 0.371 | 0.457 | metalloprotease |
| HLA-A | P01892 | HLA class I histocompatibility antigen, A-2 alpha chain | 64.38 | 15 | 0.465 | 0.509 | 0.415 | 0.447 | immunoglobulin receptor superfamily |
| NDRG1 | Q92597 | Protein NDRG1 | 27.41 | 6 | 0.468 | 0.541 | 0.260 | 0.246 | stress-responsive protein |
| STAT2 | P52630 | Signal transducer and activator of transcription 2 | 10.93 | 5 | 0.478 | 0.617 | 0.347 | 0.442 | transcription factor, nucleic acid binding |
| HLA-E | P13747 | HLA class I histocompatibility antigen, alpha chain E | 24.02 | 2 | 0.479 | 0.319 | 0.472 | 0.433 | immunoglobulin receptor superfamily |
| ATP6V0C | P27449 | V-type proton ATPase 16 kDa proteolipid subunit | 11.61 | 1 | 0.484 | 0.353 | 0.906 | 0.805 | hydrolase, ATP synthase |
| UCHL3 | P15374 | Ubiquitin carboxyl-terminal hydrolase isozyme L3 | 15.65 | 2 | 0.492 | 0.459 | 0.558 | 0.220 | cysteine protease |
| EPN2 | F6PQP6 | Epsin-2 (Fragment) | 19.56 | 7 | 0.496 | 0.576 | 0.262 | 0.295 | endocytosis |
| DBI | P07108 | Acyl-CoA-binding protein | 65.52 | 6 | 0.501 | 0.502 | 0.263 | 0.148 | transfer/carrier protein |
| SP110 | G5E9C0 | SP110 nuclear body protein, isoform CRA_b | 5.48 | 2 | 0.506 | 0.473 | 0.464 | 0.439 | HMG box transcription factor, signaling molecule |
| TCEAL3 | Q969E4 | Transcription elongation factor A protein-like 3 | 16.50 | 2 | 0.507 | 0.478 | 0.354 | 0.307 | transcription factor |
| LGALS3BP | Q08380 | Galectin-3-binding protein | 39.83 | 19 | 0.507 | 0.533 | 0.427 | 0.457 | receptor, serine protease |
| UBE2L6 | O14933 | Ubiquitin/ISG15-conjugating enzyme E2 L6 | 59.48 | 5 | 0.517 | 0.407 | 0.382 | 0.317 | ligase |
| SMYD2 | Q9NRG4 | N-lysine methyltransferase SMYD2 | 7.39 | 3 | 0.519 | 0.669 | 0.327 | 0.243 | transcription cofactor |
| TREX1 | Q9NSU2 | Three-prime repair exonuclease 1 | 7.86 | 2 | 0.526 | 0.463 | 0.482 | 0.389 | catalytic activityi |
| AK4 | P27144 | Adenylate kinase 4, mitochondrial | 49.33 | 8 | 0.529 | 0.500 | 0.381 | 0.422 | nucleotide kinase |
| FAM96B | J3KS95 | Mitotic spindle-associated MMXD complex subunit MIP18 (Fragment) | 23.58 | 2 | 0.539 | 0.421 | 0.452 | 0.473 | iron-sulfur cluster assembly |
| DPP7 | Q9UHL4 | Dipeptidyl peptidase 2 | 35.37 | 12 | 0.541 | 0.581 | 0.365 | 0.431 | serine protease |
| PML | P29590 | Protein PML | 33.79 | 22 | 0.545 | 0.558 | 0.424 | 0.392 | activator |
| AGA | P20933 | N(4)-(beta-N-acetylglucosaminyl)-l-asparaginase | 24.86 | 5 | 0.551 | 0.635 | 0.415 | 0.490 | protease |
| EPHA2 | P29317 | Ephrin type-A receptor 2 | 19.67 | 15 | 0.555 | 0.523 | 0.386 | 0.395 | nervous system development |
| SERPINI1 | Q99574 | Neuroserpin | 8.78 | 3 | 0.564 | 0.757 | 0.257 | 0.217 | serine protease inhibitor |
| PAPSS2 | O95340 | Bifunctional 3′-phosphoadenosine 5′-phosphosulfate synthase 2 | 37.30 | 18 | 0.567 | 0.544 | 0.336 | 0.276 | nucleotidyltransferase |
| IDUA | P35475 | Alpha-l-iduronidase | 31.85 | 16 | 0.572 | 0.605 | 0.416 | 0.479 | glycosidase |
| GLA | P06280 | Alpha-galactosidase A | 25.64 | 8 | 0.574 | 0.625 | 0.464 | 0.435 | glycosidase, hydrolase |
| SGSH | P51688 | N-sulphoglucosamine sulphohydrolase | 29.68 | 11 | 0.578 | 0.515 | 0.353 | 0.410 | hydrolase |
| GAA | P10253 | Lysosomal alpha-glucosidase | 24.37 | 19 | 0.584 | 0.657 | 0.426 | 0.461 | glucosidase |
| CHSY3 | Q70JA7 | Chondroitin sulfate synthase 3 | 6.92 | 6 | 0.587 | 0.483 | 0.361 | 0.320 | glycosyltransferase |
| ACP5 | K7EIP0 | Tartrate-resistant acid phosphatase type 5 (Fragment) | 36.54 | 1 | 0.587 | 0.544 | 0.313 | 0.246 | glycosylated monomeric metalloprotein enzyme |
| PSMB8 | P28062 | Proteasome subunit beta type-8 | 39.13 | 8 | 0.590 | 0.552 | 0.455 | 0.493 | endopeptidase activity |
| SPTBN2 | O15020 | Spectrin beta chain, non-erythrocytic 2 | 4.35 | 2 | 0.593 | 0.738 | 0.389 | 0.358 | non-motor actin binding protein |
| PGM2L1 | Q6PCE3 | Glucose 1,6-bisphosphate synthase | 48.07 | 30 | 0.598 | 0.627 | 0.492 | 0.448 | glycosyltransferase, mutase |
| SAMD9L | Q8IVG5 | Sterile alpha motif domain-containing protein 9-like | 4.67 | 6 | 0.611 | 0.547 | 0.489 | 0.416 | regulation of protein catabolic process |
| CSTB | P04080 | Cystatin-B | 45.92 | 3 | 0.615 | 0.680 | 0.328 | 0.380 | cysteine protease inhibitor |
| LGMN | Q99538 | Legumain | 10.39 | 4 | 0.618 | 0.636 | 0.483 | 0.499 | cysteine protease |
| CPQ | Q9Y646 | Carboxypeptidase Q | 20.55 | 7 | 0.620 | 0.631 | 0.406 | 0.452 | carboxypeptidase activity |
| CTSA | P10619 | Lysosomal protective protein | 18.75 | 9 | 0.626 | 0.667 | 0.418 | 0.422 | serine protease |
| NAGA | P17050 | Alpha-N-acetylgalactosaminidase | 11.92 | 3 | 0.626 | 0.648 | 0.480 | 0.287 | deacetylase |
| ENO2 | P09104 | Gamma-enolase | 60.83 | 11 | 0.631 | 0.676 | 0.436 | 0.494 | lyase |
| GALNS | P34059 | N-acetylgalactosamine-6-sulfatase | 8.62 | 5 | 0.633 | 0.687 | 0.436 | 0.377 | hydrolase |
| KCTD12 | Q96CX2 | BTB/POZ domain-containing protein KCTD12 | 33.23 | 11 | 0.634 | 0.624 | 0.446 | 0.480 | enzyme modulator |
| GOLIM4 | O00461 | Golgi integral membrane protein 4 | 18.25 | 11 | 0.638 | 0.671 | 0.391 | 0.416 | transport |
| NMRK1 | B3KN26 | Nicotinamide riboside kinase 1 | 12.26 | 1 | 0.641 | 0.527 | 0.422 | 0.402 | kinase |
| RNASET2 | D6REQ6 | Ribonuclease T2 | 19.27 | 4 | 0.643 | 0.545 | 0.398 | 0.400 | endoribonuclease activity |
| TUBB2B | Q9BVA1 | Tubulin beta-2B chain | 74.16 | 1 | 0.643 | 0.545 | 0.424 | 0.318 | tubulin |
| MTAP | Q13126 | S-methyl-5′-thioadenosine phosphorylase | 71.38 | 15 | 0.645 | 0.683 | 0.484 | 0.492 | phosphorylase |
| NAGLU | P54802 | Alpha-N-acetylglucosaminidase | 26.11 | 13 | 0.646 | 0.706 | 0.466 | 0.478 | glycosidase, hydrolase |
| TXNIP | Q9H3M7 | Thioredoxin-interacting protein | 16.11 | 6 | 0.650 | 0.441 | 0.479 | 0.351 | transcription regulation, oxidative stress mediator |
| BCAR3 | O75815 | Breast cancer anti-estrogen resistance protein 3 | 7.88 | 4 | 0.652 | 0.285 | 0.162 | 0.272 | guanine-nucleotide releasing factor |
| GUSB | P08236 | Beta-glucuronidase | 26.42 | 16 | 0.678 | 0.649 | 0.495 | 0.452 | galactosidase |
| PGK1 | P00558 | Phosphoglycerate kinase 1 | 84.41 | 31 | 0.686 | 0.647 | 0.461 | 0.437 | carbohydrate kinase |
| H6PD | O95479 | GDH/6PGL endoplasmic bifunctional protein | 35.65 | 23 | 0.708 | 0.729 | 0.477 | 0.483 | dehydrogenase |
| CSRP1 | P21291 | Cysteine and glycine-rich protein 1 | 64.25 | 9 | 0.711 | 0.670 | 0.426 | 0.427 | actin family cytoskeletal protein |
| CPVL | Q9H3G5 | Probable serine carboxypeptidase CPVL | 21.22 | 9 | 0.711 | 0.640 | 0.480 | 0.453 | serine protease |
| NNMT | P40261 | Nicotinamide N-methyltransferase | 56.06 | 10 | 0.713 | 0.667 | 0.335 | 0.333 | methyltransferase |
| EXTL3 | O43909 | Exostosin-like 3 | 15.34 | 13 | 0.737 | 0.807 | 0.433 | 0.472 | glycosyltransferase |
| VLDLR | P98155 | Very low-density lipoprotein receptor | 8.48 | 6 | 0.738 | 0.699 | 0.460 | 0.478 | receptor, extracellular matrix glycoprotein |
| MMP14 | P50281 | Matrix metalloproteinase-14 | 19.76 | 11 | 0.748 | 0.800 | 0.344 | 0.361 | hydrolase, metalloprotease, protease |
| OSTF1 | Q92882 | Osteoclast-stimulating factor 1 | 53.74 | 9 | 0.750 | 0.649 | 0.459 | 0.463 | signal transduction |
| AKAP2 | Q9Y2D5 | A-kinase anchor protein 2 | 15.83 | 7 | 0.751 | 0.773 | 0.410 | 0.468 | regulation of cell cycle, apoptosis process |
| SIAE | Q9HAT2 | Sialate O-acetylesterase | 12.05 | 5 | 0.769 | 0.725 | 0.346 | 0.294 | esterase |
| MRC2 | Q9UBG0 | C-type mannose receptor 2 | 11.36 | 13 | 0.778 | 0.867 | 0.379 | 0.454 | receptor |
| IDS | P22304 | Iduronate 2-sulfatase | 23.82 | 9 | 0.784 | 0.812 | 0.425 | 0.465 | hydrolase |
| CNTNAP1 | P78357 | Contactin-associated protein 1 | 2.02 | 2 | 0.792 | 0.638 | 0.403 | 0.436 | transporter, membrane-bound signaling molecule, receptor |
| AKR1C3 | S4R3Z2 | Aldo-keto reductase family 1 member C3 | 6.67 | 1 | 0.828 | 0.645 | 0.305 | 0.337 | reductase |
| AMDHD2 | Q9Y303 | Putative N-acetylglucosamine-6-phosphate deacetylase | 8.07 | 2 | 0.840 | 0.548 | 0.465 | 0.473 | deacetylase |
| MANBA | O00462 | Beta-mannosidase | 6.60 | 3 | 0.901 | 0.799 | 0.356 | 0.482 | galactosidase |
| SH3BP5L | Q7L8J4 | SH3 domain-binding protein 5-like | 5.09 | 2 | 0.912 | 0.931 | 0.354 | 0.411 | protein kinase inhibitor |
| LRP1 | Q07954 | Prolow-density lipoprotein receptor-related protein 1 | 0.62 | 3 | 1.068 | 0.617 | 0.381 | 0.478 | receptor, extracellular matrix glycoprotein |
| ATF7IP | F5GYR7 | Activating transcription factor 7-interacting protein 1 (Fragment) | 9.38 | 1 | ND | 0.447 | 0.441 | 0.146 | transcription regulation |
| VPS29 | Q9UBQ0 | Vacuolar protein sorting-associated protein 29 | 56.04 | 1 | ND | 0.922 | 0.473 | 0.474 | vesicle coat protein |
| Pathways | p Value | Differentially Expressed Proteins Involved in This Pathway |
|---|---|---|
| Lysosome | 1.11 × 10−8 | SGSH, AGA, NAGLU, GUSB, LGMN, ACP5, CTSA, MANBA, ATP6V0C, GLA, IDS, GALNS, NAGA, GAA, IDUA |
| Glycosaminoglycan degradation | 2.53 × 10−5 | SGSH, NAGLU, IDS, GUSB, GALNS, IDUA |
| Antigen processing and presentation | 5.99 × 10−4 | TAP2, LGMN, TAP1, HLA-A, HLA-B, HLA-E, TAPBP, B2M |
| DNA replication | 3.51 × 10−3 | MCM7, MCM2, MCM4, MCM5, MCM6 |
| Type I diabetes mellitus | 3.75 × 10−2 | INS, HLA-A, HLA-B, HLA-E |
| Glycolysis/Gluconeogenesis | 8.93 × 10−2 | TPI1, ENO2, PGK1, ENO1 |
| Gel No. | Cy2 | Cy3 | Cy5 |
|---|---|---|---|
| Gel 01 | Standard | A1 | B2 |
| Gel 02 | Standard | B1 | C3 |
| Gel 03 | Standard | C2 | D3 |
| Gel 04 | Standard | D2 | A2 |
| Gel 05 | Standard | A3 | C1 |
| Gel 06 | Standard | B3 | D1 |
| Samples | Forward | Reverse |
|---|---|---|
| control group | Heavy (H) | Light (L) |
| 10 μg/mL dose group | Light (L) | Medium (M) |
| 40 μg/mL dose group | Medium (M) | Heavy (H) |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Dai, J.; Tang, Y.; Li, L.; Jin, G. Quantitative Proteomic Profiling of Tachyplesin I Targets in U251 Gliomaspheres. Mar. Drugs 2017, 15, 20. https://doi.org/10.3390/md15010020
Li X, Dai J, Tang Y, Li L, Jin G. Quantitative Proteomic Profiling of Tachyplesin I Targets in U251 Gliomaspheres. Marine Drugs. 2017; 15(1):20. https://doi.org/10.3390/md15010020
Chicago/Turabian StyleLi, Xuan, Jianguo Dai, Yongjun Tang, Lulu Li, and Gang Jin. 2017. "Quantitative Proteomic Profiling of Tachyplesin I Targets in U251 Gliomaspheres" Marine Drugs 15, no. 1: 20. https://doi.org/10.3390/md15010020
APA StyleLi, X., Dai, J., Tang, Y., Li, L., & Jin, G. (2017). Quantitative Proteomic Profiling of Tachyplesin I Targets in U251 Gliomaspheres. Marine Drugs, 15(1), 20. https://doi.org/10.3390/md15010020