Discovering the Biological Target of 5-epi-Sinuleptolide Using a Combination of Proteomic Approaches





 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
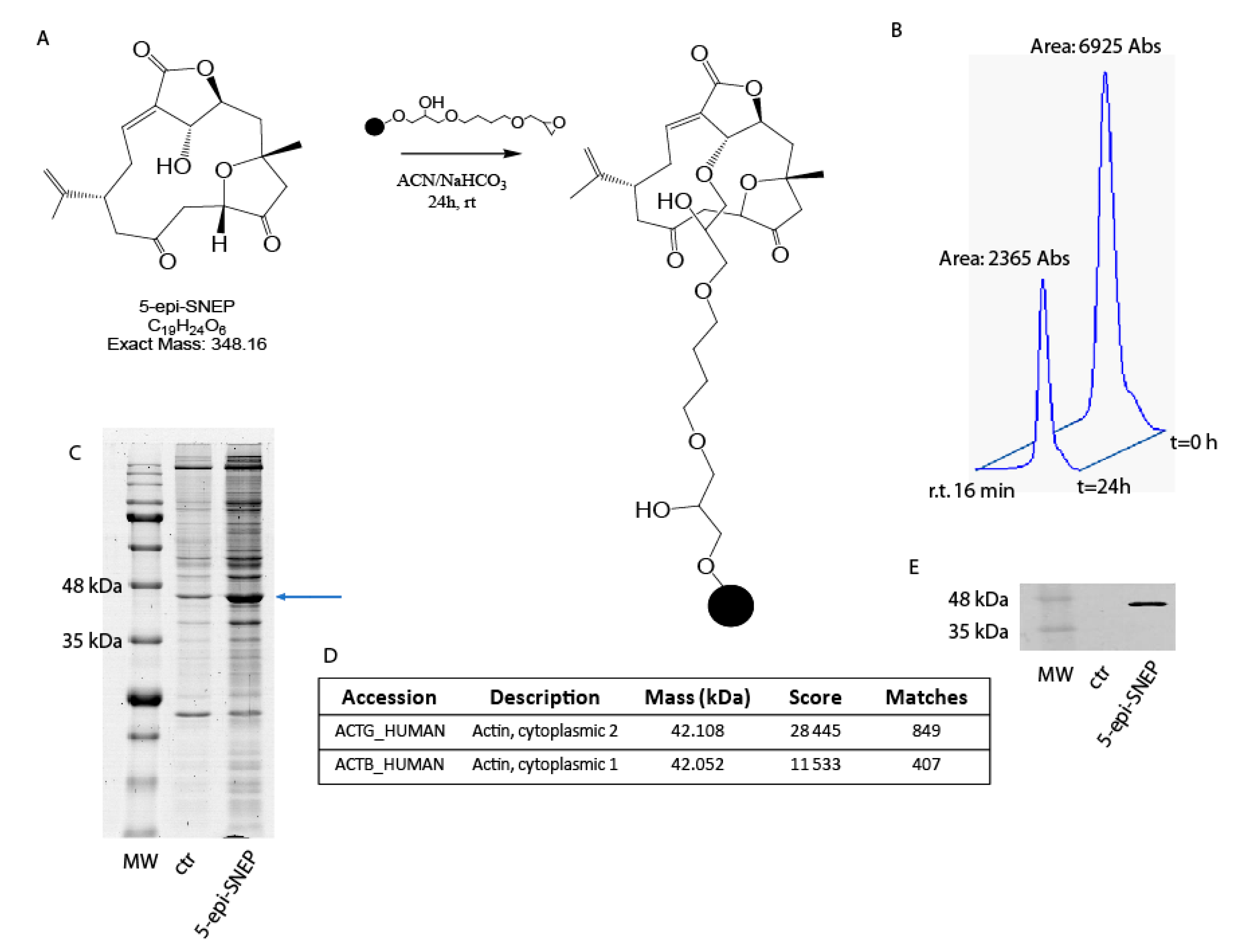
2.1. Generation of a Functional 5-epi-SNEP Affinity Matrix
2.2. Identification of 5-epi-SNEPinteractome by Nano-LC-MSMS Analysis and Western Blotting
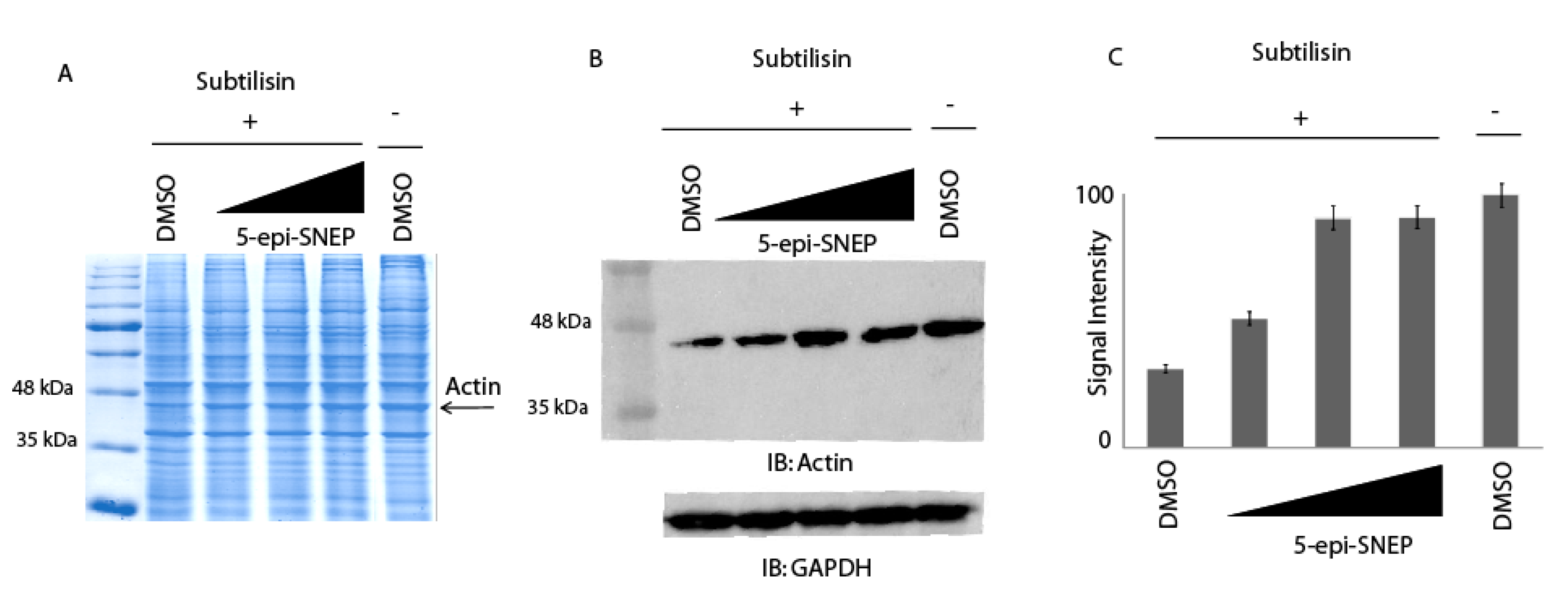
2.3. Application of DARTS Protocol on 5-epi-SNEP Target Protein
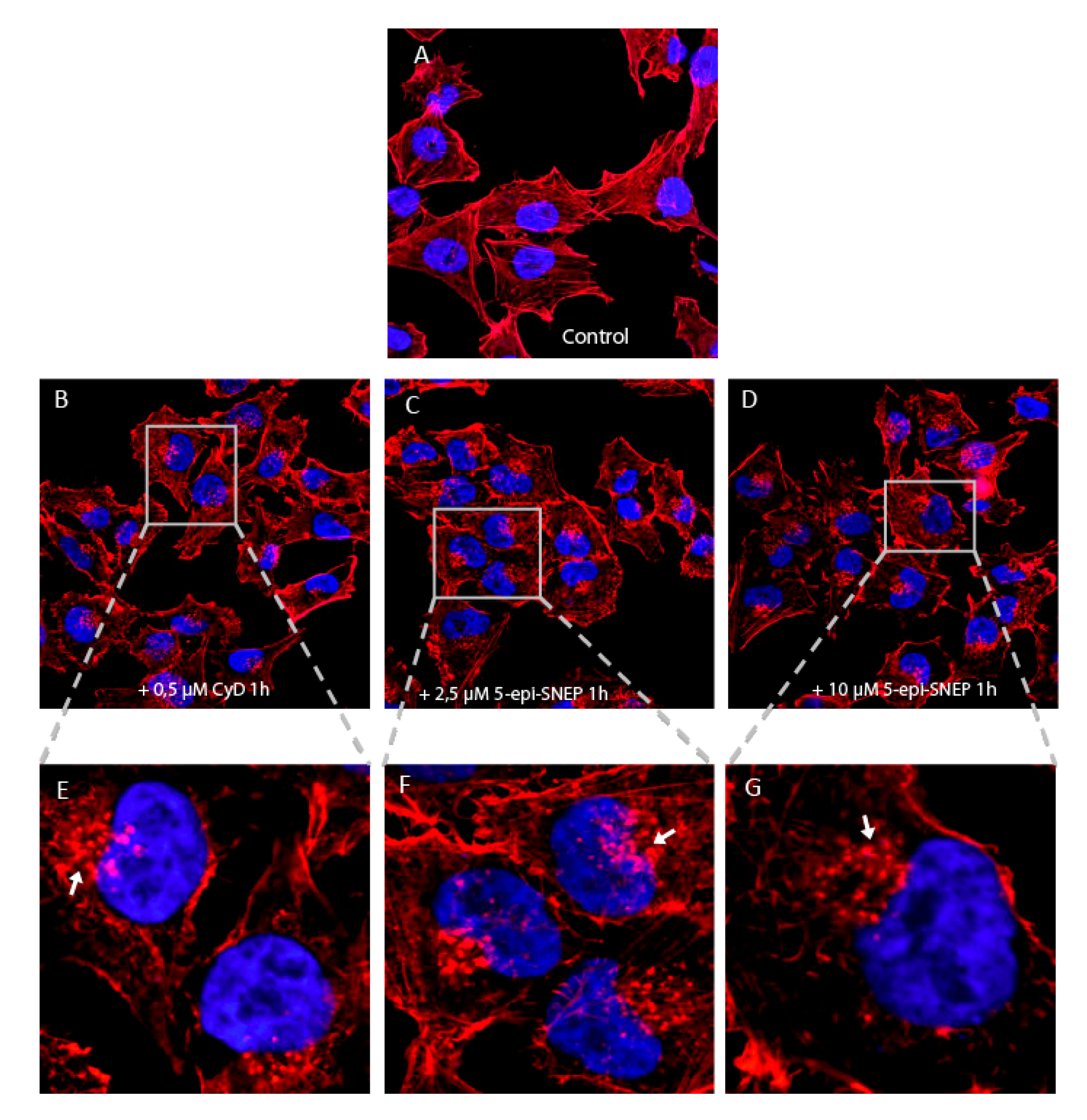
2.4. Biological Evaluation of 5-epi-SNEP Action on Actin by in-Cell Assays
3. Discussion and Conclusions
4. Materials and Methods
4.1. Generation of a Functional 5-epi-SNEP Affinity Matrix
4.2. Identification of 5-epi-SNEP Interactome by Nano-LC-MSMS Analysis and Western Blotting
4.3. Optimization of DARST Protocol on 5-epi-SNEP Target Protein
4.4. Biological Evaluation of 5-epi-SNEP Action by in-Cell Assays
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jha, R.K.; Xu, Z.R. Biomedical Compounds from Marine organisms. Mar. Drugs 2004, 2, 123–146. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Marine-Sourced Anti-Cancer and Cancer Pain Control Agents in Clinical and Late Preclinical Development. Mar. Drugs 2014, 12, 255–278. [Google Scholar] [CrossRef] [PubMed]
- Radhika, P.; Rao, P.R.; Archana, J.; Rao, N.K. Anti-inflammatory activity of a new sphingosine derivative and cembrenoid diterpene (lobohedleolide) isolated from marine soft corals of Sinularia crassa TIXIER-DURIVAULT and Lobophytum species of the Andaman and Nicobar Islands. Biol. Pharm. Bull. 2005, 28, 1311–1313. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.H.; Wang, G.H.; Hung, W.J.; Lin, R.J.; Cheng, D.L.; Chou, T.H. Apoptosis effect of Sinularia leptoclados, S. depressan and S. inflate extracts in human oral squamous cell carcinomas. J. Taiwan Inst. Chem. Eng. 2010, 41, 86–91. [Google Scholar] [CrossRef]
- Kamel, H.N.; Slattery, M. Terpenoids of Sinularia: Chemistry and Biomedical Applications. Pharm. Biol. 2005, 43, 253–269. [Google Scholar] [CrossRef]
- Takaki, H.; Koganemaru, R.; Iwakawa, Y.; Higuchi, R.; Miyamoto, T. Inhibitory effect of norditerpenes on LPS-induced TNF-alpha production from the Okinawan soft coral, Sinularia sp. Biol. Pharm. Bull. 2003, 26, 380–382. [Google Scholar] [CrossRef] [PubMed]
- Sheu, J.H.; Ahmed, A.F.; Shiue, R.T.; Dai, C.F.; Kuo, Y.H. Scabrolides A-D, four new norditerpenoids isolated from the soft coral Sinularia scabra. J. Nat. Prod. 2002, 65, 1904–1908. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.F.; Shiue, R.T.; Wang, G.H.; Dai, C.F.; Kuo, Y.H.; Sheu, J.H. Five novel norcembranoids from Sinularia leptoclados and S. parva. Tetrahedron 2003, 59, 7337–7344. [Google Scholar] [CrossRef]
- Chang, Y.T.; Huang, C.Y.; Li, K.T.; Li, R.N.; Liaw, C.C.; Wu, S.H.; Liu, J.R.; Sheu, J.H.; Chang, H.W. Sinuleptolide inhibits proliferation of oral cancer Ca9-22 cells involving apoptosis, oxidative stress, and DNA damage. Arch. Oral Biol. 2016, 66, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.H.; Wang, G.H.; Chou, T.H.; Wang, S.H.; Lin, R.J.; Chan, L.P.; So, E.C.; Sheu, J.H. 5-epi-Sinuleptolide induces cell cycle arrest and apoptosis through tumor necrosis factor/mitochondria-mediated caspase signaling pathway in human skin cancer cells. Biochim. Biophys. Acta 2012, 1820, 1149–1157. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.J.; Chen, Y.C.; El-Shazly, M.; Du, Y.C.; Su, J.H.; Tsao, C.W.; Yen, W.H.; Chang, W.B.; Su, Y.D.; Yeh, Y.T.; et al. 5-Episinuleptolide acetate, a norcembranoidal diterpene from the formosan soft coral Sinularia sp., induces leukemia cell apoptosis through Hsp90 inhibition. Molecules 2013, 18, 2924–2933. [Google Scholar] [CrossRef] [PubMed]
- Lillsunde, K.E.; Festa, C.; Adel, H.; De Marino, S.; Lombardi, V.; Tilvi, S.; Nawrot, D.A.; Zampella, A.; D’Souza, L.; D’Auria, M.V.; et al. Bioactive cembrane derivatives from the Indian Ocean soft coral, Sinularia kavarattiensis. Mar. Drugs 2014, 12, 4045–4068. [Google Scholar] [CrossRef] [PubMed]
- Margarucci, L.; Monti, M.C.; Fontanella, B.; Riccio, R.; Casapullo, A. Chemical proteomics reveals bolinaquinone as a clathrin-mediated endocytosis inhibitor. Mol. Biosyst. 2011, 7, 480–485. [Google Scholar] [CrossRef] [PubMed]
- Margarucci, L.; Monti, M.C.; Cassiano, C.; Mozzicafreddo, M.; Angeletti, M.; Riccio, R.; Tosco, A.; Casapullo, A. Chemical proteomics-driven discovery of oleocanthal as an Hsp90 inhibitor. Chem. Commun. 2013, 49, 5844–5846. [Google Scholar] [CrossRef] [PubMed]
- Lomenick, B.; Hao, R.; Jonai, N.; Chin, R.M.; Aghajan, M.; Warburton, S.; Wang, J.; Wu, R.P.; Gomez, F.; Loo, J.A.; et al. Target identification using drug affinity responsive target stability (DARTS). Proc. Natl. Acad. Sci. USA 2009, 106, 21984–21989. [Google Scholar] [CrossRef] [PubMed]
- Pai, M.Y.; Lomenick, B.; Hwang, H.; Schiestl, R.; McBride, W.; Loo, J.A.; Huang, J. Drug affinity responsive target stability (DARTS) for small-molecule target identification. Methods Mol. Biol. 2015, 1263, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, A.; Tomas, H.; Havlis, J.; Olsen, J.V.; Mann, M. In-gel digestion for mass spectrometric characterization of proteins and proteomes. Nat. Protoc. 2007, 1, 2856–2860. [Google Scholar] [CrossRef] [PubMed]
- Wakatsuki, T.; Schwab, B.; Thompson, N.C.; Elson, E.L. Effects of cytochalasin D and latrunculin B on mechanical properties of cells. J. Cell Sci. 2001, 114, 1025–1029. [Google Scholar] [PubMed]
- Suresh, S. Biomechanics and biophysics of cancer cells. Acta Biomater. 2007, 3, 413–438. [Google Scholar] [CrossRef] [PubMed]
- Weissmann, C.; Brandt, R. Mechanisms of neurodegenerative diseases: Insights from live cell imaging. J. Neurosci. Res. 2008, 86, 504–511. [Google Scholar] [CrossRef] [PubMed]
- Lázaro-Diéguez, F.; Aguado, C.; Mato, E.; Sanchez-Ruiz, Y.; Esteban, I.; Alberch, J.; Knecht, E.; Egea, G. Dynamics of an F-actin aggresome generated by the actin-stabilizing toxin jasplakinolide. J. Cell Sci. 2008, 121, 1415–1425. [Google Scholar] [CrossRef] [PubMed]




© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morretta, E.; Esposito, R.; Festa, C.; Riccio, R.; Casapullo, A.; Monti, M.C. Discovering the Biological Target of 5-epi-Sinuleptolide Using a Combination of Proteomic Approaches. Mar. Drugs 2017, 15, 312. https://doi.org/10.3390/md15100312
Morretta E, Esposito R, Festa C, Riccio R, Casapullo A, Monti MC. Discovering the Biological Target of 5-epi-Sinuleptolide Using a Combination of Proteomic Approaches. Marine Drugs. 2017; 15(10):312. https://doi.org/10.3390/md15100312
Chicago/Turabian StyleMorretta, Elva, Roberta Esposito, Carmen Festa, Raffaele Riccio, Agostino Casapullo, and Maria Chiara Monti. 2017. "Discovering the Biological Target of 5-epi-Sinuleptolide Using a Combination of Proteomic Approaches" Marine Drugs 15, no. 10: 312. https://doi.org/10.3390/md15100312
APA StyleMorretta, E., Esposito, R., Festa, C., Riccio, R., Casapullo, A., & Monti, M. C. (2017). Discovering the Biological Target of 5-epi-Sinuleptolide Using a Combination of Proteomic Approaches. Marine Drugs, 15(10), 312. https://doi.org/10.3390/md15100312