The Holo-Transcriptome of the Zoantharian Protopalythoa variabilis (Cnidaria: Anthozoa): A Plentiful Source of Enzymes for Potential Application in Green Chemistry, Industrial and Pharmaceutical Biotechnology

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Biodiversity in the P. variabilis Holobiont
2.2. Annotation and Functional Prediction
2.2.1. Functional Classification of P. variabilis Predicted Enzymes
2.2.2. Assignment of Metabolic Pathways of the P. variabilis Predicted Enzymes
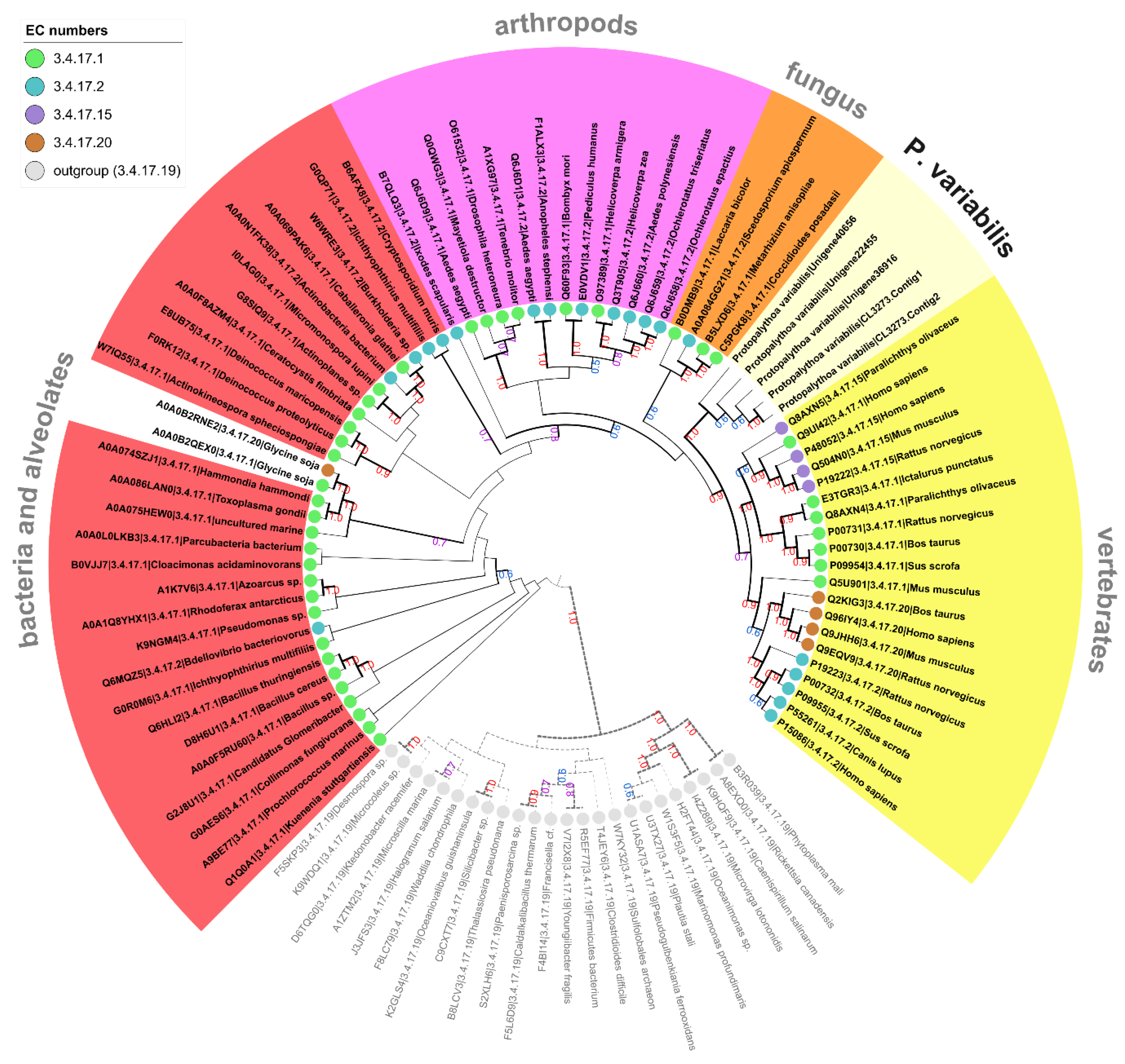
2.3. Biotechnologically Relevant Enzymes from the P. variabilis Holo-Transcriptome
2.3.1. Relevance in the Treatment of Rare Diseases and Other Biomedical and Clinical Applications
2.3.2. Relevance in Colorant, Aromas, Flavor, Fragrance, Cosmetic, and Hygienic Industries
2.3.3. Relevance in Agrochemical, Food and Feed Industries
2.3.4. Relevance in Bioconversion and Biopolymer Synthesis
2.3.5. Relevance in the Cleaning and Recovery Industries
2.3.6. Relevance in Molecular Biology and Analytical Applications
2.4. Prediction of Enzymes with Two or More Activities
2.5. Annotation of Novel Predicted Enzyme Sequences with Partial EC Number
3. Materials and Methods
3.1. Biological Sample
3.2. RNA Library Construction and Origin of Zoantharian RNA Sequences
3.3. Assessment of the Biodiversity Composition of the P. variabilis Holobiont
3.4. Sequence Annotations for Enzyme Precursors in the P. variabilis Holobiont
3.5. Gene Ontology, Enzyme Codes and KEGG Pathway Assignments
3.6. Sequence Alignment and Phylogenetic Inference
3.7. Prediction of Enzymes with Two or More Activities
3.8. Analysis of Predicted Enzyme with Partial EC Number
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Enzymes Market by Type (Industrial, Specialty), by Product (Carbohydrases, Proteases, Lipases, Polymerases & Nucleases), by Application (Food & Beverages, Detergents, Animal Feed, Textile, Paper & Pulp, Nutraceutical, Personal Care & Cosmetics, Wastewater, Research & Biotechnology, Diagnostics, Biocatalyst) and Segment Forecasts to 2024. Available online: https://www.grandviewresearch.com/industry-analysis/enzymes-industry (accessed on 13 June 2018).
- Molecular Biology Enzymes and Kits & Reagents Market Analysis by Product (Kits and Reagents, Enzymes), by Application (PCR, Sequencing, Cloning, Epigenetics), by End User, and Segment Forecasts, 2018–2025. Available online: https://www.grandviewresearch.com/industry-analysis/molecular-biology-enzymes-kits-reagents-market (accessed on 13 June 2018).
- Itoh, T.; Hanefeld, U. Enzyme catalysis in organic synthesis. Green Chem. 2017, 19, 331–332. [Google Scholar] [CrossRef]
- Wenda, S.; Illner, S.; Mell, A.; Kragl, U. Industrial biotechnology-the future of green chemistry? Green Chem. 2011, 13, 3007–3047. [Google Scholar] [CrossRef]
- Yamaguchi, S. The quest for industrial enzymes from microorganisms. Biosci. Biotechnol. Biochem. 2017, 81, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, S.; Ogawa, J.; Kataoka, M.; Kobayashi, M. Screening of novel microbial enzymes for the production of biologically and chemically useful compounds. Adv. Biochem. Eng. Biotechnol. 1997, 58, 45–87. [Google Scholar] [PubMed]
- Wang, J.B.; Li, G.; Reetz, M.T. Enzymatic site-selectivity enabled by structure-guided directed evolution. Chem. Commun. 2017, 53, 3916–3928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hibbert, E.G.; Dalby, P.A. Directed evolution strategies for improved enzymatic performance. Microb. Cell Fact. 2005, 4, 29. [Google Scholar] [CrossRef] [PubMed]
- Madhavan, A.; Sindhu, R.; Parameswaran, B.; Sukumaran, R.K.; Pandey, A. Metagenome Analysis: A Powerful Tool for Enzyme Bioprospecting. Appl. Biochem. Biotechnol. 2017, 183, 636–651. [Google Scholar] [CrossRef] [PubMed]
- Trincone, A. Marine biocatalysts: Enzymatic features and applications. Mar. Drugs 2011, 9, 478–499. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.R.M.; Uria, A.R.; Helfrich, E.J.N.; Milbredt, D.; van Pee, K.H.; Piel, J.; Goss, R.J.M. An Unusual Flavin-Dependent Halogenase from the Metagenome of the Marine Sponge Theonella swinhoei WA. ACS Chem. Biol. 2017, 12, 1281–1287. [Google Scholar] [CrossRef] [PubMed]
- Sarian, F.D.; Janecek, S.; Pijning, T.; Ihsanawati; Nurachman, Z.; Radjasa, O.K.; Dijkhuizen, L.; Natalia, D.; van der Maarel, M.J. A new group of glycoside hydrolase family 13 alpha-amylases with an aberrant catalytic triad. Sci. Rep. 2017, 7, 44230. [Google Scholar] [CrossRef] [PubMed]
- Lima, R.N.; Porto, A.L. Recent Advances in Marine Enzymes for Biotechnological Processes. Adv. Food Nutr. Res. 2016, 78, 153–192. [Google Scholar] [PubMed]
- Trincone, A. Enzymatic Processes in Marine Biotechnology. Mar. Drugs 2017, 15, 93. [Google Scholar] [CrossRef] [PubMed]
- Leal, M.C.; Puga, J.; Serodio, J.; Gomes, N.C.; Calado, R. Trends in the discovery of new marine natural products from invertebrates over the last two decades—Where and what are we bioprospecting? PLoS ONE 2012, 7, e30580. [Google Scholar] [CrossRef] [PubMed]
- Lindequist, U. Marine-Derived Pharmaceuticals—Challenges and Opportunities. Biomol. Ther. 2016, 24, 561–571. [Google Scholar] [CrossRef] [PubMed]
- Ryu, T.; Seridi, L.; Moitinho-Silva, L.; Oates, M.; Liew, Y.J.; Mavromatis, C.; Wang, X.; Haywood, A.; Lafi, F.F.; Kupresanin, M.; et al. Hologenome analysis of two marine sponges with different microbiomes. BMC Genomics 2016, 17, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macintyre, L.; Zhang, T.; Viegelmann, C.; Martinez, I.J.; Cheng, C.; Dowdells, C.; Abdelmohsen, U.R.; Gernert, C.; Hentschel, U.; Edrada-Ebel, R. Metabolomic tools for secondary metabolite discovery from marine microbial symbionts. Mar. Drugs 2014, 12, 3416–3448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunlap, W.C.; Battershill, C.N.; Liptrot, C.H.; Cobb, R.E.; Bourne, D.G.; Jaspars, M.; Long, P.F.; Newman, D.J. Biomedicinals from the phytosymbionts of marine invertebrates: A molecular approach. Methods 2007, 42, 358–376. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Y.; Sung, P.J.; Uvarani, C.; Su, J.H.; Lu, M.C.; Hwang, T.L.; Dai, C.F.; Wu, S.L.; Sheu, J.H. Glaucumolides A and B, Biscembranoids with New Structural Type from a Cultured Soft Coral Sarcophyton glaucum. Sci. Rep. 2015, 5, 15624. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Y.; Su, J.H.; Liaw, C.C.; Sung, P.J.; Chiang, P.L.; Hwang, T.L.; Dai, C.F.; Sheu, J.H. Bioactive Steroids with Methyl Ester Group in the Side Chain from a Reef Soft Coral Sinularia brassica Cultured in a Tank. Mar. Drugs 2017, 15, 280. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, T.H.; Sung, C.S.; Lan, Y.H.; Wang, Y.C.; Lu, M.C.; Wen, Z.H.; Wu, Y.C.; Sung, P.J. New Anti-Inflammatory Cembranes from the Cultured Soft Coral Nephthea columnaris. Mar. Drugs 2015, 13, 3443–3453. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, M.; Martinez-Martinez, M.; Bargiela, R.; Streit, W.R.; Golyshina, O.V.; Golyshin, P.N. Estimating the success of enzyme bioprospecting through metagenomics: Current status and future trends. Microb. Biotechnol. 2016, 9, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Morlighem, J.R.; Zhou, H.; Lima, E.P.; Gomes, P.B.; Cai, J.; Lou, I.; Perez, C.D.; Lee, S.M.; Radis-Baptista, G. The Transcriptome of the Zoanthid Protopalythoa variabilis (Cnidaria, Anthozoa) Predicts a Basal Repertoire of Toxin-like and Venom-Auxiliary Polypeptides. Genome Biol. Evol. 2016, 8, 3045–3064. [Google Scholar] [CrossRef] [PubMed]
- Rabelo, E.F.; Soares, M.D.O.; Bezerra, L.E.A.; Matthews-Cascon, H. Distribution pattern of zoanthids (Cnidaria: Zoantharia) on a tropical reef. Mar. Biol. Res. 2015, 11, 584–592. [Google Scholar] [CrossRef]
- Rabelo, E.; Rocha, L.; Colares, G.; Araújo Bomfim, T.; Lúcia Rodrigues Nogueira, V.; Katzenberger, M.; Matthews-Cascon, H.; Melo, V. Symbiodinium Diversity Associated with Zoanthids (Cnidaria: Hexacorallia) in Northeastern Brazil; Springer: Dordrecht, The Netherlands, 2015. [Google Scholar]
- Sun, W.; Zhang, F.; He, L.; Li, Z. Pyrosequencing reveals diverse microbial community associated with the zoanthid Palythoa australiae from the South China Sea. Microb. Ecol. 2014, 67, 942–950. [Google Scholar] [CrossRef] [PubMed]
- Chu, N.D.; Vollmer, S.V. Caribbean corals house shared and host-specific microbial symbionts over time and space. Environ. Microbiol. Rep. 2016, 8, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M.; Snader, K.M. The influence of natural products upon drug discovery. Nat. Prod. Rep. 2000, 17, 215–234. [Google Scholar] [CrossRef] [PubMed]
- Urbarova, I.; Karlsen, B.O.; Okkenhaug, S.; Seternes, O.M.; Johansen, S.D.; Emblem, A. Digital marine bioprospecting: Mining new neurotoxin drug candidates from the transcriptomes of cold-water sea anemones. Mar. Drugs 2012, 10, 2265–2279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, D.A.; Walz, M.E.; Weil, E.; Tonellato, P.; Smith, M.C. RNA-Seq of the Caribbean reef-building coral Orbicella faveolata (Scleractinia-Merulinidae) under bleaching and disease stress expands models of coral innate immunity. PeerJ 2016, 4, e1616. [Google Scholar] [CrossRef] [PubMed]
- Ponce, D.; Brinkman, D.L.; Potriquet, J.; Mulvenna, J. Tentacle Transcriptome and Venom Proteome of the Pacific Sea Nettle, Chrysaora fuscescens (Cnidaria: Scyphozoa). Toxins 2016, 8, 102. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, Y.; Watanabe, T.; Takazawa, M.; Ohara, O.; Kubota, S. De Novo Assembly of the Transcriptome of Turritopsis, a Jellyfish that Repeatedly Rejuvenates. Zool. Sci. 2016, 33, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Valmsen, K.; Jarving, I.; Boeglin, W.E.; Varvas, K.; Koljak, R.; Pehk, T.; Brash, A.R.; Samel, N. The origin of 15R-prostaglandins in the Caribbean coral Plexaura homomalla: Molecular cloning and expression of a novel cyclooxygenase. Proc. Natl. Acad. Sci. USA 2001, 98, 7700–7705. [Google Scholar] [CrossRef] [PubMed]
- Uria, A.R.; Zilda, D.S. Metagenomics-Guided Mining of Commercially Useful Biocatalysts from Marine Microorganisms. Adv. Food Nutr. Res. 2016, 78, 1–26. [Google Scholar] [PubMed]
- Popovic, A.; Tchigvintsev, A.; Tran, H.; Chernikova, T.N.; Golyshina, O.V.; Yakimov, M.M.; Golyshin, P.N.; Yakunin, A.F. Metagenomics as a Tool for Enzyme Discovery: Hydrolytic Enzymes from Marine-Related Metagenomes. Adv. Exp. Med. Biol. 2015, 883, 1–20. [Google Scholar] [PubMed]
- Sukul, P.; Schakermann, S.; Bandow, J.E.; Kusnezowa, A.; Nowrousian, M.; Leichert, L.I. Simple discovery of bacterial biocatalysts from environmental samples through functional metaproteomics. Microbiome 2017, 5, 28. [Google Scholar] [CrossRef] [PubMed]
- Sturmberger, L.; Wallace, P.W.; Glieder, A.; Birner-Gruenberger, R. Synergism of proteomics and mRNA sequencing for enzyme discovery. J. Biotechnol. 2016, 235, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Berglund, P. Transaminase biocatalysis: Optimization and application. Green Chem. 2017, 19, 333–360. [Google Scholar] [CrossRef]
- McGrath, B.M.; Walsh, G. Directory of Therapeutic Enzymes; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2005. [Google Scholar]
- Sun, Z.; Ilie, A.; Reetz, M.T. Towards the Production of Universal Blood by Structure-Guided Directed Evolution of Glycoside Hydrolases. Angew. Chem. 2015, 54, 9158–9160. [Google Scholar] [CrossRef] [PubMed]
- Greiner-Tollersrud, O.K.; Berg, T. Lysosomal Storage Disorders. In Madame Curie Bioscience Database [Internet]; Landes Bioscience: Austin, TX, USA, 2013. [Google Scholar]
- Desnick, R.J.; Schuchman, E.H. Enzyme replacement therapy for lysosomal diseases: Lessons from 20 years of experience and remaining challenges. Annu. Rev. Genomics Hum. Genet. 2012, 13, 307–335. [Google Scholar] [CrossRef] [PubMed]
- Kloosterman, W.M.J.; Roest, S.; Priatna, S.R.; Stavila, E.; Loos, K. Chemo-enzymatic synthesis route to poly(glucosyl-acrylates) using glucosidase from almonds. Green Chem. 2014, 16, 1837–1846. [Google Scholar] [CrossRef]
- Vellard, M. The enzyme as drug: Application of enzymes as pharmaceuticals. Curr. Opin. Biotechnol. 2003, 14, 444–450. [Google Scholar] [CrossRef]
- Olsson, M.L.; Clausen, H. Modifying the red cell surface: Towards an ABO-universal blood supply. Br. J. Haematol. 2008, 140, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Dozier, J.K.; Distefano, M.D. Site-Specific PEGylation of Therapeutic Proteins. Int. J. Mol. Sci. 2015, 16, 25831–25864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Zhu, Y.; Chen, J. Microbial transglutaminase production: Understanding the mechanism. Biotechnol. Genet. Eng. Rev. 2010, 26, 205–222. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lei, X.; Zhu, H.; Zhang, H.; Guan, C.; Chen, Z.; Zheng, W.; Fu, L.; Zheng, T. Chitinase producing bacteria with direct algicidal activity on marine diatoms. Sci. Rep. 2016, 6, 21984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollmann, F.; Arends, I.W.C.E.; Holtmann, D. Enzymatic reductions for the chemist. Green Chem. 2011, 13, 2285–2314. [Google Scholar] [CrossRef]
- Han, Y.; Yang, B.; Zhang, F.; Miao, X.; Li, Z. Characterization of antifungal chitinase from marine Streptomyces sp. DA11 associated with South China Sea sponge Craniella australiensis. Mar. Biotechnol. 2009, 11, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Hein, S.; Wang, K.; Stevens, W.F.; Kjems, J. Chitosan composites for biomedical applications: Status, challenges and perspectives. Mater. Sci. Technol. 2008, 24, 1053–1061. [Google Scholar] [CrossRef]
- Lyu, Y.; Laborda, P.; Huang, K.; Cai, Z.P.; Wang, M.; Aimin, L.; Doherty, C.; Liu, L.; Flitsch, S.; Voglmeir, J. Highly Efficient and Selective Biocatalytic Production of Glucosamine from Chitin. Green Chem. 2016, 19, 527–535. [Google Scholar]
- Shearer, T.L.; Snell, T.W.; Hay, M.E. Gene expression of corals in response to macroalgal competitors. PLoS ONE 2014, 9, e114525. [Google Scholar] [CrossRef] [PubMed]
- Cobbett, C.; Goldsbrough, P. Phytochelatins and metallothioneins: Roles in heavy metal detoxification and homeostasis. Annu. Rev. Plant Biol. 2002, 53, 159–182. [Google Scholar] [CrossRef] [PubMed]
- Bundy, J.G.; Kille, P. Metabolites and metals in Metazoa—What role do phytochelatins play in animals? Metallomics 2014, 6, 1576–1582. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Kang, S.H.; Lee, Y.-I.; Choa, Y.-H.; Mulchandani, A.; Myung, N.V.; Chen, W. Enzyme mediated synthesis of phytochelatin-capped CdS nanocrystals. Appl. Phys. Lett. 2010, 97, 123703. [Google Scholar] [CrossRef]
- Kang, S.H.; Singh, S.; Kim, J.Y.; Lee, W.; Mulchandani, A.; Chen, W. Bacteria metabolically engineered for enhanced phytochelatin production and cadmium accumulation. Appl. Environ. Microbiol. 2007, 73, 6317–6320. [Google Scholar] [CrossRef] [PubMed]
- Archelas, A.; Furstoss, R. Synthetic applications of epoxide hydrolases. Curr. Opin. Chem. Biol. 2001, 5, 112–119. [Google Scholar] [CrossRef]
- Van Loo, B.; Kingma, J.; Arand, M.; Wubbolts, M.G.; Janssen, D.B. Diversity and biocatalytic potential of epoxide hydrolases identified by genome analysis. Appl. Environ. Microbiol. 2006, 72, 2905–2917. [Google Scholar] [CrossRef] [PubMed]
- Yamada, H.; Shimizu, S.; Kobayashi, M. Hydratases involved in nitrile conversion: Screening, characterization and application. Chem. Rec. 2001, 1, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Yamada, H.; Kobayashi, M. Nitrile hydratase and its application to industrial production of acrylamide. Biosci. Biotechnol. Biochem. 1996, 60, 1391–1400. [Google Scholar] [CrossRef] [PubMed]
- Park, J.M.; Trevor Sewell, B.; Benedik, M.J. Cyanide bioremediation: The potential of engineered nitrilases. Appl. Microbiol. Biotechnol. 2017, 101, 3029–3042. [Google Scholar] [CrossRef] [PubMed]
- Eun, H.-M. 3-Nucleases. In Enzymology Primer for Recombinant DNA Technology; Academic Press: San Diego, CA, USA, 1996; pp. 145–232. [Google Scholar]
- Kim, I.; Hur, N.W.; Shin, H.D.; Park, B.L.; Cheong, H.S.; Bae, S.C. Associations of DNase IV polymorphisms with autoantibodies in patients with systemic lupus erythematosus. Rheumatology 2008, 47, 996–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholson, A.W. Ribonuclease III mechanisms of double-stranded RNA cleavage. Wiley Interdiscip. Rev. RNA 2014, 5, 31–48. [Google Scholar] [CrossRef] [PubMed]
- Eun, H.-M. 6-DNA Polymerases. In Enzymology Primer for Recombinant DNA Technology; Academic Press: San Diego, CA, USA, 1996; pp. 345–489. [Google Scholar]
- Eun, H.-M. 7-RNA Polymerases. In Enzymology Primer for Recombinant DNA Technology; Academic Press: San Diego, CA, USA, 1996; pp. 491–565. [Google Scholar]
- Bachman, J. Reverse-transcription PCR (RT-PCR). Methods Enzymol. 2013, 530, 67–74. [Google Scholar] [PubMed]
- Eun, H.-M. 2-Ligases. In Enzymology Primer for Recombinant DNA Technology; Academic Press: San Diego, CA, USA, 1996; pp. 109–144. [Google Scholar]
- Spreitzer, R.J.; Salvucci, M.E. Rubisco: Structure, regulatory interactions, and possibilities for a better enzyme. Annu. Rev. Plant Biol. 2002, 53, 449–475. [Google Scholar] [CrossRef] [PubMed]
- Moglia, A.; Lanteri, S.; Comino, C.; Hill, L.; Knevitt, D.; Cagliero, C.; Rubiolo, P.; Bornemann, S.; Martin, C. Dual catalytic activity of hydroxycinnamoyl-coenzyme A quinate transferase from tomato allows it to moonlight in the synthesis of both mono- and dicaffeoylquinic acids. Plant Physiol. 2014, 166, 1777–1787. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Z.; Zhang, Y.H.; Ren, H.; Wang, Y.L.; Jiang, W.; Fang, B.S. Strategies and perspectives of assembling multi-enzyme systems. Crit. Rev. Biotechnol. 2017, 37, 1024–1037. [Google Scholar] [CrossRef] [PubMed]
- Kupski, L.; Queiroz, M.I.; Badiale-Furlong, E. Application of carboxypeptidase A to a baking process to mitigate contamination of wheat flour by ochratoxin A. Process Biochem. 2018, 64, 248–254. [Google Scholar] [CrossRef]
- Reimer, J.D.; Ono, S.; Takishita, K.; Tsukahara, J.; Maruyama, T. Molecular evidence suggesting species in the zoanthid genera Palythoa and Protopalythoa (Anthozoa: Hexacorallia) are congeneric. Zool. Sci. 2006, 23, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Swain, T.D. Revisiting the phylogeny of Zoanthidea (Cnidaria: Anthozoa): Staggered alignment of hypervariable sequences improves species tree inference. Mol. Phyl. Evol. 2018, 118, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Quattrini, A.M.; Faircloth, B.C.; Duenas, L.F.; Bridge, T.C.L.; Brugler, M.R.; Calixto-Botia, I.F.; DeLeo, D.M.; Foret, S.; Herrera, S.; Lee, S.M.Y.; et al. Universal target-enrichment baits for anthozoan (Cnidaria) phylogenomics: New approaches to long-standing problems. Mol. Ecol. Resour. 2018, 18, 281–295. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protocols 2013, 8, 1494–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Gotz, S.; Garcia-Gomez, J.M.; Terol, J.; Talon, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Fang, L.; Zheng, H.; Zhang, Y.; Chen, J.; Zhang, Z.; Wang, J.; Li, S.; Li, R.; Bolund, L.; et al. WEGO: A web tool for plotting GO annotations. Nucleic Acids Res. 2006, 34, W293–W297. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 2016, 44, D457–D462. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lassmann, T.; Frings, O.; Sonnhammer, E.L. Kalign2: High-performance multiple alignment of protein and nucleotide sequences allowing external features. Nucleic Acids Res. 2009, 37, 858–865. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.; Binns, D.; Chang, H.Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [PubMed]
- Hill, D.P.; Davis, A.P.; Richardson, J.E.; Corradi, J.P.; Ringwald, M.; Eppig, J.T.; Blake, J.A. PROGRAM DESCRIPTION: Strategies for Biological Annotation of Mammalian Systems: Implementing Gene Ontologies in Mouse Genome Informatics. Genomics 2001, 74, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25. [Google Scholar] [CrossRef] [PubMed]
- The Gene Ontology Consortium. Expansion of the Gene Ontology knowledgebase and resources. Nucleic Acids Res. 2017, 45, D331–D338. [Google Scholar]
- Placzek, S.; Schomburg, I.; Chang, A.; Jeske, L.; Ulbrich, M.; Tillack, J.; Schomburg, D. BRENDA in 2017: New perspectives and new tools in BRENDA. Nucleic Acids Res. 2017, 45, D380–D388. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Unigene | ECs | Activities | Substrate | Product |
|---|---|---|---|---|
| CL12403.Contig1 | 1.5.1.20 | methylenetetrahydrofolate reductase (NAD(P)H) | 5-methyltetrahydrofolate | 5,10-methylenetetrahydrofolate |
| 2.1.2.1 | glycine hydroxymethyltransferase | 5,10-methylenetetrahydrofolate | tetrahydrofolate | |
| CL12403.Contig2 | 1.5.1.20 | methylenetetrahydrofolate reductase (NAD(P)H) | 5-methyltetrahydrofolate | 5,10-methylenetetrahydrofolate |
| 2 | transferase | ? | ? | |
| CL2444.Contig1 | 4.2.1.11 | phosphopyruvate hydratase | 2-phospho-d-glycerate | phosphoenolpyruvate |
| 5.3.1.1 | triose-phosphate isomerase | d-glyceraldehyde 3-phosphate | glycerone phosphate | |
| Unigene12818 | 1.3.99.1 | succinate dehydrogenase | succinate | fumarate |
| 1.8.4 | oxidoreductase, acting on a sulfur group of donors | ? | ? | |
| 5.3.4.1 | protein disulfide isomerase | -S-S- bonds rearrangement | none | |
| Unigene14615 | 2.4.2.30 | NAD ADP-ribosyltransferase | NAD+ | nicotinamide |
| 2.4.2.31 | NAD(P)+protein-arginine ADP-ribosyltransferase | NAD+ & protein-l-arginine | nicotinamide & protein-omega-N-(ADP-d-ribosyl)-l-arginine | |
| Unigene28009 | 2.5.1.21 | farnesyl-diphosphate farnesyltransferase | (2E,6E)-farnesyl diphosphate | squalene |
| 1.14 | oxidoreductase | ? | ? | |
| Unigene32504 | 2.1.1 | O-methyltransferase | ? | ? |
| 2.6.1.42 | branched-chain-amino-acid transaminase | L-leucine | 4-methyl-2-oxopentanoate | |
| Unigene33780 | 2.7.7.9 | UTP:glucose-1-phosphate uridylyltransferase | alpha-d-glucose 1-phosphate | UDP-glucose |
| 5.3.1.1 | triose-phosphate isomerase | d-glyceraldehyde 3-phosphate | glycerone phosphate | |
| Unigene34807 | 3.4.21 | serine-type endopeptidase | ? | ? |
| 3.4.24 | metalloendopeptidase | ? | ? | |
| Unigene38918 | 2.7.2.3 | phosphoglycerate kinase | 3-phospho-d-glycerate | 3-phospho-d-glyceroyl phosphate |
| 4.2.1.11 | phosphopyruvate hydratase | 2-phospho-d-glycerate | phosphoenolpyruvate | |
| Unigene52468 | 2.1.1 | methyltransferase | ? | ? |
| 2.5.1 | transferase, transferring alkyl or aryl groups | ? | ? | |
| Unigene9562 | 2.1.2.11 | 3-methyl-2-oxobutanoate hydroxymethyltransferase | 5,10-methylenetetrahydrofolate & 3-methyl-2-oxobutanoate | tetrahydrofolate & 2-dehydropantoate |
| 6.3.2.1 | pantoate-beta-alanine ligase | (R)-pantoate | (R)-pantothenate | |
| Unigene9804 | 2.5.1.21 | farnesyl-diphosphate farnesyltransferase | (2E,6E)-farnesyl diphosphate | squalene |
| 5.3.3.2 | isopentenyl-diphosphate delta-isomerase | isopentenyl diphosphate | dimethylallyl diphosphate |
| Class | Sub-Subclass EC Number | Sub-Subclass Principal Enzyme Type(s) | Sub-Subclass Known Enzyme Entries | P. variabilis Number Sequences |
|---|---|---|---|---|
| Oxidoreductases | 1.13.11 | dioxygenase | 80 | 26 |
| 1.14.11 | dioxygenase, hydroxylase, demethylase | 56 | 9 | |
| 1.14.12 | dioxygenase | 23 | 1 | |
| 1.14.13 | monooxygenase, hydroxylase | 235 | 1 | |
| 1.14.16 | monooxygenase | 7 | 3 | |
| 1.14.17 | monooxygenase | 3 | 13 | |
| 1.14.19 | desaturase | 51 | 5 | |
| Transferases | 2.7.10 | protein-tyrosine kinase | 2 | 72 |
| 2.7.11 | protein-serine/threonine kinase | 33 | 100 | |
| Hydrolases | 3.1.13 | exoribonuclease | 5 | 3 |
| 3.1.21 | endodeoxyribonuclease | 9 | 1 | |
| 3.4.11 | aminopeptidase | 23 | 25 | |
| 3.4.13 | dipeptidase | 12 | 7 | |
| 3.4.15 | peptidyl-dipeptidase | 4 | 1 | |
| 3.4.16 | serine-type carboxypeptidase | 4 | 10 | |
| 3.4.17 | metallocarboxypeptidase | 20 | 25 | |
| 3.4.19 | omega peptidase | 12 | 9 | |
| 3.4.21 | serine endopeptidase | 100 | 136 | |
| 3.4.22 | cysteine endopeptidase | 58 | 20 | |
| 3.4.23 | aspartic endopeptidase | 41 | 70 | |
| 3.4.24 | metalloendopeptidase | 83 | 120 | |
| 3.4.25 | threonine endopeptidase | 2 | 34 | |
| Lyases | 4.1.99 | carbon-carbon lyases | 16 | 3 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
R. L. Morlighem, J.-É.; Huang, C.; Liao, Q.; Braga Gomes, P.; Daniel Pérez, C.; De Brandão Prieto-da-Silva, Á.R.; Ming-Yuen Lee, S.; Rádis-Baptista, G. The Holo-Transcriptome of the Zoantharian Protopalythoa variabilis (Cnidaria: Anthozoa): A Plentiful Source of Enzymes for Potential Application in Green Chemistry, Industrial and Pharmaceutical Biotechnology. Mar. Drugs 2018, 16, 207. https://doi.org/10.3390/md16060207
R. L. Morlighem J-É, Huang C, Liao Q, Braga Gomes P, Daniel Pérez C, De Brandão Prieto-da-Silva ÁR, Ming-Yuen Lee S, Rádis-Baptista G. The Holo-Transcriptome of the Zoantharian Protopalythoa variabilis (Cnidaria: Anthozoa): A Plentiful Source of Enzymes for Potential Application in Green Chemistry, Industrial and Pharmaceutical Biotechnology. Marine Drugs. 2018; 16(6):207. https://doi.org/10.3390/md16060207
Chicago/Turabian StyleR. L. Morlighem, Jean-Étienne, Chen Huang, Qiwen Liao, Paula Braga Gomes, Carlos Daniel Pérez, Álvaro Rossan De Brandão Prieto-da-Silva, Simon Ming-Yuen Lee, and Gandhi Rádis-Baptista. 2018. "The Holo-Transcriptome of the Zoantharian Protopalythoa variabilis (Cnidaria: Anthozoa): A Plentiful Source of Enzymes for Potential Application in Green Chemistry, Industrial and Pharmaceutical Biotechnology" Marine Drugs 16, no. 6: 207. https://doi.org/10.3390/md16060207
APA StyleR. L. Morlighem, J. -É., Huang, C., Liao, Q., Braga Gomes, P., Daniel Pérez, C., De Brandão Prieto-da-Silva, Á. R., Ming-Yuen Lee, S., & Rádis-Baptista, G. (2018). The Holo-Transcriptome of the Zoantharian Protopalythoa variabilis (Cnidaria: Anthozoa): A Plentiful Source of Enzymes for Potential Application in Green Chemistry, Industrial and Pharmaceutical Biotechnology. Marine Drugs, 16(6), 207. https://doi.org/10.3390/md16060207