Marine Microalgae, Spirulina maxima-Derived Modified Pectin and Modified Pectin Nanoparticles Modulate the Gut Microbiota and Trigger Immune Responses in Mice

 , ,
, , 
Abstract
:1. Introduction
2. Results
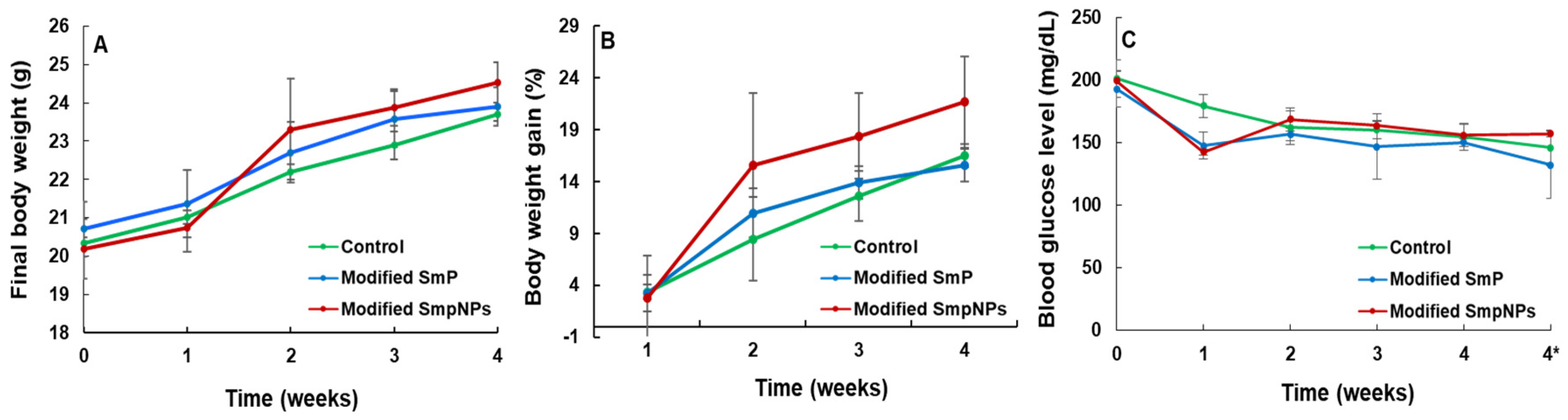
2.1. Growth Performance and Blood Glucose Levels of Mice upon Modified SmP and SmPNPs Supplementations
2.2. Effects of Modified SmP and SmPNPs Supplementations on the Mouse Gut Microbiota
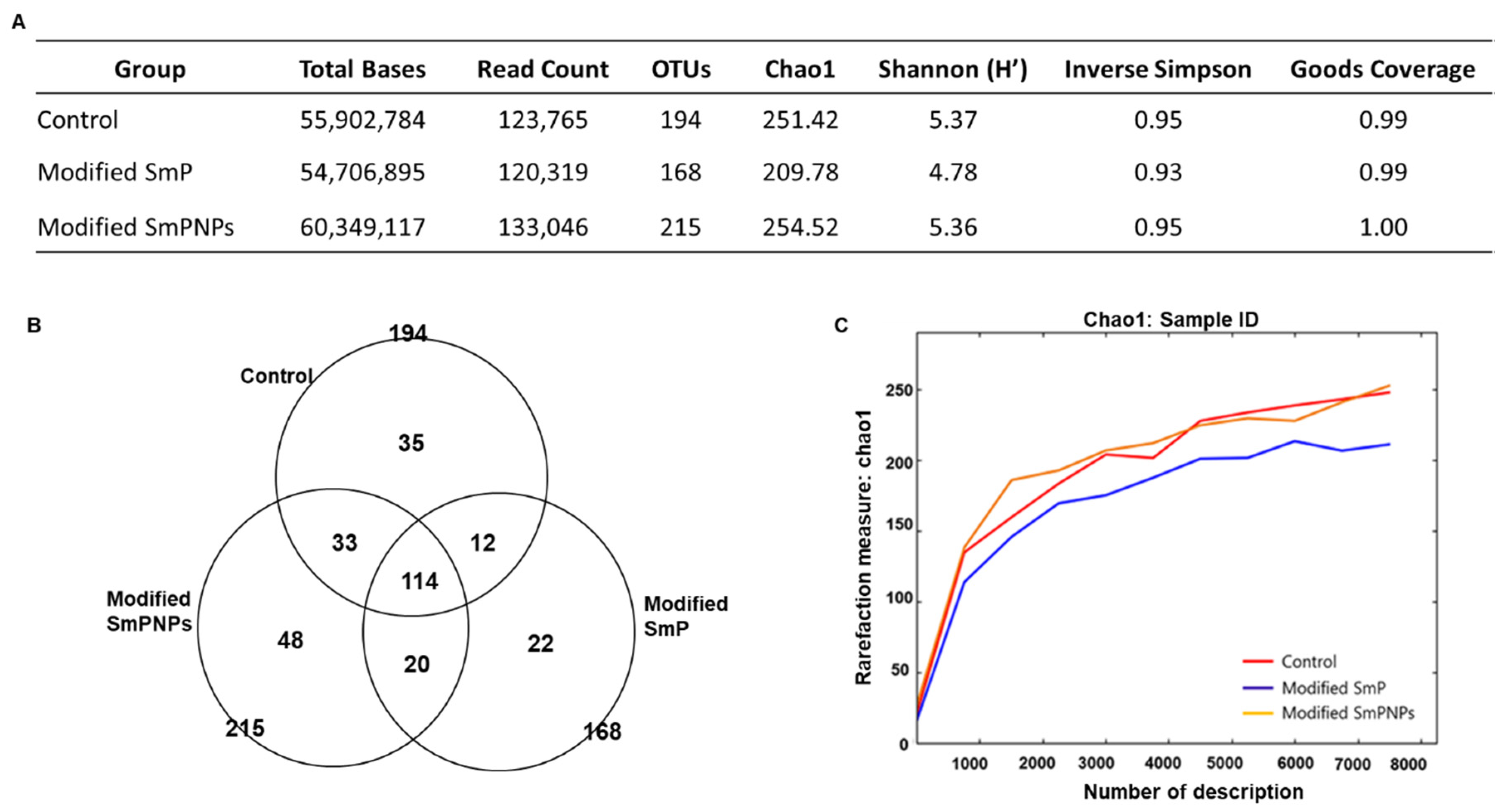
2.2.1. Metagenomic Sequencing and Diversity Analysis
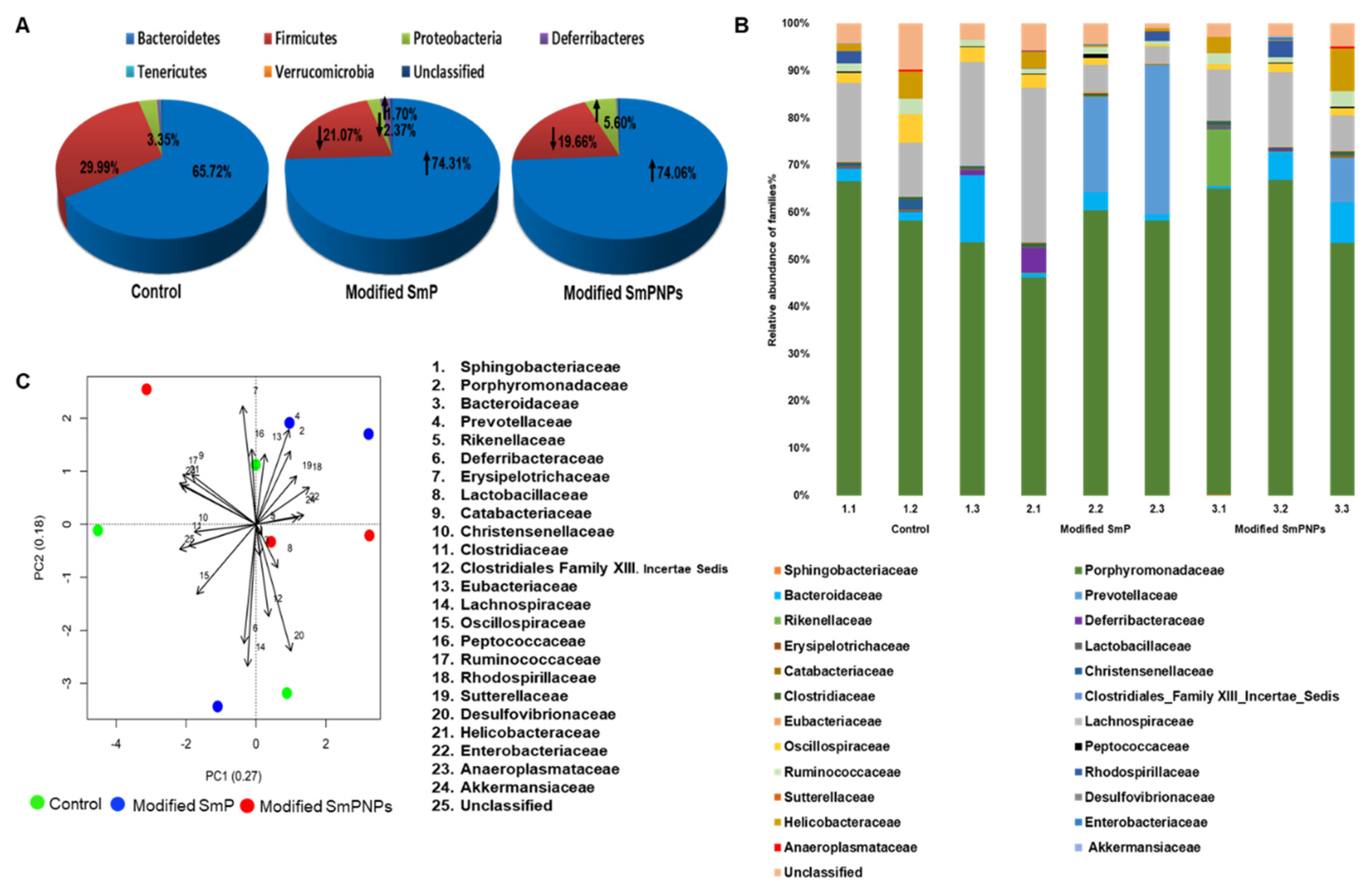
2.2.2. Taxonomic Analysis
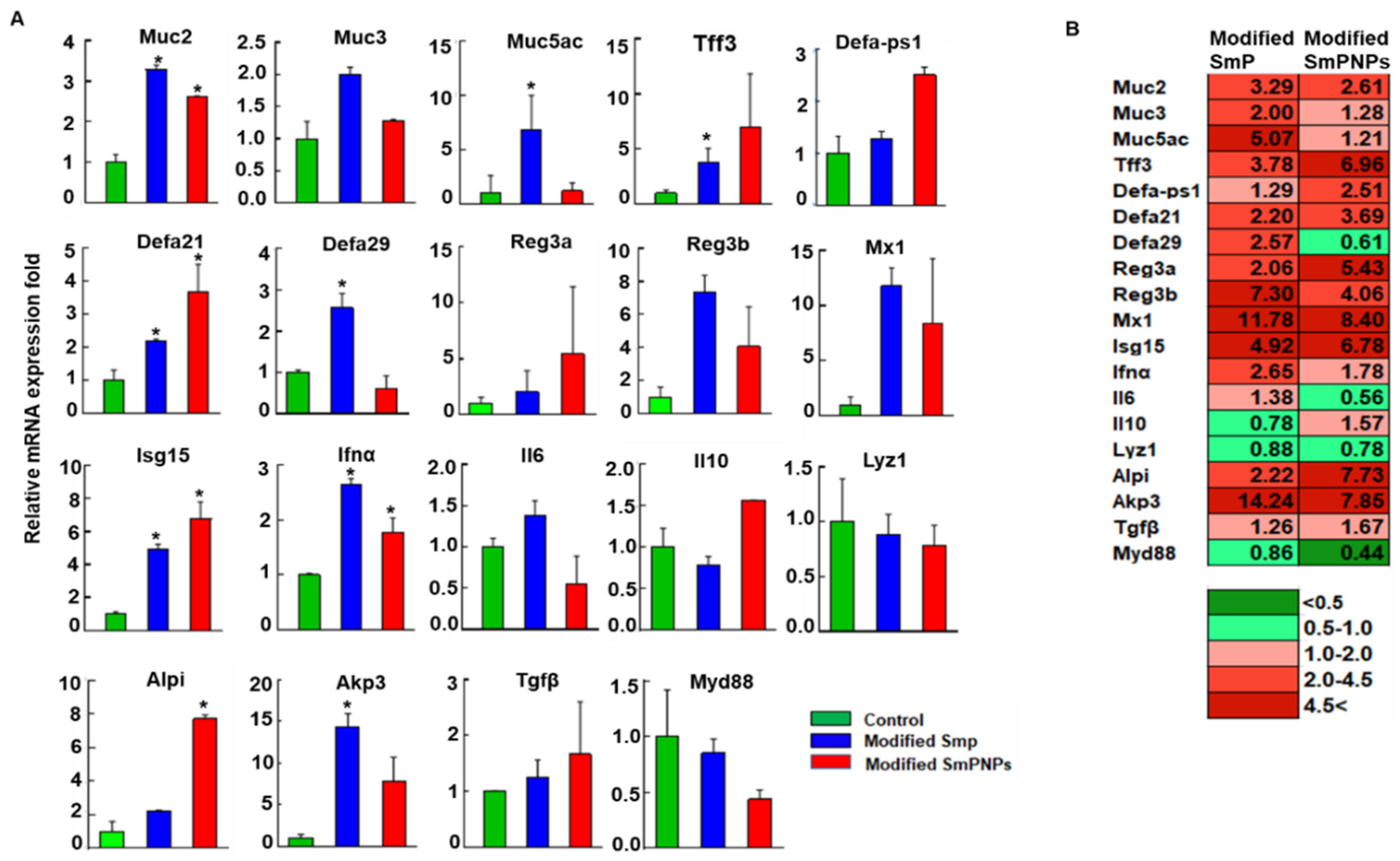
2.3. Modified SmP and SmPNPs Supplementations Displayed Immunostimulation in Mice
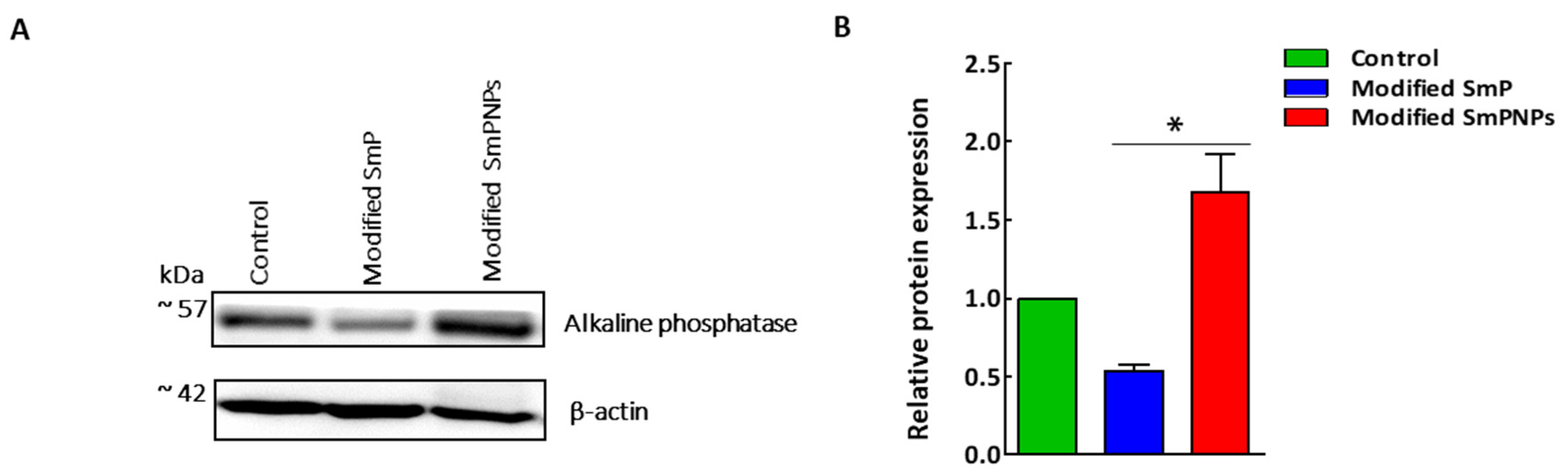
2.4. Intestine Alkaline Phosphatase (IAP) Expression by Immunoblotting Analysis
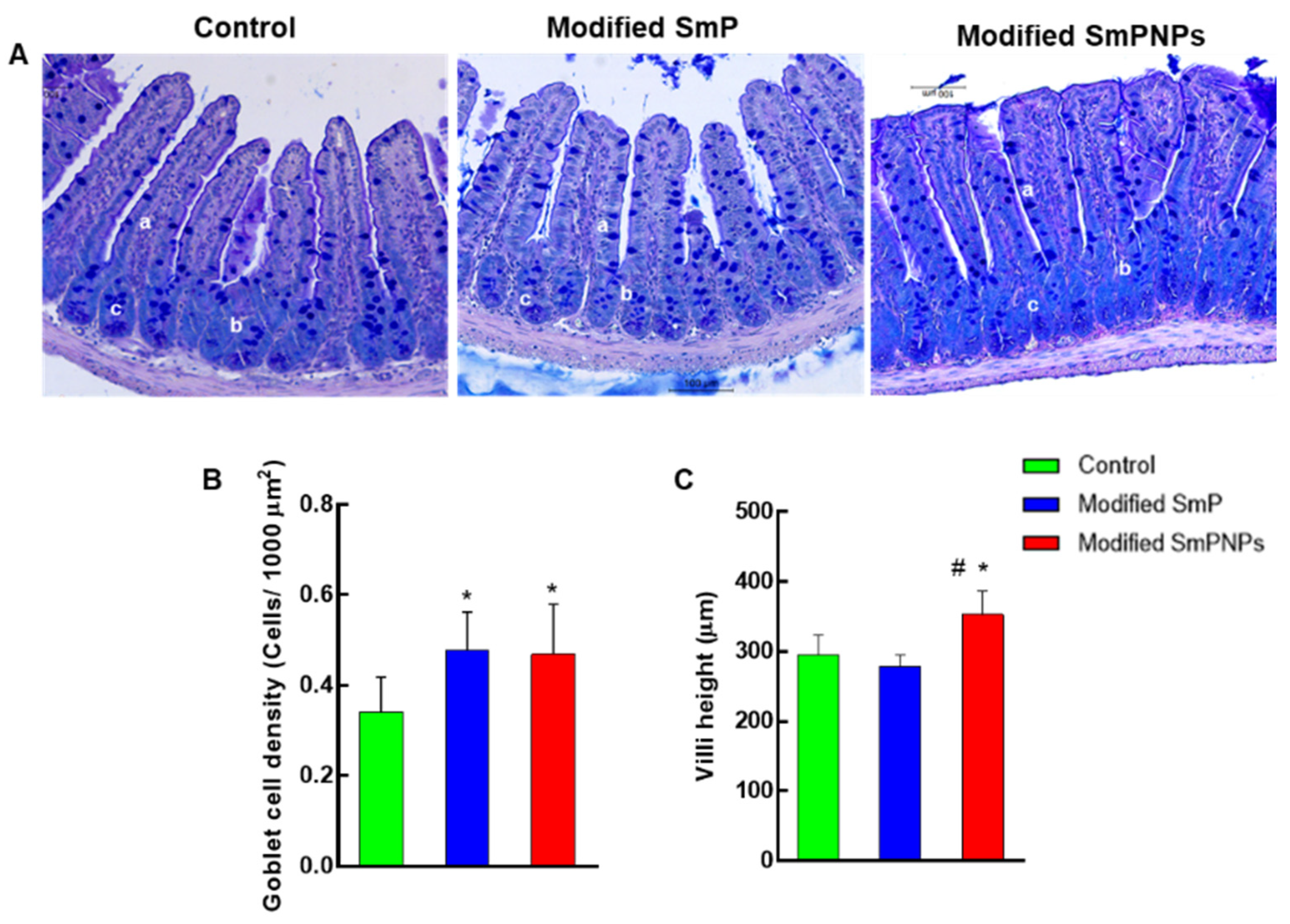
2.5. Histological Analysis
3. Discussion
4. Materials and Methods
4.1. Preparation and Characterization of Modified SmP and Modified SmPNPs
4.2. Animals and Experimental Design
4.3. Sample Collection, Genomic DNA Extraction, and Library Construction
4.4. Next Generation Sequencing (NGS) and Metagenomics Analysis
4.5. RNA Extraction and qRT-PCR Analysis
4.6. Immunoblot Analysis for IAP Expression
4.7. Histological Analysis
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Veronese, N.; Solmi, M.; Caruso, M.G.; Giannelli, G.; Osella, A.R.; Evangelou, E.; Maggi, S.; Fontana, L.; Stubbs, B.; Tzoulaki, I. Dietary fiber and health outcomes: An umbrella review of systematic reviews and meta-analyses. Am. J. Clin. Nutr. 2018, 107, 436–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Field, C.J.; McBurney, M.I.; Massimino, S.; Hayek, M.G.; Sunvold, G.D. The fermentable fiber content of the diet alters the function and composition of canine gut associated lymphoid tissue. Vet. Immunol. Immunopathol. 1999, 72, 325–341. [Google Scholar] [CrossRef]
- Malys, M.K.; Campbell, L.; Malys, N. Symbiotic and antibiotic interactions between gut commensal microbiota and host immune system. Medicina 2015, 51, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Kho, Z.Y.; Lal, S.K. The human gut microbiome: A potential controller of wellness and disease. Front. Microbiol. 2018, 9, 1835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noreen, A.; Akram, J.; Rasul, I.; Mansha, A.; Yaqoob, N.; Iqbal, R.; Tabasum, S.; Zuber, M.; Zia, K.M. Pectins functionalized biomaterials; a new viable approach for biomedical applications: A review. Int. J. Biol. Macromol. 2017, 101, 254–272. [Google Scholar] [CrossRef]
- Srivastava, P.; Malviya, R. Sources of pectin, extraction and its applications in pharmaceutical industry-An overview. Indian J. Nat. Prod. Resour. 2011, 2, 10–18. [Google Scholar]
- Habib, M.A.B.; Parvin, M.; Huntington, T.C.; Hasan, M.R. A review on culture, production and use of spirulina as food for humans and feeds for domestic animals and fish. In FAO Fisheries and Aquaculture Circular, No. 1034; FAO: Rome, Italy, 2008; 33p. [Google Scholar]
- Heo, M.G.; Choung, S.Y. Anti-obesity effects of Spirulina maxima in high fat diet induced obese rats via the activation of AMPK pathway and SIRT1. Food Funct. 2018, 9, 4906–4915. [Google Scholar] [CrossRef]
- Løbner, M.; Walsted, A.; Larsen, R.; Bendtzen, K.; Nielsen, C.H. Enhancement of human adaptive immune responses by administration of a high-molecular-weight polysaccharide extract from the Cyanobacterium Arthrospira platensis. J. Med. Food 2008, 11, 313–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.W.; Yang, T.S.; Chen, M.J.; Chang, Y.C.; Eugene, I.; Wang, C.; Ho, C.L.; Lai, Y.J.; Yu, C.C.; Chou, J.C.; et al. Purification and immunomodulating activity of C-phycocyanin from Spirulina platensis cultured using power plant flue gas. Process Biochem. 2014, 49, 1337–1344. [Google Scholar] [CrossRef] [Green Version]
- Edirisinghe, S.L.; Dananjaya, S.H.S.; Nikapitiya, C.; Liyanage, T.D.; Lee, K.A.; Oh, C.; Kang, D.H.; De Zoysa, M. Novel pectin isolated from Spirulina maxima enhances the disease resistance and immune responses in zebrafish against Edwardsiella piscicida and Aeromonas hydrophila. Fish Shellfish Immunol. 2019, 94, 558–565. [Google Scholar] [CrossRef]
- O’Rourke, C.; Gregson, T.; Murray, L.; Sadler, I.H.; Fry, S.C. Sugar composition of the pectic polysaccharides of charophytes, the closest algal relatives of land-plants: Presence of 3-O-methyl-d-galactose residues. Ann. Bot. 2015, 116, 225–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoiczyk, E.; Hansel, A. Cyanobacterial Cell Walls: News from an unusual prokaryotic envelope. J. Bacteriol. 2000, 182, 1191–1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shpigelman, A.; Kyomugasho, C.; Christiaens, S.; Loey, A.M.V.; Hendrickx, M.E. The effect of high pressure homogenization on pectin: Importance of pectin source and pH. Food Hydrocoll. 2015, 43, 189–198. [Google Scholar] [CrossRef]
- De Roeck, A.; Duvetter, T.; Fraeye, I.; Van der Plancken, I.; Sila, D.N.; Van Loey, A.; Hendrickx, M. Effect of high-pressure/high temperature processing on chemical pectin conversions in relation to fruit and vegetable texture. Food Chem. 2009, 115, 207–213. [Google Scholar] [CrossRef]
- Gurry, T.; Gibbons, S.M.; Kearney, S.M.; Ananthakrishnan, A.; Jiang, X.; Duvallet, C.; Kassam, Z.; Alm, E.J. Predictability and persistence of prebiotic dietary supplementation in a healthy human cohort. Sci. Rep. 2018, 8, 12699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Xu, C.; Huang, R.; Song, J.; Li, D.; Xia, M. Butyrate from pectin fermentation inhibits intestinal cholesterol absorption and attenuates atherosclerosis in apolipoprotein E-deficient mice. J. Nutr. Biochem. 2018, 56, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Adam, C.L.; Thomson, L.M.; Williams, P.A.; Ross, A.W. Soluble fermentable dietary fibre (Pectin) decreases caloric intake, adiposity and lipidaemia in high-fat diet-induced obese rats. PLoS ONE 2015, 10, e0140392. [Google Scholar] [CrossRef] [Green Version]
- Nelson, R.W.; Ihle, S.L.; Lewis, L.D.; Salisbury, S.K.; Miller, T.; Bergdall, V.; Bottoms, G.D. Effects of dietary fiber supplementation on glycemic control in dogs with alloxan-induced diabetes mellitus. Am. J. Vet. Res. 1991, 52, 2060–2066. [Google Scholar]
- Fraher, M.H.; O’Toole, P.W.; Quigley, E.M.M. Techniques used to characterize the gut microbiota: A guide for the clinician. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 312–322. [Google Scholar] [CrossRef]
- Jiang, T.; Gao, X.; Wu, C.; Tian, F.; Lei, Q.; Bi, J.; Xie, B.; Wang, H.Y.; Chen, S.; Wang, X. Apple-derived pectin modulates gut microbiota, improves gut barrier function, and attenuates metabolic endotoxemia in rats with diet-induced obesity. Nutrients 2016, 8, 126. [Google Scholar] [CrossRef] [Green Version]
- Chung, W.S.F.; Walker, A.W.; Louis, P.; Parkhill, J.; Vermeiren, J.; Bosscher, D.; Duncan, S.H.; Flint, H.J. Modulation of the human gut microbiota by dietary fibres occurs at the species level. BMC Biol. 2016, 14, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P.; et al. A core gut microbiome in obese and lean twins. Nature 2009, 457, 480–484. [Google Scholar] [CrossRef] [Green Version]
- Shtriker, M.G.; Hahn, M.; Taieb, E.; Nyska, A.; Moallem, U.; Tirosh, O.; Madar, Z. Fenugreek galactomannan and citrus pectin improve several parameters associated with glucose metabolism and modulate gut microbiota in mice. Nutrition 2018, 46, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Kelly, W.J.; Leahy, S.C.; Kamke, J.; Soni, P.; Koike, S.; Mackie, R.; Seshadri, R.; Cook, G.M.; Morales, S.E.; Greening, C.; et al. Occurrence and expression of genes encoding methyl-compound production in rumen bacteria. Anim. Microbiome 2019, 1, 15. [Google Scholar] [CrossRef]
- Kononen, E.; Asikainen, S.; Saarela, M.; Karjalainen, J.; Jousimies-Somer, H. The oral gram-negative anaerobic microflora in young children: Longitudinal changes from edentulous to dentate mouth. Oral Microbiol. Immunol. 1994, 9, 136–141. [Google Scholar] [CrossRef]
- Yang, Y.; Millán, J.L.; Mecsas, J.; Guillemin, K. Intestinal alkaline phosphatase deficiency leads to lipopolysaccharide desensitization and faster weight gain. Infect. Immun. 2015, 83, 247–258. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, N.; Emoto, T.; Yamashita, T.; Watanabe, H.; Hayashi, T.; Tabata, T.; Hoshi, N.; Hatano, N.; Ozawa, G.; Sasaki, N.; et al. Bacteroides vulgatus and Bacteroides dorei reduce gut microbial lipopolysaccharide production and inhibit Atherosclerosis. Circulation 2018, 138, 2486–2498. [Google Scholar] [CrossRef]
- Karunasena, E.; Kurkure, P.C.; Lackey, R.D.; McMahon, K.W.; Kiernan, E.P.; Graham, S.; Alabady, M.S.; Campos, D.L.; Tatum, O.L.; Brashears, M.M. Effects of the probiotic Lactobacillus animalis in murine Mycobacterium avium subspecies paratuberculosis infection. BMC Microbiol. 2013, 13, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Desai, M.S.; Seekatz, A.M.; Koropatkin, N.M.; Kamada, N.; Hickey, C.A.; Wolter, M.; Pudlo, N.A.; Kitamoto, S.; Terrapon, N.; Muller, A.; et al. A dietary fiber-deprived gut microbiota degrades the colonic mucus barrier and enhances pathogen susceptibility. Cell 2016, 167, 1339–1353. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.G.; Ge, Z.; Whary, M.T.; Erdman, S.E.; Horwitz, B.H. Helicobacter hepaticus infection in mice: Models for understanding lower bowel inflammation and cancer. Mucosal Immunol. 2011, 4, 22–30. [Google Scholar] [CrossRef] [Green Version]
- Mehmood, M.; Jaffar, N.A.; Nazim, M.; Khasawneh, F.A. Bacteremic skin and soft tissue infection caused by Prevotella loescheii. BMC Infect. Dis. 2014, 14, 2–5. [Google Scholar] [CrossRef] [Green Version]
- Popov, S.V.; Ovodov, Y.S. Polypotency of the immunomodulatory effect of pectins. Biochemistry 2013, 78, 823–835. [Google Scholar] [CrossRef] [PubMed]
- Hino, S.; Sonoyama, K.; Bito, H.; Kawagishi, H.; Aoe, S.; Morita, T. Low-methoxyl pectin stimulates small intestinal mucin secretion irrespective of goblet cell proliferation and is characterized by jejunum Muc2 upregulation in rats. J. Nutr. 2013, 143, 34–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aihara, E.; Engevik, K.A.; Montrose, M.H. Trefoil factor peptides and gastrointestinal function. Annu. Rev. Physiol. 2016, 79, 357–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, K.; Sakuragi, N.; Takakuwa, A.; Ayabe, T. Paneth cell α-defensins and enteric microbiota in health and disease. Biosci. Microbiota Food Health 2016, 35, 57–67. [Google Scholar] [CrossRef] [Green Version]
- Darnaud, M.; Dos Santos, A.; Gonzalez, P.; Augui, S.; Lacoste, C.; Desterke, C.; De Hertogh, G.; Valentino, E.; Braun, E.; Zheng, J.; et al. Enteric delivery of regenerating family member 3 alpha alters the intestinal microbiota and controls inflammation in mice with colitis. Gastroenterology 2018, 154, 1009–1023. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Luo, J.; Yu, J.; Mao, X.; Luo, Y.; Zheng, P.; He, J.; Yu, B.; Chen, D. Manipulation of intestinal antiviral innate immunity and immune evasion strategies of porcine epidemic diarrhea virus. BioMed. Res. Int. 2019, 3, 1862531. [Google Scholar] [CrossRef]
- Mizumori, M.; Ham, M.; Guth, P.H.; Engel, E.; Kaunitz, J.D.; Akiba, Y. Intestinal alkaline phosphatase regulates protective surface microclimate pH in rat duodenum. J. Physiol. 2009, 587, 3651–3663. [Google Scholar] [CrossRef]
- Bates, J.M.; Akerlund, J.; Mittge, E.; Guillemin, K. Intestinal alkaline phosphatase detoxifies lipopolysaccharide and prevents inflammation in zebrafish in response to the gut microbiota. Cell. Host Microbe. 2007, 2, 371–382. [Google Scholar] [CrossRef] [Green Version]
- Rahmatnejad, E.; Saki, A.A. Effect of dietary fibres on small intestine histomorphology and lipid metabolism in young broiler chickens. J. Anim. Physiol. Anim. Nutr. 2016, 100, 665–672. [Google Scholar] [CrossRef]
- Togashi, Y.; Shirakawa, J.; Okuyama, T.; Yamazaki, S.; Kyohara, M.; Miyazawa, A.; Suzuki, T.; Hamada, M.; Terauchi, Y. Evaluation of the appropriateness of using glucometers for measuring the blood glucose levels in mice. Sci. Rep. 2016, 6, 25465. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C (T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Escaffre, A.M.; Kaushik, S.; Mambrini, M. Morphometric evaluation of changes in the digestive tract of rainbow trout (Oncorhynchus mykiss) due to fish meal replacement with soy protein concentrate. Aquaculture 2007, 273, 127–138. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Control | Modified SmP | Modified SmPNPs |
|---|---|---|---|
| Initial BW (g) | 20.33 (0.15) | 20.70 (0.72) | 20.18 (0.79) |
| Final BW (g) | 23.69a (0.17) | 23.91a (0.51) | 24.53a (0.53) |
| Water intake (mL/mouse/day) | 4.05 (0.07) | 4.33* (0.07) | 4.34* (0.07) |
| Total pectin intake (g/4 weeks) | 0 | 45.36 | 45.64 |
| Weight gain (g/4 weeks) | 3.36 (0.12) | 3.21 (0.12) | 4.35 (0.44) |
| Weight gain (%/4 weeks) | 16.52 (0.7) | 15.54 (0.9) | 21.65 (2.5) |
| SGR% (% BW gain/day) | 0.55 (0.02) | 0.51 (0.03) | 0.70 (0.07) |
| Blood glucose level (mg/dL) | 171.25 (19.00) | 158.72 (20.00) | 165.82 (21.00) |
| Fasting blood glucose level (mg/mL) at 4th week | 145.75 (14.00) | 131.83 (26.00) | 156.91 (2.00) |
| Species of bacteria | Modified SmP vs. control | Modified SmPNPs vs. control | Possible effects | Reference |
|---|---|---|---|---|
| Bacteroides acidifaciens | Decrease | Increase | Prevents obesity. Improves insulin sensitivity. | [27] |
| Bacteroides vulgatus | Decrease | Increase | Reduce gut microbial lipopolysaccharide production. Inhibit Atherosclerosis. | [28] |
| Lactobacillus animalis | Decrease | Increase | Act as probiotic. Reduce chronic inflammation caused by Mycobacterium avium. | [29] |
| Akkermansia muciniphila | Decrease | Increase | Adheres to enterocytes. Strengthens the integrity of the epithelial cell layer. | [30] |
| Helicobacter hepaticus | Decrease | Decrease | Cause H. hepaticus-induced colitis. Triggers intestinal inflammation. | [31] |
| Prevotella loescheii | NA | Increase | Cause sepsis and soft tissue infections. | [32] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chandrarathna, H.P.S.U.; Liyanage, T.D.; Edirisinghe, S.L.; Dananjaya, S.H.S.; Thulshan, E.H.T.; Nikapitiya, C.; Oh, C.; Kang, D.-H.; De Zoysa, M. Marine Microalgae, Spirulina maxima-Derived Modified Pectin and Modified Pectin Nanoparticles Modulate the Gut Microbiota and Trigger Immune Responses in Mice. Mar. Drugs 2020, 18, 175. https://doi.org/10.3390/md18030175
Chandrarathna HPSU, Liyanage TD, Edirisinghe SL, Dananjaya SHS, Thulshan EHT, Nikapitiya C, Oh C, Kang D-H, De Zoysa M. Marine Microalgae, Spirulina maxima-Derived Modified Pectin and Modified Pectin Nanoparticles Modulate the Gut Microbiota and Trigger Immune Responses in Mice. Marine Drugs. 2020; 18(3):175. https://doi.org/10.3390/md18030175
Chicago/Turabian StyleChandrarathna, H.P.S.U., T.D. Liyanage, S.L. Edirisinghe, S.H.S. Dananjaya, E.H.T. Thulshan, Chamilani Nikapitiya, Chulhong Oh, Do-Hyung Kang, and Mahanama De Zoysa. 2020. "Marine Microalgae, Spirulina maxima-Derived Modified Pectin and Modified Pectin Nanoparticles Modulate the Gut Microbiota and Trigger Immune Responses in Mice" Marine Drugs 18, no. 3: 175. https://doi.org/10.3390/md18030175
APA StyleChandrarathna, H. P. S. U., Liyanage, T. D., Edirisinghe, S. L., Dananjaya, S. H. S., Thulshan, E. H. T., Nikapitiya, C., Oh, C., Kang, D. -H., & De Zoysa, M. (2020). Marine Microalgae, Spirulina maxima-Derived Modified Pectin and Modified Pectin Nanoparticles Modulate the Gut Microbiota and Trigger Immune Responses in Mice. Marine Drugs, 18(3), 175. https://doi.org/10.3390/md18030175