
Influence of Polyunsaturated Fatty Acid Intake on Kidney Functions of Rats with Chronic Renal Failure

 ,
,  and
and
Abstract
:1. Introduction
2. Results
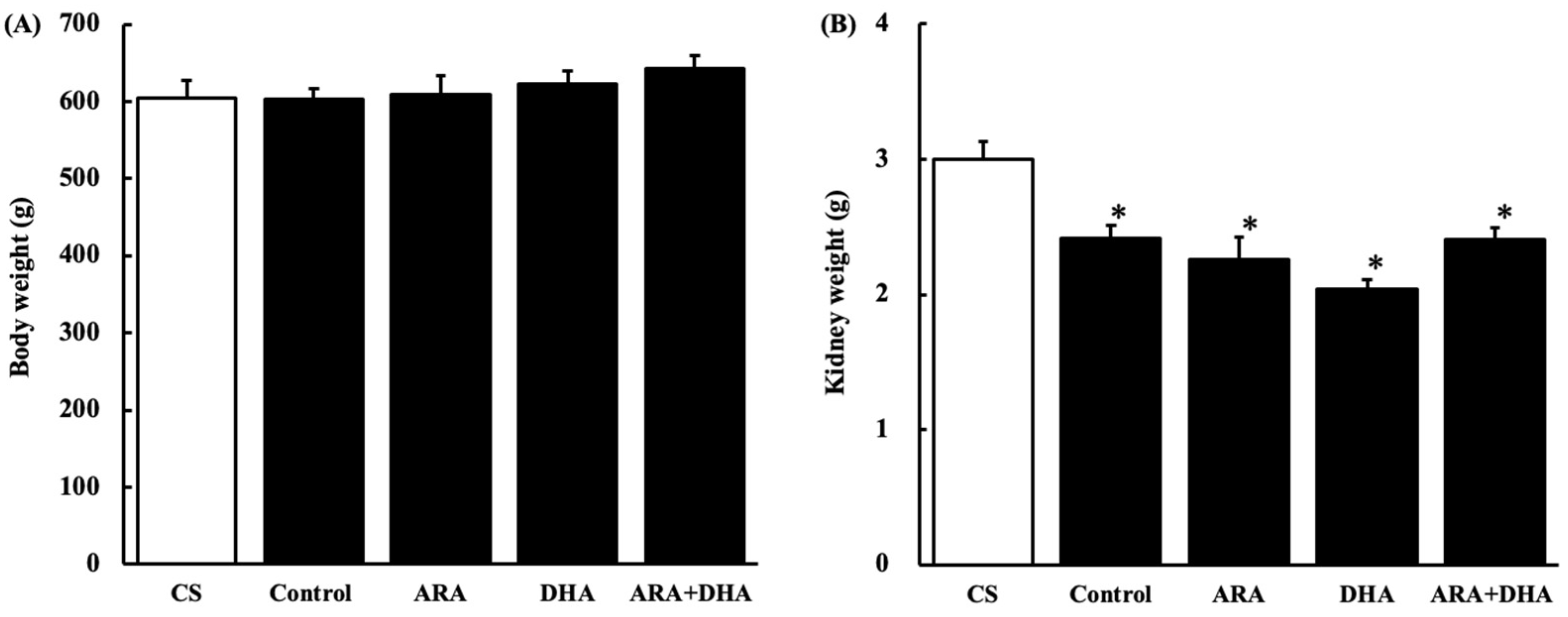
2.1. Body and Kidney Weight
2.2. Renal Parameter
2.3. Fatty Acid Composition of Total Lipids in the Plasma and Kidneys
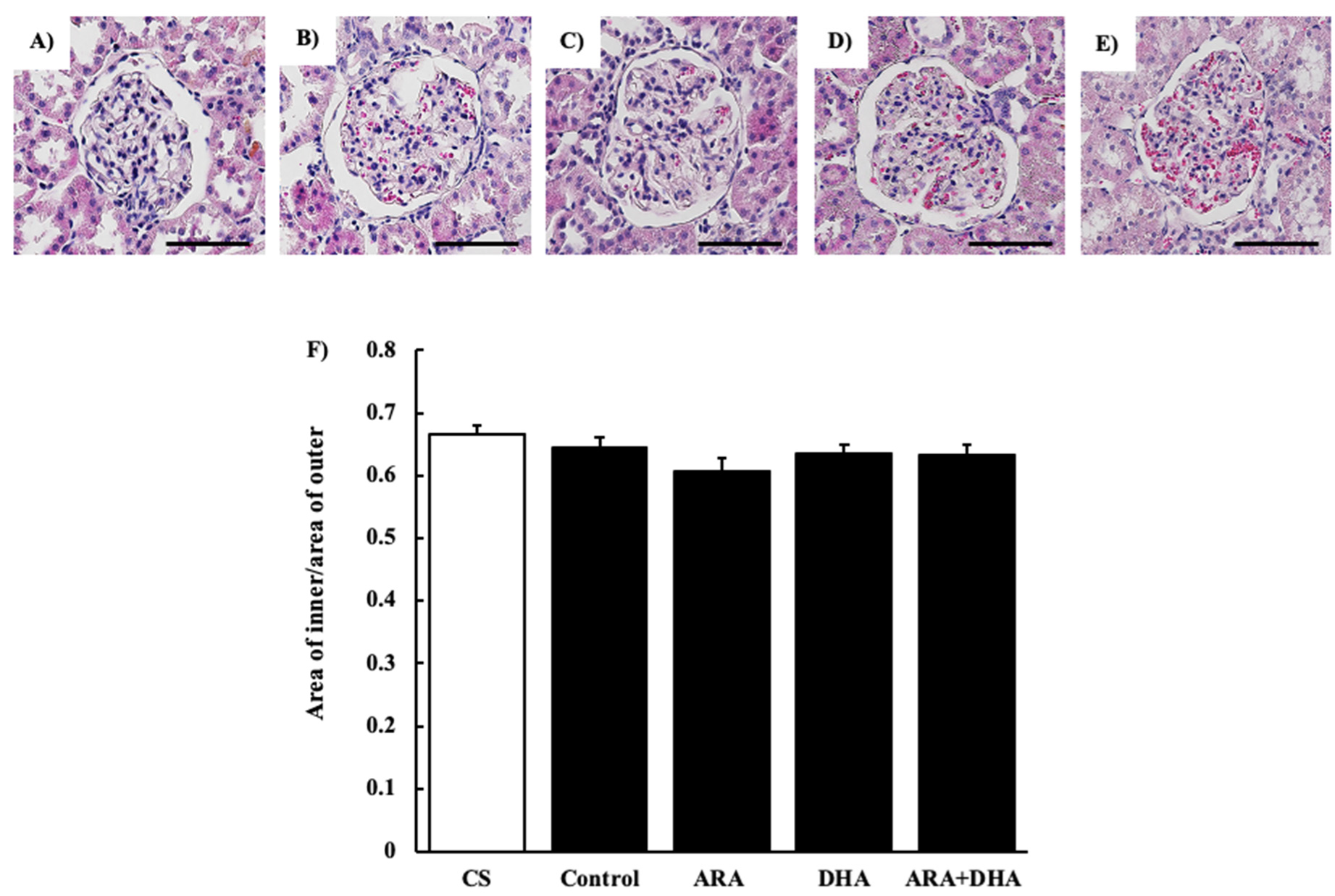
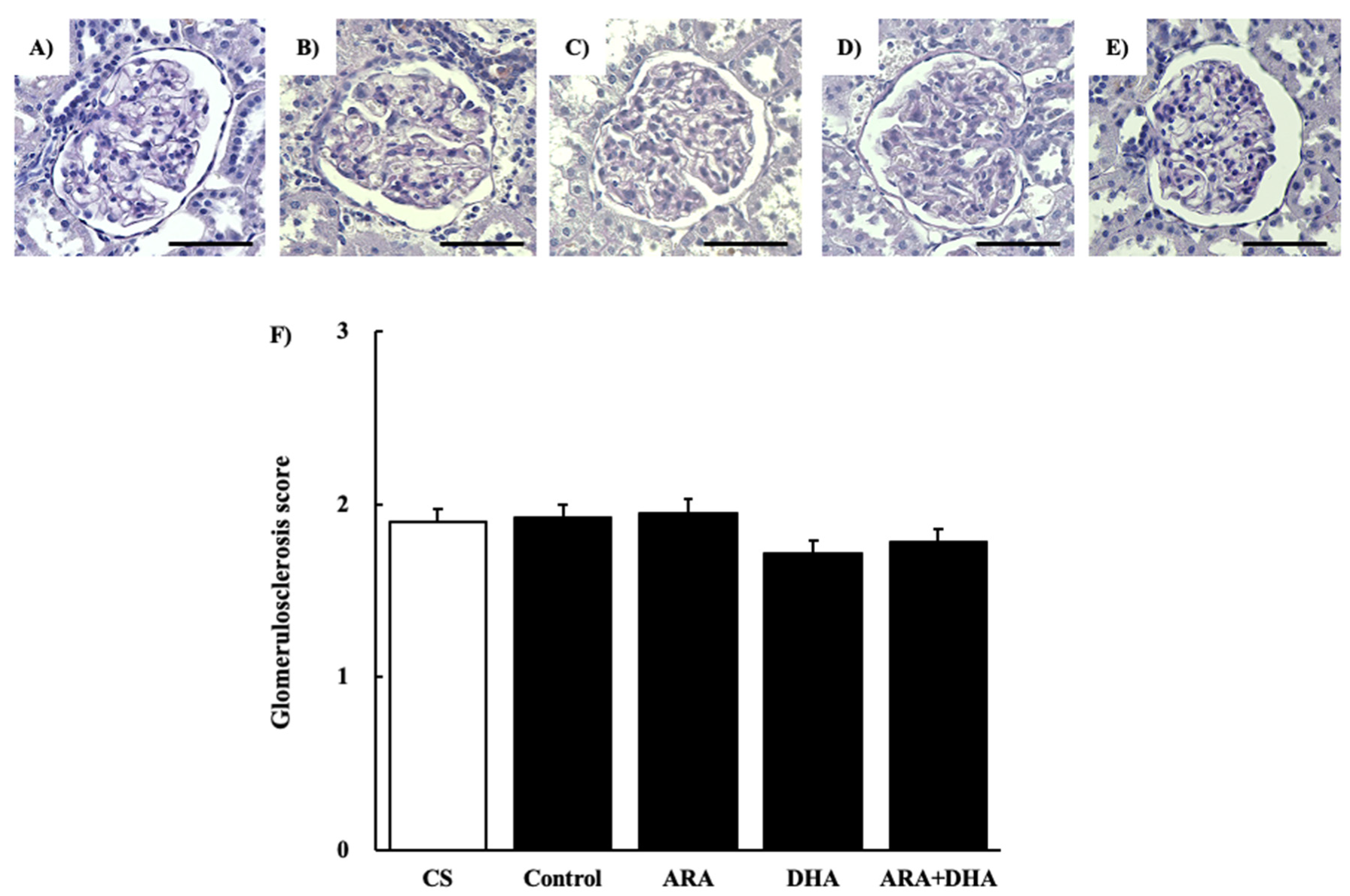
2.4. Histological Evaluation
2.4.1. Hematoxylin and Eosin (H&E) Staining
2.4.2. Periodic Acid Schiff (PAS) Staining
2.4.3. Periodic Acid Methenamine-Silver (PAM) Staining
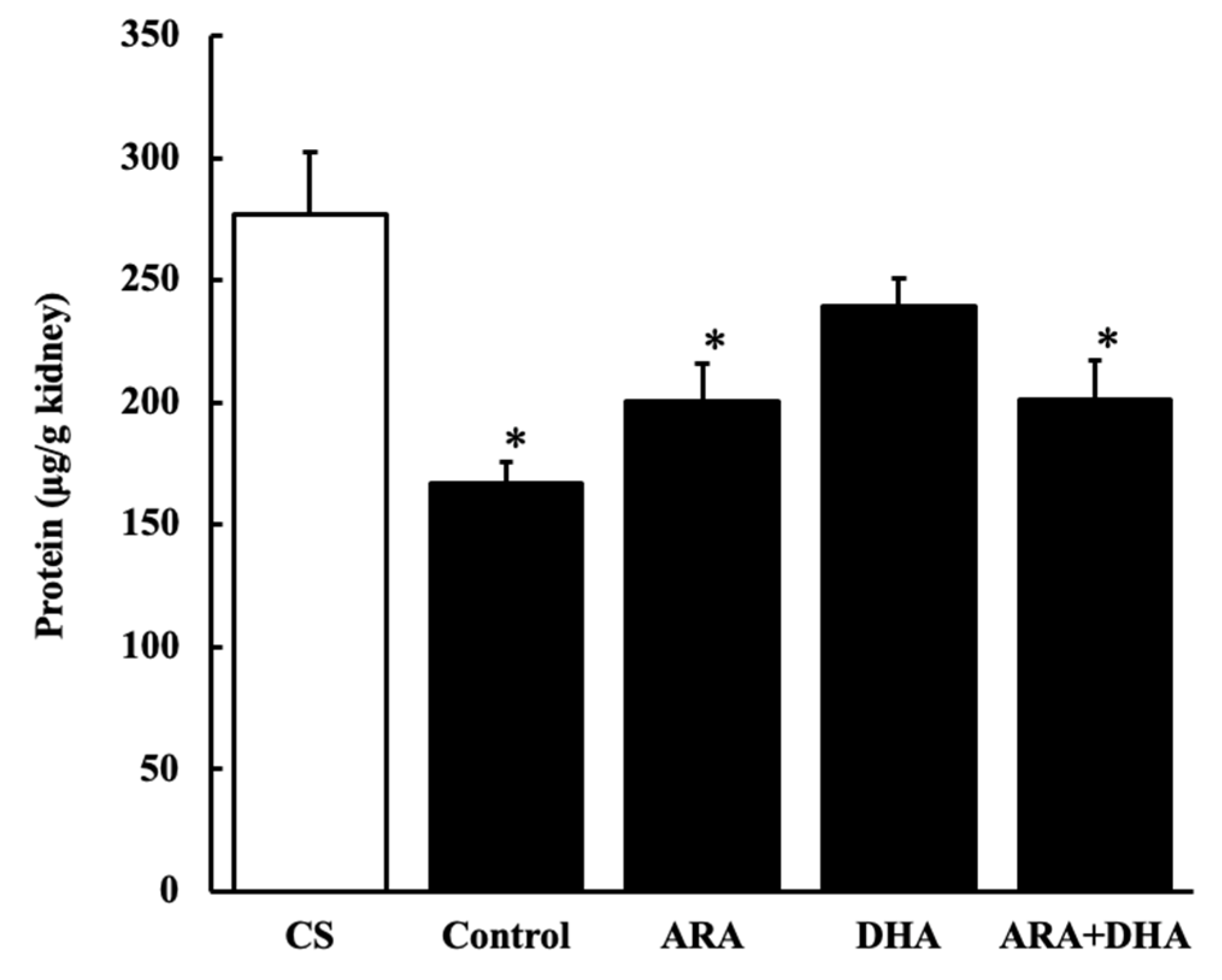
2.5. Protein Levels in Kidney
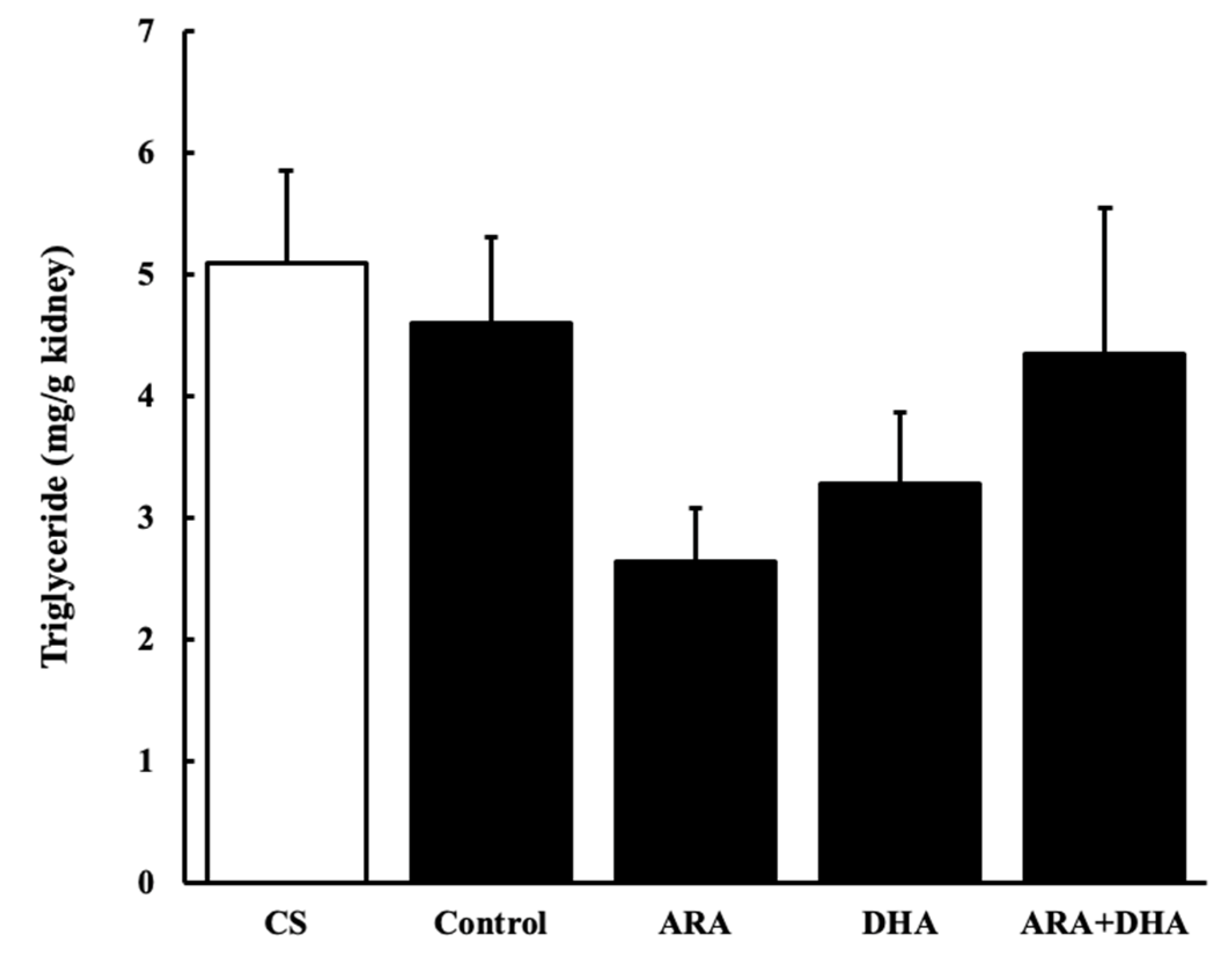
2.6. Triglycerides Levels in the Kidney
2.7. Oxidative Stress Status of the Kidney
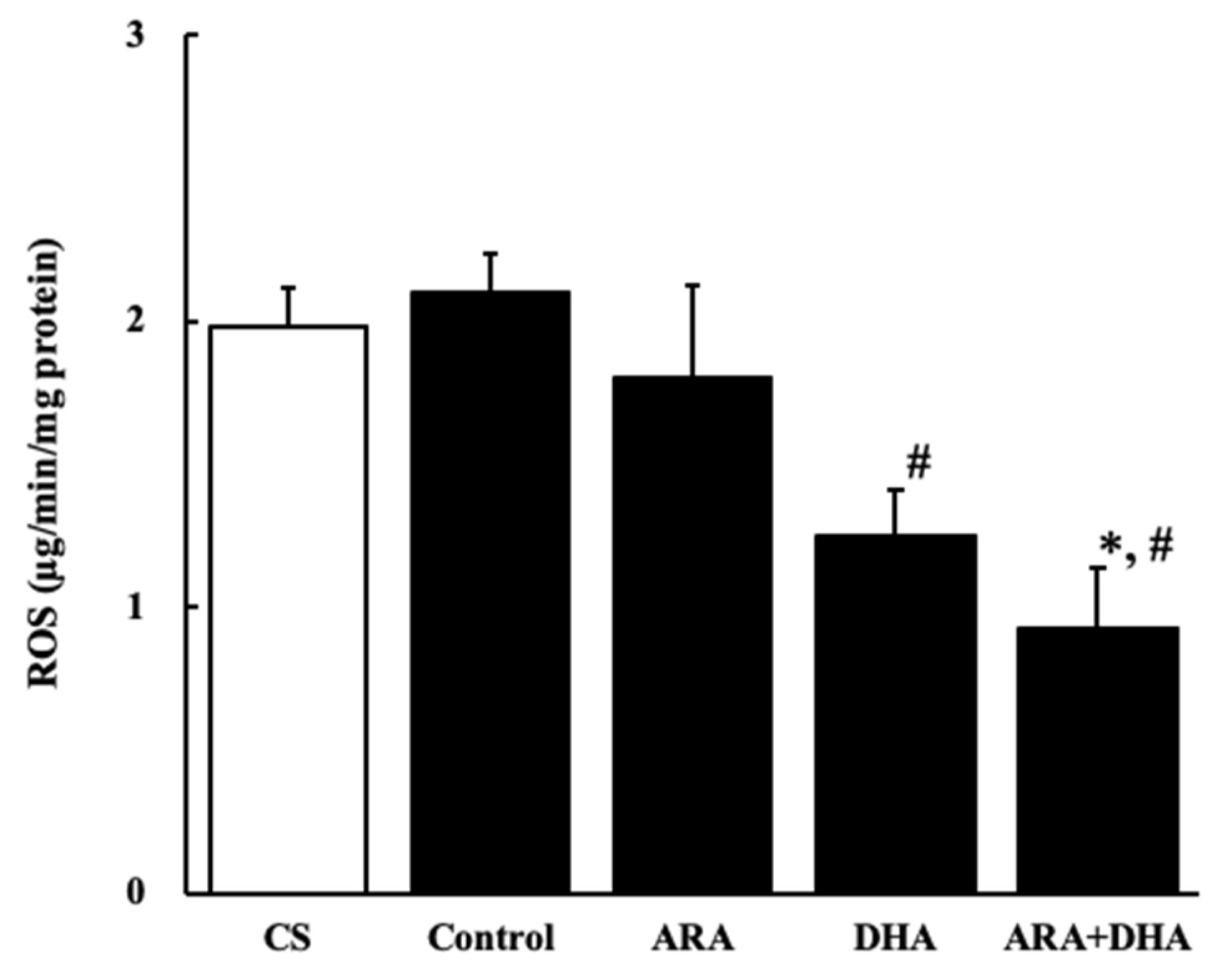
2.7.1. Reactive Oxygen Species (ROS) Level
2.7.2. Peroxynitrate (ONOO−) Level
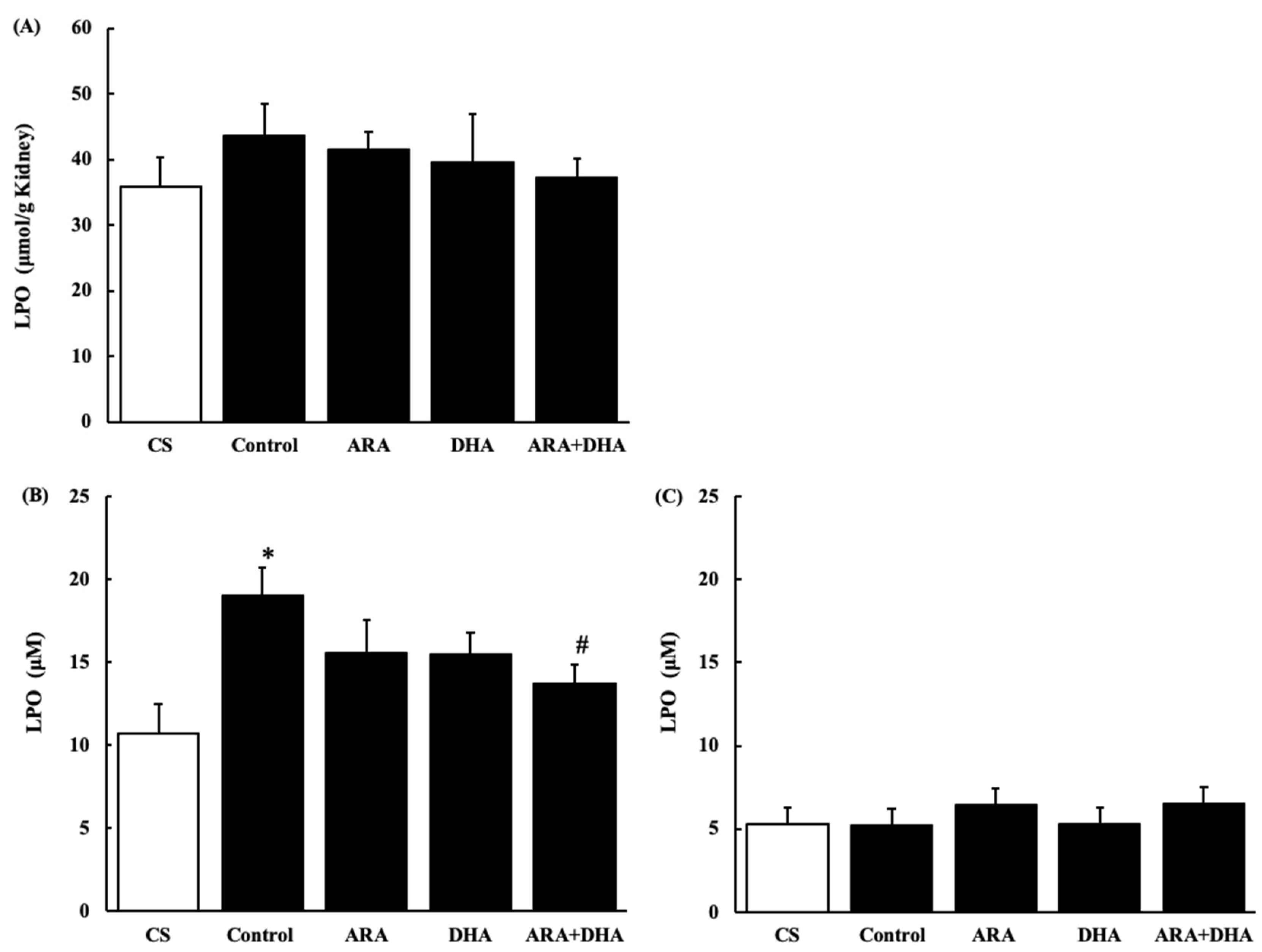
2.7.3. Lipid Peroxides (LPO) Level in the Kidney and Plasma
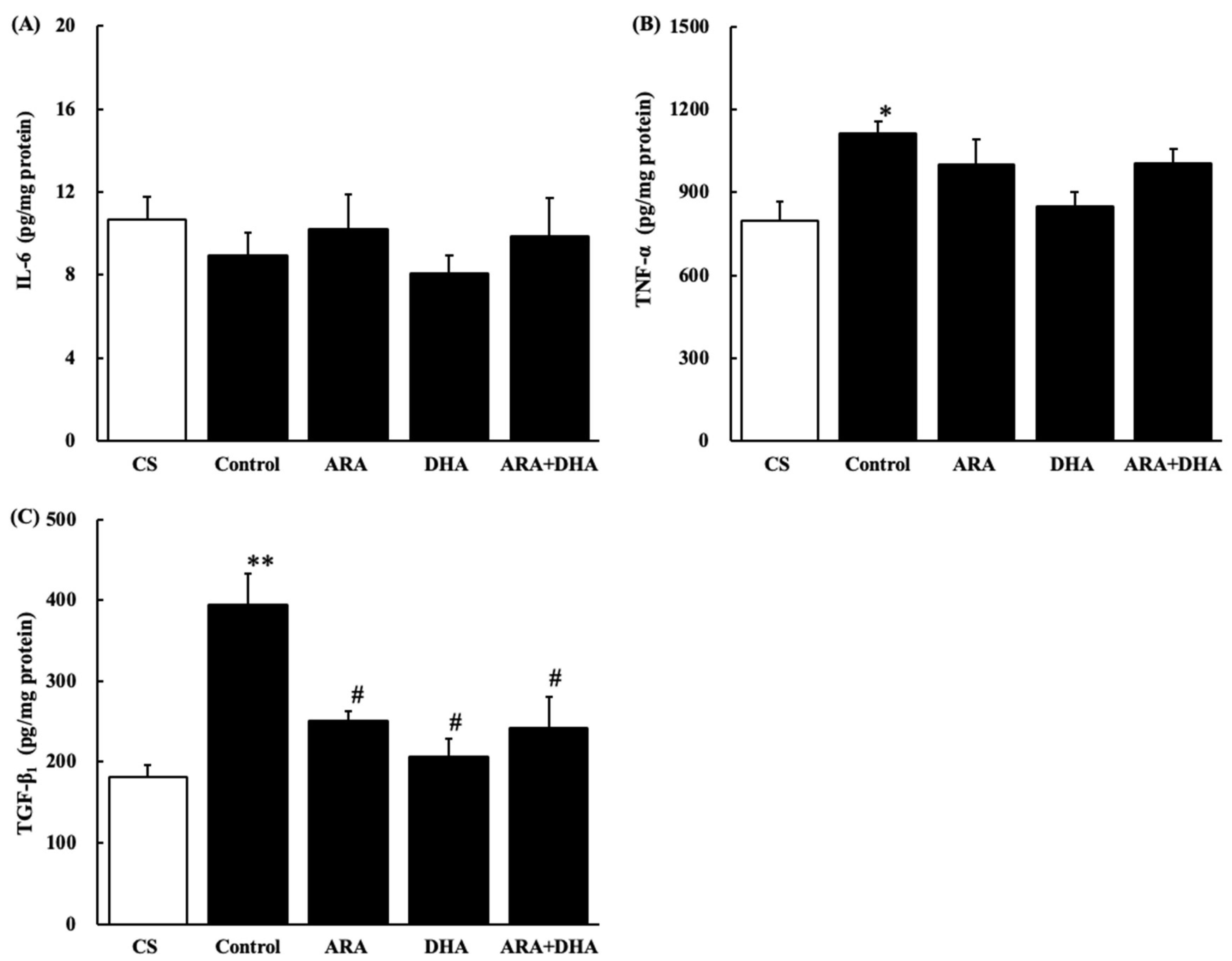
2.8. Cytokine Levels in the Kidney
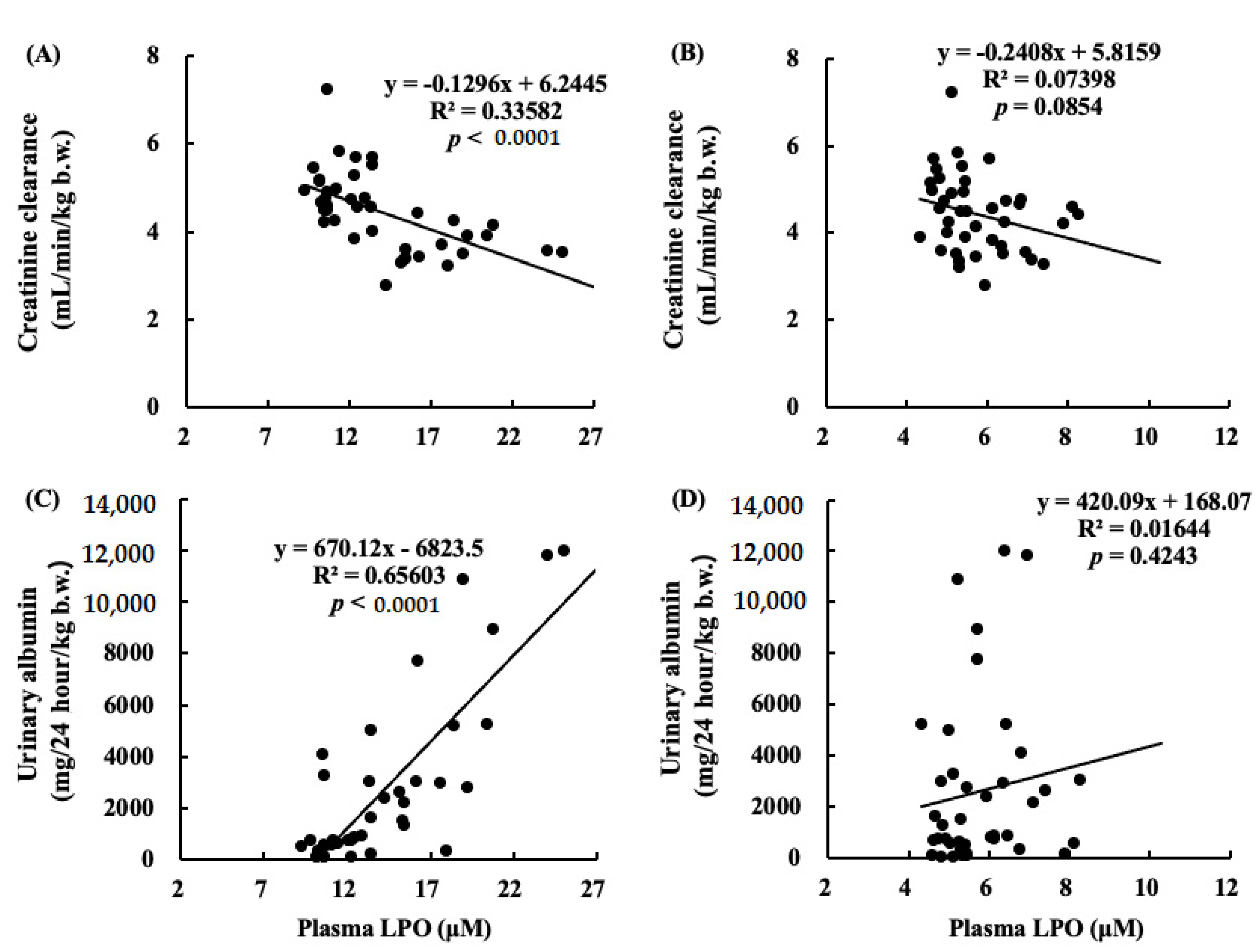
2.9. Correlations between Renal Functions and LPO Levels in Plasma
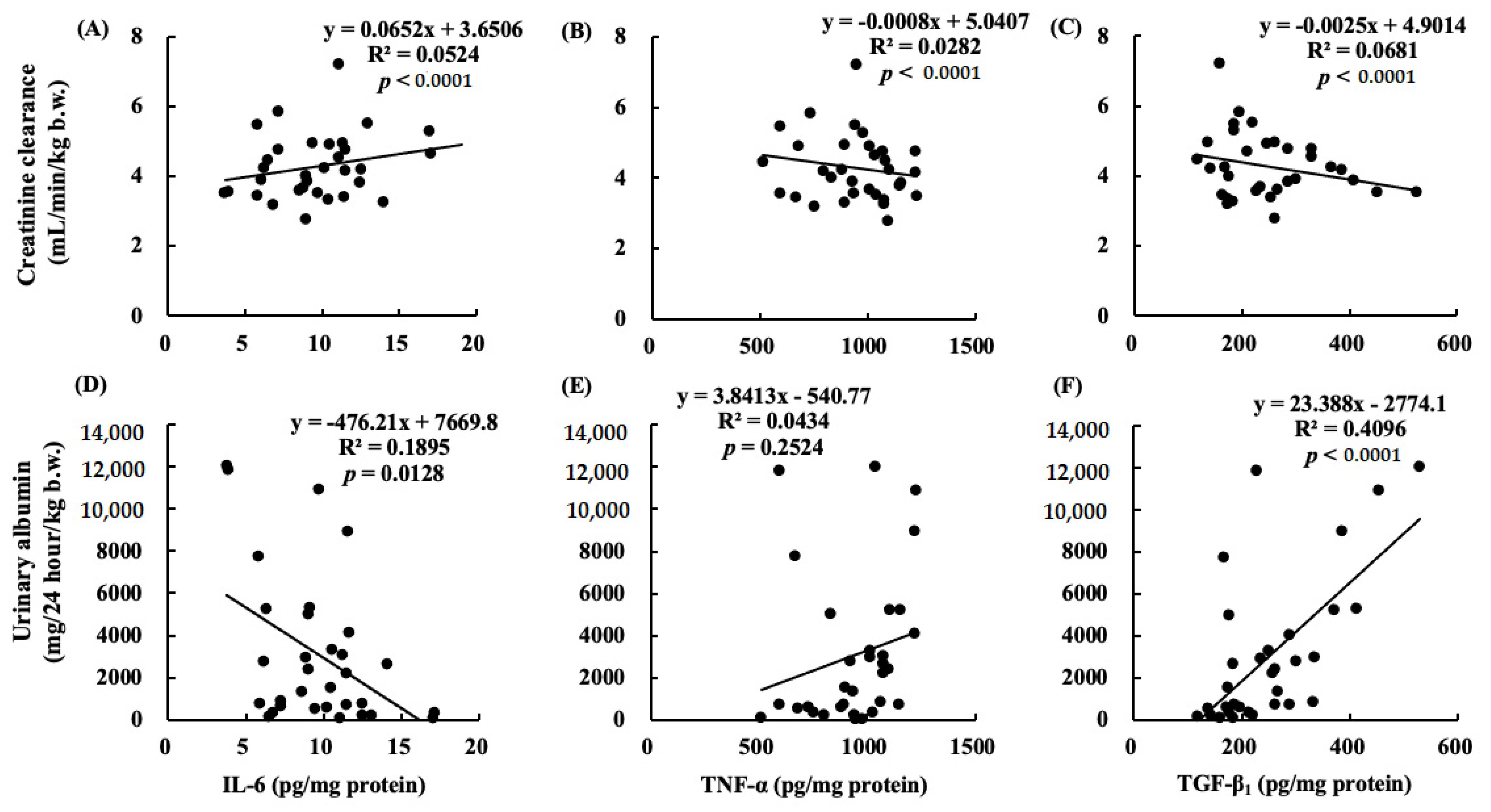
2.10. Correlations between Renal Functions and Cytokines
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Diets
4.3. Nephrectomy
4.4. Calculation of Creatinine Clearance
4.5. Sample Collection
4.6. Histological Evaluation
4.7. Sample Preparation
4.8. Analysis of Fatty Acids Composition
4.8.1. Sample Preparation for Gas Chromatography
4.8.2. GC Analysis
4.9. Quantification of Protein Level in the Kidneys
4.10. Quantification TG in Plasma and Kidneys
4.11. Analysis of Oxidative Stress Status
4.11.1. ROS Levels in the Kidney
4.11.2. ONOO− levels in the Kidney
4.11.3. LPO Levels in Plasma and Kidney
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, X.J.; Rakheja, D.; Yu, X.; Saxena, R.; Vaziri, N.D.; Silva, F.G. The aging kidney. Kidney Int. 2008, 74, 710–720. [Google Scholar] [CrossRef] [Green Version]
- Fukui, A.; Yokoo, T.; Nangaku, M.; Kashihara, N. New measures against chronic kidney diseases in Japan since 2018. Clin. Exp. Nephrol. 2019, 23, 1263–1271. [Google Scholar] [CrossRef] [Green Version]
- Katakura, M.; Hashimoto, M.; Inoue, T.; Mamun, A.A.; Tanabe, Y.; Arita, M.; Shido, O. Chronic arachidonic acid administration decreases docosahexaenoic acid-and eicosapentaenoic acid-derived metabolites in kidneys of aged rats. PLoS ONE 2015, 10, e0140884. [Google Scholar]
- Kielar, M.L.; Jeyarajah, D.R.; Zhou, X.J.; Lu, C.Y. Docosahexaenoic Acid Ameliorates Murine Ischemic Acute Renal Failure and Prevents Increases in mRNA Abundance for both TNF-α and Inducible Nitric Oxide Synthase. J. Am. Soc. Nephrol. 2003, 14, 389–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Q.; Zhou, H.-R.; Bennink, M.; Pestka, J.J. Nutritional Immunology Docosahexaenoic Acid Attenuates Mycotoxin-Induced Immunoglobulin A Nephropathy, Interleukin-6 Transcription, and Mitogen-Activated Protein Kinase Phosphorylation in Mice 1. J. Nutr. 2004, 134, 3343–3349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Q.; Shi, Y.; Bennink, M.B.; Pestka, J.J. Nutritional Immunology Docosahexaenoic Acid and Eicosapentaenoic Acid, but Not-Linolenic Acid, Suppress Deoxynivalenol-Induced Experimental IgA Nephropathy in Mice 1. J. Nutr. 2004, 134, 1353–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parinyasiri, U.; Ong-Ajyooth, L.; Parichatikanond, P.; Ong-Ajyooth, S.; Liammongkolkul, S.; Kanyog, S. Effect of fish oil on oxidative stress, lipid profile and renal function in IgA nephropathy. J. Med Assoc. Thail. Chotmaihet Thangphaet 2004, 87, 143–149. [Google Scholar]
- Mariee, A.D.; Abd-Ellah, M.D. Protective Effect of Docosahexaenoic Acid Against Cyclosporine A-Induced Nephrotoxicity in Rats: A Possible Mechanism of Action. Ren. Fail. 2011, 33, 66–71. [Google Scholar] [CrossRef]
- Thakkar, R.R.; Wang, O.-L.; Zerouga, M.; Stillwell, W.; Haq, A.; Kissling, R.; Pierce, W.M.; Smith, N.B.; Miller, F.N.; Ehringer, W.D. Docosahexaenoic acid reverses cyclosporin A-induced changes in membrane structure and function. Biochim. Biophys. Acta (BBA) Gen. Subj. 2000, 1474, 183–195. [Google Scholar] [CrossRef]
- Taneda, S.; Honda, K.; Tomidokoro, K.; Uto, K.; Nitta, K.; Oda, H. Eicosapentaenoic acid restores diabetic tubular injury through regulating oxidative stress and mitochondrial apoptosis. Am. J. Physiol. Physiol. 2010, 299, F1451–F1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chin, H.J.; Fu, Y.Y.; Ahn, J.M.; Na, K.Y.; Kim, Y.S.; Kim, S.; Chae, D.-W. Omacor(R), n-3 polyunsaturated fatty acid, attenuated albuminuria and renal dysfunction with decrease of SREBP-1 expression and triglyceride amount in the kidney of type II diabetic animals. Nephrol. Dial. Transplant. 2009, 25, 1450–1457. [Google Scholar] [CrossRef] [PubMed]
- Bays, H. Clinical Overview of Omacor: A Concentrated Formulation of Omega-3 Polyunsaturated Fatty Acids. Am. J. Cardiol. 2006, 98, 71–76. [Google Scholar] [CrossRef]
- Katakura, M.; Hashimoto, M.; Inoue, T.; Al Mamun, A.; Tanabe, Y.; Iwamoto, R.; Arita, M.; Tsuchikura, S.; Shido, O. Omega-3 Fatty Acids Protect Renal Functions by Increasing Docosahexaenoic Acid-Derived Metabolite Levels in SHR.Cg-Leprcp/NDmcr Rats, a Metabolic Syndrome Model. Molecules 2014, 19, 3247–3263. [Google Scholar] [CrossRef] [Green Version]
- Garman, J.H.; Mulroney, S.; Manigrasso, M.; Flynn, E.; Maric, C. Omega-3 fatty acid rich diet prevents diabetic renal disease. Am. J. Physiol. Physiol. 2009, 296, F306–F316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, D.B.; Christman, J.W.; Badr, K.F. Fifteen-S-hydroxyeicosatetraenoic acid (15-S-HETE) specifically antagonizes the chemotactic action and glomerular synthesis of leukotriene B4 in the rat. Kidney Int. 1992, 41, 1155–1160. [Google Scholar] [CrossRef] [Green Version]
- Ling, X.C.; Kuo, K.-L. Oxidative stress in chronic kidney disease. Ren. Replace. Ther. 2018, 4, 53. [Google Scholar] [CrossRef] [Green Version]
- Konta, T. Clinical significance of proteinuria and renal function: Findings from a population-based cohort, the Takahata study. Rinsho Byori. Jpn. J. Clin. Pathol. 2013, 61, 629–634. [Google Scholar]
- Jefferson, J.; Shankland, S.; Pichler, R. Proteinuria in diabetic kidney disease: A mechanistic viewpoint. Kidney Int. 2008, 74, 22–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garg, P. A Review of Podocyte Biology. Am. J. Nephrol. 2018, 47, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, S.; Kobori, H.; Ohashi, N.; Urushihara, M.; Nishiyama, A.; Mori, T.; Ishizuka, T.; Nako, K.; Ito, S. Angiotensin II Type 1 Receptor Blockers Reduce Urinary Angiotensinogen Excretion and the Levels of Urinary Markers of Oxidative Stress and Inflammation in Patients with Type 2 Diabetic Nephropathy. Biomark. Insights 2009, 4, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Martens, C.R.; Edwards, D.G. Peripheral Vascular Dysfunction in Chronic Kidney Disease. Cardiol. Res. Pract. 2011, 2011, 267257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attman, P.-O.; Nyberg, G.; William-Olsson, T.; Knight-Gibson, C.; Alaupovic, P. Dyslipoproteinemia in diabetic renal failure. Kidney Int. 1992, 42, 1381–1389. [Google Scholar] [CrossRef] [Green Version]
- Rutledge, J.C.; Ng, K.F.; Aung, H.H.; Wilson, D.W. Role of triglyceride-rich lipoproteins in diabetic nephropathy. Nat. Rev. Nephrol. 2010, 6, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Cheung, W.W.; Paik, K.H.; Mak, R.H. Inflammation and cachexia in chronic kidney disease. Pediatr. Nephrol. 2010, 25, 711–724. [Google Scholar] [CrossRef] [PubMed]
- Quirin, N.; Feige, J.-J.; Zaoui, P. TGF-beta: Usage in nephrology. Nephrologie 1996, 17, 227–235. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Weeks | CS | Control | ARA | DHA | ARA + DHA | |
|---|---|---|---|---|---|---|
| 0 | 175.9 ± 11.7 | 151.2 ± 4.4 | 149.3 ± 5.8 | 138.3 ± 7.6 | 155.7 ± 2.8 | |
| Blood glucose | 4 | 170.1 ± 13.9 | 181.3 ± 6.0 | 180.8 ± 7.4 | 168.0 ± 9.1 | 176.8 ± 13.6 |
| (mg/dL) | 8 | 203.0 ± 12.0 | 187.0 ± 6.4 | 164.3 ± 8.1 | 177.7 ± 6.9 | 174.0 ± 5.8 |
| 12 | 186.9 ± 5.7 | 174.2 ± 6.5 | 177.5 ± 7.0 | 183.5 ± 4.7 | 189.5 ± 7.5 | |
| 0 | 31.2 ± 5.0 | 27.2 ± 4.1 | 24.3 ± 4.4 | 20.0 ± 2.1 | 26.5 ± 8.1 | |
| Urinary glucose | 4 | 37.7 ± 8.3 | 34.5 ± 5.3 | 34.3 ± 4.4 | 31.8 ± 4.1 | 33.8 ± 2.2 |
| (mg/dL) | 8 | 43.7 ± 5.3 | 41.7 ± 3.5 | 28.5 ± 2.8 | 35.0 ± 2.1 | 34.8 ± 1.2 |
| 12 | 37.7 ± 8.6 | 29.5 ± 3.6 | 40.7 ± 6.8 | 37.8 ± 5.2 | 38.3 ± 4.3 | |
| 0 | 7.7 ± 0.2 a | 4.9 ± 0.2 b | 4.7 ± 0.3 b | 4.3 ± 0.2 b | 4.6 ± 0.3 b | |
| Creatinine clearance | 4 | 5.9 ± 0.2 | 5.2 ± 0.2 | 5.1 ± 0.2 | 4.5 ± 0.2 | 4.3 ± 0.2 |
| (mL/min/kg b.w.) | 8 | 5.9 ± 0.3 a | 4.3 ± 0.3 b | 4.5 ± 0.2 b | 3.9 ± 0.2 b | 4.6 ± 0.2 b |
| 12 | 5.1 ± 0.2 a | 3.9 ± 0.2 b | 4.1 ± 0.3 b | 3.8 ± 0.3 b | 3.9 ± 0.3 b | |
| 0 | 0.1 ± 0.0 | 0.2 ± 0.1 | 0.1 ± 0.0 | 0.1 ± 0.0 | 0.1 ± 0.0 | |
| Urinary albumin | 4 | 0.1 ± 0.9 | 2.1 ± 0.9 | 1.4 ± 1.0 | 0.6 ± 0.0 | 0.6 ± 0.3 |
| (g/24 hour/kg) | 8 | 0.2 ± 0.1 a | 5.0 ± 1.6 b | 1.9 ± 0.8 a | 1.3 ± 0.1 a | 1.2 ± 0.5 a |
| 12 | 0.6 ± 0.2 a | 11.4 ± 3.0 b | 5.9 ± 2.6 a,b,c | 5.6 ± 0.3 a,b,c | 3.0 ± 1.2 a,c |
| (%) | CS | Control | ARA | DHA | ARA + DHA |
|---|---|---|---|---|---|
| C16:0 | 26.3 ± 1.9 | 27.8 ± 0.9 | 23.7 ± 4.1 | 27.9 ± 0.7 | 27.7 ± 0.3 |
| C16:1 | 0.7 ± 0.4 | 0.4 ± 0.1 | 0.6 ± 0.4 | 0.4 ± 0.0 | 0.3 ± 0.1 |
| C18:0 | 10.4 ± 1.0 | 11.5 ± 0.5 | 11.2 ± 0.8 | 12.0 ± 0.8 | 9.7 ± 0.3 |
| C18:1 | 21.0 ± 1.8 a | 18.2 ± 1.4 a,b | 16.0 ± 1.1 a,b | 15.3 ± 0.6 b | 13.1 ± 0.4 b |
| C18:2ω-6 | 10.7 ± 0.9 b,c | 12.5 ± 0.3 a,b | 8.4 ± 0.6 c | 13.7 ± 0.3 a | 8.6 ± 0.3 c |
| C20:0 | 1.1 ± 0.2 | 0.9 ± 0.3 | 1.0 ± 0.4 | 0.4 ± 0.2 | 0.3 ± 0.1 |
| C20:4ω-6 | 26.5 ± 5.8 | 25.3 ± 1.9 | 36.3 ± 3.0 | 22.7 ± 1.4 | 34.2 ± 0.7 |
| C20:5ω-3 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.3 ± 0.2 | 0.0 ± 0.0 | 0.4 ± 0.2 |
| C22:0 | 0.2 ± 0.2 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| C22:6ω-3 | 3.2 ± 0.3 b | 3.4 ± 0.7 b | 2.4 ± 0.5 b | 7.6 ± 0.4 a | 5.7 ± 0.4 a |
| ω-6/ω-3 | 12.2 ± 6.2 | 5.0 ± 1.0 | 10.1 ± 2.2 | 3.1 ± 0.3 | 5.7 ± 0.4 |
| (%) | CS | Control | ARA | DHA | ARA + DHA |
|---|---|---|---|---|---|
| C8:0 | 0.3 ± 0.3 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.1 ± 0.1 |
| C10:0 | 0.8 ± 0.3 | 0.7 ± 0.0 | 0.4 ± 0.0 | 0.7 ± 0.1 | 0.6 ± 0.1 |
| C12:0 | 0.5 ± 0.4 | 0.1 ± 0.0 | 0.1 ± 0.0 | 0.1 ± 0.1 | 0.3 ± 0.1 |
| C14:0 | 1.0 ± 0.4 | 0.7 ± 0.1 | 0.7 ± 0.1 | 0.6 ± 0.1 | 1.0 ± 0.2 |
| C16:0 | 26.0 ± 0.8 | 27.7 ± 0.3 | 27.6 ± 0.4 | 27.3 ± 0.4 | 28.2 ± 0.9 |
| C16:1 | 1.7 ± 0.2 | 1.5 ± 0.2 | 2.0 ± 0.3 | 1.4 ± 0.3 | 2.4 ± 0.6 |
| C18:0 | 16.9 ± 0.7 | 16.5 ± 0.7 | 16.9 ± 0.7 | 17.0 ± 0.6 | 15.6 ± 1.3 |
| C18:1 | 15.6 ± 1.2 | 16.3 ± 1.3 | 16.5 ± 1.4 | 14.4 ± 1.4 | 16.1 ± 3.2 |
| C18:2ω-6 | 8.0 ± 0.3 b | 8.3 ± 0.2 b | 5.2 ± 0.3 c | 9.7 ± 0.2 a | 6.1 ± 0.2 c |
| C18:3ω-3 | 0.8 ± 0.2 | 0.7 ± 0.1 | 0.8 ± 0.1 | 0.3 ± 0.1 | 0.6 ± 0.1 |
| C20:0 | 1.0 ± 0.4 | 0.7 ± 0.1 | 0.7 ± 0.1 | 0.7 ± 0.1 | 0.8 ± 0.2 |
| C20:4ω-6 | 18.9 ± 1.3 | 17.9 ± 1.1 | 22.3 ± 1.3 | 17.0 ± 1.0 | 19.6 ± 2.3 |
| C22:0 | 2.5 ± 0.3 a,b | 2.7 ± 0.2 a,b | 1.8 ± 0.1 b | 3.5 ± 0.2 a | 2.3 ± 0.3 b |
| C24:0 | 3.1 ± 0.3 | 2.8 ± 0.2 | 2.7 ± 0.2 | 3.1 ± 0.2 | 2.7 ± 0.3 |
| C22:6ω-3 | 2.7 ± 0.3 b,c | 3.2 ± 0.2 a,b,c | 2.2 ± 0.2 c | 4.2 ± 0.2 a | 3.7 ± 0.5 a,b |
| ω-6/ω-3 | 8.0 ± 0.8 a,b | 6.7 ± 0.3 b,c | 8.9 ± 0.2 a | 6.1 ± 0.2 c | 6.0 ± 0.3 c |
| (%) | Control | ARA | DHA | ARA + DHA |
|---|---|---|---|---|
| PLA (16:0) | 27.6 | 27.2 | 27.3 | 28.1 |
| STA (18:0) | 4.2 | 4.6 | 4.4 | 4.8 |
| OLA (18:1) | 31.5 | 29.8 | 29.9 | 28.8 |
| LA (18:2ω-6) | 22.3 | 17.7 | 21.7 | 16.4 |
| ALA (18:3ω-3) | 11.3 | 11.6 | 6.2 | 5.8 |
| ARA (20:4ω-6) | 0.0 | 4.1 | 0.2 | 0.8 |
| EPA (20:5ω-3) | 0.0 | 0.0 | 0.8 | 0.8 |
| DHA (22:6ω-3) | 0.0 | 0.0 | 4.0 | 4.0 |
| SFA | 33.1 | 34.2 | 33.4 | 35.6 |
| MUFA | 31.9 | 30.3 | 31.3 | 30.2 |
| PUFA | 34 | 34.6 | 33.8 | 32.7 |
| ω-6/ω-3 | 1.98 | 1.91 | 2.01 | 2.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muramatsu, H.; Akimoto, N.; Hashimoto, M.; Sugibayashi, K.; Katakura, M. Influence of Polyunsaturated Fatty Acid Intake on Kidney Functions of Rats with Chronic Renal Failure. Mar. Drugs 2021, 19, 692. https://doi.org/10.3390/md19120692
Muramatsu H, Akimoto N, Hashimoto M, Sugibayashi K, Katakura M. Influence of Polyunsaturated Fatty Acid Intake on Kidney Functions of Rats with Chronic Renal Failure. Marine Drugs. 2021; 19(12):692. https://doi.org/10.3390/md19120692
Chicago/Turabian StyleMuramatsu, Hiroki, Naoe Akimoto, Michio Hashimoto, Kenji Sugibayashi, and Masanori Katakura. 2021. "Influence of Polyunsaturated Fatty Acid Intake on Kidney Functions of Rats with Chronic Renal Failure" Marine Drugs 19, no. 12: 692. https://doi.org/10.3390/md19120692
APA StyleMuramatsu, H., Akimoto, N., Hashimoto, M., Sugibayashi, K., & Katakura, M. (2021). Influence of Polyunsaturated Fatty Acid Intake on Kidney Functions of Rats with Chronic Renal Failure. Marine Drugs, 19(12), 692. https://doi.org/10.3390/md19120692