
Investigation of Deep-Sea Ecosystems Using Marker Fatty Acids: Sources of Essential Polyunsaturated Fatty Acids in Abyssal Megafauna

Abstract
:1. Introduction: General Characteristics of the Abyssal Zone
2. Some Notes on Samples Used for Lipid Extraction and the Methods of Fatty Acid Analysis
3. Fatty Acids of Abyssal Bottom Sediments
4. Fatty Acids of Foraminifera
5. Fatty Acids of Nematodes
6. Abyssal Megafauna
6.1. Holothurians
6.2. Sea Stars, Brittle Stars, and Sea Urchins
6.3. Mollusks
6.4. Polychaetes
7. Discussion: Sources of PUFA in Abyssal Ecosystems
8. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| FA | Fatty acid(s) |
| FAME | Fatty acid methyl ester(s) |
| SFA | Saturated FA |
| MUFA | Monounsaturated FA: FA with 1 double bond |
| NMID | Non-methylene interrupted dienoic FA |
| PUFA | Polyunsaturated FA: FA with 2 or more double bonds |
| LCFA | Long-chain FA: FA with 24 or more carbon atoms |
| C18, C20, C22 | Refers to FA with the number of carbons in a molecule, e.g., C18 polyenoic FA |
| Bacterial FA | Odd and branched-chain FA, e.g., 15:0, i-17:0 |
References
- Bruun, A.F. Deep sea and abyssal depths. Geol. Soc. America Memoir. 1957, 67, 641–672. [Google Scholar]
- Smith, C.R.; De Leo, F.C.; Bernardino, A.F.; Sweetman, A.K.; Arbizu, P.M. Abyssal food limitation, ecosystem structure and climate change. Trends Ecol. Evol. 2008, 23, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Thistle, D. The Deep-Sea Floor: An Overview; Ecosystems of the World; Elsevier: Amsterdam, The Netherlands, 2003; Chapter 2; pp. 5–37. [Google Scholar]
- Smith, C.; Demopoulos, A. The Deep Pacific Ocean Floor; Ecosystems of the World; Elsevier: Amsterdam, The Netherlands, 2003; Chapter 6; Volume 27, pp. 179–218. [Google Scholar]
- Rowe, G.T.; Gardner, W. Sedimentation rates in the slope water of the northwest Atlantic Ocean measured directly with sediment traps. J. Mar. Res. 1979, 37, 581–600. [Google Scholar]
- Honjo, S.; Dymond, J.; Collier, R.; Manganini, S.J. Export production of particles to the interior of equatorial Pacific Ocean during the 1992 EqPac experiment. Deep-Sea Res. II 1995, 42, 831–870. [Google Scholar] [CrossRef]
- Smith, C.R.; Berelson, W.; Demaster, D.J.; Dobbs, F.C.; Hammond, D.; Hoover, D.J.; Pope, R.H.; Stephens, M. Latitudinal variations in benthic processes in the abyssal equatorial Pacific: Control by biogenic particle flux. Deep-Sea Res. II 1997, 44, 2295–2317. [Google Scholar] [CrossRef]
- Wakeham, S.G.; Hedges, J.I.; Lee, C.; Peterson, M.L.; Hernes, P.J. Compositions and transport of lipid biomarkers through the water column and surficial sediments of the equatorial Pacific Ocean. Deep-Sea Res. II 1997, 44, 2131–2162. [Google Scholar] [CrossRef]
- Kiriakoulakis, K.; Stutt, E.; Rowland, S.J.; Vangriesheim, A.; Lampitt, R.S.; Wolff, G.A. Controls on the organic chemical composition of settling particles in the northeast Atlantic Ocean. Prog. Oceanogr. 2001, 50, 65–87. [Google Scholar] [CrossRef]
- Azam, F.; Fenchel, T.; Field, J.G.; Gray, J.S.; Meyer-Reil, L.A.; Thingstad, F. The ecological role of water column microbes in the sea. Mar. Ecol. Prog. Ser. 1983, 10, 257–263. [Google Scholar] [CrossRef]
- Parrish, C.C. Essential Fatty Acids in Aquatic Food Webs. In Lipids in Aquatic Ecosystems; Arts, M.T., Brett, M.T., Kainz, M.J., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 309–326. [Google Scholar]
- Bell, M.V.; Tocher, D.R. Biosynthesis of Polyunsaturated Fatty Acids in Aquatic Ecosystems: General Pathways and New Directions. In Lipids in Aquatic Ecosystems; Arts, M.T., Brett, M.T., Kainz, M.J., Eds.; Springer: The Netherlands, Germany, 2008; pp. 211–236. [Google Scholar]
- Monroig, Ó.; Tocher, D.R.; Navarro, J.C. Biosynthesis of polyunsaturated fatty acids in marine invertebrates: Recent advances in molecular mechanisms. Mar Drugs. 2013, 11, 3998–4018. [Google Scholar] [CrossRef] [Green Version]
- Sinensky, M. Homeoviscous adaptation: A homeostatic process that regulates the viscosity of the membrane lipids in Escherichia coli. Proc. Natl. Acad. Sci. USA 1974, 71, 522–526. [Google Scholar] [CrossRef] [Green Version]
- Cossins, A.R.; Macdonald, A.G. Homeoviscous adaptation under pressure. III. The fatty acid composition of liver mitochondrial phospholipids of deep-sea fish. Biochim. Biophys. Acta. 1986, 860, 325–335. [Google Scholar] [CrossRef]
- De Long, E.F.; Yayanos, A.A. Adaptation of the membrane lipids of a deep-sea bacterium to changes in hydrostatic pressure. Science 1985, 228, 1101–1102. [Google Scholar] [CrossRef] [PubMed]
- Stránsky, K.; Jursik, T.; Vitek, A. Standard equivalent chain length values of monoenic and polyenic (methylene interrupted) fatty acids. J. High Resolut. Chrom. 1997, 20, 143–158. [Google Scholar] [CrossRef]
- Sebedio, J.L.; Ackman, R.G. Calculation of retention data for some accessible C20 isomeric cis-unsaturated fatty acids. J. Chromatogr. Sci. 1982, 20, 231–234. [Google Scholar] [CrossRef]
- Wolff, R.L.; Christie, W.W. Structures, practical sources (gymnosperm seeds), gas-liquid chromatographic data (equivalent chain lengths), and mass spectrometric characteristics of all-cis ∆5-olefinic acids. Eur. J. Lipid Sci. Technol. 2002, 104, 234–244. [Google Scholar] [CrossRef]
- Kharlamenko, V.I.; Svetashev, V.I.; Tarasova, T.S. New and uncommon fatty acids in lipids of deep-sea foraminifera. Lipids 2017, 52, 345–352. [Google Scholar] [CrossRef]
- Kharlamenko, V.I.; Odintsova, N.A. Unusual methylene-interrupted polyunsaturated fatty acids of abyssal and hadal invertebrates. Progr. Oceanogr. 2019, 178, 102132. [Google Scholar] [CrossRef]
- Svetashev, V.I.; Kharlamenko, V.I. Fatty Acids of Abyssal Echinodermata, the Sea Star Eremicaster vicinus and the Sea Urchin Kamptosoma abyssale: A New Polyunsaturated Fatty Acid Detected, 22:6(n-2). Lipids 2020, 55, 291–296. [Google Scholar] [CrossRef]
- Svetashev, V.I. Mild method for preparation of 4,4-dimethyloxazoline derivatives of polyunsaturated fatty acids for GC–MS. Lipids 2011, 46, 463–467. [Google Scholar] [CrossRef]
- Christie, W.W. Mass Spectrometry of Fatty Acid Derivatives. The LipidWeb. Available online: http://lipidhome.co.uk/ms/masspec.html (accessed on 22 November 2016).
- Svetashev, V.I.; Imbs, A.B. Isomerization of octadecapentaenoic acid (18:5n-3) in algal lipid samples under derivatization for GC and GC-MS analysis. J. Phycol. 2014, 50, 322–327. [Google Scholar] [CrossRef]
- Rowe, G.; Subuet, M.; Deming, J.; Khripounoff, A.; Tietjen, J.; Macko, S.; Theroux, R. ‘Total’ sediment biomass and preliminary estimates of organic carbon residence time in deep-sea benthos. Mar. Ecol. Prog. Ser. 1991, 79, 99–114. [Google Scholar] [CrossRef]
- Fowler, S.W.; Knauer, G.A. Role of Large Particles in the Transport of Elements and Organic Compounds Through the Oceanic Water Column. Prog. Oceanog. 1986, 16, 147–194. [Google Scholar] [CrossRef]
- Pinturier-Geiss, L.; Laureillard, J.; Riaux-Gobin, C.; Fillaux, J.; Saliot, A. Lipids and pigments in deep-sea surface sediments and interfacial particles from the Western Crozet Basin. Mar. Chem. 2001, 75, 249–266. [Google Scholar] [CrossRef]
- Laureillard, J.; Méjanelle, L.; Sibuet, M. Use of lipids to study the trophic ecology of deep-sea xenophyophores. Mar. Ecol. Prog. Ser. 2004, 270, 129–140. [Google Scholar] [CrossRef]
- Würzberg, L.; Peters, J.; Schuller, M.; Brandt, A. Diet insights of deep-sea polychaetes derived from fatty acid analyses. Deep-Sea Res. Part II—Top. Stud. Oceanogr. 2011, 58, 153–162. [Google Scholar] [CrossRef]
- Würzberg, L.; Peters, J.; Brandt, A. Fatty acid patterns of Southern Ocean shelf and deep sea peracarid crustaceans and a possible food source, foraminiferans. Deep-Sea Res. Part II—Top. Stud. Oceanogr. 2011, 58, 2027–2035. [Google Scholar] [CrossRef]
- Leduc, D.; Brown, J.C.S.; Bury, S.J.; Lörz, A.-N. High intraspecific variability in the diet of a deep-sea nematode: Stable isotope and fatty acid analyses of Deontostoma tridentum on Chatham Rise, Southwest Pacific. Deep Sea Res. Part I 2015, 97, 10–18. [Google Scholar] [CrossRef]
- Bernhard, J.M.; Sen Gupta, D.K.; Baguley, J.G. Benthic foraminifera living in Gulf of Mexico bathyal and abyssal sediments: Community analysis and comparison to metazoan meiofaunal biomass and density. Deep-Sea Res. II 2008, 55, 2617–2626. [Google Scholar] [CrossRef] [Green Version]
- Gooday, A.J.; Levin, L.A.; Linke, P.; Heeger, T. The Role of Benthic Foraminifera in Deep-Sea Food Webs and Carbon Cycling. In Deep-Sea Food Chains and the Global Carbon Cycle; Rowe, G.T., Pariente, V., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1992; pp. 63–91. [Google Scholar]
- Gooday, A.J.; Schoenle, A.; Dolan, J.R.; Arndt, H. Protist diversity and function in the dark ocean—Challenging the paradigms of deep-sea ecology with special emphasis on foraminiferans and naked protists. Eur. J. Protistol. 2020, 75, 125721. [Google Scholar] [CrossRef]
- Gooday, A.J.; Nomaki, H.; Kitazato, H. Modern deep-sea benthic foraminifera: A brief review of their morphology-based biodiversity and trophic diversity. Geol. Soc. London Spec. Public. 2008, 303, 97–119. [Google Scholar] [CrossRef]
- Moodley, L.; Middelburg, J.J.; Boschker, H.T.S.; Duineveld, G.C.A.; Pel, R.; Herman, P.M.J.; Heip, C.H.R. Bacteria and foraminifera: Key players in a short-term deep-sea benthic response to phytodetritus. Mar. Ecol. Prog. Ser. 2002, 236, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Suhr, S.B.; Stephen, P.; Alexander, S.P.; Gooday, A.J.; Pond, D.W.; Bowser, S.S. Trophic modes of large Antarctic Foraminifera: Roles of carnivory, omnivory, and detritivory. Mar. Ecol. Prog. Ser. 2008, 371, 155–164. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, S.T.; Corliss, B.H. Deposit feeding in selected deep-sea and shallow-water benthic foraminifera. Deep-Sea Res. I 1994, 41, 229–241. [Google Scholar] [CrossRef]
- Nomaki, N.; Ogawa, N.O.; Ohkouchi, N.; Suga, H.; Toyofuku, T.; Shimanaga, M.; Nakatsuka, T.; Kitazato, H. Benthic foraminifera as trophic links between phytodetritus and benthic metazoans: Carbon and nitrogen isotopic evidence. Mar. Ecol. Prog. Ser. 2008, 357, 153–164. [Google Scholar] [CrossRef] [Green Version]
- Haynert, K.; Gluderer, F.; Pollierer, M.M.; Scheu, S.; Wehrmann, A. Food Spectrum and Habitat-Specific Diets of Benthic Foraminifera from the Wadden Sea—A Fatty Acid Biomarker Approach. Front. Mar. Sci. 2020, 7, 510288. [Google Scholar] [CrossRef]
- Gooday, A.J.; Pond, D.W.; Bowser, S.S. Ecology and nutrition of the large agglutinated foraminiferan Bathysiphon capillare in the bathyal NE Atlantic: Distribution within the sediment profile and lipid biomarker composition. Mar. Ecol. Prog. Ser. 2002, 245, 69–82. [Google Scholar] [CrossRef]
- Zhukova, N.V. The pathway of the biosynthesis of non-methylene-interrupted dienoic fatty acids in molluscs. Comp. Biochem. Physiol. 1991, 100B, 801–804. [Google Scholar] [CrossRef]
- Fullarton, J.G.; Dando, P.R.; Sargent, J.R.; Southward, A.J.; Southward, E.C. Fatty acids of hydrothermal vent Ridgeia piscesae and inshore bivalves containing symbiotic bacteria. J. Mar. Biol. Assoc. UK 1995, 75, 455–468. [Google Scholar] [CrossRef]
- Larkin, K.E.; Gooday, A.J.; Woulds, C.; Jeffreys, R.; Schwartz, M.; Cowie, G.; Whitcraft, C.; Levin, L.; Dick, J.R.; Pond, D.W. Uptake of algal carbon and the synthesis of an “essential” fatty acid by Uvigerina ex. gr. semiornata (Foraminifera) within the Pakistan margin oxygen minimum zone: Evidence from fatty acid biomarker and 13C tracer experiments. Biogeosciences 2014, 11, 3738–3739. [Google Scholar] [CrossRef] [Green Version]
- LeKieffre, C.; Spangenberg, J.E.; Mabilleau, G.; Escrig, S.; Meibom, A.; Geslin, E. Surviving anoxia in marine sediments: The metabolic response of ubiquitous benthic foraminifera (Ammonia tepida). PLoS ONE 2017, 12, e0177604. [Google Scholar] [CrossRef] [Green Version]
- Suhr, S.B.; Pond, D.W.; Gooday, A.J.; Smith, C.R. Selective feeding by benthic foraminifera on phytodetritus on the western Antarctic Peninsula shelf: Evidence from fatty acid biomarker analysis. Mar. Ecol. Prog. Ser. 2003, 262, 153–162. [Google Scholar] [CrossRef]
- Paradis, M.; Ackman, R. Localization of a marine source of odd chain-length fatty acids I: The amphipod Pontoporeia femorata (Kröyer). Lipids 1976, 11, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.J.L.; Mansour, M.P.; Dunstan, G.A.; Blackburn, S.I.; Koutoulis, A.; Nichols, P.D. Odd-chain polyunsaturated fatty acids in thraustochytrids. Phytochemistry 2011, 72, 1460–1465. [Google Scholar] [CrossRef]
- Kharlamenko, V.I. Abyssal foraminifera as the main source of rare and new polyunsaturated fatty acids in deep-sea ecosystems. Deep-Sea Res. Part II 2018, 154, 358–364. [Google Scholar] [CrossRef]
- Ramirez-Llodra, E.; Brandt, A.; Danovaro, R.; De Mol, B.; Escobar, E.; German, C.R.; Levin, L.A.; Martínez Arbizu, P.; Menot, L.; Buhl-Mortensen, P.; et al. Deep, diverse and definitely different: Unique attributes of the world’s largest ecosystem. Biogeosciences. 2010, 7, 2851–2899. [Google Scholar] [CrossRef] [Green Version]
- Vincx, M.; Bett, B.J.; Dinet, A.; Ferrero, T.; Gooday, A.J.; Lambshead, P.J.D.; Pfannkuche, O.; Soltwedel, T.; Vanreusel, A. Meiobenthos of the Deep Northeast Atlantic. Adv. Mar. Biol. 1994, 30, 1–88. [Google Scholar] [CrossRef] [Green Version]
- Moens, T.; Braeckman, U.; Derycke, S.; Fonseca, G.; Gallucci, F.; Gingold, R.; Guilini, K.; Ingels, J.; Leduc, D.; Vanaverbeke, J.; et al. Ecology of free-living marine nematodes. In Handbook of Zoology; Nematoda, A., Ed.; De Gruyter: Berlin, Germany, 2014; Volume 3, pp. 109–152. [Google Scholar] [CrossRef]
- Tchesunov, A.V.; Ingels, J.; Popova, E.V. Marine free-living nematodes associated with symbiotic bacteria in deep-sea canyons of north-east Atlantic Ocean. J. Mar. Biol. Ass. UK 2012, 92, 1257–1274. [Google Scholar] [CrossRef]
- Rothstein, M.; Gotz, P. Biosynthesis of Fatty Acids in the Free-living Nematode, Turbatrix aceti. Arch. Biochem. Biophys. 1968, 126, 131–140. [Google Scholar] [CrossRef]
- Hutzell, P.A.; Krusberg, L.R. Fatty acid compositions of Caenorhabditis elegans and C. briggsae. Comp. Biochem. Physiol. 1982, 73B, 517–520. [Google Scholar] [CrossRef]
- Schlechtriem, C.; Tocher, D.R.; Dick, J.; Becker, K. Incorporation and metabolism of fatty acids by desaturation and elongation in the nematode, Panagrellus redivivus. Nematology 2004, 6, 783–795. [Google Scholar]
- Fodor, A.; Dey, I.; Farkas, T.; Chitwood, D.J. Effects of Temperature and Dietary Lipids on Phospholipid Fatty Acids and Membrane Fluidity in Steinernema carpocapsae. J. Nematol. 1994, 26, 278–285. [Google Scholar]
- Tanaka, T.; Ikita, K.; Ashida, T.; Motoyama, Y.; Yamaguchi, Y.; Satouchi, K. Effects of Growth Temperature on the Fatty Acid Composition of the Free-Living Nematode Caenorhabditis elegans. Lipids 1996, 31, 1173–1178. [Google Scholar] [CrossRef]
- Watts, J.L.; Browse, J. Genetic dissection of polyunsaturated fatty acid synthesis in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2002, 99, 5854–5859. [Google Scholar] [CrossRef] [Green Version]
- Braeckman, U.; Provoost, P.; Sabbe, K.; Soetaert, K.; Middelburg, J.J.; Vincx, M.; Vanaverbeke, J. Temporal dynamics in a shallow coastal benthic food web: Insights from fatty acid biomarkers and their stable isotopes. Mar. Environ. Res. 2015, 108, 55–68. [Google Scholar] [CrossRef]
- Leduc, D.; Probert, P.K. The effect of bacterivorous nematodes on detritus incorporation by macrofaunal detritivores: A study using stable isotope and fatty acid analyses. J. Exp. Mar. Biol. Ecol. 2009, 371, 130–139. [Google Scholar] [CrossRef]
- Guilini, K.; Veit-Köhler, G.; De Troch, M.; Van Gansbeke, D.; Vanreusel, A. Latitudinal and temporal variability in the community structure and fatty acid composition of deep-sea nematodes in the Southern Ocean. Prog. Oceanogr. 2013, 110, 80–92. [Google Scholar] [CrossRef]
- Lins, L.; da Silva, M.C.; Hauquier, F.; Esteves, A.M.; Vanreusel, A. Nematode community composition and feeding shaped by contrasting productivity regimes in the Southern Ocean. Prog. Oceanogr. 2015, 134, 356–369. [Google Scholar] [CrossRef]
- Van Campenhout, J.; Vanreusel, A. Closely related intertidal and deep-sea Halomonhystera species have distinct fatty acid compositions. Helgol. Mar. Res. 2016, 70, 8. [Google Scholar] [CrossRef] [Green Version]
- Mordukhovich, V.V.; Kiyashko, S.I.; Kharlamenko, V.I.; Fadeeva, N.P. Determination of food sources for nematodes in the Kuril Basin and eastern slope of the Kuril Islands by stable isotope and fatty acid analyses. Deep Sea Res. Part II: Top. Stud. Oceanogr. 2018, 154, 365–373. [Google Scholar] [CrossRef]
- Zhou, X.-R.; Green, A.G.; Singh, S.P. Caenorhabditis elegans Δ12-Desaturase FAT-2 is a bifunctional desaturase able to desaturate a diverse range of fatty acid substrates at the Δ12 and Δ15 positions. J. Biol. Chem. 2011, 286, 43644–43650. [Google Scholar] [CrossRef] [Green Version]
- Kabeya, N.; Fonseca, M.M.; Ferrier, D.E.K.; Navarro, J.C.; Bay, L.K.; Francis, D.S.; Tocher, D.R.; Castro, L.F.C.; Monroig, O. Genes for de novo biosynthesis of omega-3 polyunsaturated fatty acids are widespread in animals. Sci. Adv. 2018, 4, eaar6849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokolova, M.N. Trophic Structure of Abyssal Macrobenthos. Adv. Mar. Biol. 1992, 32, 427–525. [Google Scholar]
- Billett, D.S.M.; Bett, B.J.; Rice, A.L.; Thurston, M.H.; Galeron, J.; Sibuet, M.; Wolff, G.A. Long-term change in the megabenthos of the Porcupine Abyssal Plain (NE Atlantic). Prog. Oceanogr. 2001, 50, 325–348. [Google Scholar] [CrossRef]
- Amaro, A.; Danovaro, R.; Matsui, Y.; Eugenio Rastelli, E.; George, A.; Wolff, G.A.; Nomaki, H. Possible links between holothurian lipid compositions and differences in organic matter (OM) supply at the western Pacific abyssal plains. Deep-Sea Res. Part I 2019, 152, 103085. [Google Scholar] [CrossRef]
- Amaro, T.; Bianchelli, S.; Billett, D.S.M.; Cunha, M.R.; Pusceddu, A.; Danovaro, R. The trophic biology of the holothurian Molpadia musculus: Implications for organic matter cycling and ecosystem functioning in a deep submarine canyon. Biogeosciences 2010, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Amaro, T.; Luna, G.M.; Danovaro, R.; Billett, D.S.M.; Cunha, M.R. High prokaryotic biodiversity associated with gut contents of the holothurian Molpadia musculus from the Nazaré Canyon (NE Atlantic). Deep-Sea Res. I 2012, 63, 82–90. [Google Scholar] [CrossRef]
- Lewis, R.W. Fatty Acid Composition of Some Marine Animals from Various Depths. J. Fish. Res. 1967, 24, 1101–1115. [Google Scholar] [CrossRef]
- Ginger, M.L.; Santos, V.L.C.S.; Wolff, G.A. A preliminary investigation of the lipids of abyssal holothurians from the north-East Atlantic Ocean. J. Exp. Marine Biol. Ecol. 2000, 80, 139–146. [Google Scholar] [CrossRef]
- Hudson, I.R.; Pond, D.W.; Billett, D.S.M.; Tyler, P.A.; Lampitt, R.S.; Wolff, G.A. Temporal variations in fatty acid composition of deep-sea holothurians: Evidence of bentho-pelagic coupling. Mar. Ecol. Prog. Ser. 2004, 281, 109–120. [Google Scholar] [CrossRef]
- Drazen, J.C.; Phleger, C.F.; Guest, M.A.; Nichols, P.D. Lipid, sterols and fatty acid composition of holothurians and ophiuroids from the north-East Pacific Ocean: Food web implications. Comp. Biochem. Physiol. B. 2008, 151, 79–87. [Google Scholar] [CrossRef]
- Kaneniwa, M.; Itabashi, Y.; Endo, S.; Takagi, T. Fatty acids in Holothuroidea: Occurrence of cis-14- tricosenoic acid. Comp. Biochem. Physiol. 1986, 84B, 451–455. [Google Scholar] [CrossRef]
- Kharlamenko, V.I.; Stepanov, V.G.; Borisovets, E.E.; Kiyashko, S.I.; Svetashev, V.I. The fatty acid composition and nutrition of deep-sea holothurians from the Sea of Okhotsk. Russ. J. Mar. Biol. 2015, 41, 448–455. [Google Scholar] [CrossRef]
- Kharlamenko, V.I.; Maiorova, A.S.; Ermolenko, E.V. Fatty acid composition as an indicator of the trophic position of abyssal megabenthic deposit feeders in the Kuril Basin of the Sea of Okhotsk. Deep Sea Res. Part II Top. Stud. Oceanogr. 2018, 154, 374–382. [Google Scholar] [CrossRef]
- Howell, K.L.; Pond, D.W.; Billett, D.S.M.; Tyler, P.A. Feeding ecology of deep-sea seastars (Echinodermata: Asteroidea): A fatty-acid biomarker approach. Mar. Ecol. Prog. Ser. 2003, 255, 193–206. [Google Scholar] [CrossRef] [Green Version]
- Pearson, M.; Gage, J.D. Diets of some deep-sea brittle stars in the Rockall Trough. Mar. Biol. 1984, 82, 247–258. [Google Scholar] [CrossRef]
- Svetashev, V.I.; Kharlamenko, V.I. Occurrence of hexacosapolyenoic acids 26:7(n-3), 26:6(n-3), 26:6(n-6) and 26:5 (n-3) in deep sea brittle stars from near the Kuril Islands. Lipids 2015, 50, 691–696. [Google Scholar] [CrossRef]
- Kiyashko, S.I.; Kharlamenko, V.I.; Sanamyan, K.; Alalykina, A.L.; Würzberg, L. Trophic structure of the abyssal benthic community in the Sea of Japan inferred from stable isotope and fatty acid analyses. Mar. Ecol. Prog. Ser. 2014, 500, 121–137. [Google Scholar] [CrossRef]
- Kelly, J.R.; Scheibling, R.E.; Iverson, S.J.; Gagnon, P. Fatty acid profiles in the gonads of the sea urchin Strongylocentrotus droebachiensis on natural algal diets. Mar. Ecol. Prog. Ser. 2008, 373, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Rieley, G.; Van Dovel, C.L.; Eglinton, G. Fatty acids as sensitive tracers of sewage sludge carbon in a deep-sea ecosystem. Environ. Sci. Technol. 1997, 31, 1018–1023. [Google Scholar] [CrossRef]
- Kharlamenko, V.I.; Würzberg, L.; Peters, J.; Borisovets, E.E. Fatty acid compositions and trophic relationships of shelled molluscs from the Kuril-Kamchatka Trench and the adjacent abyssal plain. Deep-Sea Res. II 2015, 111, 389–398. [Google Scholar] [CrossRef]
- Dalsgaard, J.; St. John, M.; Kattner, G.; Muller-Navarra, D.; Hagen, W. Fatty acid trophic markers in the pelagic marine environment. Adv. Mar. Biol. 2003, 46, 225–340. [Google Scholar] [PubMed]
- Vasskog, T.; Andersen, J.H.; Hansen, E.; Svenson, J. Characterization and cytotoxicity studies of the rare 21:4n-7 acid and other polyunsaturated fatty acids from the marine Opisthobranch Scaphander lignarius, isolated using bioassay guided fractionation. Mar. Drugs. 2012, 10, 2676–2690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drazen, J.C.; Phleger, C.F.; Guest, M.A.; Nichols, P.D. Lipid, sterols and fatty acids of abyssal polychaetes, crustaceans, and a cnidarian from the northeast Pacific Ocean: Food web implications. Mar. Ecol. Prog. Ser. 2008, 372, 157–167. [Google Scholar] [CrossRef] [Green Version]
- Wirsen, C.O.; Jannasch, H.W.; Wakeham, S.G.; Canuel, E.A. Membrane Lipids of a Psychrophilic and Barophilic Deep-sea Bacterium. Curr. Microbiol. 1987, 14, 319–322. [Google Scholar] [CrossRef]
- Fang, J.; Chan, O.; Kato, C.; Sato, T.; Peeples, T.; Niggemeyer, K. Phospholipid FA of Piezophilic Bacteria from the Deep Sea. Lipids 2003, 38, 885–887. [Google Scholar] [CrossRef]
- De Long, E.F.; Yayanos, A.A. Biochemical function and ecological significance of novel bacterial lipids in deep-sea procaryotes. Appl. Environ. Microbiol. 1986, 51, 730–737. [Google Scholar] [CrossRef] [Green Version]
- Santos, V.; Billett, D.S.M.; Rice, A.L.; Wolff, G.A. Organic matter in deep-sea sediments from the Porcupine Abyssal Plain in the north-east Atlantic Ocean. I-Lipids. Deep-Sea Res. I 1994, 41, 787–819. [Google Scholar] [CrossRef]
- Guezennec, J.; Fiala-Medioni, A. Bacterial abundance and diversity in the Barbados Trench determined by phospholipid analysis. FEMS Microbiol. Ecol. 1996, 19, 83–93. [Google Scholar] [CrossRef]
- Baird, B.H.; White, D.C. Biomass and community structure of the abyssal microbiota determined from the ester-linked phospholipids recovered from Venezuela Basin and Puerto Rico Trench sediments. Mar. Geol. 1985, 68, 217–231. [Google Scholar] [CrossRef]
- Mills, C.T.; Dias, R.F.; Graham, D.; Mandernack, K.W. Determination of phospholipid fatty acid structures and stable carbon isotope compositions of deep-sea sediments of the Northwest Pacific, ODP site 1179. Mar. Chem. 2006, 98, 197–209. [Google Scholar] [CrossRef]
- Lipps, J.H.; Valentine, J.W. The role of Foraminifera in the trophic structure of marine communities. Lethaia 1970, 3, 279–286. [Google Scholar] [CrossRef]
- Zhukova, N.V.; Kharlamenko, V.I. Sources of essential fatty acids in the marine microbial loop. Aquat. Microb. Ecol. 1999, 17, 153–157. [Google Scholar] [CrossRef] [Green Version]
- Desvilettes, C.; Bec, A. Formation and Transfer of Fatty Acids in Aquatic Microbial Food Webs: Role of Heterotrophic Protists. In Lipids in Aquatic Ecosystems; Kainz, M., Brett, M.T., Arts, M.T., Eds.; Springer: New York, NY, USA, 2009; pp. 25–42. [Google Scholar]
- Veloza, A.J.; Chu, F.-L.E.; Tang, K.W. Trophic modification of essential fatty acids by heterotrophic protists and its effects on the fatty acid composition of the copepod Acartia tonsa. Mar. Biol. 2006, 148, 779–788. [Google Scholar] [CrossRef]
- Lambshead, P.J.D. Recent developments in marine benthic biodiversity research. Oceanis 1993, 19, 5–24. [Google Scholar]
- Gems, D. Longevity and ageing in parasitic and free-living nematodes. Biogerontology 2000, 1, 289–307. [Google Scholar] [CrossRef]


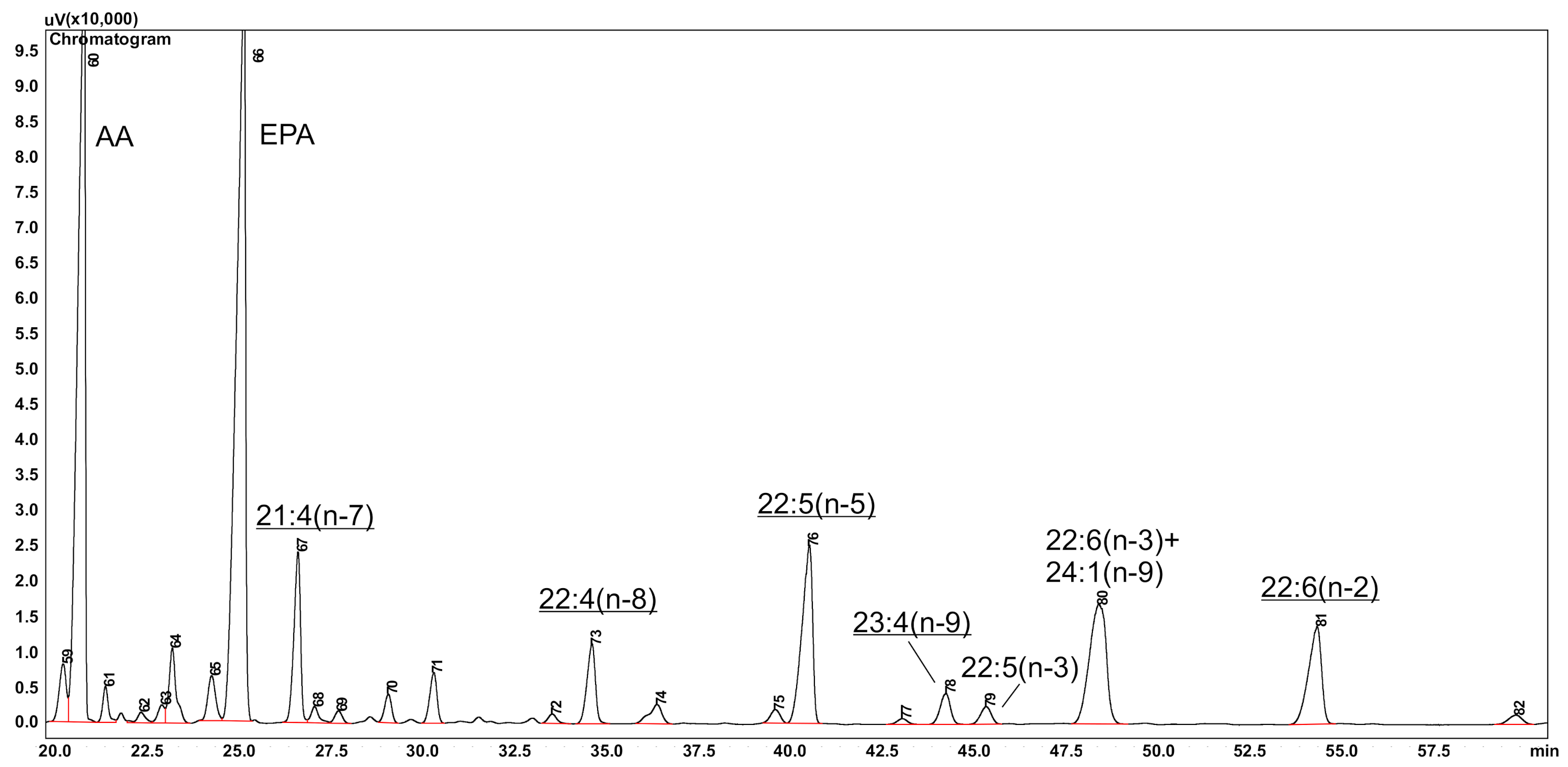{kind=link}
| FA | % | Species | Place and Depth | References |
|---|---|---|---|---|
| Foraminifera | ||||
| 21:4(n-7) | 2.4–4.8% | Bathysiphon lanosum, B. major Rhabdammina abyssorum, Rh. Parabyssorum | Kuril Basin, Sea of Okhotsk, 3307–3386 m | [20] |
| 22:4(n-8) | 1.2–2.0% | |||
| 23:4(n-9) | 1.7–2.3% | |||
| ∆4-i-16:1 | 0.6–1.1% | |||
| ∆7-i-21:1 | 0.4% | |||
| ∆7-ai-21:1 | 0.3–0.8% | |||
| ∆4,11–18:2 | 0.3–0.9% | |||
| ∆4,7–20:2 | ||||
| ∆7,12–20:2 | 0.4–0.6% | |||
| ∆4,7–21:2 | 0.6% | |||
| ∆7,12–22:2 | 6.4–2.1% | |||
| 20:3(n-5) | 1.2% | Reophax nodulosus | Kuril Basin, Sea of Okhotsk, and the adjacent abyssal area of the Pacific Ocean, including the slope of the Kuril-Kamchatka Trench, 3500 m | [21] |
| 22:3(n-5) | 2.4% | |||
| 22:5(n-5) | 18.3% | |||
| 23:4(n-7) | 0.4% | Pyrgo sp. | ||
| 24:4(n-8) | 0.4% | |||
| 24:5(n-9) | 1.7% | |||
| 26:4(n-6) | 0.9% | |||
| 26:5(n-3) | 0.9% | |||
| Echinodermata | ||||
| 22:6(n-2) | 1.6–0.33% | sea urchin Eremicaster vicinus sea star Kamptosoma abyssale | Kuril–Kamchatka Trench, 5200 and 6300 m | [22] |
| 22:5(n-5) | 2.8% | sea star Eremicaster sp. | [50] | |
| 24:6(n-3) | 20.5% | ophiuran Ophiopenia vicina | ||
| 26:7(n-3) | 14.5% | ophiuran Amphiophiura ponderosa | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Svetashev, V.I. Investigation of Deep-Sea Ecosystems Using Marker Fatty Acids: Sources of Essential Polyunsaturated Fatty Acids in Abyssal Megafauna. Mar. Drugs 2022, 20, 17. https://doi.org/10.3390/md20010017
Svetashev VI. Investigation of Deep-Sea Ecosystems Using Marker Fatty Acids: Sources of Essential Polyunsaturated Fatty Acids in Abyssal Megafauna. Marine Drugs. 2022; 20(1):17. https://doi.org/10.3390/md20010017
Chicago/Turabian StyleSvetashev, Vasily I. 2022. "Investigation of Deep-Sea Ecosystems Using Marker Fatty Acids: Sources of Essential Polyunsaturated Fatty Acids in Abyssal Megafauna" Marine Drugs 20, no. 1: 17. https://doi.org/10.3390/md20010017
APA StyleSvetashev, V. I. (2022). Investigation of Deep-Sea Ecosystems Using Marker Fatty Acids: Sources of Essential Polyunsaturated Fatty Acids in Abyssal Megafauna. Marine Drugs, 20(1), 17. https://doi.org/10.3390/md20010017