
Marine Neurotoxins’ Effects on Environmental and Human Health: An OMICS Overview


Abstract
:1. Introduction
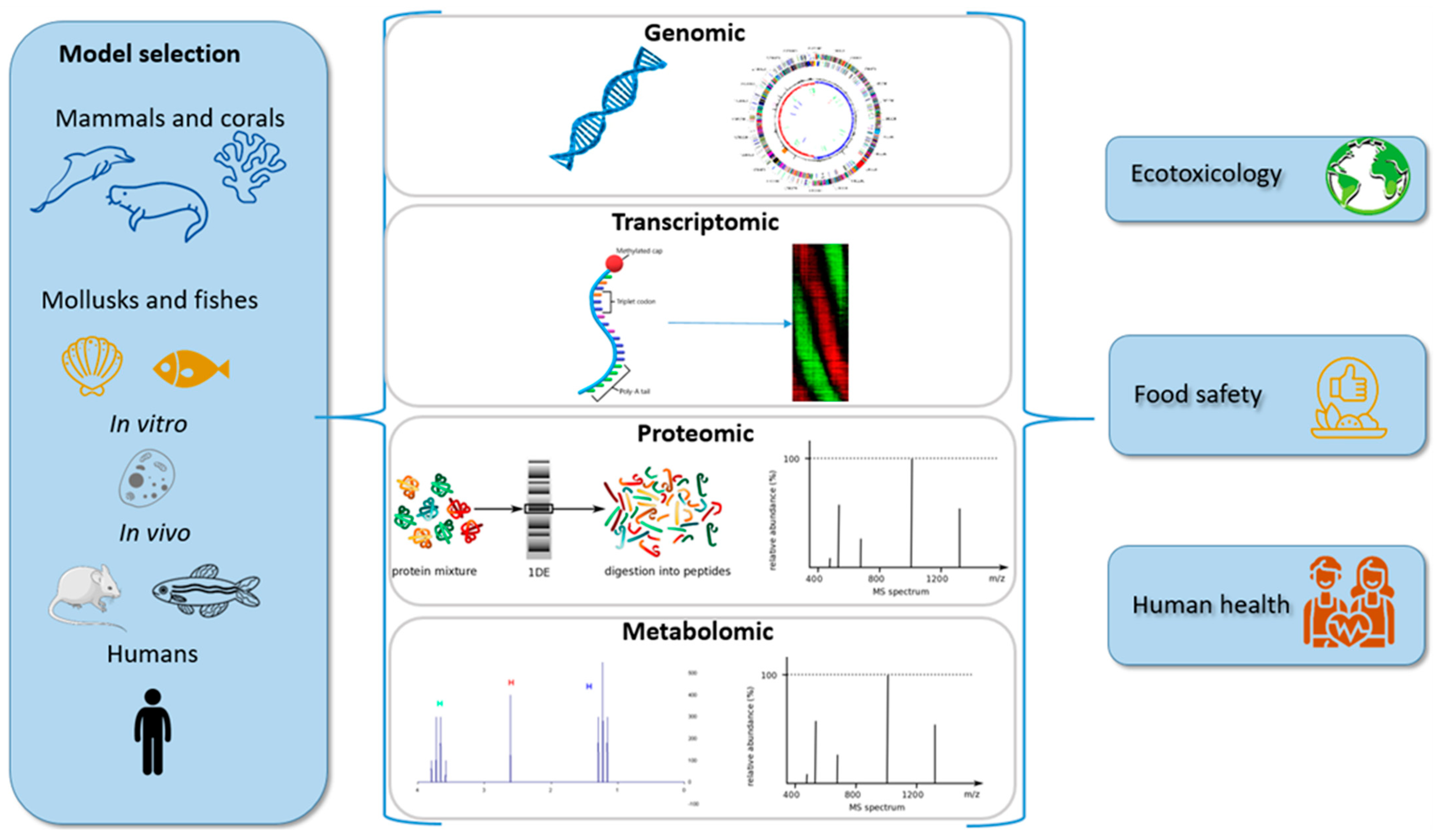
2. OMICS Overview
2.1. Transcriptomics
2.2. Proteomics
2.3. Metabolomics
3. Paralytic Shellfish Poisoning (PSP)
3.1. Food Safety
3.2. In Vitro Studies
3.3. In Vivo Studies
4. Amnesic Shellfish Poisoning (ASP)
4.1. Ecotoxicology
4.2. Food Safety
4.3. In Vivo Studies
5. Neurotoxic Shellfish Poisoning (NSP)
5.1. Ecotoxicology
5.2. In Vivo Studies
6. Ciguatera Fish Poisoning (CFP)
6.1. In Vitro Studies
6.2. In Vivo Studies
6.3. Human Toxicity
7. β-N-Methylamino-L-alanine (BMAA)
7.1. In Vitro Studies
7.2. In Vivo Studies
8. Tetrodotoxin (TTX) Poisoning
8.1. Ecotoxicology
8.2. Food Safety
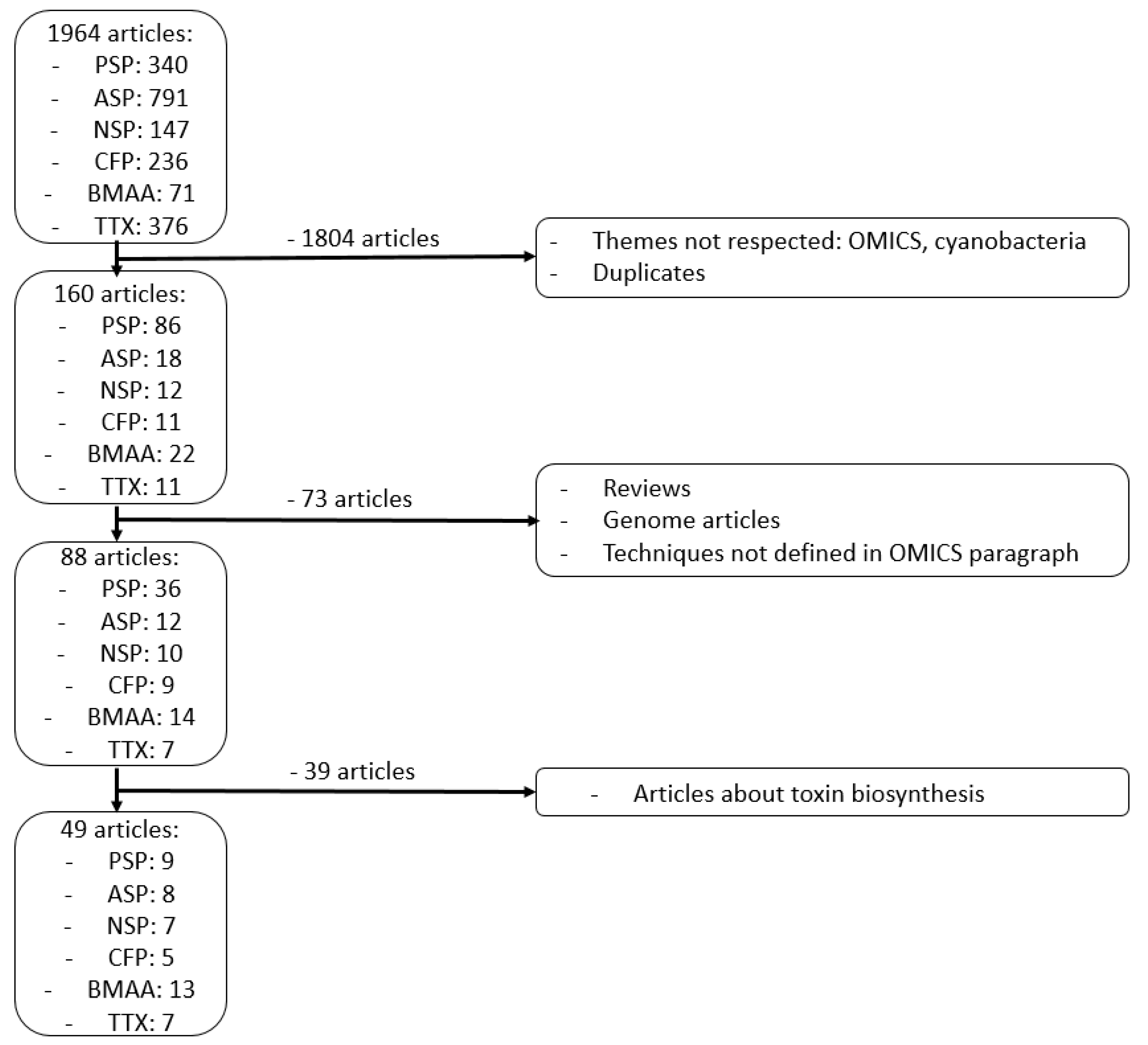
9. Methods
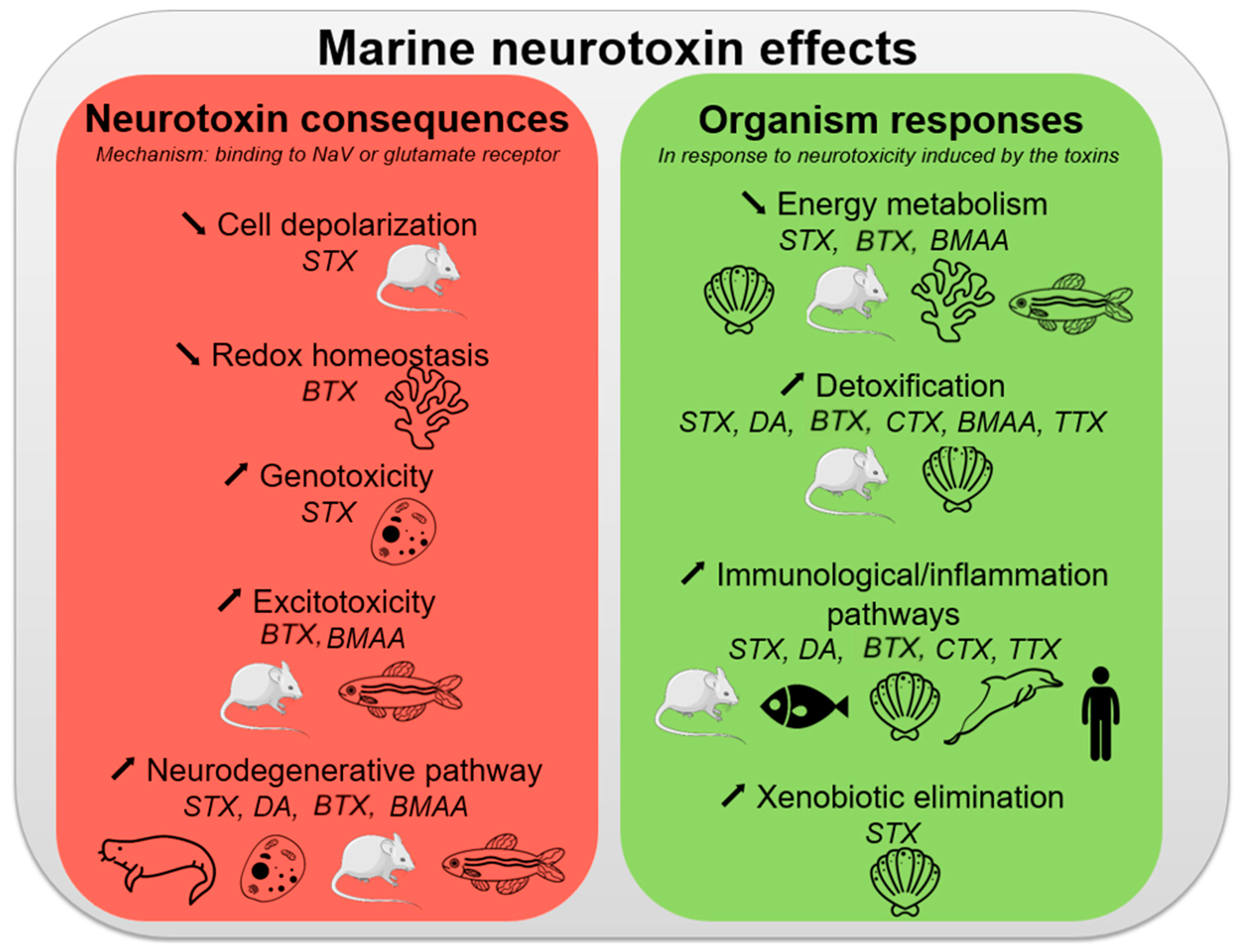
10. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Vilariño, N.; Louzao, M.C.; Abal, P.; Cagide, E.; Carrera, C.; Vieytes, M.R.; Botana, L.M. Human Poisoning from Marine Toxins: Unknowns for Optimal Consumer Protection. Toxins 2018, 10, 324. [Google Scholar] [CrossRef] [Green Version]
- Young, N.; Sharpe, R.A.; Barciela, R.; Nichols, G.; Davidson, K.; Berdalet, E.; Fleming, L.E. Marine Harmful Algal Blooms and Human Health: A Systematic Scoping Review. Harmful Algae 2020, 98, 101901. [Google Scholar] [CrossRef] [PubMed]
- Delcourt, N.; Lagrange, E.; Abadie, E.; Fessard, V.; Frémy, J.-M.; Vernoux, J.-P.; Peyrat, M.-B.; Maignien, T.; Arnich, N.; Molgó, J.; et al. Pinnatoxins’ Deleterious Effects on Cholinergic Networks: From Experimental Models to Human Health. Mar. Drugs 2019, 17, 425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banack, S.A.; Cox, P.A. Biomagnification of Cycad Neurotoxins in Flying Foxes: Implications for ALS-PDC in Guam. Neurology 2003, 61, 387–389. [Google Scholar] [CrossRef] [PubMed]
- Delcourt, N.; Claudepierre, T.; Maignien, T.; Arnich, N.; Mattei, C. Cellular and Molecular Aspects of the β-N-Methylamino-l-Alanine (BMAA) Mode of Action within the Neurodegenerative Pathway: Facts and Controversy. Toxins 2017, 10, 6. [Google Scholar] [CrossRef] [Green Version]
- Campos, A.; Freitas, M.; de Almeida, A.M.; Martins, J.C.; Domínguez-Pérez, D.; Osório, H.; Vasconcelos, V.; Reis Costa, P. OMICs Approaches in Diarrhetic Shellfish Toxins Research. Toxins 2020, 12, 493. [Google Scholar] [CrossRef] [PubMed]
- Lauritano, C.; Ferrante, M.I.; Rogato, A. Marine Natural Products from Microalgae: An -Omics Overview. Mar. Drugs 2019, 17, 269. [Google Scholar] [CrossRef] [Green Version]
- Otto, C.; Reiche, K.; Hackermüller, J. Detection of Differentially Expressed Segments in Tiling Array Data. Bioinformatics 2012, 28, 1471–1479. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A Revolutionary Tool for Transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Roncalli, V.; Cieslak, M.C.; Lenz, P.H. Transcriptomic Responses of the Calanoid Copepod Calanus Finmarchicus to the Saxitoxin Producing Dinoflagellate Alexandrium Fundyense. Sci. Rep. 2016, 6, 25708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Detree, C.; Núñez-Acuña, G.; Roberts, S.; Gallardo-Escárate, C. Uncovering the Complex Transcriptome Response of Mytilus Chilensis against Saxitoxin: Implications of Harmful Algal Blooms on Mussel Populations. PLoS ONE 2016, 11, e0165231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Astuya, A.; Carrera, C.; Ulloa, V.; Aballay, A.; Núñez-Acuña, G.; Hégaret, H.; Gallardo-Escárate, C. Saxitoxin Modulates Immunological Parameters and Gene Transcription in Mytilus Chilensis Hemocytes. Int. J. Mol. Sci. 2015, 16, 15235–15250. [Google Scholar] [CrossRef] [Green Version]
- Gerdol, M.; De Moro, G.; Manfrin, C.; Milandri, A.; Riccardi, E.; Beran, A.; Venier, P.; Pallavicini, A. RNA Sequencing and de Novo Assembly of the Digestive Gland Transcriptome in Mytilus Galloprovincialis Fed with Toxinogenic and Non-Toxic Strains of Alexandrium Minutum. BMC Res. Notes 2014, 7, 722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hlaing, S.M.M.; Lou, J.; Cheng, J.; Xun, X.; Li, M.; Lu, W.; Hu, X.; Bao, Z. Tissue-Biased and Species-Specific Regulation of Glutathione Peroxidase (GPx) Genes in Scallops Exposed to Toxic Dinoflagellates. Toxins 2020, 13, 21. [Google Scholar] [CrossRef]
- Lian, S.; Zhao, L.; Xun, X.; Lou, J.; Li, M.; Li, X.; Wang, S.; Zhang, L.; Hu, X.; Bao, Z. Genome-Wide Identification and Characterization of SODs in Zhikong Scallop Reveals Gene Expansion and Regulation Divergence after Toxic Dinoflagellate Exposure. Mar. Drugs 2019, 17, 700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xun, X.; Cheng, J.; Wang, J.; Li, Y.; Li, X.; Li, M.; Lou, J.; Kong, Y.; Bao, Z.; Hu, X. Solute Carriers in Scallop Genome: Gene Expansion and Expression Regulation after Exposure to Toxic Dinoflagellate. Chemosphere 2020, 241, 124968. [Google Scholar] [CrossRef]
- Lefebvre, K.A.; Tilton, S.C.; Bammler, T.K.; Beyer, R.P.; Srinouanprachan, S.; Stapleton, P.L.; Farin, F.M.; Gallagher, E.P. Gene Expression Profiles in Zebrafish Brain after Acute Exposure to Domoic Acid at Symptomatic and Asymptomatic Doses. Toxicol. Sci. 2009, 107, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.-H.; Kokame, K.; Tsukamoto, Y.; Yutani, C.; Kato, H.; Miyata, T. Characterization of the Human NDRG Gene Family: A Newly Identified Member, NDRG4, Is Specifically Expressed in Brain and Heart. Genomics 2001, 73, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Mancia, A.; Ryan, J.C.; Chapman, R.W.; Wu, Q.; Warr, G.W.; Gulland, F.M.D.; Van Dolah, F.M. Health Status, Infection and Disease in California Sea Lions (Zalophus Californianus) Studied Using a Canine Microarray Platform and Machine-Learning Approaches. Dev. Comp. Immunol. 2012, 36, 629–637. [Google Scholar] [CrossRef] [Green Version]
- Pazos, A.J.; Ventoso, P.; Martínez-Escauriaza, R.; Pérez-Parallé, M.L.; Blanco, J.; Triviño, J.C.; Sánchez, J.L. Transcriptional Response after Exposure to Domoic Acid-Producing Pseudo-Nitzschia in the Digestive Gland of the Mussel Mytilus Galloprovincialis. Toxicon 2017, 140, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Ventoso, P.; Pazos, A.J.; Pérez-Parallé, M.L.; Blanco, J.; Triviño, J.C.; Sánchez, J.L. RNA-Seq Transcriptome Profiling of the Queen Scallop (Aequipecten Opercularis) Digestive Gland after Exposure to Domoic Acid-Producing Pseudo-Nitzschia. Toxins 2019, 11, 97. [Google Scholar] [CrossRef] [Green Version]
- Ventoso, P.; Pazos, A.J.; Blanco, J.; Pérez-Parallé, M.L.; Triviño, J.C.; Sánchez, J.L. Transcriptional Response in the Digestive Gland of the King Scallop (Pecten Maximus) After the Injection of Domoic Acid. Toxins 2021, 13, 339. [Google Scholar] [CrossRef]
- Lazensky, R.; Hunter, M.E.; Moraga Amador, D.; Al-Khedery, B.; Yu, F.; Walsh, C.; Gitzendanner, M.A.; Tripp, K.; Walsh, M.T.; Denslow, N.D. Investigating the Gene Expression Profiles of Rehabilitated Florida Manatees (Trichechus Manatus Latirostris) Following Red Tide Exposure. PLoS ONE 2020, 15, e0234150. [Google Scholar] [CrossRef]
- Cammen, K.M.; Schultz, T.F.; Rosel, P.E.; Wells, R.S.; Read, A.J. Genomewide Investigation of Adaptation to Harmful Algal Blooms in Common Bottlenose Dolphins (Tursiops Truncatus). Mol. Ecol. 2015, 24, 4697–4710. [Google Scholar] [CrossRef]
- Walsh, P.J.; Bookman, R.J.; Zaias, J.; Mayer, G.D.; Abraham, W.; Bourdelais, A.J.; Baden, D.G. Toxicogenomic Effects of Marine Brevetoxins in Liver and Brain of Mouse. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2003, 136, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Rubiolo, J.A.; Vale, C.; Boente-Juncal, A.; Hirama, M.; Yamashita, S.; Camiña, M.; Vieytes, M.R.; Botana, L.M. Transcriptomic Analysis of Ciguatoxin-Induced Changes in Gene Expression in Primary Cultures of Mice Cortical Neurons. Toxins 2018, 10, 192. [Google Scholar] [CrossRef] [Green Version]
- Ryan, J.C.; Morey, J.S.; Bottein, M.-Y.D.; Ramsdell, J.S.; Van Dolah, F.M. Gene Expression Profiling in Brain of Mice Exposed to the Marine Neurotoxin Ciguatoxin Reveals an Acute Anti-Inflammatory, Neuroprotective Response. BMC Neurosci. 2010, 11, 107. [Google Scholar] [CrossRef] [Green Version]
- Ryan, J.C.; Bottein Dechraoui, M.-Y.; Morey, J.S.; Rezvani, A.; Levin, E.D.; Gordon, C.J.; Ramsdell, J.S.; Van Dolah, F.M. Transcriptional Profiling of Whole Blood and Serum Protein Analysis of Mice Exposed to the Neurotoxin Pacific Ciguatoxin-1. Neurotoxicology 2007, 28, 1099–1109. [Google Scholar] [CrossRef] [PubMed]
- Morey, J.S.; Ryan, J.C.; Bottein Dechraoui, M.-Y.; Rezvani, A.H.; Levin, E.D.; Gordon, C.J.; Ramsdell, J.S.; Van Dolah, F.M. Liver Genomic Responses to Ciguatoxin: Evidence for Activation of Phase I and Phase II Detoxification Pathways Following an Acute Hypothermic Response in Mice. Toxicol. Sci. 2008, 103, 298–310. [Google Scholar] [CrossRef] [Green Version]
- Ryan, J.C.; Wu, Q.; Shoemaker, R.C. Transcriptomic Signatures in Whole Blood of Patients Who Acquire a Chronic Inflammatory Response Syndrome (CIRS) Following an Exposure to the Marine Toxin Ciguatoxin. BMC Med. Genom. 2015, 8, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, A.S.; Braidy, N.; Marçal, H.; Welch, J.H.; Gehringer, M.M.; Guillemin, G.J.; Neilan, B.A. Global Cellular Responses to β-Methyl-Amino-L-Alanine (BMAA) by Olfactory Ensheathing Glial Cells (OEC). Toxicon 2015, 99, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Whitelaw, B.L.; Cooke, I.R.; Finn, J.; da Fonseca, R.R.; Ritschard, E.A.; Gilbert, M.T.P.; Simakov, O.; Strugnell, J.M. Adaptive Venom Evolution and Toxicity in Octopods Is Driven by Extensive Novel Gene Formation, Expansion, and Loss. GigaScience 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Ishizaki, S.; Nagashima, Y. Differential Gene Expression Profile in the Liver of the Marine Puffer Fish Takifugu Rubripes Induced by Intramuscular Administration of Tetrodotoxin. Toxicon 2011, 57, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Feroudj, H.; Matsumoto, T.; Kurosu, Y.; Kaneko, G.; Ushio, H.; Suzuki, K.; Kondo, H.; Hirono, I.; Nagashima, Y.; Akimoto, S.; et al. DNA Microarray Analysis on Gene Candidates Possibly Related to Tetrodotoxin Accumulation in Pufferfish. Toxicon 2014, 77, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Feroudj, H.; Kikuchi, R.; Kawana, Y.; Kondo, H.; Hirono, I.; Mochizuki, T.; Nagashima, Y.; Kaneko, G.; Ushio, H.; et al. DNA Microarray Analysis on the Genes Differentially Expressed in the Liver of the Pufferfish, Takifugu Rubripes, Following an Intramuscular Administration of Tetrodotoxin. Microarrays 2014, 3, 226–244. [Google Scholar] [CrossRef]
- Zou, S. Comparative Transcriptome Analysis of Toxic and Non-Toxic Nassarius Communities and Identification of Genes Involved in TTX-Adaptation. Toxins 2020, 12, 761. [Google Scholar] [CrossRef]
- Yıldırım, Y.; Anderson, M.J.; Hansson, B.; Patel, S.; Millar, C.D.; Rainey, P.B. Genetic Structure of the Grey Side-Gilled Sea Slug (Pleurobranchaea Maculata) in Coastal Waters of New Zealand. PLoS ONE 2018, 13, e0202197. [Google Scholar] [CrossRef] [Green Version]
- Mann, M. Origins of Mass Spectrometry-Based Proteomics. Nat. Rev. Mol. Cell Biol. 2016, 17, 678. [Google Scholar] [CrossRef] [PubMed]
- Müller, J.B.; Geyer, P.E.; Colaço, A.R.; Treit, P.V.; Strauss, M.T.; Oroshi, M.; Doll, S.; Virreira Winter, S.; Bader, J.M.; Köhler, N.; et al. The Proteome Landscape of the Kingdoms of Life. Nature 2020, 582, 592–596. [Google Scholar] [CrossRef]
- Ünlü, M.; Morgan, M.E.; Minden, J.S. Difference Gel Electrophoresis. A Single Gel Method for Detecting Changes in Protein Extracts. Electrophoresis 1997, 18, 2071–2077. [Google Scholar] [CrossRef]
- Chen, X.; Sun, Y.; Huang, H.; Liu, W.; Hu, P.; Huang, X.; Zou, F.; Liu, J. Uncovering the Proteome Response of Murine Neuroblastoma Cells against Low-Dose Exposure to Saxitoxin. Toxicol. Mech. Methods 2018, 28, 335–344. [Google Scholar] [CrossRef]
- Neely, B.A.; Ferrante, J.A.; Chaves, J.M.; Soper, J.L.; Almeida, J.S.; Arthur, J.M.; Gulland, F.M.D.; Janech, M.G. Proteomic Analysis of Plasma from California Sea Lions (Zalophus Californianus) Reveals Apolipoprotein E as a Candidate Biomarker of Chronic Domoic Acid Toxicosis. PLoS ONE 2014, 10, e0123295. [Google Scholar] [CrossRef] [Green Version]
- Tian, L.; Wang, M.; Li, X.; Lam, P.K.S.; Wang, M.; Wang, D.; Chou, H.N.; Li, Y.; Chan, L.L. Proteomic Modification in Gills and Brains of Medaka Fish (Oryzias Melastigma) after Exposure to a Sodium Channel Activator Neurotoxin, Brevetoxin-1. Aquat. Toxicol. Amst. Neth. 2011, 104, 211–217. [Google Scholar] [CrossRef]
- Moulder, R.; Bhosale, S.D.; Goodlett, D.R.; Lahesmaa, R. Analysis of the plasma proteome using iTRAQ and TMT-based Isobaric labeling. Mass Spectrom. Rev. 2018, 37, 583–606. [Google Scholar] [CrossRef]
- Reynolds, D.A.; Yoo, M.-J.; Dixson, D.L.; Ross, C. Exposure to the Florida Red Tide Dinoflagellate, Karenia Brevis, and Its Associated Brevetoxins Induces Ecophysiological and Proteomic Alterations in Porites Astreoides. PLoS ONE 2020, 15, e0228414. [Google Scholar] [CrossRef] [Green Version]
- Ong, S.-E.; Blagoev, B.; Kratchmarova, I.; Kristensen, D.B.; Steen, H.; Pandey, A.; Mann, M. Stable Isotope Labeling by Amino Acids in Cell Culture, SILAC, as a Simple and Accurate Approach to Expression Proteomics. Mol. Cell. Proteomics 2002, 1, 376–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beri, J.; Nash, T.; Martin, R.M.; Bereman, M.S. Exposure to BMAA Mirrors Molecular Processes Linked to Neurodegenerative Disease. Proteomics 2017, 17. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Chen, X.; Liu, W.; Li, S.; Zhou, Y.; Yang, X.; Liu, J. Effects of Long-Term Low Dose Saxitoxin Exposure on Nerve Damage in Mice. Aging 2021, 13, 17211–17226. [Google Scholar] [CrossRef]
- Doerr, S.; Harvey, M.J.; Noé, F.; Fabritiis, G.D. HTMD: High-throughput molecular dynamics for molecular discovery. J. Chem. Theory Comput. 2016, 12, 1845–1852. [Google Scholar] [CrossRef]
- Neely, B.A.; Soper, J.L.; Gulland, F.M.D.; Bell, P.D.; Kindy, M.; Arthur, J.M.; Janech, M.G. Proteomic Analysis of Cerebrospinal Fluid in California Sea Lions (Zalophus Californianus) with Domoic Acid Toxicosis Identifies Proteins Associated with Neurodegeneration. Proteomics 2015, 15, 4051–4063. [Google Scholar] [CrossRef]
- Glover, W.B.; Mash, D.C.; Murch, S.J. The Natural Non-Protein Amino Acid N-β-Methylamino-L-Alanine (BMAA) Is Incorporated into Protein during Synthesis. Amino Acids 2014, 46, 2553–2559. [Google Scholar] [CrossRef] [PubMed]
- Nauser, T.; Steinmann, D.; Koppenol, W.H. Why Do Proteins Use Selenocysteine Instead of Cysteine? Amino Acids 2012, 42, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.M.; Stallrich, J.; Bereman, M.S. Mixture Designs to Investigate Adverse Effects upon Co-Exposure to Environmental Cyanotoxins. Toxicology 2019, 421, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, O.; Kultima, K.; Wadensten, H.; Nilsson, A.; Roman, E.; Andrén, P.E.; Brittebo, E.B. Neurotoxin-Induced Neuropeptide Perturbations in Striatum of Neonatal Rats. J. Proteome Res. 2013, 12, 1678–1690. [Google Scholar] [CrossRef]
- Karlsson, O.; Bergquist, J.; Andersson, M. Quality Measures of Imaging Mass Spectrometry Aids in Revealing Long-Term Striatal Protein Changes Induced by Neonatal Exposure to the Cyanobacterial Toxin β-N-Methylamino-L-Alanine (BMAA). Mol. Cell. Proteomics MCP 2014, 13, 93–104. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, O.; Berg, A.-L.; Hanrieder, J.; Arnerup, G.; Lindström, A.-K.; Brittebo, E.B. Intracellular Fibril Formation, Calcification, and Enrichment of Chaperones, Cytoskeletal, and Intermediate Filament Proteins in the Adult Hippocampus CA1 Following Neonatal Exposure to the Nonprotein Amino Acid BMAA. Arch. Toxicol. 2015, 89, 423–436. [Google Scholar] [CrossRef]
- Whitelaw, B.L.; Strugnell, J.M.; Faou, P.; da Fonseca, R.R.; Hall, N.E.; Norman, M.; Finn, J.; Cooke, I.R. Combined Transcriptomic and Proteomic Analysis of the Posterior Salivary Gland from the Southern Blue-Ringed Octopus and the Southern Sand Octopus. J. Proteome Res. 2016, 15, 3284–3297. [Google Scholar] [CrossRef]
- Fernie, A.R.; Trethewey, R.N.; Krotzky, A.J.; Willmitzer, L. Metabolite Profiling: From Diagnostics to Systems Biology. Nat. Rev. Mol. Cell Biol. 2004, 5, 763–769. [Google Scholar] [CrossRef]
- Wishart, D.S.; Tzur, D.; Knox, C.; Eisner, R.; Guo, A.C.; Young, N.; Cheng, D.; Jewell, K.; Arndt, D.; Sawhney, S.; et al. HMDB: The Human Metabolome Database. Nucleic Acids Res. 2007, 35, D521–D526. [Google Scholar] [CrossRef]
- Beckonert, O.; Keun, H.C.; Ebbels, T.M.D.; Bundy, J.; Holmes, E.; Lindon, J.C.; Nicholson, J.K. Metabolic Profiling, Metabolomic and Metabonomic Procedures for NMR Spectroscopy of Urine, Plasma, Serum and Tissue Extracts. Nat. Protoc. 2007, 2, 2692–2703. [Google Scholar] [CrossRef]
- Chan, E.C.Y.; Pasikanti, K.K.; Nicholson, J.K. Global Urinary Metabolic Profiling Procedures Using Gas Chromatography–Mass Spectrometry. Nat. Protoc. 2011, 6, 1483–1499. [Google Scholar] [CrossRef]
- Wishart, D.S. NMR Metabolomics: A Look Ahead. J. Magn. Reson. 2019, 306, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Yau, M.-S.; Lei, E.N.-Y.; Ng, I.H.-M.; Yuen, C.K.-K.; Lam, J.C.-W.; Lam, M.H.-W. Changes in the Neurotransmitter Profile in the Central Nervous System of Marine Medaka (Oryzias Melastigma) after Exposure to Brevetoxin PbTx-1-A Multivariate Approach to Establish Exposure Biomarkers. Sci. Total Environ. 2019, 673, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Annunziato, M.; Eeza, M.N.H.; Bashirova, N.; Lawson, A.; Matysik, J.; Benetti, D.; Grosell, M.; Stieglitz, J.D.; Alia, A.; Berry, J.P. An Integrated Systems-Level Model of the Toxicity of Brevetoxin Based on High-Resolution Magic-Angle Spinning Nuclear Magnetic Resonance (HRMAS NMR) Metabolic Profiling of Zebrafish Embryos. Sci. Total Environ. 2022, 803, 149858. [Google Scholar] [CrossRef]
- Engskog, M.K.R.; Ersson, L.; Haglöf, J.; Arvidsson, T.; Pettersson, C.; Brittebo, E. β-N-Methylamino-L-Alanine (BMAA) Perturbs Alanine, Aspartate and Glutamate Metabolism Pathways in Human Neuroblastoma Cells as Determined by Metabolic Profiling. Amino Acids 2017, 49, 905–919. [Google Scholar] [CrossRef]
- Engskog, M.K.R.; Karlsson, O.; Haglöf, J.; Elmsjö, A.; Brittebo, E.; Arvidsson, T.; Pettersson, C. The Cyanobacterial Amino Acid β-N-Methylamino-l-Alanine Perturbs the Intermediary Metabolism in Neonatal Rats. Toxicology 2013, 312, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Veenstra, T.D. Omics in Systems Biology: Current Progress and Future Outlook. Proteomics 2021, 21, 2000235. [Google Scholar] [CrossRef] [PubMed]
- Roy, U.; Conklin, L.; Schiller, J.; Matysik, J.; Berry, J.P.; Alia, A. Metabolic Profiling of Zebrafish (Danio Rerio) Embryos by NMR Spectroscopy Reveals Multifaceted Toxicity of β-Methylamino-L-Alanine (BMAA). Sci. Rep. 2017, 7, 17305. [Google Scholar] [CrossRef] [Green Version]
- Waidyanatha, S.; Ryan, K.; Sanders, J.M.; McDonald, J.D.; Wegerski, C.J.; Doyle-Eisle, M.; Garner, C.E. Disposition of β-N-Methylamino-l-Alanine (L-BMAA), a Neurotoxin, in Rodents Following a Single or Repeated Oral Exposure. Toxicol. Appl. Pharmacol. 2018, 339, 151–160. [Google Scholar] [CrossRef]
- Farabegoli, F.; Blanco, L.; Rodríguez, L.P.; Vieites, J.M.; Cabado, A.G. Phycotoxins in Marine Shellfish: Origin, Occurrence and Effects on Humans. Mar. Drugs 2018, 16, 188. [Google Scholar] [CrossRef] [Green Version]
- Hallegraeff, G.M. A Review of Harmful Algal Blooms and Their Apparent Global Increase. Phycologia 1993, 32, 79–99. [Google Scholar] [CrossRef] [Green Version]
- Samson, J.C.; Shumway, S.E.; Weis, J.S. Effects of the Toxic Dinoflagellate, Alexandrium Fundyense on Three Species of Larval Fish: A Food-Chain Approach. J. Fish Biol. 2008, 72, 168–188. [Google Scholar] [CrossRef]
- Shumway, S.E.; Burkholder, J.M.; Springer, J. Effects of the Estuarine Dinoflagellate Pfiesteria Shumwayae (Dinophyceae) on Survival and Grazing Activity of Several Shellfish Species. Harmful Algae 2006, 5, 442–458. [Google Scholar] [CrossRef]
- Wang, C.; Huan, P.; Yue, X.; Yan, M.; Liu, B. Molecular Characterization of a Glutathione Peroxidase Gene and Its Expression in the Selected Vibrio-Resistant Population of the Clam Meretrix Meretrix. Fish Shellfish Immunol. 2011, 30, 1294–1302. [Google Scholar] [CrossRef] [PubMed]
- Cong, M.; Zhang, L.; Zhang, L.; Zhao, J.; Wu, H.; Chen, H.; Kong, J. Molecular Characterization of a Se-Containing Glutathione Peroxidases Gene and Its Expressions to Heavy Metals Compared with Non-Se-Containing Glutathione Peroxidases in Venerupis Philippinarum. Agri Gene 2016, 1, 46–52. [Google Scholar] [CrossRef]
- Estrada, N.; de Jesús Romero, M.; Campa-Córdova, A.; Luna, A.; Ascencio, F. Effects of the Toxic Dinoflagellate, Gymnodinium Catenatum on Hydrolytic and Antioxidant Enzymes, in Tissues of the Giant Lions-Paw Scallop Nodipecten Subnodosus. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2007, 146, 502–510. [Google Scholar] [CrossRef]
- Deeds, J.; Landsberg, J.; Etheridge, S.; Pitcher, G.; Longan, S. Non-Traditional Vectors for Paralytic Shellfish Poisoning. Mar. Drugs 2008, 6, 308–348. [Google Scholar] [CrossRef]
- Li, Y.; Sun, X.; Hu, X.; Xun, X.; Zhang, J.; Guo, X.; Jiao, W.; Zhang, L.; Liu, W.; Wang, J.; et al. Scallop Genome Reveals Molecular Adaptations to Semi-Sessile Life and Neurotoxins. Nat. Commun. 2017, 8, 1721. [Google Scholar] [CrossRef]
- César-Razquin, A.; Snijder, B.; Frappier-Brinton, T.; Isserlin, R.; Gyimesi, G.; Bai, X.; Reithmeier, R.A.; Hepworth, D.; Hediger, M.A.; Edwards, A.M.; et al. A Call for Systematic Research on Solute Carriers. Cell 2015, 162, 478–487. [Google Scholar] [CrossRef] [Green Version]
- Parveen, F.; Bender, D.; Law, S.-H.; Mishra, V.K.; Chen, C.-C.; Ke, L.-Y. Role of Ceramidases in Sphingolipid Metabolism and Human Diseases. Cells 2019, 8, 1573. [Google Scholar] [CrossRef] [Green Version]
- Tammilehto, A.; Nielsen, T.G.; Krock, B.; Møller, E.F.; Lundholm, N. Induction of Domoic Acid Production in the Toxic Diatom Pseudo-Nitzschia Seriata by Calanoid Copepods. Aquat. Toxicol. 2015, 159, 52–61. [Google Scholar] [CrossRef] [Green Version]
- Wells, M.L.; Trainer, V.L.; Smayda, T.J.; Karlson, B.S.O.; Trick, C.G.; Kudela, R.M.; Ishikawa, A.; Bernard, S.; Wulff, A.; Anderson, D.M.; et al. Harmful Algal Blooms and Climate Change: Learning from the Past and Present to Forecast the Future. Harmful Algae 2015, 49, 68–93. [Google Scholar] [CrossRef] [Green Version]
- Petroff, R.; Hendrix, A.; Shum, S.; Grant, K.S.; Lefebvre, K.A.; Burbacher, T.M. Public Health Risks Associated with Chronic, Low-Level Domoic Acid Exposure: A Review of the Evidence. Pharmacol. Ther. 2021, 227, 107865. [Google Scholar] [CrossRef]
- Cook, P.F.; Reichmuth, C.; Rouse, A.A.; Libby, L.A.; Dennison, S.E.; Carmichael, O.T.; Kruse-Elliott, K.T.; Bloom, J.; Singh, B.; Fravel, V.A.; et al. Algal Toxin Impairs Sea Lion Memory and Hippocampal Connectivity, with Implications for Strandings. Science 2015, 350, 1545–1547. [Google Scholar] [CrossRef]
- Beltrán, A.S.; Palafox-Uribe, M.; Grajales-Montiel, J.; Cruz-Villacorta, A.; Ochoa, J.L. Sea Bird Mortality at Cabo San Lucas, Mexico: Evidence That Toxic Diatom Blooms Are Spreading. Toxicon 1997, 35, 447–453. [Google Scholar] [CrossRef]
- Hsiao, K.; Chapman, P.; Nilsen, S.; Eckman, C.; Harigaya, Y.; Younkin, S.; Yang, F.; Cole, G. Correlative memory deficits, Abeta elevation, and amyloid plaques in transgenic mice. Science 1996, 274, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Maina, V.; Cotena, A.; Doni, A.; Nebuloni, M.; Pasqualini, F.; Milner, C.M.; Day, A.J.; Mantovani, A.; Garlanda, C. Coregulation in Human Leukocytes of the Long Pentraxin PTX3 and TSG-6. J. Leukoc. Biol. 2009, 86, 123–132. [Google Scholar] [CrossRef] [Green Version]
- Sando, S.B.; Melquist, S.; Cannon, A.; Hutton, M.L.; Sletvold, O.; Saltvedt, I.; White, L.R.; Lydersen, S.; Aasly, J.O. APOE Epsilon 4 Lowers Age at Onset and Is a High Risk Factor for Alzheimer’s Disease; a Case Control Study from Central Norway. BMC Neurol. 2008, 8, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gulland, F.M.D.; Hall, A.J.; Greig, D.J.; Frame, E.R.; Colegrove, K.M.; Booth, R.K.N.; Wasser, S.K.; Scott-Moncrieff, J.C.R. Evaluation of Circulating Eosinophil Count and Adrenal Gland Function in California Sea Lions Naturally Exposed to Domoic Acid. J. Am. Vet. Med. Assoc. 2012, 241, 943–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finehout, E.J.; Franck, Z.; Lee, K.H. Complement Protein Isoforms in CSF as Possible Biomarkers for Neurodegenerative Disease. Dis. Markers 2005, 21, 93–101. [Google Scholar] [CrossRef] [Green Version]
- Novaczek, I.; Madhyastha, M.S.; Ablett, R.F.; Johnson, G.; Nijjar, M.S.; Sims, D.E. Uptake, Disposition and Depuration of Domoic Acid by Blue Mussels (Mytilus Edulis). Aquat. Toxicol. 1991, 21, 103–118. [Google Scholar] [CrossRef]
- Livingstone, D.R. The Fate of Organic Xenobiotics in Aquatic Ecosystems: Quantitative and Qualitative Differences in Biotransformation by Invertebrates and Fish. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 1998, 120, 43–49. [Google Scholar] [CrossRef]
- Di Dato, V.; Musacchia, F.; Petrosino, G.; Patil, S.; Montresor, M.; Sanges, R.; Ferrante, M.I. Transcriptome Sequencing of Three Pseudo-Nitzschia Species Reveals Comparable Gene Sets and the Presence of Nitric Oxide Synthase Genes in Diatoms. Sci. Rep. 2015, 5, 12329. [Google Scholar] [CrossRef] [PubMed]
- Hiolski, E.M.; Kendrick, P.S.; Frame, E.R.; Myers, M.S.; Bammler, T.K.; Beyer, R.P.; Farin, F.M.; Wilkerson, H.-W.; Smith, D.R.; Marcinek, D.J.; et al. Chronic Low-Level Domoic Acid Exposure Alters Gene Transcription and Impairs Mitochondrial Function in the CNS. Aquat. Toxicol. Amst. Neth. 2014, 155, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Gauthier, J.; Siddiqui, T.J.; Huashan, P.; Yokomaku, D.; Hamdan, F.F.; Champagne, N.; Lapointe, M.; Spiegelman, D.; Noreau, A.; Lafrenière, R.G.; et al. Truncating Mutations in NRXN2 and NRXN1 in Autism Spectrum Disorders and Schizophrenia. Hum. Genet. 2011, 130, 563–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.G.; Morey, J.S.; Neely, M.G.; Ryan, J.C.; Van Dolah, F.M. Transcriptome Remodeling Associated with Chronological Aging in the Dinoflagellate, Karenia Brevis. Mar. Genom. 2012, 5, 15–25. [Google Scholar] [CrossRef]
- Granéli, E.; Turner, J.T. Ecology of Harmful Algae: 15 Tables; Springer: Berlin, Germany, 2006; pp. 1–413. [Google Scholar]
- HAB Monitoring Database, Florida Fish and Wildlife Conservation Commission. Available online: http://myfwc.com/research/redtide/monitoring/database/ (accessed on 29 July 2021).
- Hilderbrand, S.C.; Murrell, R.N.; Gibson, J.E.; Brown, J.M. Marine Brevetoxin Induces IgE-Independent Mast Cell Activation. Arch. Toxicol. 2011, 85, 135–141. [Google Scholar] [CrossRef]
- Ahmed, M.Y.; Chioza, B.A.; Rajab, A.; Schmitz-Abe, K.; Al-Khayat, A.; Al-Turki, S.; Baple, E.L.; Patton, M.A.; Al-Memar, A.Y.; Hurles, M.E.; et al. Loss of PCLO Function Underlies Pontocerebellar Hypoplasia Type III. Neurology 2015, 84, 1745–1750. [Google Scholar] [CrossRef] [Green Version]
- Fenster, S.D.; Garner, C.C. Gene Structure and Genetic Localization of the PCLO Gene Encoding the Presynaptic Active Zone Protein Piccolo. Int. J. Dev. Neurosci. 2002, 20, 161–171. [Google Scholar] [CrossRef]
- van Hooidonk, R.; Maynard, J.A.; Manzello, D.; Planes, S. Opposite Latitudinal Gradients in Projected Ocean Acidification and Bleaching Impacts on Coral Reefs. Glob. Change Biol. 2014, 20, 103–112. [Google Scholar] [CrossRef]
- Puga, A.; Maier, A.; Medvedovic, M. The Transcriptional Signature of Dioxin in Human Hepatoma HepG2 Cells. Biochem. Pharmacol. 2000, 60, 1129–1142. [Google Scholar] [CrossRef]
- Kales, S.C.; Bols, N.C.; Dixon, B. Calreticulin in Rainbow Trout: A Limited Response to Endoplasmic Reticulum (ER) Stress. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2007, 147, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Eijkelkamp, N.; Linley, J.E.; Baker, M.D.; Minett, M.S.; Cregg, R.; Werdehausen, R.; Rugiero, F.; Wood, J.N. Neurological Perspectives on Voltage-Gated Sodium Channels. Brain J. Neurol. 2012, 135, 2585–2612. [Google Scholar] [CrossRef]
- Friedman, M.A.; Fernandez, M.; Backer, L.C.; Dickey, R.W.; Bernstein, J.; Schrank, K.; Kibler, S.; Stephan, W.; Gribble, M.O.; Bienfang, P.; et al. An Updated Review of Ciguatera Fish Poisoning: Clinical, Epidemiological, Environmental, and Public Health Management. Mar. Drugs 2017, 15, 72. [Google Scholar] [CrossRef]
- Pearn, J.H. Chronic Fatigue Syndrome: Chronic Ciguatera Poisoning as a Differential Diagnosis. Med. J. Aust. 1997, 166, 309–310. [Google Scholar] [CrossRef]
- Tamura, Y.; Shintani, M.; Nakamura, A.; Monden, M.; Shiomi, H. Phase-Specific Central Regulatory Systems of Hibernation in Syrian Hamsters. Brain Res. 2005, 1045, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Marone, G.; Granata, F.; Spadaro, G.; Genovese, A.; Triggiani, M. The Histamine-Cytokine Network in Allergic Inflammation. J. Allergy Clin. Immunol. 2003, 112, S83–S88. [Google Scholar] [CrossRef]
- Denham, J.M.; Hill, I.D. Celiac Disease and Autoimmunity: Review and Controversies. Curr. Allergy Asthma Rep. 2013, 13, 347–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Didierlaurent, A.; Brissoni, B.; Velin, D.; Aebi, N.; Tardivel, A.; Käslin, E.; Sirard, J.C.; Angelov, G.; Tschopp, J.; Burns, K. Tollip Regulates Proinflammatory Responses to Interleukin-1 and Lipopolysaccharide. Mol. Cell. Biol. 2006, 26, 735–742. [Google Scholar] [CrossRef] [Green Version]
- Vega, A.; Bell, E.A. α-Amino-β-methylaminopropionic acid, a new amino acid from seeds of Cycas circinalis. Phytochemistry 1967, 6, 759–762. [Google Scholar] [CrossRef]
- Jiang, L.; Eriksson, J.; Lage, S.; Jonasson, S.; Shams, S.; Mehine, M.; Ilag, L.L.; Rasmussen, U. Diatoms: A Novel Source for the Neurotoxin BMAA in Aquatic Environments. PLoS ONE 2014, 9, e84578. [Google Scholar] [CrossRef] [PubMed]
- Lage, S.; Costa, P.R.; Moita, T.; Eriksson, J.; Rasmussen, U.; Rydberg, S.J. BMAA in Shellfish from Two Portuguese Transitional Water Bodies Suggests the Marine Dinoflagellate Gymnodinium Catenatum as a Potential BMAA Source. Aquat. Toxicol. 2014, 152, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Lage, S.; Annadotter, H.; Rasmussen, U.; Rydberg, S. Biotransfer of β-N-Methylamino-l-Alanine (BMAA) in a Eutrophicated Freshwater Lake. Mar. Drugs 2015, 13, 1185–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masseret, E.; Banack, S.; Boumédiène, F.; Abadie, E.; Brient, L.; Pernet, F.; Juntas-Morales, R.; Pageot, N.; Metcalf, J.; Cox, P.; et al. Dietary BMAA Exposure in an Amyotrophic Lateral Sclerosis Cluster from Southern France. PLoS ONE 2013, 8, e83406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenthal, G.A.; Dahlman, D.L. L-Canavanine and Protein Synthesis in the Tobacco Hornworm Manduca Sexta. Proc. Natl. Acad. Sci. USA 1986, 83, 14–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H.; Shi, Y.; Hanson, K.A.; Williams, L.M.; Sakasai, R.; Bowler, M.J.; Tibbetts, R.S. Potentiation of Amyotrophic Lateral Sclerosis (ALS)-Associated TDP-43 Aggregation by the Proteasome-Targeting Factor, Ubiquilin 1. J. Biol. Chem. 2009, 284, 8083–8092. [Google Scholar] [CrossRef] [Green Version]
- Rush, T.; Liu, X.; Lobner, D. Synergistic Toxicity of the Environmental Neurotoxins Methylmercury and β-N-Methylamino-L-Alanine. NeuroReport 2012, 23, 216–219. [Google Scholar] [CrossRef] [PubMed]
- Snyder, S.E.; Pintar, J.E.; Salton, S.R.J. Developmental expression of VGF mRNA in the prenatal and postnatal rat. J. Comp. Neurol. 1998, 394, 64–90. [Google Scholar] [CrossRef]
- Mahata, M.; Mahata, S.K.; Fischer-Colbrie, R.; Winkler, H. Ontogenic Development and Distribution of MRNAs of Chromogranin A and B, Secretogranin II, P65 and Synaptin/Synaptophysin in Rat Brain. Brain Res. Dev. Brain Res. 1993, 76, 43–58. [Google Scholar] [CrossRef]
- Phokeo, V.; Kwiecien, J.M.; Ball, A.K. Characterization of the Optic Nerve and Retinal Ganglion Cell Layer in the Dysmyelinated Adult Long Evans Shaker Rat: Evidence for Axonal Sprouting. J. Comp. Neurol. 2002, 451, 213–224. [Google Scholar] [CrossRef]
- Myhrer, T. Neurotransmitter Systems Involved in Learning and Memory in the Rat: A Meta-Analysis Based on Studies of Four Behavioral Tasks. Brain Res. Rev. 2003, 41, 268–287. [Google Scholar] [CrossRef]
- Frøyset, A.K.; Khan, E.A.; Fladmark, K.E. Quantitative Proteomics Analysis of Zebrafish Exposed to Sub-Lethal Dosages of β-Methyl-Amino-L-Alanine (BMAA). Sci. Rep. 2016, 6, 29631. [Google Scholar] [CrossRef] [Green Version]
- Bilsland, L.G.; Dick, J.R.T.; Pryce, G.; Petrosino, S.; Di Marzo, V.; Baker, D.; Greensmith, L.; Bilsland, L.G.; Dick, J.R.T.; Pryce, G.; et al. Increasing Cannabinoid Levels by Pharmacological and Genetic Manipulation Delays Disease Progression in SOD1 Mice. FASEB J. 2006, 20, 1003–1005. [Google Scholar] [CrossRef] [PubMed]
- Andres, R.H.; Ducray, A.D.; Schlattner, U.; Wallimann, T.; Widmer, H.R. Functions and Effects of Creatine in the Central Nervous System. Brain Res. Bull. 2008, 76, 329–343. [Google Scholar] [CrossRef] [PubMed]
- Chau, R.; Kalaitzis, J.A.; Neilan, B.A. On the Origins and Biosynthesis of Tetrodotoxin. Aquat. Toxicol. 2011, 104, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Hort, V.; Arnich, N.; Guérin, T.; Lavison-Bompard, G.; Nicolas, M. First Detection of Tetrodotoxin in Bivalves and Gastropods from the French Mainland Coasts. Toxins 2020, 12, 599. [Google Scholar] [CrossRef] [PubMed]
- Kemparaju, K.; Girish, K.S. Snake Venom Hyaluronidase: A Therapeutic Target. Cell Biochem. Funct. 2006, 24, 7–12. [Google Scholar] [CrossRef]
- Grisley, M.S.; Boyle, P.R. Chitinase, a New Enzyme in Octopus Saliva. Comp. Biochem. Physiol. Part B Comp. Biochem. 1990, 95, 311–316. [Google Scholar] [CrossRef]
- Jost, M.C.; Hillis, D.M.; Lu, Y.; Kyle, J.W.; Fozzard, H.A.; Zakon, H.H. Toxin-Resistant Sodium Channels: Parallel Adaptive Evolution across a Complete Gene Family. Mol. Biol. Evol. 2008, 25, 1016–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldman, C.R.; Brodie, E.D.; Brodie, E.D.; Pfrender, M.E. Constraint Shapes Convergence in Tetrodotoxin-Resistant Sodium Channels of Snakes. Proc. Natl. Acad. Sci. USA 2012, 109, 4556–4561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Kondo, H.; Sato, S.; Akimoto, S.; Saito, T.; Kodama, M.; Watabe, S. Identification of Novel Genes Related to Tetrodotoxin Intoxication in Pufferfish. Toxicon 2007, 49, 939–953. [Google Scholar] [CrossRef] [PubMed]
- Moshage, H. Cytokines and the Hepatic Acute Phase Response. J. Pathol. 1997, 181, 257–266. [Google Scholar] [CrossRef]
- Isom, L.L.; Ragsdale, D.S.; De Jongh, K.S.; Westenbroek, R.E.; Reber, B.F.; Scheuer, T.; Catterall, W.A. Structure and Function of the p2 Subunit of Brain Sodium Channels, a Transmembrane Glycoprotein with a CAM Motif. Cell 1995, 83, 433–442. [Google Scholar] [CrossRef] [Green Version]
- Malhotra, J.D.; Kazen-Gillespie, K.; Hortsch, M.; Isom, L.L. Sodium Channel β Subunits Mediate Homophilic Cell Adhesion and Recruit Ankyrin to Points of Cell-Cell Contact. J. Biol. Chem. 2000, 275, 11383–11388. [Google Scholar] [CrossRef] [Green Version]
- Khor, S.; Wood, S.; Salvitti, L.; Taylor, D.; Adamson, J.; McNabb, P.; Cary, S. Investigating Diet as the Source of Tetrodotoxin in Pleurobranchaea Maculata. Mar. Drugs 2013, 12, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henikoff, S.; Greally, J.M. Epigenetics, Cellular Memory and Gene Regulation. Curr. Biol. CB 2016, 26, R644–R648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aebersold, R.; Mann, M. Mass-Spectrometric Exploration of Proteome Structure and Function. Nature 2016, 537, 347–355. [Google Scholar] [CrossRef]
- Canzler, S.; Schor, J.; Busch, W.; Schubert, K.; Rolle-Kampczyk, U.E.; Seitz, H.; Kamp, H.; von Bergen, M.; Buesen, R.; Hackermüller, J. Prospects and Challenges of Multi-Omics Data Integration in Toxicology. Arch. Toxicol. 2020, 94, 371–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.; Chaudary, K.; Garmire, L. More is better: Recent progress in multi-omics data integration methods. Front. Genet. 2017, 8, 84. [Google Scholar] [CrossRef] [Green Version]
- Cantini, L.; Zakeri, P.; Hernandez, C.; Naldi, A.; Thieffry, D.; Remy, E.; Baudot, A. Benchmarking joint multi-omics dimensionality reduction approaches for the study of cancer. Nat. Commun. 2021, 12, 124–136. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Toxins | Species | Consequences | Technique | Article |
|---|---|---|---|---|
| STX | Calanus finmarchicus | Less energy: downregulation of lipid biosynthesis, growth, and reproduction, and of antioxidants enzymes. | RNA-Seq | [10] |
| Mytilus chilensis | Involvement of immune response with PRRs. | RNA-Seq and RT-qPCR | [11] | |
| Mytilus chilensis | SOD and CAT as early potential biomarkers of pollution/involvement of immune response. | qPCR | [12] | |
| Mytilus galloprovincialis | Involvement of immune response. | RNA-Seq | [13] | |
| Chlamys farreri, Patinopecten yessoensis | GPx as protectors in the kidneys against the oxidative stress. | RNA-Seq | [14] | |
| Chlamys farreri | Upregulation of SODs in the hepatopancreas and the kidneys. | RNA-Seq | [15] | |
| Patinopecten yessoensis, Crassostrea gigas, Lottia gigantean | Close relation between the expression of SLCs and STX accumulation in the hepatopancreas. | RNA-Seq | [16] | |
| DA | Danio rerio | Upregulation of neurodegeneration, especially memory functions. | Microarray and RT-qPCR | [17] |
| Danio rerio | Five genes of interest involved in glutamate receptor, neurodegeneration, anti-inflammatory, and apoptotic responses. | RT-PCR | [18] | |
| Zalophus californianus | Three dysregulated genes already found in AD studies. | Microarray | [19] | |
| Mytilus galloprovincialis | Upregulation of detoxification processes, the response against the oxidative stress, and immunological processes. | RNA-Seq | [20] | |
| Aequipecten opercularis | Upregulation of detoxification and immunological processes. | RNA-Seq | [21] | |
| Pecten maximus | Upregulation of detoxification and immunological processes. | RNA-Seq | [22] | |
| BTX | Trichechus manatus latirostris | Upregulation of the immune and neurodegenerative pathways. | RNA-Seq | [23] |
| Tursiops truncatus | BTX as haptens that induce an inflammatory immune response. | Restriction site-associated DNA sequencing | [24] | |
| Mus musculus (BALB/C) | No direct binding between BTX6 and AhR. | DNA microarray | [25] | |
| CTX | Mus musculus neurons (primary cultures of mice cortical neurons) | Activation of the mu1 opioid related to the hypothermia induced by CTX treatment. | Microarray | [26] |
| Mus musculus (C57BL6) | Inflammatory response. | Microarray and RT-qPCR | [27] | |
| Mus musculus (C57BL6) | Histamine mediating inflammatory response may cause asthma-like symptoms/dysregulation in detoxification metabolism. | Microarray and RT-qPCR | [28] | |
| Mus musculus (C57BL6) | Detoxification metabolism in the hepatocytes/dysregulation of immune and inflammation systems. | Microarray and RT-qPCR | [29] | |
| Homo sapiens | Inflammatory response, haplotypes DQ2, and DQ8 over-represented. | Microarray and RT-qPCR | [30] | |
| BMAA | Rattus norvegicus cells (OEC) | Dysregulations of apoptosis, excitotoxic pathway, aggregation and degradation of proteins, and cell homeostasis/upregulation of VDAC1. | Microarray and RT-qPCR | [31] |
| TTX | Octopus bimaculoides | TTX resistance: hypothesis of NaV domain 1 mutation. | DNA sequencing and RNA-Seq | [32] |
| Takifugu rubripes | Involvement of the immune system in the liver. | RT-PCR | [33] | |
| Takifugu rubripes | Involvement of the immune system in the liver. | Microarray | [34] | |
| Takifugu rubripes | Modulation of the NaV kinetic. | Microarray and RT-qPCR | [35] | |
| Nassarius succinctus, Nassarius variciferus | Dysregulations of detoxification and immune systems/NaV mutation. | RNA-Seq | [36] | |
| Pleurobranchaea maculata | Difference in TTX levels due to exogenous factors. | PCR amplification and genotyping | [37] |
| Toxins | Species | Consequences | Technique | Article |
|---|---|---|---|---|
| STX | Mus musculus cells (N2A) | Dysregulated proteins in accordance with genotoxicity and neurotoxicity induced by STX/downregulation of proteins suggesting membrane depolarization. | 2D DIGE and MALDI-TOF-MS | [41] |
| Mus musculus (C57BL/6NJ) | Close relationship between dysregulated proteins in long-term effects and neurodegenerative diseases. | TMT labeling and LC-MS/MS | [48] | |
| DA | Zalophus californianus | ApoE as indicator of chronic DAT. | 2D DIGE and LC-MS/MS | [42] |
| Zalophus californianus | Upregulation of CSF proteins involved in neurodegenerative pathway and antiapoptotic response. | Label-free LC-MS/MS | [50] | |
| BTX | Oryzias melastigma | Dysregulation of calcium homeostasis/downregulation of proteins involved in tissue integrity | 2D DIGE and MALDI-TOF/TOF | [43] |
| Porites astreoides | Downregulation of redox homeostasis, energy metabolism, and ROS production. | iTRAQ and LC-MS/MS | [45] | |
| BMAA | Danio rerio | Neurocytotoxic effect/dysregulation of endocannabinoid system. | nanoLC-MS/MS | [51] |
| Homo sapiens tissue (human brain) | Misincorporation of BMAA in de novo synthesis. | LC-MS/MS | [52] | |
| Mus musculus cells (NSC-34) | Link between neurodegenerative diseases and dysregulation of NRF2. | SILAC and nanoLC-MS/MS | [47] | |
| Rattus norvegicus cells (OEC) | Dysregulation of apoptosis, excitotoxic, cell homeostasis pathways/link with proteins involved in AD and ALS. | nanoLC-MS | [31] | |
| Mus musculus cells (NSC-34) | Dysregulation of apoptosis, excitotoxic, cell homeostasis pathways. | LC-MS/MS | [53] | |
| Rattus norvegicus (wistar) | Dose-dependent increase in neuronal pathways and sex-dependent increase in neuropeptides. | nanoLC-MS/MS | [54] | |
| Rattus norvegicus (male wistar) | Memory impairments: decrease in MBP. | MALDI IMS | [55] | |
| Rattus norvegicus (male wistar) | Downstream response to the BMAA-induced intracellular formation of fibrils. | LC-MS/MS | [56] | |
| TTX | Octopus kaurna, Hapalochlaena maculosa | Hyaluronidase as an indicator of TTX production. | LC-MS/MS | [57] |
| Toxins | Species | Consequences | Technique | Article |
|---|---|---|---|---|
| BTX | Oryzias melastigma | Dysregulation of metabolites involved in neural excitotoxicity. | LC-MS/MS | [63] |
| Danio rerio | Dysregulation of excitotoxic, carbohydrate, and energy metabolisms related to BTX mechanism. | HR-MAS-NMR | [64] | |
| BMAA | Homo sapiens cells (SH-SY5Y) | Interference of BMAA with fundamental metabolic pathways related to neurotransmission. | NMR and LC-MS | [65] |
| Rattus norvegicus (male wistar) | Reorganization of metabolic program to increase energy. | NMR | [66] | |
| Danio rerio | Reorganization of metabolic program to increase energy/ROS production and decrease in protection against excitotoxicity and oxidative stress. | HR-MAS-NMR | [68] | |
| Rattus norvegicus (HSD), Mus musculus (B6C3F1/N) | Misincorporation into proteins not proven in urine: excretion problem? | LC-MS/MS | [69] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guillotin, S.; Delcourt, N. Marine Neurotoxins’ Effects on Environmental and Human Health: An OMICS Overview. Mar. Drugs 2022, 20, 18. https://doi.org/10.3390/md20010018
Guillotin S, Delcourt N. Marine Neurotoxins’ Effects on Environmental and Human Health: An OMICS Overview. Marine Drugs. 2022; 20(1):18. https://doi.org/10.3390/md20010018
Chicago/Turabian StyleGuillotin, Sophie, and Nicolas Delcourt. 2022. "Marine Neurotoxins’ Effects on Environmental and Human Health: An OMICS Overview" Marine Drugs 20, no. 1: 18. https://doi.org/10.3390/md20010018
APA StyleGuillotin, S., & Delcourt, N. (2022). Marine Neurotoxins’ Effects on Environmental and Human Health: An OMICS Overview. Marine Drugs, 20(1), 18. https://doi.org/10.3390/md20010018