
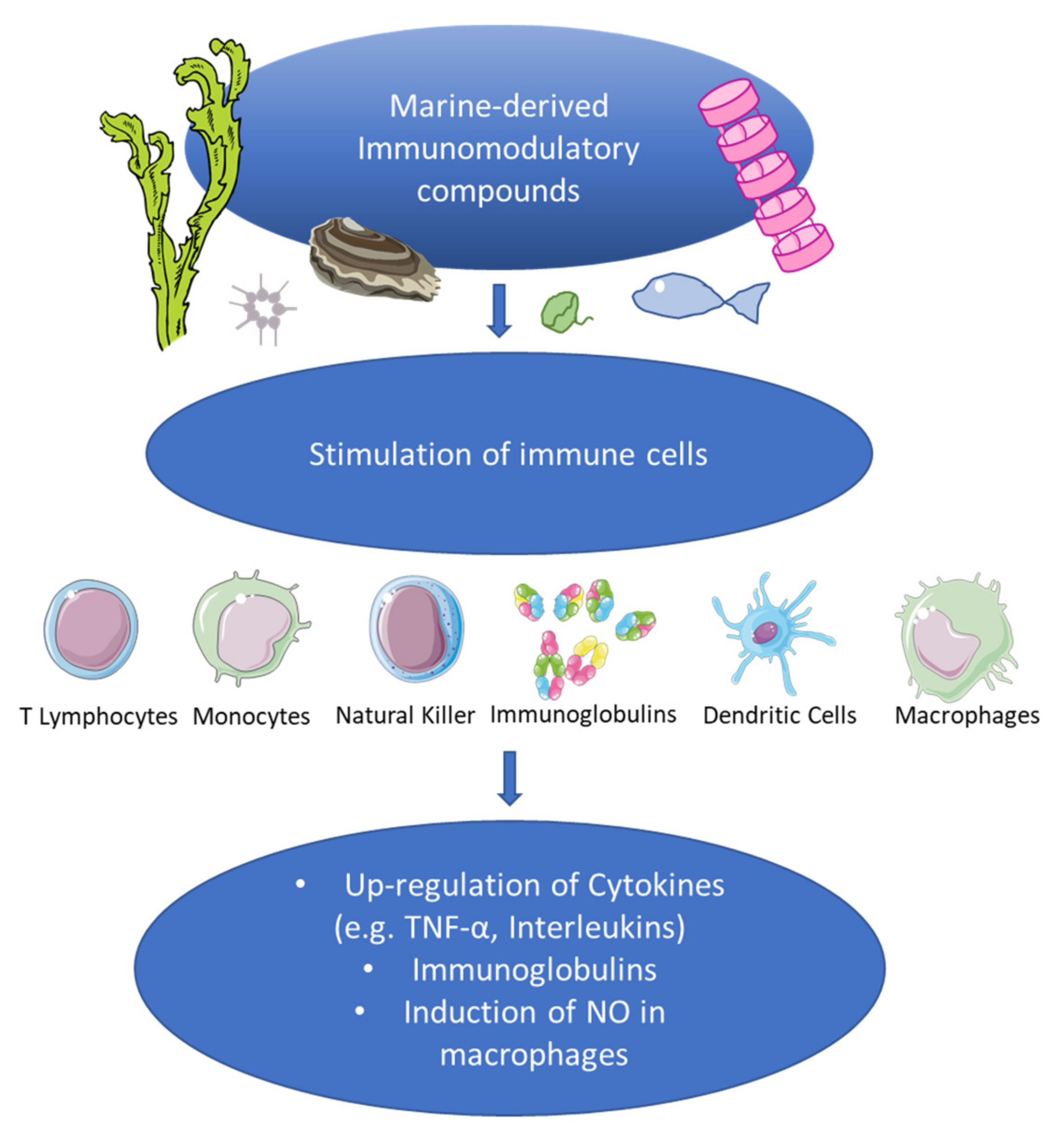
Recent Discoveries on Marine Organism Immunomodulatory Activities




Abstract
:1. Introduction
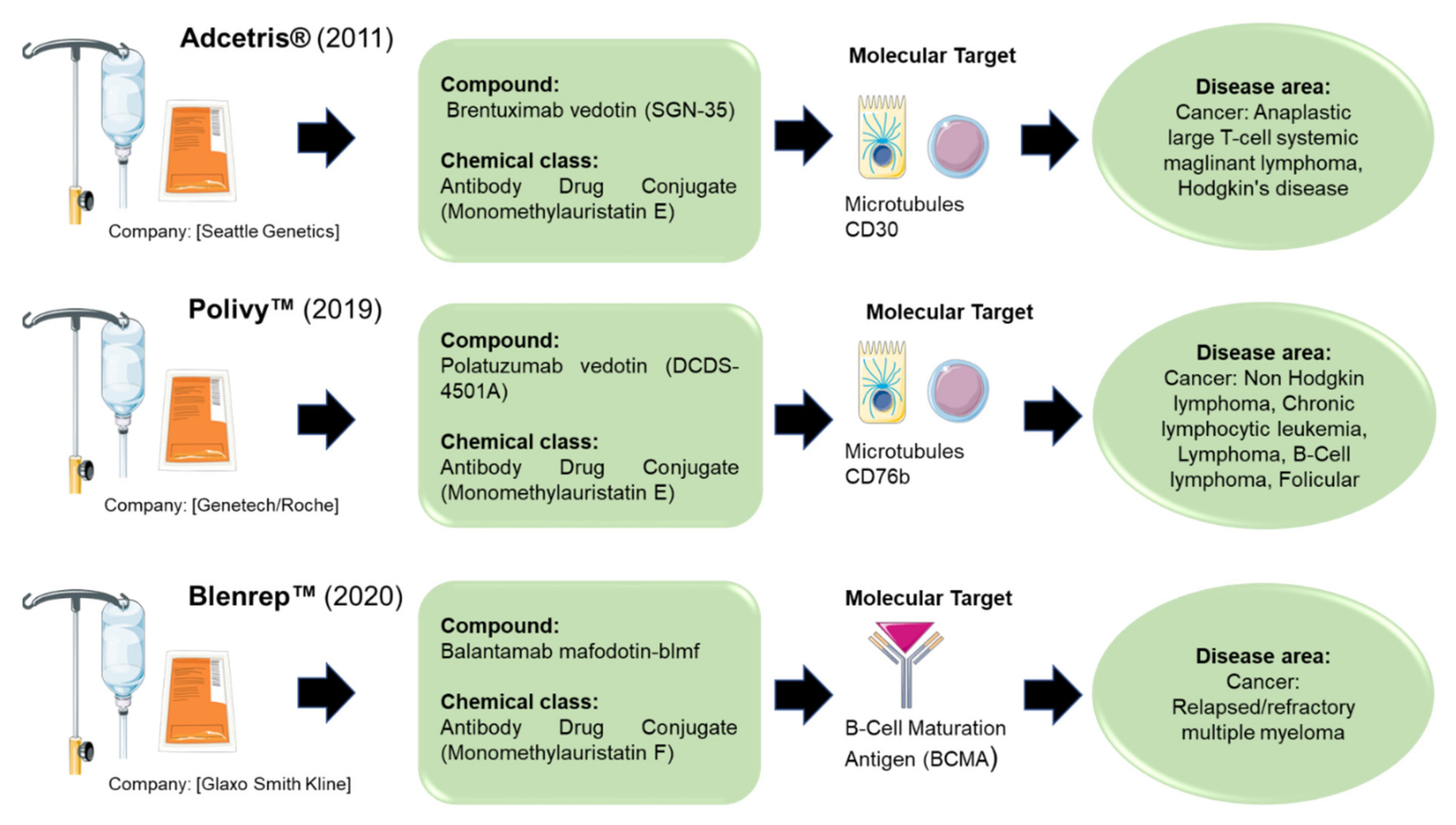
2. Compounds Available on the Market
3. Marine Microorganisms as Source of Immunomodulatory Compounds
3.1. Fungi and Bacteria
3.2. Microalgae
4. Macroorganisms as Source of Immunomodulatory Compounds
4.1. Macroalgae
4.2. Sponges
4.3. Other Species
4.3.1. Mollusks
4.3.2. Corals
4.3.3. Fishes
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Santos, J.D.; Vitorino, I.; Reyes, F.; Vicente, F.; Lage, O.M. From Ocean to Medicine: Pharmaceutical Applications of Metabolites from Marine Bacteria. Antibiotics 2020, 9, 455. [Google Scholar] [CrossRef] [PubMed]
- Cutignano, A.; Nuzzo, G.; Ianora, A.; Luongo, E.; Romano, G.; Gallo, C.; Sansone, C.; Aprea, S.; Mancini, F.; D’Oro, U.; et al. Development and Application of a Novel SPE-Method for Bioassay-Guided Fractionation of Marine Extracts. Mar. Drugs 2015, 13, 5736–5749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotter, A.; Barbier, M.; Bertoni, F.; Bones, A.M.; Cancela, M.L.; Carlsson, J.; Carvalho, M.F.; Cegłowska, M.; Chirivella-Martorell, J.; Conk Dalay, M.; et al. The Essentials of Marine Biotechnology. Front. Mar. Sci. 2021, 8, 629629. [Google Scholar] [CrossRef]
- Ameen, F.; AlNadhari, S.; Al-Homaidan, A.A. Marine Microorganisms as an Untapped Source of Bioactive Compounds. Saudi J. Biol. Sci. 2021, 28, 224–231. [Google Scholar] [CrossRef]
- Alves, A.; Sousa, E.; Kijjoa, A.; Pinto, M. Marine-Derived Compounds with Potential Use as Cosmeceuticals and Nutricosmetics. Molecules 2020, 25, 2536. [Google Scholar] [CrossRef]
- Riccio, G.; Lauritano, C. Microalgae with Immunomodulatory Activities. Mar. Drugs 2019, 18, 2. [Google Scholar] [CrossRef] [Green Version]
- Rubtsova, K.; Rubtsov, A.V.; Thurman, J.M.; Mennona, J.M.; Kappler, J.W.; Marrack, P. B Cells Expressing the Transcription Factor T-Bet Drive Lupus-like Autoimmunity. J. Clin. Investig. 2017, 127, 1392–1404. [Google Scholar] [CrossRef] [Green Version]
- Lauritano, C.; Ianora, A. Grand Challenges in Marine Biotechnology: Overview of Recent EU-Funded Projects. In Grand Challenges in Marine Biotechnology; Rampelotto, P.H., Trincone, A., Eds.; Grand Challenges in Biology and Biotechnology; Springer International Publishing: Cham, Switzerland, 2018; pp. 425–449. ISBN 978-3-319-69074-2. [Google Scholar]
- Mayer, A.M.S.; Rodríguez, A.D.; Berlinck, R.G.S.; Hamann, M.T. Marine Pharmacology in 2003–4: Marine Compounds with Anthelmintic Antibacterial, Anticoagulant, Antifungal, Anti-Inflammatory, Antimalarial, Antiplatelet, Antiprotozoal, Antituberculosis, and Antiviral Activities; Affecting the Cardiovascular, Immune and Nervous Systems, and Other Miscellaneous Mechanisms of Action. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2007, 145, 553–581. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, D.L.; Gange, S.J.; Rose, N.R.; Graham, N.M.H. Epidemiology and Estimated Population Burden of Selected Autoimmune Diseases in the United States. Clin. Immunol. Immunopathol. 1997, 84, 223–243. [Google Scholar] [CrossRef] [Green Version]
- Annual Congress on Radiology. Nature 1962, 193, 1030. [CrossRef]
- Markets and Markets. Available online: https://www.marketsandmarkets.com/Market-Reports/autoimmune-disease-diagnosis-market-174826519.html?gclid=CjwKCAjwquWVBhBrEiwAt1KmwpOR-wBOWHUujkgG8GZW_lyfLcpI2B6MfZfRtby7xVsba0E6PxpNWRoCZ5gQAvD_BwE (accessed on 22 June 2022).
- Da Silva Barbosa, J.; Sabry, D.A.; Silva, C.H.F.; Gomes, D.L.; Santana-Filho, A.P.; Sassaki, G.L.; Rocha, H.A.O. Immunostimulatory Effect of Sulfated Galactans from the Green Seaweed Caulerpa cupressoides Var. flabellata. Mar. Drugs 2020, 18, 234. [Google Scholar] [CrossRef]
- Catanzaro, M.; Corsini, E.; Rosini, M.; Racchi, M.; Lanni, C. Immunomodulators Inspired by Nature: A Review on Curcumin and Echinacea. Molecules 2018, 23, 2778. [Google Scholar] [CrossRef] [Green Version]
- Bergman, P.; Raqib, R.; Rekha, R.S.; Agerberth, B.; Gudmundsson, G.H. Host Directed Therapy Against Infection by Boosting Innate Immunity. Front. Immunol. 2020, 11, 1209. [Google Scholar] [CrossRef]
- Lee, J.S.; Kwon, D.S.; Lee, K.R.; Park, J.M.; Ha, S.-J.; Hong, E.K. Mechanism of Macrophage Activation Induced by Polysaccharide from Cordyceps militaris Culture Broth. Carbohydr. Polym. 2015, 120, 29–37. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Inflammation, Metaflammation and Immunometabolic Disorders. Nature 2017, 542, 177–185. [Google Scholar] [CrossRef]
- Ayeka, P.A. Potential of Mushroom Compounds as Immunomodulators in Cancer Immunotherapy: A Review. Evid. Based Complement. Altern. Med. 2018, 2018, 7271509. [Google Scholar] [CrossRef]
- Sorokina, M.; Steinbeck, C. Review on Natural Products Databases: Where to Find Data in 2020. J. Cheminform. 2020, 12, 20. [Google Scholar] [CrossRef] [Green Version]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, B.; Shah, M.; Choi, S. Oceans as a Source of Immunotherapy. Mar. Drugs 2019, 17, 282. [Google Scholar] [CrossRef] [Green Version]
- Luesch, H.; Moore, R.E.; Paul, V.J.; Mooberry, S.L.; Corbett, T.H. Isolation of Dolastatin 10 from the Marine Cyanobacterium Symploca Species VP642 and Total Stereochemistry and Biological Evaluation of Its Analogue Symplostatin 1. J. Nat. Prod. 2001, 64, 907–910. [Google Scholar] [CrossRef]
- Advani, R.H.; Moskowitz, A.J.; Bartlett, N.L.; Vose, J.M.; Ramchandren, R.; Feldman, T.A.; LaCasce, A.S.; Christian, B.A.; Ansell, S.M.; Moskowitz, C.H.; et al. Brentuximab Vedotin in Combination with Nivolumab in Relapsed or Refractory Hodgkin Lymphoma: 3-Year Study Results. Blood 2021, 138, 427–438. [Google Scholar] [CrossRef]
- Newman, D.J. The “Utility” of Highly Toxic Marine-Sourced Compounds. Mar. Drugs 2019, 17, 324. [Google Scholar] [CrossRef] [Green Version]
- Ekholm, F.; Ruokonen, S.-K.; Redón, M.; Pitkänen, V.; Vilkman, A.; Saarinen, J.; Helin, J.; Satomaa, T.; Wiedmer, S. Hydrophilic Monomethyl Auristatin E Derivatives as Novel Candidates for the Design of Antibody-Drug Conjugates. Separations 2018, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Okazaki, M.; Luo, Y.; Han, T.; Yoshida, M.; Seon, B.K. Three New Monoclonal Antibodies That Define a Unique Antigen Associated with Prolymphocytic Leukemia/Non-Hodgkin’s Lymphoma and Are Effectively Internalized after Binding to the Cell Surface Antigen. Blood 1993, 81, 84–94. [Google Scholar] [CrossRef] [Green Version]
- Sehn, L.H.; Herrera, A.F.; Flowers, C.R.; Kamdar, M.K.; McMillan, A.; Hertzberg, M.; Assouline, S.; Kim, T.M.; Kim, W.S.; Ozcan, M.; et al. Polatuzumab Vedotin in Relapsed or Refractory Diffuse Large B-Cell Lymphoma. JCO 2020, 38, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Behren Ketchum, E.; Clarke, A.; Clemmons, A.B. Belantamab Mafodotin-Blmf: A Novel Antibody-Drug Conjugate for Treatment of Patients With Relapsed/Refractory Multiple Myeloma. JADPRO 2022, 13, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Yi, M.; Ding, L.; He, S. A Review of Anti-Inflammatory Compounds from Marine Fungi, 2000–2018. Mar. Drugs 2019, 17, 636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.-Y.; Cai, S.-X.; Zhang, W.-J.; Tang, X.-L.; Shin, H.-Y.; Lee, J.-Y.; Gu, Q.-Q.; Park, H. Semi-Vioxanthin Isolated from Marine-Derived Fungus Regulates Tumor Necrosis Factor-.ALPHA., Cluster of Differentiation (CD) 80, CD86, and Major Histocompatibility Complex Class II Expression in RAW264.7 Cells via Nuclear Factor-.KAPPA.B and Mitogen-Activated Protein Kinase Signaling Pathways. Biol. Pharm. Bull. 2008, 31, 2228–2233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.-X.; Crews, M.S.; Draskovic, M.; Sohn, J.; Johnson, T.A.; Tenney, K.; Valeriote, F.A.; Yao, X.-J.; Bjeldanes, L.F.; Crews, P. Azonazine, a Novel Dipeptide from a Hawaiian Marine Sediment-Derived Fungus, Aspergillus Insulicola. Org. Lett. 2010, 12, 4458–4461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldring, S.R.; Goldring, M.B. The Role of Cytokines in Cartilage Matrix Degeneration in Osteoarthritis. Clin. Orthop. Relat. Res. 2004, 427, S27–S36. [Google Scholar] [CrossRef]
- Yang, X.; Gao, X.; Han, F.; Xu, B.; Song, Y.; Tan, R. Purification, Characterization and Enzymatic Degradation of YCP, a Polysaccharide from Marine Filamentous Fungus YS4108. Biochimie 2005, 87, 747–754. [Google Scholar] [CrossRef]
- Liu, W.; Tang, S.; Zhao, Q.; Zhang, W.; Li, K.; Yao, W.; Gao, X. The α-D-Glucan from Marine Fungus Phoma herbarum YS4108 Ameliorated Mice Colitis by Repairing Mucosal Barrier and Maintaining Intestinal Homeostasis. Int. J. Biol. Macromol. 2020, 149, 1180–1188. [Google Scholar] [CrossRef]
- Kamada, N.; Seo, S.-U.; Chen, G.Y.; Núñez, G. Role of the Gut Microbiota in Immunity and Inflammatory Disease. Nat. Rev. Immunol. 2013, 13, 321–335. [Google Scholar] [CrossRef]
- Marchese, P.; Young, R.; O’Connell, E.; Afoullouss, S.; Baker, B.J.; Allcock, A.L.; Barry, F.; Murphy, J.M. Deep-Sea Coral Garden Invertebrates and Their Associated Fungi Are Genetic Resources for Chronic Disease Drug Discovery. Mar. Drugs 2021, 19, 390. [Google Scholar] [CrossRef]
- Klemm, E.J.; Wong, V.K.; Dougan, G. Emergence of Dominant Multidrug-Resistant Bacterial Clades: Lessons from History and Whole-Genome Sequencing. Proc. Natl. Acad. Sci. USA. 2018, 115, 12872–12877. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Atienza, E.; Gómez-Sala, B.; Araújo, C.; Campanero, C.; del Campo, R.; Hernández, P.E.; Herranz, C.; Cintas, L.M. Antimicrobial Activity, Antibiotic Susceptibility and Virulence Factors of Lactic Acid Bacteria of Aquatic Origin Intended for Use as Probiotics in Aquaculture. BMC Microbiol. 2013, 13, 15. [Google Scholar] [CrossRef] [Green Version]
- De Azevedo, R.V.; Fosse Filho, J.C.; Cardoso, L.D.; Mattos, D.d.C.; Vidal Júnior, M.V.; Andrade, D.R.d. Economic Evaluation of Prebiotics, Probiotics and Symbiotics in Juvenile Nile tilapia. Rev. Ciênc. Agron. 2015, 46, 72–79. [Google Scholar] [CrossRef] [Green Version]
- Standen, B.T.; Rawling, M.D.; Davies, S.J.; Castex, M.; Foey, A.; Gioacchini, G.; Carnevali, O.; Merrifield, D.L. Probiotic Pediococcus Acidilactici Modulates Both Localised Intestinal- and Peripheral-Immunity in Tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2013, 35, 1097–1104. [Google Scholar] [CrossRef]
- Wasana, W.P.; Senevirathne, A.; Nikapitiya, C.; Eom, T.-Y.; Lee, Y.; Lee, J.-S.; Kang, D.-H.; Oh, C.; De Zoysa, M. A Novel Pseudoalteromonas xiamenensis Marine Isolate as a Potential Probiotic: Anti-Inflammatory and Innate Immune Modulatory Effects against Thermal and Pathogenic Stresses. Mar. Drugs 2021, 19, 707. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics Consensus Statement on the Scope and Appropriate Use of the Term Probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Ramesh, D.; Souissi, S. Effects of Potential Probiotic Bacillus Subtilis KADR1 and Its Subcellular Components on Immune Responses and Disease Resistance in Labeo Rohita. Aquac. Res. 2018, 49, 367–377. [Google Scholar] [CrossRef]
- Meidong, R.; Khotchanalekha, K.; Doolgindachbaporn, S.; Nagasawa, T.; Nakao, M.; Sakai, K.; Tongpim, S. Evaluation of Probiotic Bacillus aerius B81e Isolated from Healthy Hybrid Catfish on Growth, Disease Resistance and Innate Immunity of Pla-Mong Pangasius bocourti. Fish Shellfish Immunol. 2018, 73, 1–10. [Google Scholar] [CrossRef]
- Verschuere, L.; Rombaut, G.; Sorgeloos, P.; Verstraete, W. Probiotic Bacteria as Biological Control Agents in Aquaculture. Microbiol. Mol. Biol. Rev. 2000, 64, 655–671. [Google Scholar] [CrossRef] [Green Version]
- Lauritano, C.; Andersen, J.H.; Hansen, E.; Albrigtsen, M.; Escalera, L.; Esposito, F.; Helland, K.; Hanssen, K.Ø.; Romano, G.; Ianora, A. Bioactivity Screening of Microalgae for Antioxidant, Anti-Inflammatory, Anticancer, Anti-Diabetes, and Antibacterial Activities. Front. Mar. Sci. 2016, 3, 68. [Google Scholar] [CrossRef] [Green Version]
- Heo, S.-J.; Yoon, W.-J.; Kim, K.-N.; Oh, C.; Choi, Y.-U.; Yoon, K.-T.; Kang, D.-H.; Qian, Z.-J.; Choi, I.-W.; Jung, W.-K. Anti-Inflammatory Effect of Fucoxanthin Derivatives Isolated from Sargassum siliquastrum in Lipopolysaccharide-Stimulated RAW 264.7 Macrophage. Food Chem. Toxicol. 2012, 50, 3336–3342. [Google Scholar] [CrossRef]
- Costa, J.A.V.; Lucas, B.F.; Alvarenga, A.G.P.; Moreira, J.B.; de Morais, M.G. Microalgae Polysaccharides: An Overview of Production, Characterization, and Potential Applications. Polysaccharides 2021, 2, 759–772. [Google Scholar] [CrossRef]
- Faculty of Fisheries and Marine Sciences; Mutmainnah, N.; Risjani, Y.; Hertika, A.M.S. Growth Rate and Chemical Composition of Secondary Metabolite Extracellular Polysaccharide (EPS) in Microalga Porphyridium cruentum. JELS 2018, 8, 97–102. [Google Scholar] [CrossRef]
- Risjani, Y.; Mutmainnah, N.; Manurung, P.; Wulan, S.N. Yunianta Exopolysaccharide from Porphyridium cruentum (purpureum) Is Not Toxic and Stimulates Immune Response against Vibriosis: The Assessment Using Zebrafish and White Shrimp Litopenaeus vannamei. Mar. Drugs 2021, 19, 133. [Google Scholar] [CrossRef] [PubMed]
- Sathasivam, R.; Radhakrishnan, R.; Hashem, A.; Abd_Allah, E.F. Microalgae Metabolites: A Rich Source for Food and Medicine. Saudi J. Biol. Sci. 2019, 26, 709–722. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.C.; Ferreira, I.C.F.R.; Dias, M.M.; Barreiro, M.F. Microalgae-Derived Pigments: A 10-Year Bibliometric Review and Industry and Market Trend Analysis. Molecules 2020, 25, 3406. [Google Scholar] [CrossRef]
- Chang, M.X.; Xiong, F. Astaxanthin and Its Effects in Inflammatory Responses and Inflammation-Associated Diseases: Recent Advances and Future Directions. Molecules 2020, 25, 5342. [Google Scholar] [CrossRef]
- Speranza, L.; Pesce, M.; Patruno, A.; Franceschelli, S.; de Lutiis, M.A.; Grilli, A.; Felaco, M. Astaxanthin Treatment Reduced Oxidative Induced Pro-Inflammatory Cytokines Secretion in U937: SHP-1 as a Novel Biological Target. Mar. Drugs 2012, 10, 890–899. [Google Scholar] [CrossRef]
- Tan, K.; Zhang, H.; Lim, L.-S.; Ma, H.; Li, S.; Zheng, H. Roles of Carotenoids in Invertebrate Immunology. Front. Immunol. 2020, 10, 3041. [Google Scholar] [CrossRef]
- Widyaningrum, D.; Oktafika, R.A.; Cecilia, D. Microalgae Pigments as a Promising Immunomodulating Food Ingredient: In Silico Study. IOP Conf. Ser. Earth Environ. Sci. 2022, 998, 012056. [Google Scholar] [CrossRef]
- Pantsar, T.; Poso, A. Binding Affinity via Docking: Fact and Fiction. Molecules 2018, 23, 1899. [Google Scholar] [CrossRef] [Green Version]
- Fuster, J.J.; Walsh, K. The Good, the Bad, and the Ugly of Interleukin-6 Signaling. EMBO J. 2014, 33, 1425–1427. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in Inflammation, Immunity, and Disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef]
- Zia, K.; Ashraf, S.; Jabeen, A.; Saeed, M.; Nur-e-Alam, M.; Ahmed, S.; Al-Rehaily, A.J.; Ul-Haq, Z. Identification of Potential TNF-α Inhibitors: From in Silico to in Vitro Studies. Sci. Rep. 2020, 10, 20974. [Google Scholar] [CrossRef]
- Shen, C.; Liu, H.; Wang, X.; Lei, T.; Wang, E.; Xu, L.; Yu, H.; Li, D.; Yao, X. Importance of Incorporating Protein Flexibility in Molecule Modeling: A Theoretical Study on Type I1/2 NIK Inhibitors. Front. Pharmacol. 2019, 10, 345. [Google Scholar] [CrossRef]
- Reis, B.; Gonçalves, A.T.; Santos, P.; Sardinha, M.; Conceição, L.E.C.; Serradeiro, R.; Pérez-Sánchez, J.; Calduch-Giner, J.; Schmid-Staiger, U.; Frick, K.; et al. Immune Status and Hepatic Antioxidant Capacity of Gilthead Seabream Sparus aurata Juveniles Fed Yeast and Microalga Derived β-Glucans. Mar. Drugs 2021, 19, 653. [Google Scholar] [CrossRef]
- Kim, S.M.; Jung, Y.-J.; Kwon, O.-N.; Cha, K.H.; Um, B.-H.; Chung, D.; Pan, C.-H. A Potential Commercial Source of Fucoxanthin Extracted from the Microalga Phaeodactylum tricornutum. Appl. Biochem. Biotechnol. 2012, 166, 1843–1855. [Google Scholar] [CrossRef] [PubMed]
- Gügi, B.; Le Costaouec, T.; Burel, C.; Lerouge, P.; Helbert, W.; Bardor, M. Diatom-Specific Oligosaccharide and Polysaccharide Structures Help to Unravel Biosynthetic Capabilities in Diatoms. Mar. Drugs 2015, 13, 5993–6018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert-López, B.; Barranco, A.; Herrero, M.; Cifuentes, A.; Ibáñez, E. Development of New Green Processes for the Recovery of Bioactives from Phaeodactylum tricornutum. Food Res. Int. 2017, 99, 1056–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, S.K.; Beck, B.R.; Kim, D.; Park, J.; Kim, J.; Kim, H.D.; Ringø, E. Prebiotics as Immunostimulants in Aquaculture: A Review. Fish Shellfish Immunol. 2014, 40, 40–48. [Google Scholar] [CrossRef]
- Meena, D.K.; Das, P.; Kumar, S.; Mandal, S.C.; Prusty, A.K.; Singh, S.K.; Akhtar, M.S.; Behera, B.K.; Kumar, K.; Pal, A.K.; et al. Beta-Glucan: An Ideal Immunostimulant in Aquaculture (a Review). Fish Physiol. Biochem. 2013, 39, 431–457. [Google Scholar] [CrossRef]
- Soltanian, S.; Stuyven, E.; Cox, E.; Sorgeloos, P.; Bossier, P. Beta-Glucans as Immunostimulant in Vertebrates and Invertebrates. Crit. Rev. Microbiol. 2009, 35, 109–138. [Google Scholar] [CrossRef]
- Ainsworth, A.J. A β-Glucan Inhibitable Zymosan Receptor on Channel Catfish Neutrophils. Vet. Immunol. Immunopathol. 1994, 41, 141–152. [Google Scholar] [CrossRef]
- Chang, C.-S.; Huang, S.-L.; Chen, S.; Chen, S.-N. Innate Immune Responses and Efficacy of Using Mushroom Beta-Glucan Mixture (MBG) on Orange-Spotted Grouper, Epinephelus coioides, Aquaculture. Fish Shellfish Immunol. 2013, 35, 115–125. [Google Scholar] [CrossRef]
- Bagni, M.; Archetti, L.; Amadori, M.; Marino, G. Effect of Long-Term Oral Administration of an Immunostimulant Diet on Innate Immunity in Sea Bass (Dicentrarchus labrax). J. Vet. Med. Series B 2000, 47, 745–751. [Google Scholar] [CrossRef]
- Paredes, M.; Gonzalez, K.; Figueroa, J.; Montiel-Eulefi, E. Immunomodulatory Effect of Prolactin on Atlantic Salmon (Salmo salar) Macrophage Function. Fish Physiol. Biochem. 2013, 39, 1215–1221. [Google Scholar] [CrossRef]
- Hwang, J.; Yadav, D.; Lee, P.C.; Jin, J. Immunomodulatory Effects of Polysaccharides from Marine Algae for Treating Cancer, Infectious Disease, and Inflammation. Phytother. Res. 2022, 36, 761–777. [Google Scholar] [CrossRef]
- Manzo, E.; Cutignano, A.; Pagano, D.; Gallo, C.; Barra, G.; Nuzzo, G.; Sansone, C.; Ianora, A.; Urbanek, K.; Fenoglio, D.; et al. A New Marine-Derived Sulfoglycolipid Triggers Dendritic Cell Activation and Immune Adjuvant Response. Sci. Rep. 2017, 7, 6286. [Google Scholar] [CrossRef]
- De Jesus Raposo, M.; de Morais, A.; de Morais, R. Marine Polysaccharides from Algae with Potential Biomedical Applications. Mar. Drugs 2015, 13, 2967–3028. [Google Scholar] [CrossRef]
- Hafting, J.T.; Craigie, J.S.; Stengel, D.B.; Loureiro, R.R.; Buschmann, A.H.; Yarish, C.; Edwards, M.D.; Critchley, A.T. Prospects and Challenges for Industrial Production of Seaweed Bioactives. J. Phycol. 2015, 51, 821–837. [Google Scholar] [CrossRef]
- Wells, M.L.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.S.; Helliwell, K.E.; Smith, A.G.; Camire, M.E.; Brawley, S.H. Algae as Nutritional and Functional Food Sources: Revisiting Our Understanding. J. Appl. Phycol. 2017, 29, 949–982. [Google Scholar] [CrossRef]
- Dillehay, T.D.; Ramírez, C.; Pino, M.; Collins, M.B.; Rossen, J.; Pino-Navarro, J.D. Monte Verde: Seaweed, Food, Medicine, and the Peopling of South America. Science 2008, 320, 784–786. [Google Scholar] [CrossRef] [Green Version]
- Mazarrasa, I.; Olsen, Y.S.; Mayol, E.; Marbà, N.; Duarte, C.M. Global Unbalance in Seaweed Production, Research Effort and Biotechnology Markets. Biotechnol. Adv. 2014, 32, 1028–1036. [Google Scholar] [CrossRef]
- Parker, K.H.; Beury, D.W.; Ostrand-Rosenberg, S. Myeloid-Derived Suppressor Cells. In Advances in Cancer Research; Elsevier: Amsterdam, The Netherlands, 2015; Volume 128, pp. 95–139. ISBN 978-0-12-802316-7. [Google Scholar]
- Faury, G.; Ruszova, E.; Molinari, J.; Mariko, B.; Raveaud, S.; Velebny, V.; Robert, L. The α-l-Rhamnose Recognizing Lectin Site of Human Dermal Fibroblasts Functions as a Signal Transducer. Biochim. Et Biophys. Acta BBA Gen. Subj. 2008, 1780, 1388–1394. [Google Scholar] [CrossRef]
- Kidgell, J.T.; Glasson, C.R.K.; Magnusson, M.; Vamvounis, G.; Sims, I.M.; Carnachan, S.M.; Hinkley, S.F.R.; Lopata, A.L.; de Nys, R.; Taki, A.C. The Molecular Weight of Ulvan Affects the in Vitro Inflammatory Response of a Murine Macrophage. Int. J. Biol. Macromol. 2020, 150, 839–848. [Google Scholar] [CrossRef]
- Mizuno, M.; Sakaguchi, K.; Sakane, I. Oral Administration of Fucoidan Can Exert Anti-Allergic Activity after Allergen Sensitization by Enhancement of Galectin-9 Secretion in Blood. Biomolecules 2020, 10, 258. [Google Scholar] [CrossRef] [Green Version]
- Chandía, N.P.; Matsuhiro, B. Characterization of a Fucoidan from Lessonia vadosa (Phaeophyta) and Its Anticoagulant and Elicitor Properties. Int. J. Biol. Macromol. 2008, 42, 235–240. [Google Scholar] [CrossRef]
- Yang, C.; Chung, D.; Shin, I.-S.; Lee, H.; Kim, J.; Lee, Y.; You, S. Effects of Molecular Weight and Hydrolysis Conditions on Anticancer Activity of Fucoidans from Sporophyll of Undaria pinnatifida. Int. J. Biol. Macromol. 2008, 43, 433–437. [Google Scholar] [CrossRef]
- Li, B.; Lu, F.; Wei, X.; Zhao, R. Fucoidan: Structure and Bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar] [CrossRef] [Green Version]
- Cumashi, A.; Ushakova, N.A.; Preobrazhenskaya, M.E.; D’Incecco, A.; Piccoli, A.; Totani, L.; Tinari, N.; Morozevich, G.E.; Berman, A.E.; Bilan, M.I.; et al. A Comparative Study of the Anti-Inflammatory, Anticoagulant, Antiangiogenic, and Antiadhesive Activities of Nine Different Fucoidans from Brown Seaweeds. Glycobiology 2007, 17, 541–552. [Google Scholar] [CrossRef] [Green Version]
- Kwak, J.-Y. Fucoidan as a Marine Anticancer Agent in Preclinical Development. Mar. Drugs 2014, 12, 851–870. [Google Scholar] [CrossRef]
- Jin, J.-O.; Zhang, W.; Du, J.-Y.; Wong, K.-W.; Oda, T.; Yu, Q. Fucoidan Can Function as an Adjuvant In Vivo to Enhance Dendritic Cell Maturation and Function and Promote Antigen-Specific T Cell Immune Responses. PLoS ONE 2014, 9, e99396. [Google Scholar] [CrossRef]
- Peng, Y.; Song, Y.; Wang, Q.; Hu, Y.; He, Y.; Ren, D.; Wu, L.; Liu, S.; Cong, H.; Zhou, H. In Vitro and in Vivo Immunomodulatory Effects of Fucoidan Compound Agents. Int. J. Biol. Macromol. 2019, 127, 48–56. [Google Scholar] [CrossRef]
- Zhang, W.; Park, H.-B.; Yadav, D.; Hwang, J.; An, E.-K.; Eom, H.-Y.; Kim, S.-J.; Kwak, M.; Lee, P.C.-W.; Jin, J.-O. Comparison of Human Peripheral Blood Dendritic Cell Activation by Four Fucoidans. Int. J. Biol. Macromol. 2021, 174, 477–484. [Google Scholar] [CrossRef]
- Ngo, D.-H.; Kim, S.-K. Sulfated Polysaccharides as Bioactive Agents from Marine Algae. Int. J. Biol. Macromol. 2013, 62, 70–75. [Google Scholar] [CrossRef]
- Liu, Q.; Zhou, Y.; Ma, L.; Gu, F.; Liao, K.; Liu, Y.; Zhang, Y.; Liu, H.; Hong, Y.; Cao, M.; et al. Sulfate Oligosaccharide of Gracilaria Lemaneiformis Modulates Type 1 Immunity by Restraining T Cell Activation. Carbohydr. Polym. 2022, 288, 119377. [Google Scholar] [CrossRef]
- Autoimmune Diseases: A Leading Cause of Death among Young and Middle-Aged Women in the United States. Am. J. Public Health 2000, 90, 1463–1466. [CrossRef] [PubMed] [Green Version]
- Anjum, K.; Abbas, S.Q.; Shah, S.A.A.; Akhter, N.; Batool, S.; ul Hassan, S.S. Marine Sponges as a Drug Treasure. Biomol. Ther. 2016, 24, 347–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, A.M.S.; Glaser, K.B.; Cuevas, C.; Jacobs, R.S.; Kem, W.; Little, R.D.; McIntosh, J.M.; Newman, D.J.; Potts, B.C.; Shuster, D.E. The Odyssey of Marine Pharmaceuticals: A Current Pipeline Perspective. Trends Pharmacol. Sci. 2010, 31, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Talley, R.W.; O’Bryan, R.M.; Tucker, W.G.; Loo, R.V. Clinical Pharmacology and Human Antitumor Activity of Cytosine Arabinoside. Cancer 1967, 20, 809–816. [Google Scholar] [CrossRef]
- Pejin, B.; Iodice, C.; Kojic, V.; Jakimov, D.; Lazovic, M.; Tommonaro, G. In Vitro Evaluation of Cytotoxic and Mutagenic Activity of Avarol. Nat. Prod. Res. 2016, 30, 1293–1296. [Google Scholar] [CrossRef]
- Altmann, K.-H. Microtubule-Stabilizing Agents: A Growing Class of Important Anticancer Drugs. Curr. Opin. Chem. Biol. 2001, 5, 424–431. [Google Scholar] [CrossRef]
- Rangel, M.; Falkenberg, M. An Overview of the Marine Natural Products in Clinical Trials and on the Market. J. Coast Life Med. 2015, 3, 421–428(8). [Google Scholar] [CrossRef]
- Liu, K.K.-C.; Sakya, S.M.; O’Donnell, C.J.; Flick, A.C.; Ding, H.X. Synthetic Approaches to the 2010 New Drugs. Bioorganic Med. Chem. 2012, 20, 1155–1174. [Google Scholar] [CrossRef]
- de Almeida Leone, P.; Redburn, J.; Hooper, J.N.A.; Quinn, R.J. Polyoxygenated Dysidea Sterols That Inhibit the Binding of [I125] IL-8 to the Human Recombinant IL-8 Receptor Type A. J. Nat. Prod. 2000, 63, 694–697. [Google Scholar] [CrossRef]
- Gunathilake, V.; Bertolino, M.; Bavestrello, G.; Udagama, P. Immunomodulatory Activity of the Marine Sponge, Haliclona (Soestella) sp. (Haplosclerida: Chalinidae), from Sri Lanka in Wistar Albino Rats: Immunosuppression and Th1-Skewed Cytokine Response. J. Immunol. Res. 2020, 2020, 7281295. [Google Scholar] [CrossRef]
- Paolino, M.; Penninger, J.M. Cbl-b in T-Cell Activation. Semin. Immunopathol. 2010, 32, 137–148. [Google Scholar] [CrossRef]
- Jiang, W.; Wang, D.; Wilson, B.; Kang, U.; Bokesch, H.; Smith, E.; Wamiru, A.; Goncharova, E.; Voeller, D.; Lipkowitz, S.; et al. Agelasine Diterpenoids and Cbl-b Inhibitory Ageliferins from the Coralline Demosponge Astrosclera willeyana. Mar. Drugs 2021, 19, 361. [Google Scholar] [CrossRef]
- Chakrabarti, S.; Jahandideh, F.; Wu, J. Food-Derived Bioactive Peptides on Inflammation and Oxidative Stress. BioMed Res. Int. 2014, 2014, 608979. [Google Scholar] [CrossRef] [Green Version]
- Chalamaiah, M.; Yu, W.; Wu, J. Immunomodulatory and Anticancer Protein Hydrolysates (Peptides) from Food Proteins: A Review. Food Chem. 2018, 245, 205–222. [Google Scholar] [CrossRef]
- Shi, H.; Bi, S.; Li, H.; Li, J.; Li, C.; Yu, R.; Song, L.; Zhu, J. Purification and Characterization of a Novel Mixed-Linkage α,β-d-Glucan from Arca subcrenata and Its Immunoregulatory Activity. Int. J. Biol. Macromol. 2021, 182, 207–216. [Google Scholar] [CrossRef]
- Xiang, X.-W.; Wang, R.; Yao, L.-W.; Zhou, Y.-F.; Sun, P.-L.; Zheng, B.; Chen, Y.-F. Anti-Inflammatory Effects of Mytilus coruscus Polysaccharide on RAW264.7 Cells and DSS-Induced Colitis in Mice. Mar. Drugs 2021, 19, 468. [Google Scholar] [CrossRef]
- Liu, W.; Bi, S.; Li, C.; Zheng, H.; Guo, Z.; Luo, Y.; Ou, X.; Song, L.; Zhu, J.; Yu, R. Purification and Characterization of a New CRISP-Related Protein from Scapharca broughtonii and Its Immunomodulatory Activity. Mar. Drugs 2020, 18, 299. [Google Scholar] [CrossRef]
- Zhang, W.; Wei, Y.; Cao, X.; Guo, K.; Wang, Q.; Xiao, X.; Zhai, X.; Wang, D.; Huang, Z. Enzymatic Preparation of Crassostrea oyster Peptides and Their Promoting Effect on Male Hormone Production. J. Ethnopharmacol. 2021, 264, 113382. [Google Scholar] [CrossRef]
- Wang, L.; Zhu, L.; Qin, S. Gut Microbiota Modulation on Intestinal Mucosal Adaptive Immunity. J. Immunol. Res. 2019, 2019, 4735040. [Google Scholar] [CrossRef]
- Shi, N.; Li, N.; Duan, X.; Niu, H. Interaction between the Gut Microbiome and Mucosal Immune System. Military Med. Res. 2017, 4, 14. [Google Scholar] [CrossRef]
- Danese, S.; Sans, M.; Fiocchi, C. Inflammatory Bowel Disease: The Role of Environmental Factors. Autoimmun. Rev. 2004, 3, 394–400. [Google Scholar] [CrossRef]
- Li, Y.; Jin, L.; Chen, T. The Effects of Secretory IgA in the Mucosal Immune System. BioMed Res. Int. 2020, 2020, 2032057. [Google Scholar] [CrossRef]
- Wang, J.; Liang, Q.; Zhao, Q.; Tang, Q.; Ahmed, A.F.; Zhang, Y.; Kang, W. The Effect of Microbial Composition and Proteomic on Improvement of Functional Constipation by Chrysanthemum morifolium Polysaccharide. Food Chem. Toxicol. 2021, 153, 112305. [Google Scholar] [CrossRef]
- Xiang, X.-W.; Zheng, H.-Z.; Wang, R.; Chen, H.; Xiao, J.-X.; Zheng, B.; Liu, S.-L.; Ding, Y.-T. Ameliorative Effects of Peptides Derived from Oyster (Crassostrea gigas) on Immunomodulatory Function and Gut Microbiota Structure in Cyclophosphamide-Treated Mice. Mar. Drugs 2021, 19, 456. [Google Scholar] [CrossRef]
- Ciavatta, M.L.; Lefranc, F.; Carbone, M.; Mollo, E.; Gavagnin, M.; Betancourt, T.; Dasari, R.; Kornienko, A.; Kiss, R. Marine Mollusk-Derived Agents with Antiproliferative Activity as Promising Anticancer Agents to Overcome Chemotherapy Resistance: Marine Mollusk-Derived Agents With Antiproliferative Activity. Med. Res. Rev. 2017, 37, 702–801. [Google Scholar] [CrossRef]
- Benkendorff, K. Molluscan Biological and Chemical Diversity: Secondary Metabolites and Medicinal Resources Produced by Marine Molluscs. Biol. Rev. 2010, 85, 757–775. [Google Scholar] [CrossRef]
- Wright, R.M.; Strader, M.E.; Genuise, H.M.; Matz, M. Effects of Thermal Stress on Amount, Composition, and Antibacterial Properties of Coral Mucus. PeerJ 2019, 7, e6849. [Google Scholar] [CrossRef] [Green Version]
- Meikle, P.; Richards, G.N.; Yellowlees, D. Structural Determination of the Oligosaccharide Side Chains from a Glycoprotein Isolated from the Mucus of the Coral Acropora formosa. J. Biol. Chem. 1987, 262, 16941–16947. [Google Scholar] [CrossRef]
- Krediet, C.J.; Ritchie, K.B.; Cohen, M.; Lipp, E.K.; Sutherland, K.P.; Teplitski, M. Utilization of Mucus from the Coral Acropora Palmata by the Pathogen Serratia Marcescens and by Environmental and Coral Commensal Bacteria. Appl. Environ. Microbiol. 2009, 75, 3851–3858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chernikov, O.V.; Chiu, H.-W.; Li, L.-H.; Kokoulin, M.S.; Molchanova, V.I.; Hsu, H.-T.; Ho, C.-L.; Hua, K.-F. Immunomodulatory Properties of Polysaccharides from the Coral Pseudopterogorgia americana in Macrophages. Cells 2021, 10, 3531. [Google Scholar] [CrossRef] [PubMed]
- Portela, L.C.P.N.; Cahú, T.B.; Bezerra, T.S.; do Nascimento Santos, D.K.D.; Sousa, G.F.; Portela, R.W.S.; Melo, C.M.L.; de Souza Bezerra, R. Biocompatibility and Immunostimulatory Properties of Fish Collagen and Shrimp Chitosan towards Peripheral Blood Mononuclear Cells (PBMCs). Int. J. Biol. Macromol. 2022, 210, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Niyonsaba, F.; Ushio, H.; Hara, M.; Yokoi, H.; Tominaga, M.; Takamori, K.; Kajiwara, N.; Saito, H.; Nagaoka, I.; Ogawa, H.; et al. Antimicrobial Peptides Human β-Defensins and Cathelicidin LL-37 Induce the Secretion of a Pruritogenic Cytokine IL-31 by Human Mast Cells. J. Immunol. 2010, 184, 3526–3534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonawane, A.; Santos, J.C.; Mishra, B.B.; Jena, P.; Progida, C.; Sorensen, O.E.; Gallo, R.; Appelberg, R.; Griffiths, G. Cathelicidin Is Involved in the Intracellular Killing of Mycobacteria in Macrophages: The Role of Cathelicidin in Mycobacteria Killing. Cell. Microbiol. 2011, 13, 1601–1617. [Google Scholar] [CrossRef]
- Wan, M.; Soehnlein, O.; Tang, X.; Does, A.M.; Smedler, E.; Uhlén, P.; Lindbom, L.; Agerberth, B.; Haeggström, J.Z. Cathelicidin LL-37 Induces Time-resolved Release of LTB4 and TXA2 by Human Macrophages and Triggers Eicosanoid Generation in Vivo. FASEB J. 2014, 28, 3456–3467. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.H.; Lillehoj, H.S.; Min, W. Evaluation of the Immunomodulatory Activity of the Chicken NK-Lysin-Derived Peptide CNK-2. Sci. Rep. 2017, 7, 45099. [Google Scholar] [CrossRef] [Green Version]
- Hancock, R.E.W.; Sahl, H.-G. Antimicrobial and Host-Defense Peptides as New Anti-Infective Therapeutic Strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef]
- Cotton, S.; Donnelly, S.; Robinson, M.W.; Dalton, J.P.; Thivierge, K. Defense Peptides Secreted by Helminth Pathogens: Antimicrobial and/or Immunomodulator Molecules? Front. Immun. 2012, 3, 269. [Google Scholar] [CrossRef] [Green Version]
- Andraö, J.; Monreal, D.; de Tejada, G.M.; Olak, C.; Brezesinski, G.; Gomez, S.S.; Goldmann, T.; Bartels, R.; Brandenburg, K.; Moriyon, I. Rationale for the Design of Shortened Derivatives of the NK-Lysin-Derived Antimicrobial Peptide NK-2 with Improved Activity against Gram-Negative Pathogens. J. Biol. Chem. 2007, 282, 14719–14728. [Google Scholar] [CrossRef] [Green Version]
- Kumar, J.; Okada, S.; Clayberger, C.; Krensky, A.M. Granulysin: A Novel Antimicrobial. Expert Opin. Investig. Drugs 2001, 10, 321–329. [Google Scholar] [CrossRef]
- Zhang, M.; Long, H.; Sun, L. A NK-Lysin from Cynoglossus Semilaevis Enhances Antimicrobial Defense against Bacterial and Viral Pathogens. Dev. Comp. Immunol. 2013, 40, 258–265. [Google Scholar] [CrossRef]
- Pereiro, P.; Varela, M.; Diaz-Rosales, P.; Romero, A.; Dios, S.; Figueras, A.; Novoa, B. Zebrafish Nk-Lysins: First Insights about Their Cellular and Functional Diversification. Dev. Comp. Immunol. 2015, 51, 148–159. [Google Scholar] [CrossRef] [Green Version]
- Cai, S.; Wang, J.; Wang, K.; Chen, D.; Dong, X.; Liu, T.; Zeng, Y.; Wang, X.; Wu, D. Expression, Purification and Antibacterial Activity of NK-Lysin Mature Peptides from the Channel Catfish (Ictalurus punctatus). Appl. Sci. 2016, 6, 240. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.L.; Wang, M.C.; Liu, Y.L.; Zhang, Q.; Li, C.F.; Liu, P.T.; Li, E.Z.; Nie, P.; Xie, H.X. Identification, Expression Analysis, and Antibacterial Activity of NK-Lysin from Common Carp Cyprinus carpio. Fish Shellfish Immunol. 2018, 73, 11–21. [Google Scholar] [CrossRef]
- Huang, Y.; Zheng, Q.; Niu, J.; Tang, J.; Wang, B.; Abarike, E.D.; Lu, Y.; Cai, J.; Jian, J. NK-Lysin from Oreochromis niloticus Improves Antimicrobial Defence against Bacterial Pathogens. Fish Shellfish Immunol. 2018, 72, 259–265. [Google Scholar] [CrossRef]
- Pereiro, P.; Romero, A.; Díaz-Rosales, P.; Estepa, A.; Figueras, A.; Novoa, B. Nucleated Teleost Erythrocytes Play an Nk-Lysin- and Autophagy-Dependent Role in Antiviral Immunity. Front. Immunol. 2017, 8, 1458. [Google Scholar] [CrossRef]
- Acosta, J.; Roa, F.; González-Chavarría, I.; Astuya, A.; Maura, R.; Montesino, R.; Muñoz, C.; Camacho, F.; Saavedra, P.; Valenzuela, A.; et al. In Vitro Immunomodulatory Activities of Peptides Derived from Salmo Salar NK-Lysin and Cathelicidin in Fish Cells. Fish Shellfish Immunol. 2019, 88, 587–594. [Google Scholar] [CrossRef]
- Zhou, Q.-J.; Wang, J.; Mao, Y.; Liu, M.; Su, Y.-Q.; Ke, Q.-Z.; Chen, J.; Zheng, W.-Q. Molecular Structure, Expression and Antibacterial Characterization of a Novel Antimicrobial Peptide NK-Lysin from the Large Yellow Croaker Larimichthys crocea. Aquaculture 2019, 500, 315–321. [Google Scholar] [CrossRef]
- Zhang, M.; Li, M.; Sun, L. NKLP27: A Teleost NK-Lysin Peptide That Modulates Immune Response, Induces Degradation of Bacterial DNA, and Inhibits Bacterial and Viral Infection. PLoS ONE 2014, 9, e106543. [Google Scholar] [CrossRef]
- Zhou, Q.-J.; Wang, J.; Liu, M.; Qiao, Y.; Hong, W.-S.; Su, Y.-Q.; Han, K.-H.; Ke, Q.-Z.; Zheng, W.-Q. Identification, Expression and Antibacterial Activities of an Antimicrobial Peptide NK-Lysin from a Marine Fish Larimichthys crocea. Fish Shellfish Immunol. 2016, 55, 195–202. [Google Scholar] [CrossRef]
- Lama, R.; Pereiro, P.; Costa, M.M.; Encinar, J.A.; Medina-Gali, R.M.; Pérez, L.; Lamas, J.; Leiro, J.; Figueras, A.; Novoa, B. Turbot (Scophthalmus maximus) Nk-Lysin Induces Protection against the Pathogenic Parasite Philasterides dicentrarchi via Membrane Disruption. Fish Shellfish Immunol. 2018, 82, 190–199. [Google Scholar] [CrossRef] [Green Version]
- Falco, A.; Medina-Gali, R.; Poveda, J.; Bello-Perez, M.; Novoa, B.; Encinar, J. Antiviral Activity of a Turbot (Scophthalmus maximus) NK-Lysin Peptide by Inhibition of Low-PH Virus-Induced Membrane Fusion. Mar. Drugs 2019, 17, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, F.-F.; Li, C.-H.; Chen, J. Molecular Characterization of the NK-Lysin in a Teleost Fish, Boleophthalmus pectinirostris: Antimicrobial Activity and Immunomodulatory Activity on Monocytes/Macrophages. Fish Shellfish Immunol. 2019, 92, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Dalmo, R.A.; Bøgwald, J. SS-Glucans as Conductors of Immune Symphonies. Fish Shellfish Immunol. 2008, 25, 384–396. [Google Scholar] [CrossRef] [PubMed]
- Petit, J.; Bailey, E.C.; Wheeler, R.T.; de Oliveira, C.A.F.; Forlenza, M.; Wiegertjes, G.F. Studies Into β-Glucan Recognition in Fish Suggests a Key Role for the C-Type Lectin Pathway. Front. Immunol. 2019, 10, 280. [Google Scholar] [CrossRef]
- Derwenskus, F.; Metz, F.; Gille, A.; Schmid-Staiger, U.; Briviba, K.; Schließmann, U.; Hirth, T. Pressurized Extraction of Unsaturated Fatty Acids and Carotenoids from Wet Chlorella vulgaris and Phaeodactylum tricornutum Biomass Using Subcritical Liquids. GCB Bioenergy 2019, 11, 335–344. [Google Scholar] [CrossRef]
- Mylonas, C.C.; Cardinaletti, G.; Sigelaki, I.; Polzonetti-Magni, A. Comparative Efficacy of Clove Oil and 2-Phenoxyethanol as Anesthetics in the Aquaculture of European Sea Bass (Dicentrarchus labrax) and Gilthead Sea Bream (Sparus aurata) at Different Temperatures. Aquaculture 2005, 246, 467–481. [Google Scholar] [CrossRef]
- Raveendran, A.; MV, A.; Philip, R.; Antony, S.P. β-Defensin from the Asian Sea Bass, Lates Calcarifer: Molecular Prediction and Phylogenetic Analysis. Probiotics Antimicro. Prot. 2021, 13, 1798–1807. [Google Scholar] [CrossRef]
- Neelima, S.; Archana, K.; Athira, P.P.; Anju, M.V.; Anooja, V.V.; Bright Singh, I.S.; Philip, R. Molecular Characterization of a Novel β-Defensin Isoform from the Red-Toothed Trigger Fish, Odonus niger (Ruppel, 1836). J. Genet. Eng. Biotechnol. 2021, 19, 71. [Google Scholar] [CrossRef]
- Iqbal, H.M.N. B b High-Value Compounds from Microalgae with Industrial Exploitability Ndash A Review b b. Front. Biosci. 2017, 9, 319–342. [Google Scholar] [CrossRef]
- Schadendorf, D.; Ascierto, P.A.; Haanen, J.; Espinosa, E.; Demidov, L.; Garbe, C.; Guida, M.; Lorigan, P.; Chiarion-Sileni, V.; Gogas, H.; et al. Safety and Efficacy of Nivolumab in Challenging Subgroups with Advanced Melanoma Who Progressed on or after Ipilimumab Treatment: A Single-Arm, Open-Label, Phase II Study (CheckMate 172). Eur. J. Cancer 2019, 121, 144–153. [Google Scholar] [CrossRef]
- Wang, J.; Shen, T.; Wang, Q.; Zhang, T.; Li, L.; Wang, Y.; Fang, Y. The Long-Term Efficacy of Cytokine-Induced Killer Cellular Therapy for Hepatocellular Carcinoma: A Meta-Analysis. Immunotherapy 2019, 11, 1325–1335. [Google Scholar] [CrossRef]
- Dolmatova, L.S.; Dolmatov, I.Y. Tumor-Associated Macrophages as Potential Targets for Anti-Cancer Activity of Marine Invertebrate-Derived Compounds. CPD 2021, 27, 3139–3160. [Google Scholar] [CrossRef]
- Yang, Y. Cancer Immunotherapy: Harnessing the Immune System to Battle Cancer. J. Clin. Investig. 2015, 125, 3335–3337. [Google Scholar] [CrossRef] [Green Version]
- Torres-Tiji, Y.; Fields, F.J.; Mayfield, S.P. Microalgae as a Future Food Source. Biotechnol. Adv. 2020, 41, 107536. [Google Scholar] [CrossRef]
- Matos, J.; Afonso, C.; Cardoso, C.; Serralheiro, M.L.; Bandarra, N.M. Yogurt Enriched with Isochrysis Galbana: An Innovative Functional Food. Foods 2021, 10, 1458. [Google Scholar] [CrossRef]
- Brentuximab Vedotin. Drugs R. D. 2011, 11, 85–95. [CrossRef]
- Deeks, E.D. Polatuzumab Vedotin: First Global Approval. Drugs 2019, 79, 1467–1475. [Google Scholar] [CrossRef] [Green Version]
- Markham, A. Belantamab Mafodotin: First Approval. Drugs 2020, 80, 1607–1613. [Google Scholar] [CrossRef]
- Kazamia, E.; Czesnick, H.; Nguyen, T.T.V.; Croft, M.T.; Sherwood, E.; Sasso, S.; Hodson, S.J.; Warren, M.J.; Smith, A.G. Mutualistic Interactions between Vitamin B12-Dependent Algae and Heterotrophic Bacteria Exhibit Regulation: Algal-Bacterial Interactions for Delivery of Vitamin B12. Environ. Microbiol. 2012, 14, 1466–1476. [Google Scholar] [CrossRef]
- Lutzu, G.A. Interactions of Microalgae and Other Microorganisms for Enhanced Production of High-Value Compounds. Front. Biosci. 2018, 23, 1487–1504. [Google Scholar] [CrossRef] [Green Version]
- Ruocco, N.; Esposito, R.; Bertolino, M.; Zazo, G.; Sonnessa, M.; Andreani, F.; Coppola, D.; Giordano, D.; Nuzzo, G.; Lauritano, C.; et al. A Metataxonomic Approach Reveals Diversified Bacterial Communities in Antarctic Sponges. Mar. Drugs 2021, 19, 173. [Google Scholar] [CrossRef]
- Chen, D.S.; Mellman, I. Oncology Meets Immunology: The Cancer-Immunity Cycle. Immunity 2013, 39, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.S.; Mellman, I. Elements of Cancer Immunity and the Cancer–Immune Set Point. Nature 2017, 541, 321–330. [Google Scholar] [CrossRef]
- Galon, J.; Bruni, D. Tumor Immunology and Tumor Evolution: Intertwined Histories. Immunity 2020, 52, 55–81. [Google Scholar] [CrossRef] [PubMed]
- Lauritano, C.; Ferrante, M.I.; Rogato, A. Marine Natural Products from Microalgae: An—Omics Overview. Marine Drugs 2019, 17, 269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vingiani, G.M.; De Luca, P.; Ianora, A.; Dobson, A.D.W.; Lauritano, C. Microalgal Enzymes with Biotechnological Applications. Mar. Drugs 2019, 17, 459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trivella, D.B.B.; de Felicio, R. The Tripod for Bacterial Natural Product Discovery: Genome Mining, Silent Pathway Induction, and Mass Spectrometry-Based Molecular Networking. mSystems 2018, 3, e00160-17. [Google Scholar] [CrossRef] [Green Version]
- Albarano, L.; Esposito, R.; Ruocco, N.; Costantini, M. Genome Mining as New Challenge in Natural Products Discovery. Mar. Drugs 2020, 18, 199. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Compound/Extract | Activity (Cells) | Organism | Mechanism of Action | Active Concentration | Reference |
|---|---|---|---|---|---|
| α-d-glucan | Immunomodulatory functions by regulating T lymphocytes and dendritic cells in vitro. | Marine fungus Phoma herbarum YS4108 | After 7 days, there was relief in the clinical symptoms of mice with colitis, restoration of intestinal immune homeostasis, and remission of mucosal damage. YCP blocked the overexpression of the pro-inflammatory cytokines IL-6, TNF-α and IL-1β induced by DSS in the colon. | In vivo: 40 mg/kg Intraperitoneal injection | [24,26] |
| Irish deep-sea organism extracts | Induce human mesenchymal stem cell (hMSC) differentiation. | Filamentous fungi | Reduced the production of pro-inflammatory cytokines, such as TNFα and IL-1β. | In vitro: 125 µg/mL | [27] |
| β-carotene | Anti-inflammatory | Microalgae | Docked pro-inflammatory proteins IL-6 with binding energy −7.9 Kcal/mol. | In silico study | [48] |
| Phycocyanobilin | Anti-inflammatory | Microalgae | Docked pro-inflammatory protein NF-κB inducing kinase (NIK) with binding energy −9.9 Kcal/mol. | In silico study | [48] |
| 9-cis-β-carotene | Anti-inflammatory | Microalgae | Docked pro-inflammatory protein TNF-α with binding energy −7.9 Kcal/mol. | In silico study | [48] |
| Exopolysaccharides (EPS) | Immunostimulatory Antibacterial | Microalga Porphyridium cruentum | Increase in total hemocytes (THC) value, phagocytotic activity (PA), and respiratory burst (RB). | Treatment with increasing concentration of EPS | [41] |
| β-glucans | Immunostimulatory, Anti-inflammatory and Antioxidant effects in fishes Promoting the growth of commensal microbiota | Microalga Phaeodactylum tricornutum | Elicited an inflammatory response with the downregulation of pro-inflammatory cytokines IL-1β, IL-6, and TNF-α; activated the host’s innate immune cells; increased activity of SOD and CAT in the intestine and erythrocytes in the blood; and decreased intestinal IL-1β, IL-6, and TNF-α intestinal expression. | 0.6 g β-glucans per kg of feed | [54] |
| Compound/Extract | Activity (Cells) | Organism | Mechanism of Action | Active Concentration | Reference |
|---|---|---|---|---|---|
| Ulvan | Immunomodulatory activity | Alga: Ulva ohnoi | Increases in IL-1β, IL-6, and IL-10; improves LPS-induced inflammation; and a decreases prostaglandin E2. | 100 µg/mL in vitro | [76] |
| Fucoidan | Immunostimulatory activity Pro-proliferative activity | Algae: Ecklonia cava, Macrocystis pyrifera, Undaria pinnatifida, and Fucus vesiculosus | Increases the production of IL-6, IL-12, and TNF-α in MODCs and PBDCs; induces INF-γ production. | 100 µg/mL in vitro | [85] |
| Sulphated polysaccharides: sulfated galactans and pyruvates (named SP1 and SP2) | Immunostimulatory activity | Alga: Caulerpa cupressoides | Increased production of NO, ERO, and the cytokines IL-6 and TNF-α in murine macrophages RAW 264.7. | 100 µg/mL in vitro | [8] |
| Sulphated oligosaccharide(GLSO) | Immunomodulatory activity | Alga: Gracilaria lemaneiformis | Inhibits the production of INF-γ by T cells in ovalbumin (OVA) immunized mice and in vitro activation system of OVA-specific CD4+ T cells; inhibits the activity of mTOR, glycolysis, cell cycle, and DNA replication. | [87] | |
| Crude extract | Immunomodulatory activity | Sponge: Haliclona (Soestellla) sp. | Decrease of immune cells (WBC, lymphocytes, platelets, BMC, and splenocytes) and of the splenocytic index; increase of neutrophil:lymphocyte ratio. | In vivo 15 mg/kg; 10 mg/kg; 5 mg/kg. | [97] |
| ThisAgeliferins derivates | Immunomodulatory activity | Sponge: Astrosclera willeyana | Inhibition of Cbl-b ubiquitin ligase activity (IC50 values ranging from 18 to 35 µM); | 10–50 µM | [99] |
| ASPG-2 polysaccharide | Immunomodulatory activity | Mollusk: Arca subcrenata Lischke | promotes NO secretion; increases phagocytosis in murine RAW 264.7 macrophages and activation of the TLR4-MAPK/Akt-NF-κB signaling pathway. | In vitro: 250 μg/mL and 500 μg/mL for 24 h | [102] |
| Polysaccharides | Immunomodulatory activity Anti-inflammatory activity | Mollusk: Mytilus coruscus | Promotes the abundance of some probiotics in the colon. | In vitro: 300 μg/mL and 600 μg/mL | [103] |
| Protein HPCG2 | Immunomodulatory activity | Mollusk: Scapharca broughtonii | Promotes the phosphorylation of Akt, ERK, and JNK. | In vitro: 250 μg/mL and 500 μg/mL | [104] |
| Polysaccharide PPA | Immunomodulatory activity | Coral: Pseudopterogorgia americana | Induces pro-inflammatory mediator expression in macrophages via ROS-, MAPK-, PKC-α/δ-, and NF-κB-dependent pathways. | In vitro: 10 μg/mL | [117] |
| BpNK-Lysine | Immunomodulatory activity Antibacterial activity | Fish: Beleophthalmus pectinirostris | In vivo, decreased the tissue bacterial burden of mudskipper infected by E. tarda, upregulated the mRNA expression of pro-inflammatory cytokines (IL-1β, TNF-α and IFN-γ), and downregulated the mRNA expression of anti-inflammatory cytokines (IL-10 and TGF-β) in Beleophthalmus pectinirostris. | In vivo: injection of 1.0 µg/g In vitro: 1.0 μg/mL | [139] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montuori, E.; de Pascale, D.; Lauritano, C. Recent Discoveries on Marine Organism Immunomodulatory Activities. Mar. Drugs 2022, 20, 422. https://doi.org/10.3390/md20070422
Montuori E, de Pascale D, Lauritano C. Recent Discoveries on Marine Organism Immunomodulatory Activities. Marine Drugs. 2022; 20(7):422. https://doi.org/10.3390/md20070422
Chicago/Turabian StyleMontuori, Eleonora, Donatella de Pascale, and Chiara Lauritano. 2022. "Recent Discoveries on Marine Organism Immunomodulatory Activities" Marine Drugs 20, no. 7: 422. https://doi.org/10.3390/md20070422
APA StyleMontuori, E., de Pascale, D., & Lauritano, C. (2022). Recent Discoveries on Marine Organism Immunomodulatory Activities. Marine Drugs, 20(7), 422. https://doi.org/10.3390/md20070422