
First Characterization of Ostreopsis cf. ovata (Dinophyceae) and Detection of Ovatoxins during a Multispecific and Toxic Ostreopsis Bloom on French Atlantic Coast

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
2.1. Respiratory Disorders and Cutaneous Irritations Observed during Summer Blooms on the French Basque Coast
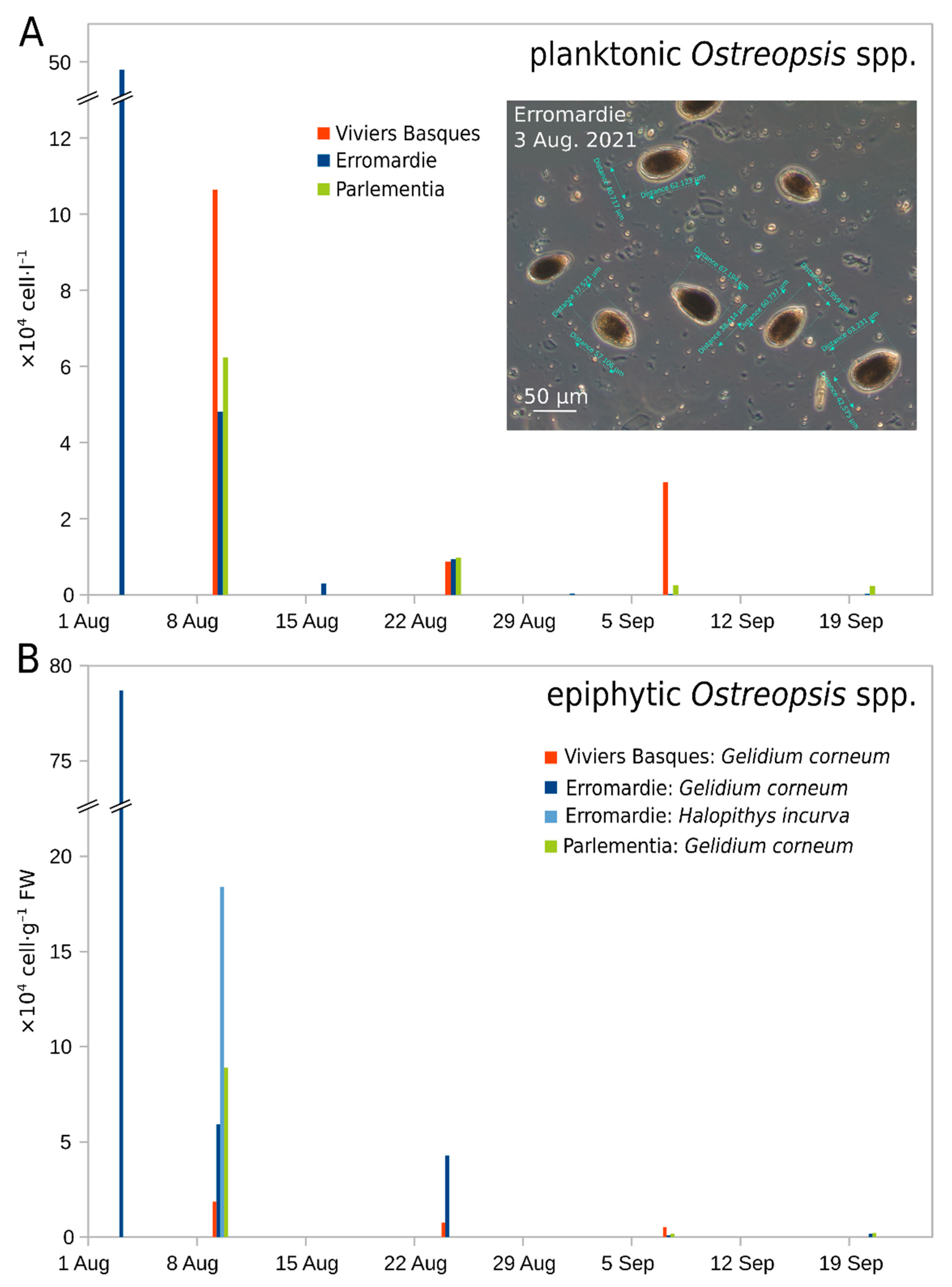
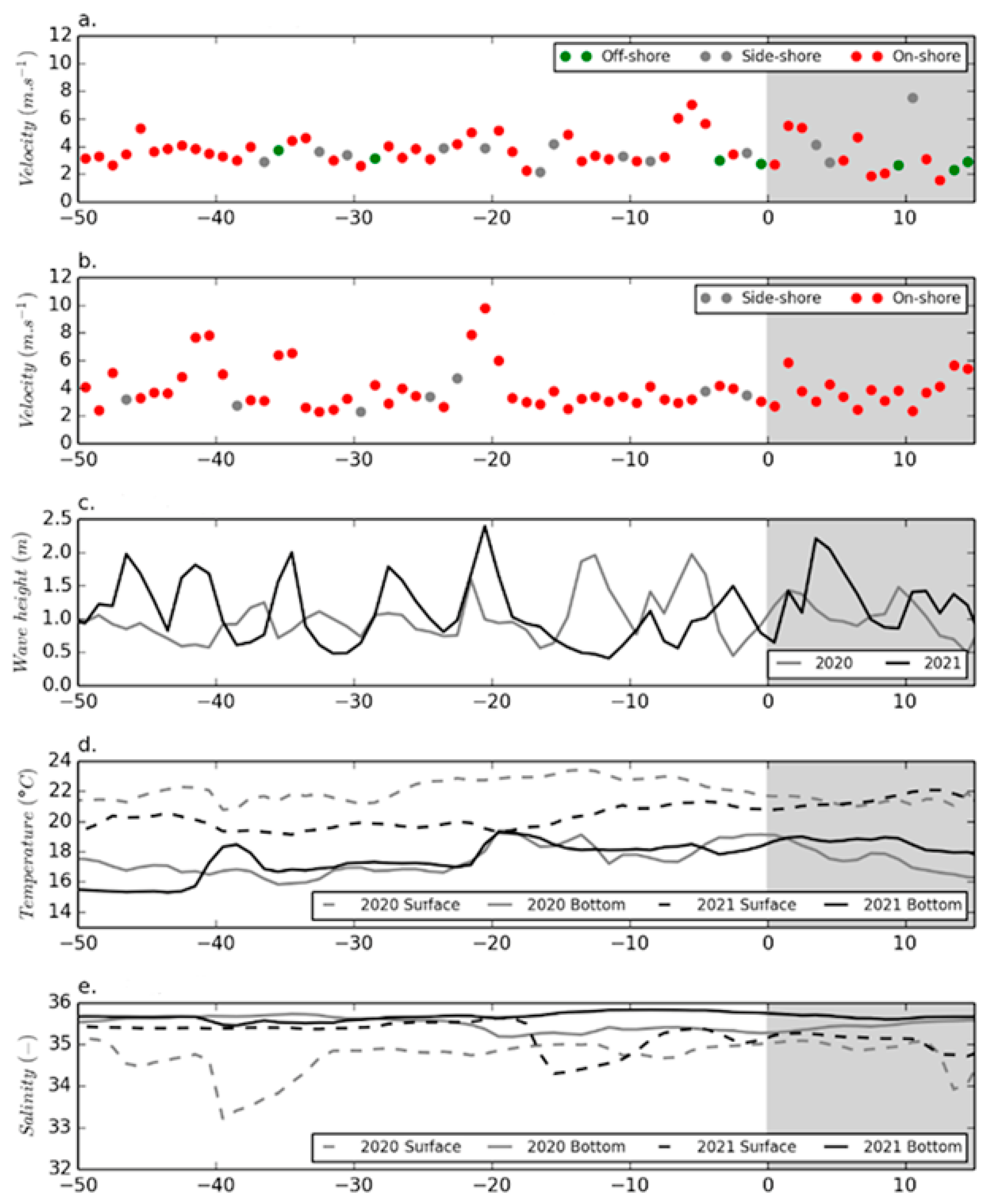
2.2. Environmental Conditions before and during the 2020 and 2021 Events
2.3. Identification of Ostreopsis Species in the Bloom Samples
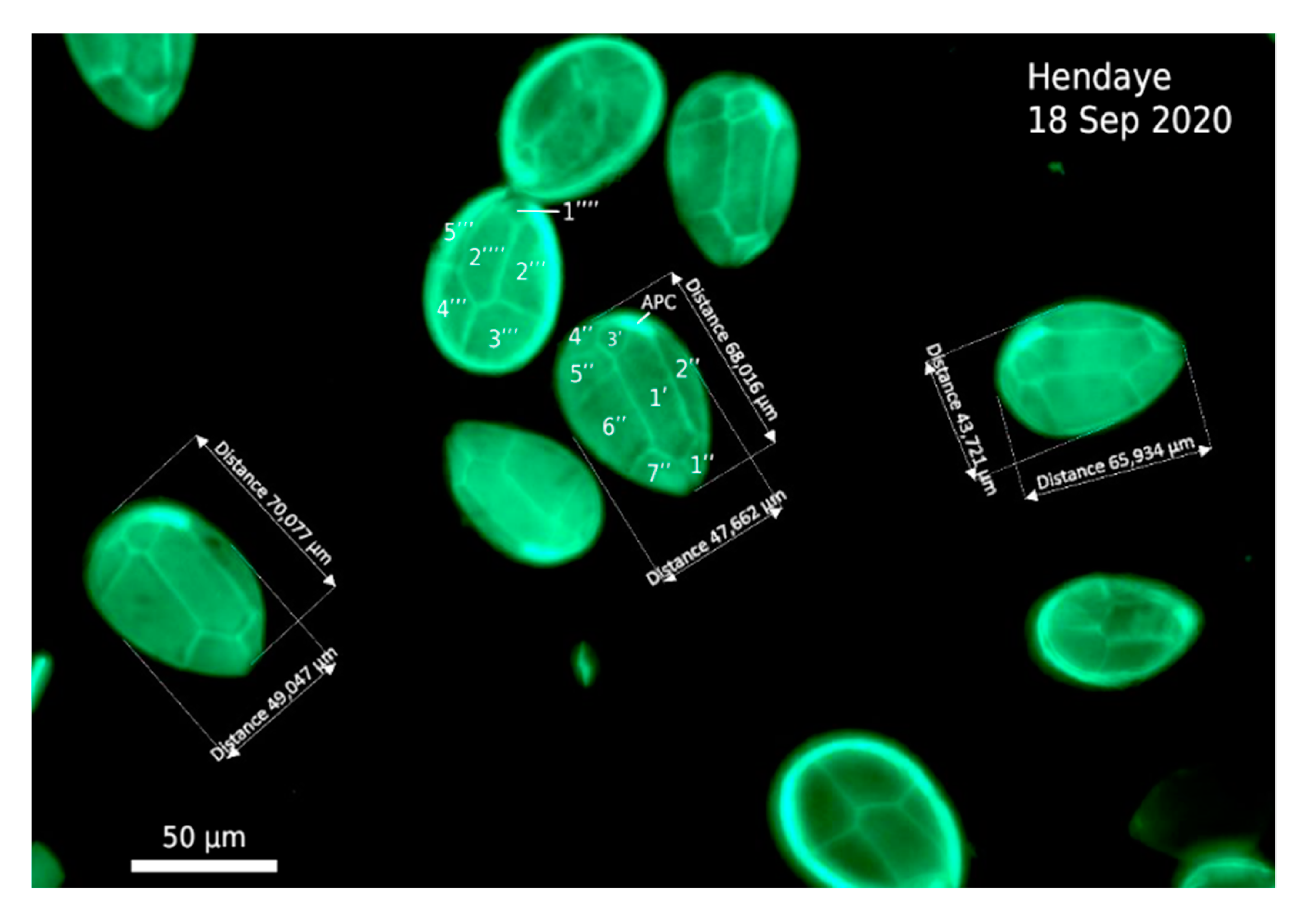
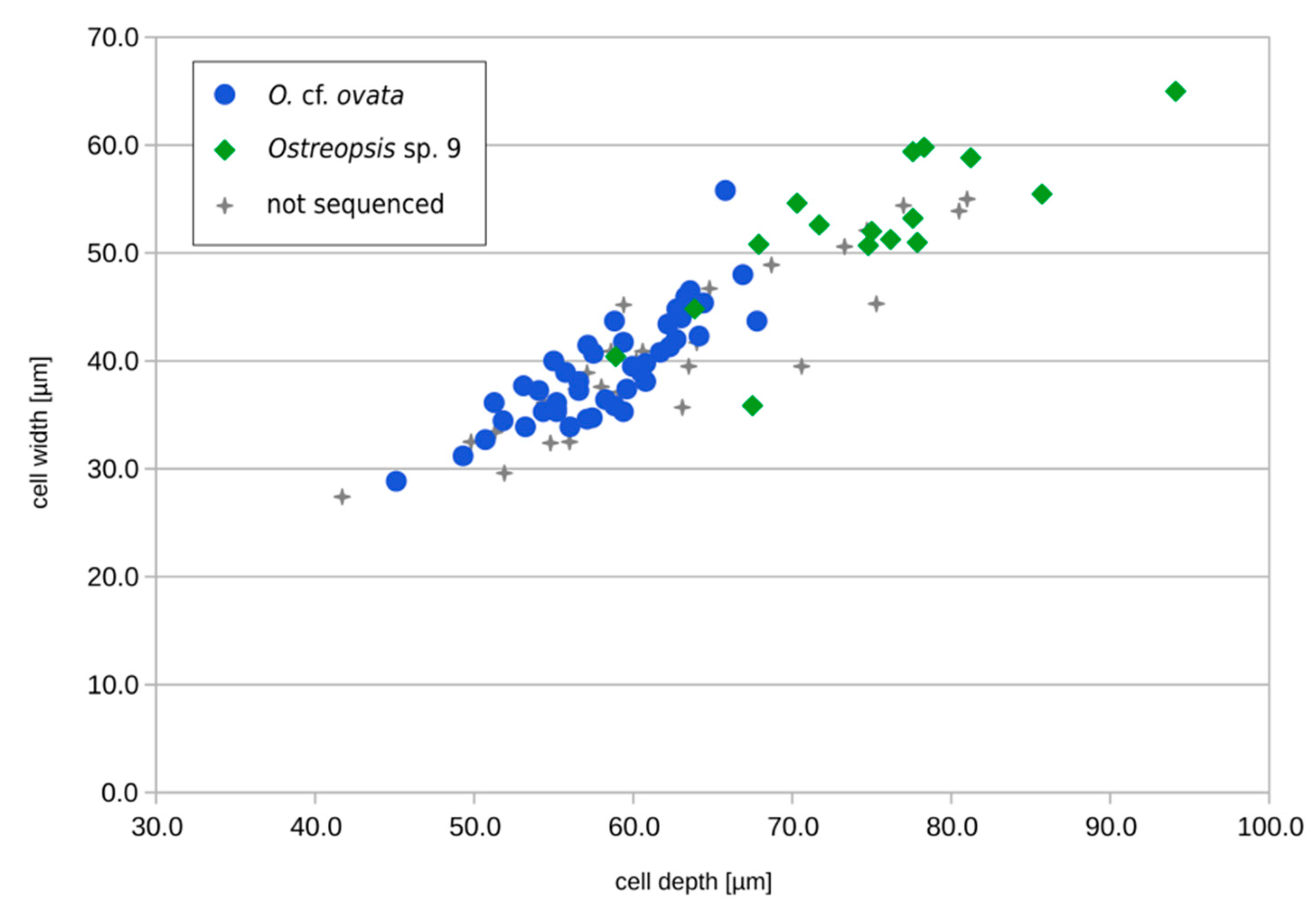
2.3.1. Cell Morphology in the 2020 Bloom Samples
2.3.2. Single Cells Isolated from the 2021 Environmental Sample and Estimation of Relative Abundances of Ostreopsis spp.
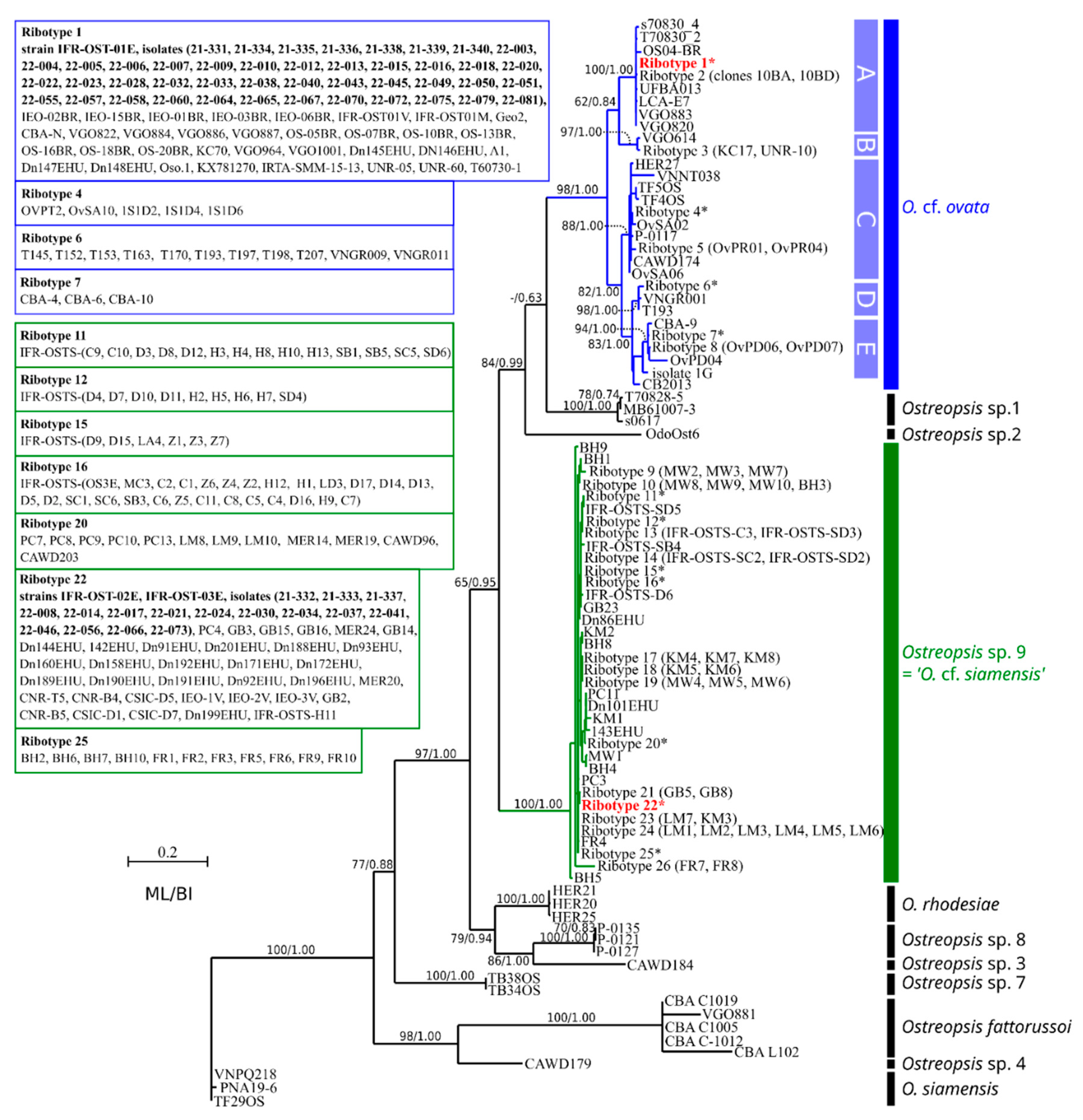
2.3.3. Molecular Phylogeny of Isolated Specimens and Cultivated Strains from the 2021 Bloom
2.4. Toxin Profiles of Ostreopsis Samples
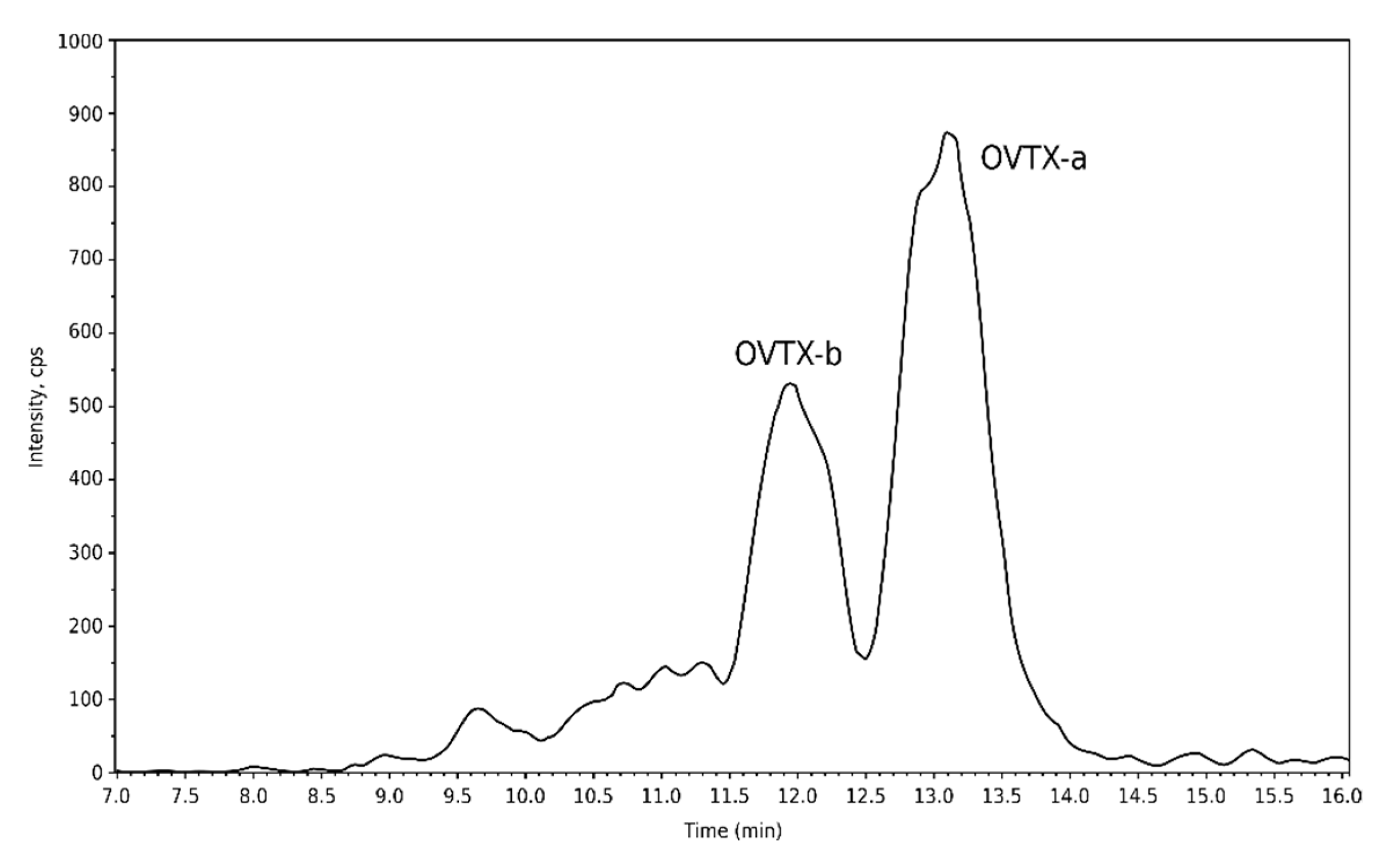
2.4.1. Toxins Detected in the 2021 Environmental Sample from Erromardie (16 August 2021)
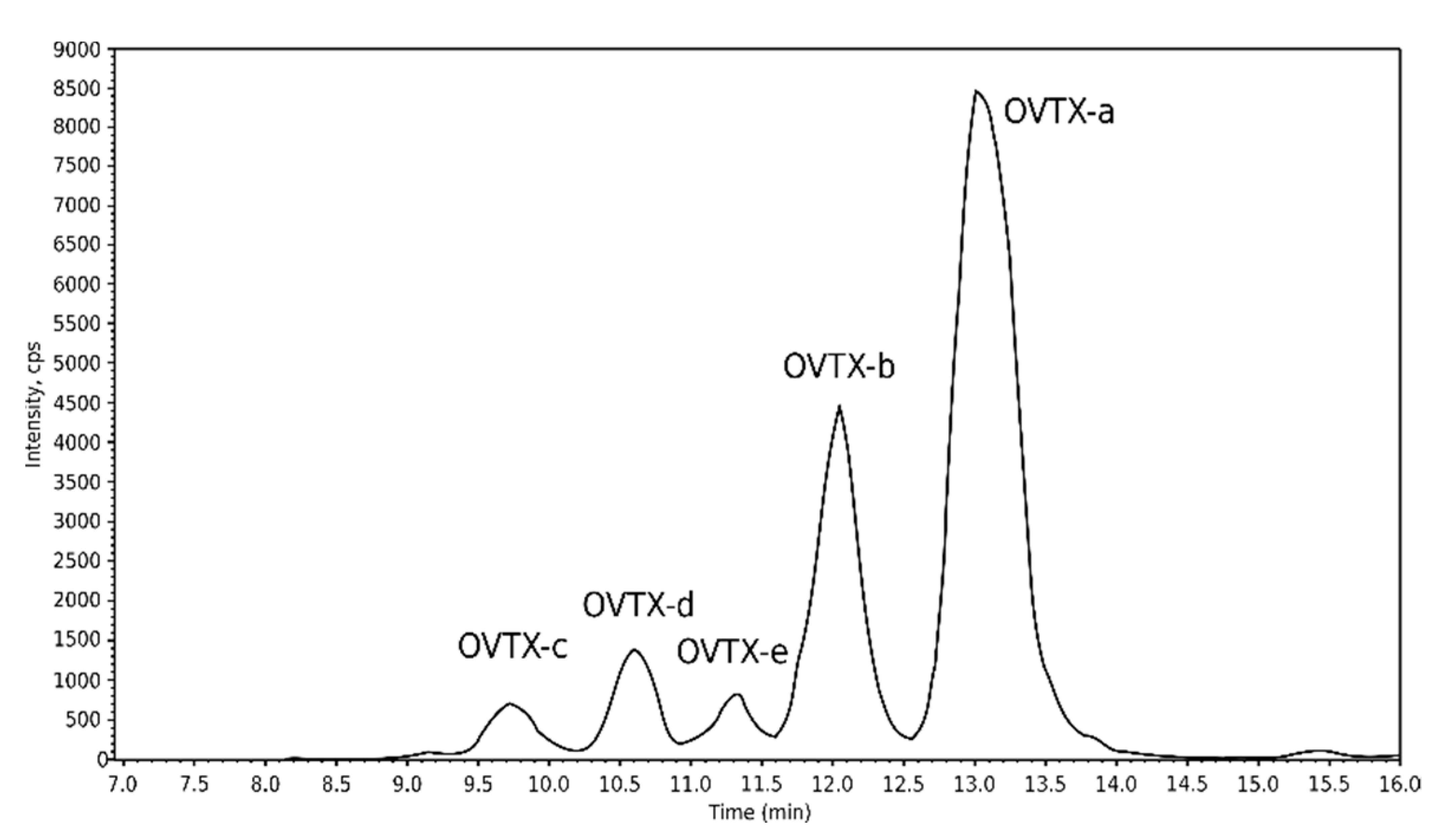
2.4.2. Toxin Profiles of Cultivated Strains
3. Discussion
3.1. Ostreopsis Bloom Events and Impact on Human Health
3.2. Environmental Conditions Associated with the 2020 and 2021 Bloom Events
3.3. Identification of Ostreopsis Species during the 2021 Toxic Event
3.4. Toxin Production by Ostreopsis Species during the 2021 Toxic Event
4. Materials and Methods
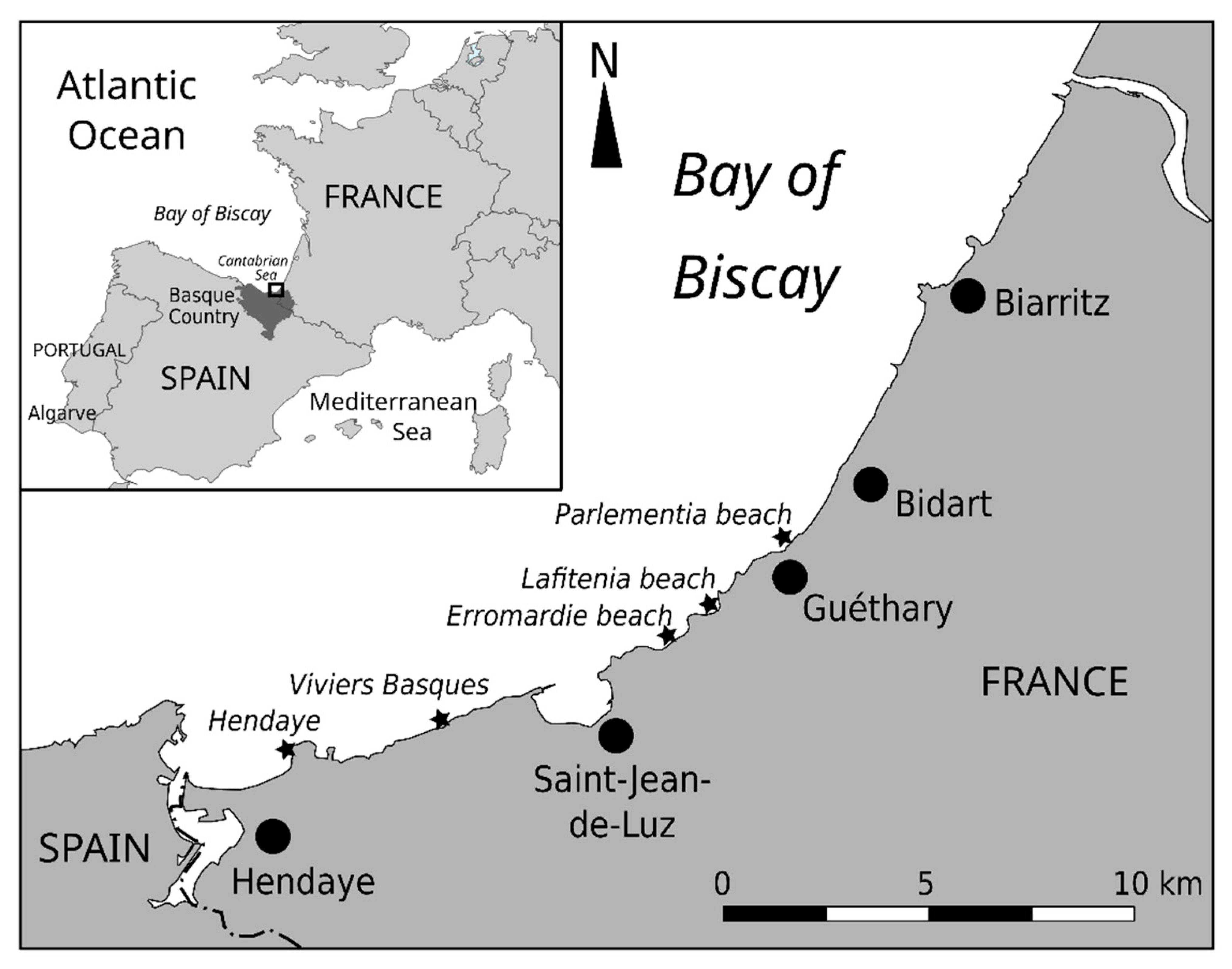
4.1. Sampling on the Basque Coast
4.2. Cell Counts and Estimation of Ostreopsis Abundances
4.3. Environmental Parameters
4.4. Strains Isolation and Cultivation
4.5. Species Identification and Estimation of Their Relative Abundances
4.5.1. Single-Cells Isolation
4.5.2. DNA Extraction from Strains
4.5.3. Amplification and Sequencing
4.5.4. Phylogenetic Analysis
4.6. Toxin Analysis by Liquid Chromatography Tandem Mass Spectrometry (LC–MS/MS)
4.6.1. Sample Preparation
- Environmental sample
- Strains in culture
4.6.2. LC–MS/MS Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schmidt, J. Flora of Koh Chang: Contribution to the Knowledge to the Vegetation in the Gulf of Siam. Part IV. Peridiniales. Bot. Tidsskr. 1901, 24, 212–221. [Google Scholar] [CrossRef]
- Tester, P.A.; Litaker, R.W.; Berdalet, E. Climate Change and Harmful Benthic Microalgae. Harmful Algae 2020, 91, 101655. [Google Scholar] [CrossRef]
- Neves, R.A.F.; Contins, M.; Nascimento, S.M. Effects of the Toxic Benthic Dinoflagellate Ostreopsis cf. ovata on Fertilization and Early Development of the Sea Urchin Lytechinus variegatus. Mar. Environ. Res. 2018, 135, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Pavaux, A.-S.; Rostan, J.; Guidi-Guilvard, L.; Marro, S.; Ternon, E.; Thomas, O.P.; Lemée, R.; Gasparini, S. Effects of the Toxic Dinoflagellate Ostreopsis cf. ovata on Survival, Feeding and Reproduction of a Phytal Harpacticoid Copepod. J. Exp. Mar. Biol. Ecol. 2019, 516, 103–113. [Google Scholar] [CrossRef]
- Tichadou, L.; Glaizal, M.; Armengaud, A.; Grossel, H.; Lemée, R.; Kantin, R.; Lasalle, J.-L.; Drouet, G.; Rambaud, L.; Malfait, P.; et al. Health impact of unicellular algae of the Ostreopsis genus blooms in the mediterranean sea: Experience of the French Mediterranean coast surveillance network from 2006 to 2009. Clin. Toxicol. 2010, 48, 839–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tubaro, A.; Durando, P.; Del Favero, G.; Ansaldi, F.; Icardi, G.; Deeds, J.R.; Sosa, S. Case Definitions for Human Poisonings Postulated to Palytoxins Exposure. Toxicon 2011, 57, 478–495. [Google Scholar] [CrossRef] [PubMed]
- Durando, P.; Ansaldi, F.; Oreste, P.; Moscatelli, P.; Marensi, L.; Grillo, C.; Gasparini, R.; Icardi, G.; Collaborative Group for the Ligurian Syndromic Algal Surveillance. Ostreopsis ovata and Human Health: Epidemiological and Clinical Features of Respiratory Syndrome Outbreaks from a Two-Year Syndromic Surveillance, 2005–2006, in North-West Italy. Euro Surveill. 2007, 12, 3212. [Google Scholar]
- Vila, M.; Abós-Herràndiz, R.; Isern-Fontanet, J.; Àlvarez, J.; Berdalet, E. Establishing the Link between Ostreopsis cf. ovata Blooms and Human Health Impacts Using Ecology and Epidemiology. Sci. Mar. 2016, 80, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Aligizaki, K.; Katikou, P.; Nikolaidis, G.; Panou, A. First Episode of Shellfish Contamination by Palytoxin-like Compounds from Ostreopsis Species (Aegean Sea, Greece). Toxicon 2008, 51, 418–427. [Google Scholar] [CrossRef]
- Amzil, Z.; Sibat, M.; Chomérat, N.; Grossel, H.; Marco-Miralles, F.; Lemée, R.; Nézan, E.; Séchet, V. Ovatoxin-a and Palytoxin Accumulation in Seafood in Relation to Ostreopsis cf. ovata Blooms on the French Mediterranean Coast. Mar. Drugs 2012, 10, 477–496. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication. Available online: http://www.algaebase.org (accessed on 25 May 2022).
- Verma, A.; Hoppenrath, M.; Dorantes-Aranda, J.J.; Harwood, D.T.; Murray, S.A. Molecular and Phylogenetic Characterization of Ostreopsis (Dinophyceae) and the Description of a New Species, Ostreopsis rhodesae sp. nov., from a Subtropical Australian Lagoon. Harmful Algae 2016, 60, 116–130. [Google Scholar] [CrossRef] [PubMed]
- Accoroni, S.; Romagnoli, T.; Penna, A.; Capellacci, S.; Ciminiello, P.; Dell’Aversano, C.; Tartaglione, L.; Abboud-Abi Saab, M.; Giussani, V.; Asnaghi, V.; et al. Ostreopsis fattorussoi Sp. Nov. (Dinophyceae), a New Benthic Toxic Ostreopsis Species from the Eastern Mediterranean Sea. J. Phycol. 2016, 52, 1064–1084. [Google Scholar] [CrossRef] [PubMed]
- Penna, A.; Vila, M.; Fraga, S.; Giacobbe, M.G.; Andreoni, F.; Riobó, P.; Vernesi, C. Characterization of Ostreopsis and Coolia (Dinophyceae) Isolates in the Western Mediterranean Sea Based on Morphology, Toxicity and Internal Transcribed Spacer 5.8S RDNA Sequences. J. Phycol. 2005, 41, 212–225. [Google Scholar] [CrossRef]
- Parsons, M.L.; Aligizaki, K.; Dechraoui Bottein, M.-Y.; Fraga, S.; Morton, S.L.; Penna, A.; Rhodes, L. Gambierdiscus and Ostreopsis: Reassessment of the State of Knowledge of Their Taxonomy, Geography, Ecophysiology, and Toxicology. Harmful Algae 2012, 14, 107–129. [Google Scholar] [CrossRef]
- Chomérat, N.; Bilien, G.; Derrien, A.; Henry, K.; Ung, A.; Viallon, J.; Darius, H.T.; Mahana iti Gatti, C.; Roué, M.; Hervé, F.; et al. Ostreopsis lenticularis Y. Fukuyo (Dinophyceae, Gonyaulacales) from French Polynesia (South Pacific Ocean): A Revisit of Its Morphology, Molecular Phylogeny and Toxicity. Harmful Algae 2019, 84, 95–111. [Google Scholar] [CrossRef] [Green Version]
- Hoppenrath, M.; Murray, S.; Chomérat, N.; Horiguchi, T. Marine Benthic Dinoflagellates—Unveiling Their Worldwide Biodiversity (Kleine Senckenberg-Reihe 54); E. Schweizerbart’sche Verlagbuchhandlung: Stuttgart, Germany, 2014. [Google Scholar]
- Chomérat, N.; Bilien, G.; Viallon, J.; Hervé, F.; Réveillon, D.; Henry, K.; Zubia, M.; Vieira, C.; Ung, A.; Gatti, C.M.; et al. Taxonomy and Toxicity of a Bloom-Forming Ostreopsis Species (Dinophyceae, Gonyaulacales) in Tahiti Island (South Pacific Ocean): One Step Further towards Resolving the Identity of O. siamensis. Harmful Algae 2020, 98, 101888. [Google Scholar] [CrossRef]
- Nguyen-Ngoc, L.; Doan-Nhu, H.; Larsen, J.; Phan-Tan, L.; Nguyen, X.; Lundholm, N.; Van Chu, T.; Huynh-Thi, D.N. Morphological and Genetic Analyses of Ostreopsis (Dinophyceae, Gonyaulacales, Ostreopsidaceae) Species from Vietnamese Waters with a Re-description of the Type Species, O. siamensis. J. Phycol. 2021, 57, 1059–1083. [Google Scholar] [CrossRef]
- Boisnoir, A.; Bilien, G.; Lemée, R.; Chomérat, N. First Insights on the Diversity of the Genus Ostreopsis (Dinophyceae, Gonyaulacales) in Guadeloupe Island, with Emphasis on the Phylogenetic Position of O. heptagona. Eur. J. Protistol. 2022, 83, 125875. [Google Scholar] [CrossRef]
- Sato, S.; Nishimura, T.; Uehara, K.; Sakanari, H.; Tawong, W.; Hariganeya, N.; Smith, K.; Rhodes, L.; Yasumoto, T.; Taira, Y.; et al. Phylogeography of Ostreopsis along West Pacific Coast, with Special Reference to a Novel Clade from Japan. PLoS ONE 2011, 6, e27983. [Google Scholar] [CrossRef]
- Tawong, W.; Nishimura, T.; Sakanari, H.; Sato, S.; Yamaguchi, H.; Adachi, M. Distribution and Molecular Phylogeny of the Dinoflagellate Genus Ostreopsis in Thailand. Harmful Algae 2014, 37, 160–171. [Google Scholar] [CrossRef]
- Chomérat, N.; Bilien, G.; Couté, A.; Quod, J.-P. Reinvestigation of Ostreopsis mascarenensis Quod (Dinophyceae, Gonyaulacales) from Réunion Island (SW Indian Ocean): Molecular Phylogeny and Emended Description. Phycologia 2020, 59, 140–153. [Google Scholar] [CrossRef]
- Poli, M.; Ruiz-Olvera, P.; Nalca, A.; Ruiz, S.; Livingston, V.; Frick, O.; Dyer, D.; Schellhase, C.; Raymond, J.; Kulis, D.; et al. Toxicity and Pathophysiology of Palytoxin Congeners after Intraperitoneal and Aerosol Administration in Rats. Toxicon 2018, 150, 235–250. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.E.; Scheuer, P.J. Palytoxin: A New Marine Toxin from a Coelenterate. Science 1971, 172, 495–498. [Google Scholar] [CrossRef] [PubMed]
- Medina-Pérez, N.I.; Dall’Osto, M.; Decesari, S.; Paglione, M.; Moyano, E.; Berdalet, E. Aerosol Toxins Emitted by Harmful Algal Blooms Susceptible to Complex Air–Sea Interactions. Environ. Sci. Technol. 2021, 55, 468–477. [Google Scholar] [CrossRef] [PubMed]
- Ciminiello, P.; Dell’Aversano, C.; Iacovo, E.D.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Benedettini, G.; Onorari, M.; Serena, F.; Battocchi, C.; et al. First Finding of Ostreopsis cf. ovata Toxins in Marine Aerosols. Environ. Sci. Technol. 2014, 48, 3532–3540. [Google Scholar] [CrossRef]
- Casabianca, S.; Casabianca, A.; Riobó, P.; Franco, J.M.; Vila, M.; Penna, A. Quantification of the Toxic Dinoflagellate Ostreopsis spp. by QPCR Assay in Marine Aerosol. Environ. Sci. Technol. 2013, 47, 3788–3795. [Google Scholar] [CrossRef]
- Ninčević Gladan, Ž.; Arapov, J.; Casabianca, S.; Penna, A.; Honsell, G.; Brovedani, V.; Pelin, M.; Tartaglione, L.; Sosa, S.; Dell’Aversano, C.; et al. Massive Occurrence of the Harmful Benthic Dinoflagellate Ostreopsis cf. ovata in the Eastern Adriatic Sea. Toxins 2019, 11, 300. [Google Scholar] [CrossRef] [Green Version]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Forino, M.; Magno, G.S.; Tartaglione, L.; Grillo, C.; Melchiorre, N. The Genoa 2005 Outbreak. Determination of Putative Palytoxin in Mediterranean Ostreopsis ovata by a New Liquid Chromatography Tandem Mass Spectrometry Method. Anal. Chem. 2006, 78, 6153–6159. [Google Scholar] [CrossRef]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Grillo, C.; Melchiorre, N. Putative Palytoxin and Its New Analogue, Ovatoxin-a, in Ostreopsis ovata Collected along the Ligurian Coasts during the 2006 Toxic Outbreak. J. Am. Soc. Mass Spectrom. 2008, 19, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; Grauso, L.; Tartaglione, L.; Guerrini, F.; Pezzolesi, L.; Pistocchi, R.; et al. Isolation and Structure Elucidation of Ovatoxin-a, the Major Toxin Produced by Ostreopsis ovata. J. Am. Chem. Soc. 2012, 134, 1869–1875. [Google Scholar] [CrossRef]
- Ciminiello, P.; Dell’Aversano, C.; Iacovo, E.D.; Fattorusso, E.; Forino, M.; Grauso, L.; Tartaglione, L.; Guerrini, F.; Pistocchi, R. Complex Palytoxin-like Profile of Ostreopsis ovata. Identification of Four New Ovatoxins by High-Resolution Liquid Chromatography/Mass Spectrometry. Rapid Commun. Mass Spectrom. 2010, 24, 2735–2744. [Google Scholar] [CrossRef] [PubMed]
- Ciminiello, P.; Dell’Aversano, C.; Iacovo, E.D.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Battocchi, C.; Crinelli, R.; Carloni, E.; Magnani, M.; et al. Unique Toxin Profile of a Mediterranean Ostreopsis cf. ovata Strain: HR LC-MS n Characterization of Ovatoxin-f, a New Palytoxin Congener. Chem. Res. Toxicol. 2012, 25, 1243–1252. [Google Scholar] [CrossRef] [PubMed]
- García-Altares, M.; Tartaglione, L.; Dell’Aversano, C.; Carnicer, O.; de la Iglesia, P.; Forino, M.; Diogène, J.; Ciminiello, P. The Novel Ovatoxin-g and Isobaric Palytoxin (so Far Referred to as Putative Palytoxin) from Ostreopsis cf. ovata (NW Mediterranean Sea): Structural Insights by LC-High Resolution MS. Anal. Bioanal. Chem. 2015, 407, 1191–1204. [Google Scholar] [CrossRef] [PubMed]
- Brissard, C.; Hervé, F.; Sibat, M.; Séchet, V.; Hess, P.; Amzil, Z.; Herrenknecht, C. Characterization of Ovatoxin-h, a New Ovatoxin Analog, and Evaluation of Chromatographic Columns for Ovatoxin Analysis and Purification. J. Chromatogr. A 2015, 1388, 87–101. [Google Scholar] [CrossRef] [Green Version]
- Tartaglione, L.; Dello Iacovo, E.; Mazzeo, A.; Casabianca, S.; Ciminiello, P.; Penna, A.; Dell’Aversano, C. Variability in Toxin Profiles of the Mediterranean Ostreopsis cf. ovata and in Structural Features of the Produced Ovatoxins. Environ. Sci. Technol. 2017, 51, 13920–13928. [Google Scholar] [CrossRef]
- Guerrini, F.; Pezzolesi, L.; Feller, A.; Riccardi, M.; Ciminiello, P.; Dell’Aversano, C.; Tartaglione, L.; Iacovo, E.D.; Fattorusso, E.; Forino, M.; et al. Comparative Growth and Toxin Profile of Cultured Ostreopsis ovata from the Tyrrhenian and Adriatic Seas. Toxicon 2010, 55, 211–220. [Google Scholar] [CrossRef]
- Brissard, C.; Herrenknecht, C.; Séchet, V.; Hervé, F.; Pisapia, F.; Harcouet, J.; Lemée, R.; Chomérat, N.; Hess, P.; Amzil, Z. Complex Toxin Profile of French Mediterranean Ostreopsis cf. ovata Strains, Seafood Accumulation and Ovatoxins Prepurification. Mar. Drugs 2014, 12, 2851–2876. [Google Scholar] [CrossRef] [Green Version]
- Tartaglione, L.; Mazzeo, A.; Dell’Aversano, C.; Forino, M.; Giussani, V.; Capellacci, S.; Penna, A.; Asnaghi, V.; Faimali, M.; Chiantore, M.; et al. Chemical, Molecular, and Eco-Toxicological Investigation of Ostreopsis sp. from Cyprus Island: Structural Insights into Four New Ovatoxins by LC-HRMS/MS. Anal. Bioanal. Chem. 2016, 408, 915–932. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Watanabe, R.; Uchida, H.; Matsushima, R.; Nagai, H.; Yasumoto, T.; Yoshimatsu, T.; Sato, S.; Adachi, M. LC-MS/MS Analysis of Novel Ovatoxin Isomers in Several Ostreopsis Strains Collected in Japan. Harmful Algae 2012, 20, 81–91. [Google Scholar] [CrossRef]
- Uchida, H.; Taira, Y.; Yasumoto, T. Structural Elucidation of Palytoxin Analogs Produced by the Dinoflagellate Ostreopsis ovata IK2 Strain by Complementary Use of Positive and Negative Ion Liquid Chromatography/Quadrupole Time-of-Flight Mass Spectrometry: Structural Elucidation of Ovatoxin-a, -d, -e IK2 by LC/QTOFMS. Rapid Commun. Mass Spectrom. 2013, 27, 1999–2008. [Google Scholar] [CrossRef]
- Usami, M.; Satake, M.; Ishida, S.; Inoue, A.; Kan, Y.; Yasumoto, T. Palytoxin Analogs from the Dinoflagellate Ostreopsis siamensis. J. Am. Chem. Soc. 1995, 117, 5389–5390. [Google Scholar] [CrossRef]
- Ukena, T.; Satake, M.; Usami, M.; Oshima, Y.; Naoki, H.; Fujita, T.; Kan, Y.; Yasumoto, T. Structure Elucidation of Ostreocin D, a Palytoxin Analog Isolated from the Dinoflagellate Ostreopsis siamensis. Biosci. Biotechnol. Biochem. 2001, 65, 2585–2588. [Google Scholar] [CrossRef] [Green Version]
- Ukena, T.; Satake, M.; Usami, M.; Oshima, Y.; Fujita, T.; Naoki, H.; Yasumoto, T. Structural Confirmation of Ostreocin-D by Application of Negative-Ion Fast-Atom Bombardment Collision-Induced Dissociation Tandem Mass Spectrometric Methods. Rapid Commun. Mass Spectrom. 2002, 16, 2387–2393. [Google Scholar] [CrossRef] [PubMed]
- Terajima, T.; Uchida, H.; Abe, N.; Yasumoto, T. Structure Elucidation of Ostreocin-A and Ostreocin-E1, Novel Palytoxin Analogs Produced by the Dinoflagellate Ostreopsis siamensis, Using LC/Q-TOF MS. Biosci. Biotech. Bioch. 2018, 83, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Terajima, T.; Uchida, H.; Abe, N.; Yasumoto, T. Simple Structural Elucidation of Ostreocin-B, a New Palytoxin Congener Isolated from the Marine Dinoflagellate Ostreopsis siamensis, Using Complementary Positive and Negative Ion Liquid Chromatography/Quadrupole Time-of-Flight Mass Spectrometry. Rapid Commun. Mass Spectrom. 2018, 32, 1001–1007. [Google Scholar] [CrossRef]
- Hoffmann, K.; Hermanns-Clausen, M.; Buhl, C.; Büchler, M.W.; Schemmer, P.; Mebs, D.; Kauferstein, S. A Case of Palytoxin Poisoning Due to Contact with Zoanthid Corals through a Skin Injury. Toxicon 2008, 51, 1535–1537. [Google Scholar] [CrossRef]
- Tartaglione, L.; Dell’Aversano, C.; Mazzeo, A.; Forino, M.; Wieringa, A.; Ciminiello, P. Determination of Palytoxins in Soft Coral and Seawater from a Home Aquarium. Comparison between Palythoa- and Ostreopsis-Related Inhalatory Poisonings. Environ. Sci. Technol. 2016, 50, 1023–1030. [Google Scholar] [CrossRef] [Green Version]
- Beau, N.; Allemand, D.; Molinatti, E.; Claessens, Y.-E. Intoxication à la palytoxine ou la légende de Limu-make-o’hana. Ann. Fr. Med. Urgence 2017, 7, 52–54. [Google Scholar] [CrossRef]
- Thakur, L.K.; Jha, K.K. Palytoxin-Induced Acute Respiratory Failure. Respir. Med. Case Rep. 2017, 20, 4–6. [Google Scholar] [CrossRef] [Green Version]
- Calon, T.; Sinno-Tellier, S.; de Haro, L.; Bloch, J. Exposition à la palytoxine des personnes manipulant des coraux mous d’aquarium d’eau de mer: Étude des cas rapportés au réseau des Centres antipoison de 2000 à 2017. Toxicol. Anal. Clin. 2019, 31, 64–76. [Google Scholar] [CrossRef]
- Schulz, M.; Łoś, A.; Szabelak, A.; Strachecka, A. Inhalation Poisoning with Palytoxin from Aquarium Coral: Case Description and Safety Advice. Arch. Ind. Hyg. Toxicol. 2019, 70, 14–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhry, N.L.; Przybek, J.; Hamilton, A.; Carley, F. Unique Case of Palytoxin-Related Keratitis. Clin. Exp. Ophthalmol. 2016, 44, 853–854. [Google Scholar] [CrossRef] [PubMed]
- Accoroni, S.; Totti, C. The Toxic Benthic Dinoflagellates of the Genus Ostreopsis in Temperate Areas: A Review. Adv. Oceanogr. Limnol. 2016, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Brescianini, C.; Grillo, C.; Melchiorre, N.; Bertolotto, R.; Ferrari, A.; Vivaldi, B.; Icardi, G.; Gramaccioni, L.; Funari, E.; Scardala, S. Ostreopsis ovata Algal Blooms Affecting Human Health in Genova, Italy, 2005 and 2006. Euro Surveill. 2006, 11, 3040. [Google Scholar] [CrossRef]
- Barroso García, P.; de la Puerta, P.R.; Parrón Carreño, T.; Marín Martínez, P.; Guillén Enríquez, J. Brote con síntomas respiratorios en la provincia de Almería por una posible exposición a microalgas tóxicas. Gac. Sanit. 2008, 22, 578–584. [Google Scholar] [CrossRef] [Green Version]
- Illoul, H.; Hernández, F.R.; Vila, M.; Adjas, N.; Younes, A.A.; Bournissa, M.; Koroghli, A.; Marouf, N.; Rabia, S.; Ameur, F.L.K. The Genus Ostreopsis along the Algerian Coastal Waters (SW Mediterranean Sea) Associated with a Human Respiratory Intoxication Episode. Cryptog. Algol. 2012, 33, 209–216. [Google Scholar] [CrossRef]
- Penna, A.; Fraga, S.; Battocchi, C.; Casabianca, S.; Giacobbe, M.G.; Riobó, P.; Vernesi, C. A Phylogeographical Study of the Toxic Benthic Dinoflagellate Genus Ostreopsis Schmidt. J. Biogeogr. 2010, 37, 830–841. [Google Scholar] [CrossRef] [Green Version]
- Tibiriçá, C.E.J.A.; Leite, I.P.; Batista, T.V.V.; Fernandes, L.F.; Chomérat, N.; Herve, F.; Hess, P.; Mafra, L.L. Ostreopsis cf. ovata Bloom in Currais, Brazil: Phylogeny, Toxin Profile and Contamination of Mussels and Marine Plastic Litter. Toxins 2019, 11, 446. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, S.M.; Neves, R.A.F.; De’Carli, G.A.L.; Borsato, G.T.; da Silva, R.A.F.; Melo, G.A.; de Morais, A.M.; Cockell, T.C.; Fraga, S.; Menezes-Salgueiro, A.D.; et al. Ostreopsis cf. ovata (Dinophyceae) Molecular Phylogeny, Morphology, and Detection of Ovatoxins in Strains and Field Samples from Brazil. Toxins 2020, 12, 70. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.; Brotas, V.; Orive, E.; Neto, A. First Records of Ostreopsis heptagona, O. cf. siamensis and O. cf. ovata—In the Azores Archipelago, Portugal. Harmful Algae News 2010, 42, 1–2. Available online: https://hdl.handle.net/10400.3/1200 (accessed on 10 July 2022).
- Bennouna, A.; El Attar, J.; Abouabdellah, R.; Palma, S.; Penna, A.; Moita, M.T. First Records of Ostreopsis cf. Siamensis in Moroccan Atlantic Upwelling Waters. Harmful Algae News 2010, 42, 1–3. Available online: https://hdl.handle.net/11576/2503674 (accessed on 10 July 2022).
- David, H.; Nascimento, P.; Melo, R.; Amorim, A.; Caeiro, M.F. Bloom of Ostreopsis cf. siamensis in Lisbon Bay. Harmful Algae News 2018, 60, 11–12. [Google Scholar]
- Amorim, A.; Veloso, V.; Penna, A. First Detection of Ostreopsis cf. siamensis in Portuguese Coastal Waters. Harmful Algae News 2010, 42, 6–7. Available online: https://hdl.handle.net/11576/2503669 (accessed on 10 July 2022).
- Laza-Martínez, A.; Orive, E.; Irati, M. Morphological and Genetic Characterization of Benthic Dinoflagellates of the Genera Coolia, Ostreopsis and Prorocentrum from South-Eastern Bay of Biscay. Eur. J. Phycol. 2011, 46, 45–65. [Google Scholar] [CrossRef]
- David, H.; Laza-Martínez, A.; Miguel, I.; Orive, E. Ostreopsis cf. siamensis and Ostreopsis cf. ovata from the Atlantic Iberian Peninsula: Morphological and Phylogenetic Characterization. Harmful Algae 2013, 30, 44–55. [Google Scholar] [CrossRef]
- Ramos, V.; Salvi, D.; Machado, J.; Vale, M.; Azevedo, J.; Vasconcelos, V. Culture-Independent Study of the Late-Stage of a Bloom of the Toxic Dinoflagellate Ostreopsis cf. ovata: Preliminary Findings Suggest Genetic Differences at the Sub-Species Level and Allow ITS2 Structure Characterization. Toxins 2015, 7, 2514–2533. [Google Scholar] [CrossRef] [Green Version]
- Santos, M.; Oliveira, P.B.; Moita, M.T.; David, H.; Caeiro, M.F.; Zingone, A.; Amorim, A.; Silva, A. Ocurrence of Ostreopsis in Two Temperate Coastal Bays (SW Iberia): Insights from the Plankton. Harmful Algae 2019, 86, 20–36. [Google Scholar] [CrossRef]
- Seoane, S.; Siano, R. New Limits of Ostreopsis Distribution in the Bay of Biscay: A First Report of Ostreopsis in Santander Bay, Cantabria (Northern Spain). Harmful Algae News 2018, 60, 10. Available online: https://unesdoc.unesco.org/ark:/48223/pf0000266184 (accessed on 10 July 2022).
- Drouet, K.; Jauzein, C.; Herviot-Heath, D.; Hariri, S.; Laza-Martinez, A.; Lecadet, C.; Plus, M.; Seoane, S.; Sourisseau, M.; Lemée, R.; et al. Current Distribution and Potential Expansion of the Harmful Benthic Dinoflagellate Ostreopsis cf. Siamensis towards the Warming Waters of the Bay of Biscay, North-East Atlantic. Environ. Microbiol. 2021, 23, 4965–4979. [Google Scholar] [CrossRef]
- Deeds, J.R.; Schwartz, M.D. Human Risk Associated with Palytoxin Exposure. Toxicon 2010, 56, 150–162. [Google Scholar] [CrossRef] [Green Version]
- Pelin, M.; Sosa, S.; Ponti, C.; Gibellini, D.; Florio, C.; Tubaro, A. Pro-Inflammatory Effects of Palytoxin on Skin Keratinocytes and Indirect Modulation of Inflammatory Cells. Toxicon 2018, 149, 97. [Google Scholar] [CrossRef]
- Mangialajo, L.; Ganzin, N.; Accoroni, S.; Asnaghi, V.; Blanfuné, A.; Cabrini, M.; Cattaneo-Vietti, R.; Chavanon, F.; Chiantore, M.; Cohu, S.; et al. Trends in Ostreopsis Proliferation along the Northern Mediterranean Coasts. Toxicon 2011, 57, 408–420. [Google Scholar] [CrossRef] [Green Version]
- Pistocchi, R.; Pezzolesi, L.; Guerrini, F.; Vanucci, S.; Dell’Aversano, C.; Fattorusso, E. A Review on the Effects of Environmental Conditions on Growth and Toxin Production of Ostreopsis ovata. Toxicon 2011, 57, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Drouet, K.; Jauzein, C.; Gasparini, S.; Pavaux, A.-S.; Berdalet, E.; Marro, S.; Davenet-Sbirrazuoli, V.; Siano, R.; Lemée, R. The Benthic Toxic Dinoflagellate Ostreopsis cf. ovata in the NW Mediterranean Sea: Relationship between Sea Surface Temperature and Bloom Phenology. Harmful Algae 2022, 112, 102184. [Google Scholar] [CrossRef] [PubMed]
- David, H.; Ganzedo, U.; Laza-Martínez, A.; Orive, E. Relationships between the Presence of Ostreopsis (Dinophyceae) in the Atlantic Coast of the Iberian Peninsula and Sea-Surface Temperature. Cryptog. Algol. 2012, 33, 199–207. [Google Scholar] [CrossRef]
- Gémin, M.-P.; Réveillon, D.; Hervé, F.; Pavaux, A.-S.; Tharaud, M.; Séchet, V.; Bertrand, S.; Lemée, R.; Amzil, Z. Toxin Content of Ostreopsis cf. ovata Depends on Bloom Phases, Depth and Macroalgal Substrate in the NW Mediterranean Sea. Harmful Algae 2020, 92, 101727. [Google Scholar] [CrossRef]
- Nascimento, S.M.; Corrêa, E.V.; Menezes, M.; Varela, D.; Paredes, J.; Morris, S. Growth and Toxin Profile of Ostreopsis cf. ovata (Dinophyta) from Rio de Janeiro, Brazil. Harmful Algae 2012, 13, 1–9. [Google Scholar] [CrossRef]
- Ciminiello, P.; Dell’Aversano, C.; Iacovo, E.D.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Yasumoto, T.; Battocchi, C.; Giacobbe, M.; Amorim, A.; et al. Investigation of Toxin Profile of Mediterranean and Atlantic Strains of Ostreopsis cf. siamensis (Dinophyceae) by Liquid Chromatography–High Resolution Mass Spectrometry. Harmful Algae 2013, 23, 19–27. [Google Scholar] [CrossRef]
- Cagide, E.; Louzao, M.C.; Espiña, B.; Vieytes, M.R.; Jaen, D.; Maman, L.; Yasumoto, T.; Botana, L.M. Production of Functionally Active Palytoxin-like Compounds by Mediterranean Ostreopsis cf. siamensis. Cell Physiol. Biochem. 2009, 23, 431–440. [Google Scholar] [CrossRef]
- Drouet, K. Impact de la température sur la Biogéographie et la Phénologie des Dinoflagellés Benthiques Toxiques du Genre Ostreopsis en Méditerranée et en Atlantique. Ph.D. Thesis, Sorbonne University, Paris, France, 2021. [Google Scholar]
- Verma, A.; Hoppenrath, M.; Harwood, T.; Brett, S.; Rhodes, L.; Murray, S. Molecular Phylogeny, Morphology and Toxigenicity of Ostreopsis cf. siamensis (Dinophyceae) from Temperate South-East Australia. Phycol. Res. 2016, 64, 146–159. [Google Scholar] [CrossRef]
- Pavaux, A.-S.; Berdalet, E.; Lemée, R. Chemical Ecology of the Benthic Dinoflagellate Genus Ostreopsis: Review of Progress and Future Directions. Front. Mar. Sci. 2020, 7, 498. [Google Scholar] [CrossRef]
- Chomérat, N.; iti Gatti, C.M.; Nézan, E.; Chinain, M. Studies on the Benthic Genus Sinophysis (Dinophysales, Dinophyceae) II. S. canaliculata from Rapa Island (French Polynesia). Phycologia 2017, 56, 193–203. [Google Scholar] [CrossRef]
- Lazure, P.; Dumas, F. An External–Internal Mode Coupling for a 3D Hydrodynamical Model for Applications at Regional Scale (MARS). Adv. Water Resour. 2008, 31, 233–250. [Google Scholar] [CrossRef]
- Guillard, R.R.L.; Hargraves, P.E. Stichochrysis immobilis is a Diatom, not a chrysophyte. Phycologia 1993, 32, 234–236. [Google Scholar] [CrossRef]
- Nunn, G.B.; Theisen, B.F.; Christensen, B.; Arctander, P. Simplicity-Correlated Size Growth of the Nuclear 28S Ribosomal RNA D3 Expansion Segment in the Crustacean Order Isopoda. J. Mol. Evol. 1996, 42, 211–223. [Google Scholar] [CrossRef]
- Nézan, E.; Siano, R.; Boulben, S.; Six, C.; Bilien, G.; Chèze, K.; Duval, A.; Le Panse, S.; Quéré, J.; Chomérat, N. Genetic Diversity of the Harmful Family Kareniaceae (Gymnodiniales, Dinophyceae) in France, with the Description of Karlodinium gentienii sp. Nov.: A New Potentially Toxic Dinoflagellate. Harmful Algae 2014, 40, 75–91. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT Online Service: Multiple Sequence Alignment, Interactive Sequence Choice and Visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian Phylogenetic Inference under Mixed Models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Uemura, D.; Hirata, Y. Studies on Palytoxins. Tetrahedron 1985, 41, 1007–1017. [Google Scholar] [CrossRef]
- Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; Grauso, L.; Tartaglione, L.; Florio, C.; Lorenzon, P.; De Bortoli, M.; et al. Stereostructure and Biological Activity of 42-Hydroxy-Palytoxin: A New Palytoxin Analogue from Hawaiian Palythoa Subspecies. Chem. Res. Toxicol. 2009, 22, 1851–1859. [Google Scholar] [CrossRef] [PubMed]
- Kerbrat, A.S.; Amzil, Z.; Pawlowiez, R.; Golubic, S.; Sibat, M.; Darius, H.T.; Chinain, M.; Laurent, D. First Evidence of Palytoxin and 42-Hydroxy-Palytoxin in the Marine Cyanobacterium Trichodesmium. Marine Drugs 2011, 9, 543–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenoir, S.; Ten-Hage, L.; Turquet, J.; Quod, J.-P.; Bernard, C.; Hennion, M.-C. First Evidence of Palytoxin Analogues from an Ostreopsis Mascarenensis (Dinophyceae) Benthic Bloom in Southwestern Indian Ocean. J. Phycol. 2004, 40, 1042–1051. [Google Scholar] [CrossRef]
- Rossi, R.; Castellano, V.; Scalco, E.; Serpe, L.; Zingone, A.; Soprano, V. New Palytoxin-like Molecules in Mediterranean Ostreopsis cf. ovata (Dinoflagellates) and in Palythoa Tuberculosa Detected by Liquid Chromatography-Electrospray Ionization Time-of-Flight Mass Spectrometry. Toxicon 2010, 56, 1381–1387. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Analyzed | Toxins | Cellular Content (fg eq. PLTX·Cell−1) |
|---|---|---|
| Bloom water sample (16 August 2021) | OVTX-a | 5.5 1 |
| OVTX-b | 2.3 1 |
| Strain | Species | Concentrations (pg eq. PLTX·Cell−1) | Cell Concentration (Cells·mL−1) | ||||
|---|---|---|---|---|---|---|---|
| IFR-OST-01E | Ostreopsis cf. ovata | OVTX-a | OVTX-b | OVTX-c | OVTX-d | OVTX-e | 80,700 |
| 4.3 | 1.6 | 0.2 | 0.3 | 0.3 | |||
| IFR-OST-02E | Ostreopsis sp. 9 (O. cf. siamensis) | No targeted toxins detected | 96,300 | ||||
| IFR-OST-03E | Ostreopsis sp. 9 (O. cf. siamensis) | No targeted toxins detected | 84,000 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chomérat, N.; Antajan, E.; Auby, I.; Bilien, G.; Carpentier, L.; Casamajor, M.-N.d.; Ganthy, F.; Hervé, F.; Labadie, M.; Méteigner, C.; et al. First Characterization of Ostreopsis cf. ovata (Dinophyceae) and Detection of Ovatoxins during a Multispecific and Toxic Ostreopsis Bloom on French Atlantic Coast. Mar. Drugs 2022, 20, 461. https://doi.org/10.3390/md20070461
Chomérat N, Antajan E, Auby I, Bilien G, Carpentier L, Casamajor M-Nd, Ganthy F, Hervé F, Labadie M, Méteigner C, et al. First Characterization of Ostreopsis cf. ovata (Dinophyceae) and Detection of Ovatoxins during a Multispecific and Toxic Ostreopsis Bloom on French Atlantic Coast. Marine Drugs. 2022; 20(7):461. https://doi.org/10.3390/md20070461
Chicago/Turabian StyleChomérat, Nicolas, Elvire Antajan, Isabelle Auby, Gwenael Bilien, Liliane Carpentier, Marie-Noëlle de Casamajor, Florian Ganthy, Fabienne Hervé, Magali Labadie, Claire Méteigner, and et al. 2022. "First Characterization of Ostreopsis cf. ovata (Dinophyceae) and Detection of Ovatoxins during a Multispecific and Toxic Ostreopsis Bloom on French Atlantic Coast" Marine Drugs 20, no. 7: 461. https://doi.org/10.3390/md20070461
APA StyleChomérat, N., Antajan, E., Auby, I., Bilien, G., Carpentier, L., Casamajor, M.-N. d., Ganthy, F., Hervé, F., Labadie, M., Méteigner, C., Paradis, C., Perrière-Rumèbe, M., Sanchez, F., Séchet, V., & Amzil, Z. (2022). First Characterization of Ostreopsis cf. ovata (Dinophyceae) and Detection of Ovatoxins during a Multispecific and Toxic Ostreopsis Bloom on French Atlantic Coast. Marine Drugs, 20(7), 461. https://doi.org/10.3390/md20070461