
Biochemical Insights into a Novel Family 2 Glycoside Hydrolase with Both β-1,3-Galactosidase and β-1,4-Galactosidase Activity from the Arctic

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
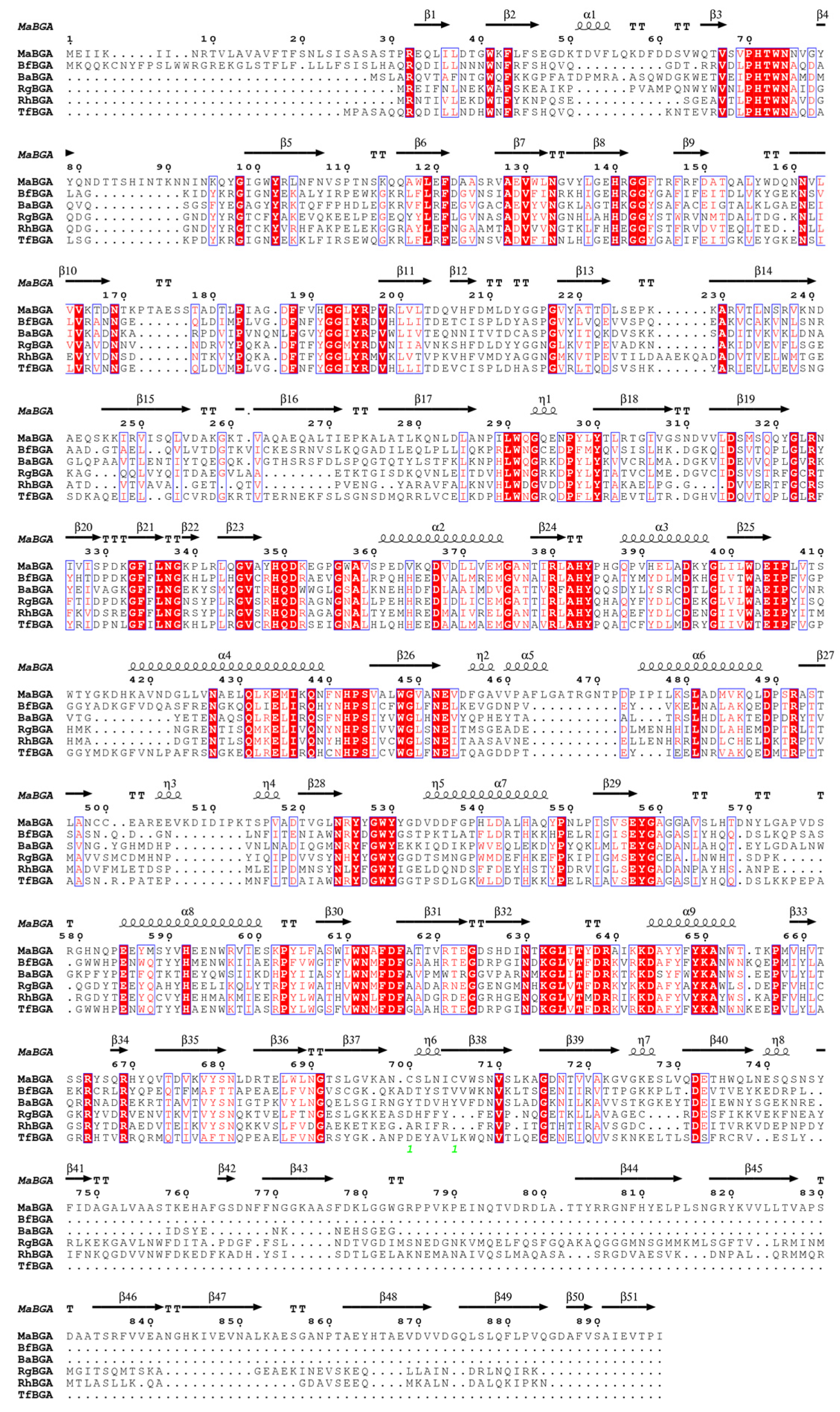
2.1. Sequence Analysis
2.2. Expression and Purification
2.3. Effects of Temperature on the Activity and Stability of the Enzyme
2.4. Effects of pH on the Activity and Stability of the Enzyme
2.5. Effects of NaCl on the Activity and Stability of the Enzyme
2.6. Effects of Metal Ions and Chemicals on the Activity of the Enzyme
2.7. Effects of Organic Solvents on the Stability of the Enzyme
2.8. Substrate Specificity of the Enzyme
2.9. Steady-State Kinetics
2.10. Thermal Denaturation Kinetics
3. Materials and Methods
3.1. Strains, Culture Conditions and Reagents
3.2. Gene Cloning and Sequence Analysis
3.3. Gene Expression and Protein Purification
3.4. Electrophoretic Analysis
3.5. β-Galactosidase Activity Assay
3.6. Effects of Temperature on the Activity and Stability of the Enzyme
3.7. Effects of pH on the Activity and Stability of the Enzyme
3.8. Effects of NaCl on the Activity and Stability of the Enzyme
3.9. Effects of Metal Ions and Chemicals on the Activity of the Enzyme
3.10. Effects of Organic Solvents on the Stability of the Enzyme
3.11. Substrate Specificity of the Enzyme
3.12. Steady-State Kinetics
3.13. Thermal Denaturation Kinetics
3.14. Analysis of Variance
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Saqib, S.; Akram, A.; Halim, S.A.; Tassaduq, R. Sources of beta-galactosidase and its applications in food industry. 3 Biotech 2017, 7, 79. [Google Scholar] [CrossRef]
- Guerrero, C.; Vera, C.; Conejeros, R.; Illanes, A. Transgalactosylation and hydrolytic activities of commercial preparations of beta-galactosidase for the synthesis of prebiotic carbohydrates. Enzyme Microb. Technol. 2015, 70, 9–17. [Google Scholar] [CrossRef]
- Movahedpour, A.; Ahmadi, N.; Ghalamfarsa, F.; Ghesmati, Z.; Khalifeh, M.; Maleksabet, A.; Shabaninejad, Z.; Taheri-Anganeh, M.; Savardashtaki, A. Beta-Galactosidase: From its source and applications to its recombinant form. Biotechnol. Appl. Biochem. 2022, 69, 612–628. [Google Scholar] [CrossRef]
- Vera, C.; Guerrero, C.; Aburto, C.; Cordova, A.; Illanes, A. Conventional and non-conventional applications of beta-galactosidases. Biochim. Biophys. Acta Proteins Proteom. 2020, 1868, 140271. [Google Scholar] [CrossRef]
- Li, D.; Li, S.; Wu, Y.; Jin, M.; Zhou, Y.; Wang, Y.; Chen, X.; Han, Y. Cloning and characterization of a new beta-galactosidase from Alteromonas sp. QD01 and its potential in synthesis of galacto-oligosaccharides. Mar. Drugs 2020, 18, 312. [Google Scholar] [CrossRef]
- Torres, D.P.M.; Goncalves, M.; Teixeira, J.A.; Rodrigues, L.R. Galacto-Oligosaccharides: Production, properties, applications, and significance as prebiotics. Compr. Rev. Food Sci. Food Saf. 2010, 9, 438–454. [Google Scholar] [CrossRef]
- Batsalova, K.; Kunchev, K.; Popova, Y.; Kozhukharova, A.; Kirova, N. Hydrolysis of lactose by β-galactosidase immobilized in polyvinylalcohol. Appl. Microbiol. Biot. 1987, 26, 227–230. [Google Scholar] [CrossRef]
- Zhou, Q.Z.K.; Chen, X.D. Effects of temperature and pH on the catalytic activity of the immobilized beta-galactosidase from Kluyveromyces lactis. Biochem. Eng. J. 2001, 9, 33–40. [Google Scholar] [CrossRef]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef]
- Yoshida, E.; Sakurama, H.; Kiyohara, M.; Nakajima, M.; Kitaoka, M.; Ashida, H.; Hirose, J.; Katayama, T.; Yamamoto, K.; Kumagai, H. Bifidobacterium longum subsp. infantis uses two different beta-galactosidases for selectively degrading type-1 and type-2 human milk oligosaccharides. Glycobiology 2012, 22, 361–368. [Google Scholar] [CrossRef]
- Kondo, T.; Nishimura, Y.; Matsuyama, K.; Ishimaru, M.; Nakazawa, M.; Ueda, M.; Sakamoto, T. Characterization of three GH35 beta-galactosidases, enzymes able to shave galactosyl residues linked to rhamnogalacturonan in pectin, from Penicillium chrysogenum 31B. Appl. Microbiol. Biot. 2020, 104, 1135–1148. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Zeng, Q.; Zhou, L.; Yu, Y.; Chen, B. Biochemical and structural insights into a novel thermostable beta-1,3-galactosidase from Marinomonas sp. BSi20414. Mar. Drugs 2017, 15, 2–15. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Liu, L.; Cui, L.; Zheng, T.; Ji, B.; Liu, K. Characterization of two beta-galactosidases LacZ and WspA1 from Nostoc flagelliforme with focus on the latter′s central active region. Sci. Rep. 2021, 11, 18448. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Guo, L.; Wang, K.; Liu, Y.; Xiao, M. Beta-Galactosidases: A great tool for synthesizing galactose-containing carbohydrates. Biotechnol. Adv. 2020, 39, 107465. [Google Scholar] [CrossRef]
- Guo, Q.; Liu, Y.; Cui, S.W. Structure, Classification and modification of polysaccharides. In Comprehensive Glycoscience; Elsevier: Amsterdam, The Netherlands, 2021; pp. 204–219. [Google Scholar]
- Kotake, T.; Dina, S.; Konishi, T.; Kaneko, S.; Igarashi, K.; Samejima, M.; Watanabe, Y.; Kimura, K.; Tsumuraya, Y. Molecular cloning of a β-galactosidase from radish that specifically hydrolyzes β-(1,3)- and β-(1,6)-galactosyl residues of arabinogalactan protein. Plant Physiol. 2005, 138, 1563–1576. [Google Scholar] [CrossRef]
- Fujimoto, H.; Miyasato, M.; Ito, Y.; Sasaki, T.; Ajisaka, K. Purification and properties of recombinant β-galactosidase from Bacillus circulans. Glycoconj. J. 1998, 15, 155–160. [Google Scholar] [CrossRef]
- Zhang, X.; Yu, G.; Leng, J.; Zhang, H.; Zhou, Y.; Yuan, Y.; Gao, J. Glycoside hydrolase family 2 exo-β-1,6-galactosidase LpGal2 from Lactobacillus plantarum: Cloning, expression, and enzymatic characterization. Process Biochem. 2021, 102, 269–274. [Google Scholar] [CrossRef]
- Viborg, A.H.; Fredslund, F.; Katayama, T.; Nielsen, S.K.; Svensson, B.; Kitaoka, M.; Lo Leggio, L.; Abou Hachem, M. A beta1-6/beta1-3 galactosidase from Bifidobacterium animalis subsp. lactis Bl-04 gives insight into sub-specificities of beta-galactoside catabolism within Bifidobacterium. Mol. Microbiol. 2014, 94, 1024–1040. [Google Scholar] [CrossRef]
- Viborg, A.H.; Katayama, T.; Abou Hachem, M.; Andersen, M.C.; Nishimoto, M.; Clausen, M.H.; Urashima, T.; Svensson, B.; Kitaoka, M. Distinct substrate specificities of three glycoside hydrolase family 42 beta-galactosidases from Bifidobacterium longum subsp. infantis ATCC 15697. Glycobiology 2014, 24, 208–216. [Google Scholar] [CrossRef]
- Zhou, A.; Yi, H.; Yan, X.; Mao, Z.; Deng, Y.; Lv, X.; Wang, D.; Zhang, X. Cloning, expression, purification and characterization of the beta-galactosidase PobetaGal35A from Penicillium oxalicum. Mol. Biotechnol. 2022, 65, 1140–1150. [Google Scholar] [CrossRef]
- Guo, B.S.; Zheng, F.; Crouch, L.; Cai, Z.P.; Wang, M.; Bolam, D.N.; Liu, L.; Voglmeir, J. Cloning, purification and biochemical characterisation of a GH35 beta-1,3/beta-1,6-galactosidase from the mucin-degrading gut bacterium Akkermansia muciniphila. Glycoconj. J. 2018, 35, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Sekimata, M.; Ogura, K.; Tsumuraya, Y.; Hashimoto, Y.; Yamamoto, S. A β-galactosidase from radish (Raphanus sativus L.) seeds. Plant Physiol. 1989, 90, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Hirano, Y.; Tsumuraya, Y.; Hashimoto, Y. Characterization of spinach leaf α-L-arabinofuranosidases and β-galactosidases and their synergistic action on an endogenous arabinogalactan-protein. Physiol. Plantarum. 1994, 92, 286–296. [Google Scholar] [CrossRef]
- Ishimaru, M.; Smith, D.L.; Mort, A.J.; Gross, K.C. Enzymatic activity and substrate specificity of recombinant tomato beta-galactosidases 4 and 5. Planta 2009, 229, 447–456. [Google Scholar] [CrossRef]
- Naundorf, A.M.C.; Ajisaka, K. Characterization of the immobilized β-galactosidase from Bacillus circulans and the production of β (1-3)-linked disaccharides. J. Agric. Chem. Soc. Jpn. 1998, 62, 1313–1317. [Google Scholar]
- Mangiagalli, M.; Lotti, M. Cold-Active beta-galactosidases: Insight into cold adaption mechanisms and biotechnological exploitation. Mar. Drugs 2021, 19, 43. [Google Scholar] [CrossRef]
- Feller, G.; Gerday, C. Psychrophilic enzymes: Hot topics in cold adaptation. Nat. Rev. Microbiol. 2003, 1, 200–208. [Google Scholar] [CrossRef]
- Li, S.; Zhu, X.; Xing, M. A new beta-galactosidase from the Antarctic bacterium Alteromonas sp. ANT48 and its potential in formation of prebiotic galacto-oligosaccharides. Mar. Drugs 2019, 17, 599. [Google Scholar] [CrossRef]
- Mangiagalli, M.; Lapi, M.; Maione, S.; Orlando, M.; Brocca, S.; Pesce, A.; Barbiroli, A.; Camilloni, C.; Pucciarelli, S.; Lotti, M.; et al. The co-existence of cold activity and thermal stability in an Antarctic GH42 beta-galactosidase relies on its hexameric quaternary arrangement. FEBS J. 2021, 288, 546–565. [Google Scholar] [CrossRef]
- Talens-Perales, D.; Gorska, A.; Huson, D.H.; Polaina, J.; Marin-Navarro, J. Analysis of domain architecture and phylogenetics of family 2 Glycoside Hydrolases (GH2). PLoS ONE 2016, 11, e0168035. [Google Scholar] [CrossRef]
- Madeira, F.; Pearce, M.; Tivey, A.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and sequence analysis tools services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. [Google Scholar] [CrossRef]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Burrin, J.M.; Price, C.P. Measurement of blood glucose. Ann. Clin. Biochem. 1985, 22, 327–342. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Wang, L.; Jiang, Y.L.; Bai, X.H.; Chu, J.; Li, Q.; Yu, G.; Liang, Q.L.; Zhou, C.Z.; Chen, Y. Structural insights into the substrate specificity of Streptococcus pneumoniae beta (1,3)-galactosidase BgaC. J. Biol. Chem. 2012, 287, 22910–22918. [Google Scholar] [CrossRef]
- Wang, E.Z. Measurement Method and Diagnosis/Measurement Kit of Galactose; Suzhou Aijie Biotechnology Co., Ltd.: Suzhou, China, 2011. [Google Scholar]
- Bhatti, H.N.; Madeeha, M.; Asgher, M.; Batool, N. Purification and thermodynamic characterization of glucose oxidase from a newly isolated strain of Aspergillus niger. Can. J. Microbiol. 2006, 52, 519–524. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metal Ions | Relative Activity (%) | Chemicals | Relative Activity (%) |
|---|---|---|---|
| Control | 100 b | Glycerol | 134.846 ± 12.909 a |
| Zn2+ | 135.857 ± 8.286 a | L-glutathione (oxidized) | 140.193 ± 3.047 a |
| Mg2+ | 142.263 ± 0.423 a | EDTA·2Na | 40.141 ± 0.488 e |
| Ni2+ | 121.218 ± 6.632 b | SDS | 45.161 ± 6.920 e |
| K+ | 86.545 ± 3.926 cd | ||
| Na+ | 85.855 ± 2.771 cd | ||
| Fe2+ | 92.927 ± 4.478 bc | ||
| Fe3+ | 78.092 ± 1.936 d | ||
| Ca2+ | 98.620 ± 4.952 b |
| Natural Substrate | Substrate Activity (%) |
|---|---|
| Lactose | 100 |
| Trehalose | 5.618 |
| Maltose | 1.053 |
| Cellobiose | <1 |
| Arabinogalactan | 0 |
| Pectic galactan | 0 |
| Substrates | Km (mM) | kcat (s−1) | kcat/Km (s−1 mM−1) |
|---|---|---|---|
| ONPG | 0.294 ± 0.044 | 13.160 ± 0.459 | 46.028 ± 8.450 |
| β-lactose | 5.265 ± 0.520 | 7.768 ± 0.212 | 1.494 ± 0.188 |
| Galβ-(1,3)-GlcNAc | 2.107 ± 0.484 | 0.025 ± 0.001 | 0.013 ± 0.004 |
| Galβ-(1,4)-GlcNAc | 7.023 ± 1.181 | 3.388 ± 0.156 | 0.500 ± 0.106 |
| Temperate (°C) | kd (h−1) | t1/2 (h) | ΔH* (kJmol−1) | ΔG* (kJmol−1) | ΔS* (Jmol−1K−1) |
|---|---|---|---|---|---|
| 20 | 0.00153 | 452.94 | 118.19 | 87.55 | 104.52 |
| 30 | 0.0035 | 196.32 | 115.67 | 85.62 | 99.14 |
| 40 | 0.0116 | 59.94 | 113.07 | 82.77 | 96.73 |
| 50 | 0.189 | 3.67 | 110.38 | 81.24 | 90.18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Wang, Z.; Yu, Y.; Li, H.; Luo, W.; Chen, B.; Niu, G.; Ding, H. Biochemical Insights into a Novel Family 2 Glycoside Hydrolase with Both β-1,3-Galactosidase and β-1,4-Galactosidase Activity from the Arctic. Mar. Drugs 2023, 21, 521. https://doi.org/10.3390/md21100521
Li D, Wang Z, Yu Y, Li H, Luo W, Chen B, Niu G, Ding H. Biochemical Insights into a Novel Family 2 Glycoside Hydrolase with Both β-1,3-Galactosidase and β-1,4-Galactosidase Activity from the Arctic. Marine Drugs. 2023; 21(10):521. https://doi.org/10.3390/md21100521
Chicago/Turabian StyleLi, Dianyi, Zheng Wang, Yong Yu, Huirong Li, Wei Luo, Bo Chen, Guoqing Niu, and Haitao Ding. 2023. "Biochemical Insights into a Novel Family 2 Glycoside Hydrolase with Both β-1,3-Galactosidase and β-1,4-Galactosidase Activity from the Arctic" Marine Drugs 21, no. 10: 521. https://doi.org/10.3390/md21100521
APA StyleLi, D., Wang, Z., Yu, Y., Li, H., Luo, W., Chen, B., Niu, G., & Ding, H. (2023). Biochemical Insights into a Novel Family 2 Glycoside Hydrolase with Both β-1,3-Galactosidase and β-1,4-Galactosidase Activity from the Arctic. Marine Drugs, 21(10), 521. https://doi.org/10.3390/md21100521