
Species-Specific N-Glycomes and Methylation Patterns of Oysters Crassostrea gigas and Ostrea edulis and Their Possible Consequences for the Norovirus–HBGA Interaction

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Release and Analysis of N-Glycans
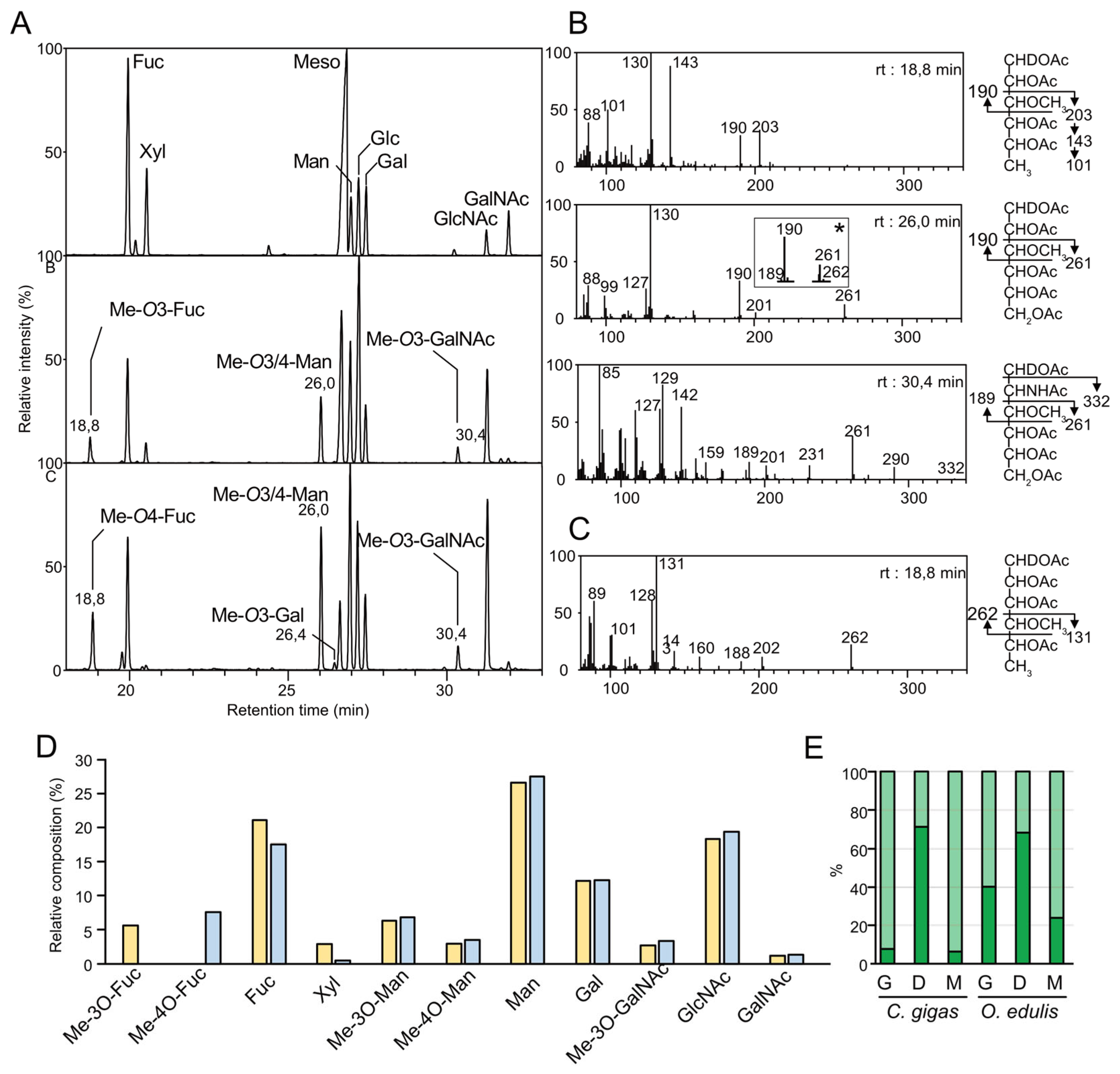
2.2. Monosaccharide Composition of N-Glycans
2.3. N-Glycome Analysis
2.3.1. N-Glycome of C. gigas
2.3.2. N-Glycome of O. edulis
2.4. Methyl N-Glycome Analysis
2.4.1. Localization of Methyl Groups on HBGAs
2.4.2. Relative Quantification of Methylated Antigens
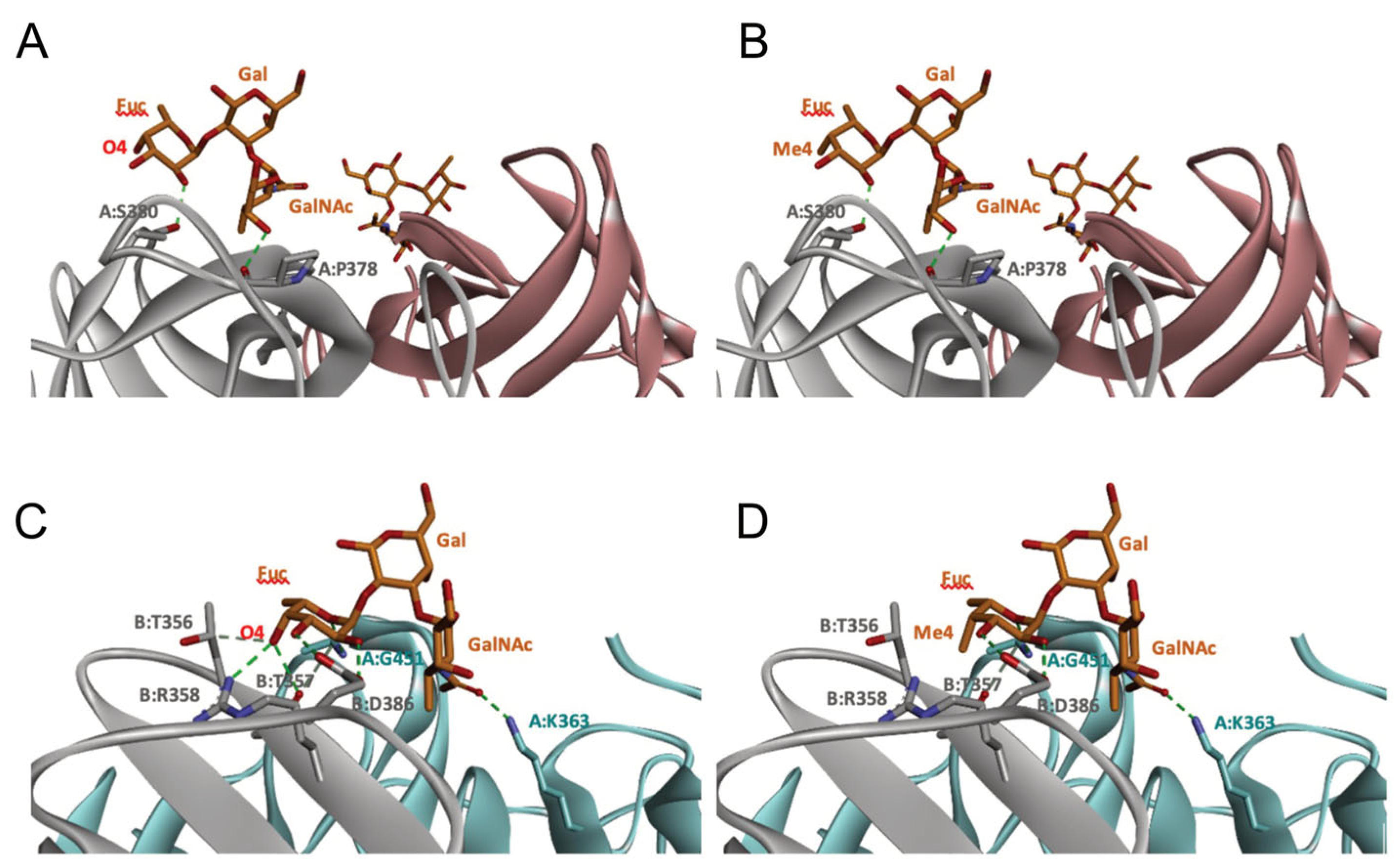
2.5. Molecular Modeling of Norovirus P Domain Interaction with Methylated A Antigens
3. Discussion
4. Materials and Methods
4.1. Preparation of Anatomic Pieces
4.2. Preparation of N-Glycans
4.3. Protein Assay
4.4. Permethylation of N-Glycans
4.5. MALDI-MS Analysis
4.6. Quantification of A and ALeb Epitopes Using NanoLC-MS/MS
4.7. Preparation of Itol-Acetate Derivatives
4.8. GC-MS Analysis Itol-Acetates and Permethylated Derivatives
4.9. Molecular Modeling
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Farahmand, M.; Moghoofei, M.; Dorost, A.; Shoja, Z.; Ghorbani, S.; Kiani, S.J.; Khales, P.; Esteghamati, A.; Sayyahfar, S.; Jafarzadeh, M.; et al. Global Prevalence and Genotype Distribution of Norovirus Infection in Children with Gastroenteritis: A Meta-analysis on 6 Years of Research from 2015 to 2020. Rev. Med. Virol. 2022, 32, e2237. [Google Scholar] [CrossRef]
- Glass, R.I.; Parashar, U.D.; Estes, M.K. Norovirus Gastroenteritis. N. Engl. J. Med. 2009, 361, 1776–1785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bányai, K.; Estes, M.K.; Martella, V.; Parashar, U.D. Viral Gastroenteritis. Lancet 2018, 392, 175–186. [Google Scholar] [CrossRef]
- Caddy, S.; Papa, G.; Borodavka, A.; Desselberger, U. Rotavirus Research: 2014–2020. Virus Res. 2021, 304, 198499. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, P.; de Graaf, M.; Parra, G.I.; Chan, M.C.-W.; Green, K.; Martella, V.; Wang, Q.; White, P.A.; Katayama, K.; Vennema, H.; et al. Updated Classification of Norovirus Genogroups and Genotypes. J. Gen. Virol. 2019, 100, 1393–1406. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xue, L.; Gao, J.; Cai, W.; Zhang, Z.; Meng, L.; Miao, S.; Hong, X.; Xu, M.; Wu, Q.; et al. A Systematic Review and Meta-Analysis Indicates a Substantial Burden of Human Noroviruses in Shellfish Worldwide, with GII.4 and GII.2 Being the Predominant Genotypes. Food Microbiol. 2023, 109, 104140. [Google Scholar] [CrossRef] [PubMed]
- King, M.-J. Blood Group Antigens on Human Erythrocytes-Distribution, Structure and Possible Functions. Biochim. Biophys. Acta (BBA)-Rev. Biomembr. 1994, 1197, 15–44. [Google Scholar] [CrossRef]
- Ewald, D.R.; Sumner, S.C.J. Blood Type Biochemistry and Human Disease. WIREs Mech. Dis. 2016, 8, 517–535. [Google Scholar] [CrossRef] [Green Version]
- Ravn, V.; Dabelsteen, E. Tissue Distribution of Histo-Blood Group Antigens. APMIS 2000, 108, 1–28. [Google Scholar] [CrossRef]
- Marionneau, S.; Cailleau-Thomas, A.; Rocher, J.; Le Moullac-Vaidye, B.; Ruvoën, N.; Clément, M.; Le Pendu, J. ABH and Lewis Histo-Blood Group Antigens, a Model for the Meaning of Oligosaccharide Diversity in the Face of a Changing World. Biochimie 2001, 83, 565–573. [Google Scholar] [CrossRef]
- Ruvoën-Clouet, N.; Belliot, G.; Le Pendu, J. Noroviruses and Histo-Blood Groups: The Impact of Common Host Genetic Polymorphisms on Virus Transmission and Evolution: Noroviruses and Herd Innate Protection. Rev. Med. Virol. 2013, 23, 355–366. [Google Scholar] [CrossRef]
- Caddy, S.; Breiman, A.; le Pendu, J.; Goodfellow, I. Genogroup IV and VI Canine Noroviruses Interact with Histo-Blood Group Antigens. J. Virol. 2014, 88, 10377–10391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.; Liu, Y.; Tan, M. Histo-Blood Group Antigens as Receptors for Rotavirus, New Understanding on Rotavirus Epidemiology and Vaccine Strategy: Rotavirus Host Receptor and Vaccine Strategy. Emerg. Microbes Infect. 2017, 6, e22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Pendu, J.; Ruvoën-Clouet, N. Fondness for Sugars of Enteric Viruses Confronts Them with Human Glycans Genetic Diversity. Hum. Genet. 2020, 139, 903–910. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Cai, H.; Hu, L.; Lei, R.; Pan, Y.; Yan, S.; Wang, Y. Molecular Epidemiology of Oyster-Related Human Noroviruses and Their Global Genetic Diversity and Temporal-Geographical Distribution from 1983 to 2014. Appl. Environ. Microbiol. 2015, 81, 7615–7624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metcalf, T.G.; Melnick, J.L.; Estes, M.K. Environmental Virology: From Detection of Virus in Sewage and Water by Isolation to Identification by Molecular Biology—A Trip of over 50 Years. Annu. Rev. Microbiol. 1995, 49, 461–487. [Google Scholar] [CrossRef]
- Lowther, J.A.; Gustar, N.E.; Powell, A.L.; O’Brien, S.; Lees, D.N. A One-Year Survey of Norovirus in UK Oysters Collected at the Point of Sale. Food Environ. Virol. 2018, 10, 278–287. [Google Scholar] [CrossRef] [Green Version]
- Jørgensen, C. Bivalve Filter Feeding Revisited. Mar. Ecol. Prog. Ser. 1996, 142, 287–302. [Google Scholar] [CrossRef]
- Maalouf, H.; Schaeffer, J.; Parnaudeau, S.; Le Pendu, J.; Atmar, R.L.; Crawford, S.E.; Le Guyader, F.S. Strain-Dependent Norovirus Bioaccumulation in Oysters. Appl. Environ. Microbiol. 2011, 77, 3189–3196. [Google Scholar] [CrossRef] [Green Version]
- Loisy, F.; Atmar, R.L.; Le Saux, J.-C.; Cohen, J.; Caprais, M.-P.; Pommepuy, M.; Le Guyader, F.S. Use of Rotavirus Virus-Like Particles as Surrogates To Evaluate Virus Persistence in Shellfish. Appl. Environ. Microbiol. 2005, 71, 6049–6053. [Google Scholar] [CrossRef] [Green Version]
- Polo, D.; Schaeffer, J.; Fournet, N.; Le Saux, J.-C.; Parnaudeau, S.; McLeod, C.; Le Guyader, F.S. Digital PCR for Quantifying Norovirus in Oysters Implicated in Outbreaks, France. Emerg. Infect. Dis. 2016, 22, 2189–2191. [Google Scholar] [CrossRef] [PubMed]
- Rupnik, A.; Doré, W.; Devilly, L.; Fahy, J.; Fitzpatrick, A.; Schmidt, W.; Hunt, K.; Butler, F.; Keaveney, S. Evaluation of Norovirus Reduction in Environmentally Contaminated Pacific Oysters during Laboratory Controlled and Commercial Depuration. Food Environ. Virol. 2021, 13, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Battistini, R.; Masotti, C.; Maurella, C.; Costa, E.; Orlandi, M.; Feletti, M.; Ercolini, C.; Serracca, L. Study on the Effect of Relaying on Norovirus Reduction from Crassostrea gigas Oysters. Microorganisms 2022, 10, 2389. [Google Scholar] [CrossRef]
- Le Guyader, F.S.; Loisy, F.; Atmar, R.L.; Hutson, A.M.; Estes, M.K.; Ruvoën-Clouet, N.; Pommepuy, M.; Le Pendu, J. Norwalk Virus–Specific Binding to Oyster Digestive Tissues. Emerg. Infect. Dis. 2006, 12, 931–936. [Google Scholar] [CrossRef] [PubMed]
- Tian, P.; Engelbrektson, A.L.; Jiang, X.; Zhong, W.; Mandrell, R.E. Norovirus Recognizes Histo-Blood Group Antigens on Gastrointestinal Cells of Clams, Mussels, and Oysters: A Possible Mechanism of Bioaccumulation. J. Food Prot. 2007, 70, 2140–2147. [Google Scholar] [CrossRef] [Green Version]
- Maalouf, H.; Zakhour, M.; Le Pendu, J.; Le Saux, J.-C.; Atmar, R.L.; Le Guyader, F.S. Distribution in Tissue and Seasonal Variation of Norovirus Genogroup I and II Ligands in Oysters. Appl. Environ. Microbiol. 2010, 76, 5621–5630. [Google Scholar] [CrossRef] [Green Version]
- Lowmoung, T.; Pombubpa, K.; Duangdee, T.; Tipayamongkholgul, M.; Kittigul, L. Distribution of Naturally Occurring Norovirus Genogroups I, II, and IV in Oyster Tissues. Food Environ. Virol. 2017, 9, 415–422. [Google Scholar] [CrossRef]
- Su, L.; Ma, L.; Liu, H.; Zhao, F.; Su, Z.; Zhou, D. Presence and Distribution of Histo-Blood Group Antigens in Pacific Oysters and the Effects of Exposure to Noroviruses GI.3 and GII.4 on Their Expression. J. Food Prot. 2018, 81, 1783–1790. [Google Scholar] [CrossRef]
- Paschinger, K.; Wilson, I.B.H. Comparisons of N-Glycans across Invertebrate Phyla. Parasitology 2019, 146, 1733–1742. [Google Scholar] [CrossRef] [Green Version]
- Kurz, S.; Jin, C.; Hykollari, A.; Gregorich, D.; Giomarelli, B.; Vasta, G.R.; Wilson, I.B.H.; Paschinger, K. Hemocytes and Plasma of the Eastern Oyster (Crassostrea Virginica) Display a Diverse Repertoire of Sulfated and Blood Group A-Modified N-Glycans*. J. Biol. Chem. 2013, 288, 24410–24428. [Google Scholar] [CrossRef] [Green Version]
- Feng, C.; Ghosh, A.; Amin, M.N.; Giomarelli, B.; Shridhar, S.; Banerjee, A.; Fernández-Robledo, J.A.; Bianchet, M.A.; Wang, L.-X.; Wilson, I.B.H.; et al. The Galectin CvGal1 from the Eastern Oyster (Crassostrea Virginica) Binds to Blood Group A Oligosaccharides on the Hemocyte Surface. J. Biol. Chem. 2013, 288, 24394–24409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varki, A. Nothing in Glycobiology Makes Sense, except in the Light of Evolution. Cell 2006, 126, 841–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delannoy, C.; Huang, C.; Coddeville, B.; Chen, J.-Y.; Mouajjah, D.; Groux-Degroote, S.; Harduin-Lepers, A.; Khoo, K.-H.; Guerardel, Y.; Elass-Rochard, E. Mycobacterium Bovis BCG Infection Alters the Macrophage N -Glycome. Mol. Omics 2020, 16, 345–354. [Google Scholar] [CrossRef]
- Lemp, D.; Haselbeck, A.; Klebl, F. Molecular Cloning and Heterologous Expression of N-Glycosidase F from Flavobacterium Meningosepticum. J. Biol. Chem. 1990, 265, 15606–15610. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N. Demonstration of a New Amidase Acting on Glycopeptides. Biochem. Biophys. Res. Commun. 1977, 76, 1194–1201. [Google Scholar] [CrossRef]
- Qin, Y.; Li, R.; Liao, Q.; Shi, G.; Zhou, Y.; Wan, W.; Li, J.; Ma, H.; Zhang, Y.; Yu, Z. Comparison of Biochemical Composition, Nutritional Quality, and Metals Concentrations between Males and Females of Three Different Crassostrea sp. Food Chem. 2023, 398, 133868. [Google Scholar] [CrossRef]
- Yu, S.-Y.; Wu, S.-W.; Khoo, K.-H. Distinctive Characteristics of MALDI-Q/TOF and TOF/TOF Tandem Mass Spectrometry for Sequencing of Permethylated Complex Type N-Glycans. Glycoconj. J. 2006, 23, 355–369. [Google Scholar] [CrossRef]
- Domon, B.; Costello, C.E. A Systematic Nomenclature for Carbohydrate Fragmentations in FAB-MS/MS Spectra of Glycoconjugates. Glycoconj. J. 1988, 5, 397–409. [Google Scholar] [CrossRef]
- Varki, A.; Cummings, R.D.; Aebi, M.; Packer, N.H.; Seeberger, P.H.; Esko, J.D.; Stanley, P.; Hart, G.; Darvill, A.; Kinoshita, T.; et al. Symbol Nomenclature for Graphical Representations of Glycans. Glycobiology 2015, 25, 1323–1324. [Google Scholar] [CrossRef] [Green Version]
- Schachter, H. The ‘Yellow Brick Road’ to Branched Complex N-Glycans. Glycobiology 1991, 1, 453–461. [Google Scholar] [CrossRef]
- Hulsen, T.; de Vlieg, J.; Alkema, W. BioVenn—A Web Application for the Comparison and Visualization of Biological Lists Using Area-Proportional Venn Diagrams. BMC Genom. 2008, 9, 488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staudacher, E. Methylation—An Uncommon Modification of Glycans. Biol. Chem. 2012, 393, 675–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsiao, C.-T.; Wang, P.-W.; Chang, H.-C.; Chen, Y.-Y.; Wang, S.-H.; Chern, Y.; Khoo, K.-H. Advancing a High Throughput Glycotope-Centric Glycomics Workflow Based on NAnoLC-MS2-Product Dependent-MS3 ANAlysis of Permethylated Glycans*. Mol. Cell. Proteom. 2017, 16, 2268–2280. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, N.; Masuda, K.; Hiraki, K.; Yoshihara, K.; Huang, H.-H.; Khoo, K.-H.; Kato, K. N-Glycan Structures of Squid Rhodopsin. Existence of the Alpha1-3 and Alpha1-6 Difucosylated Innermost GlcNAc Residue in a Molluscan Glycoprotein. Eur. J. Biochem. 2003, 270, 2627–2632. [Google Scholar] [CrossRef] [PubMed]
- Oltmanns, A.; Hoepfner, L.; Scholz, M.; Zinzius, K.; Schulze, S.; Hippler, M. Novel Insights Into N-Glycan Fucosylation and Core Xylosylation in C. Reinhardtii. Front. Plant Sci. 2020, 10, 1686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minagawa, S.; Sekiguchi, S.; Nakaso, Y.; Tomita, M.; Takahisa, M.; Yasuda, H. Identification of Core Alpha 1,3-Fucosyltransferase Gene from Silkworm: An Insect Popularly Used to Express Mammalian Proteins. J. Insect. Sci. 2015, 15, 110. [Google Scholar] [CrossRef] [Green Version]
- Yan, S.; Vanbeselaere, J.; Jin, C.; Blaukopf, M.; Wöls, F.; Wilson, I.B.H.; Paschinger, K. Core Richness of N-Glycans of Caenorhabditis Elegans: A Case Study on Chemical and Enzymatic Release. Anal. Chem. 2018, 90, 928–935. [Google Scholar] [CrossRef] [Green Version]
- Fabini, G.; Freilinger, A.; Altmann, F.; Wilson, I.B.H. Identification of Core A1,3-Fucosylated Glycans and Cloning of the Requisite Fucosyltransferase CDNA from Drosophila Melanogaster: Potential Basis of the Neural Anti-Horseradish Peroxidase Epitope *. J. Biol. Chem. 2001, 276, 28058–28067. [Google Scholar] [CrossRef] [Green Version]
- Paschinger, K.; Staudacher, E.; Stemmer, U.; Fabini, G.; Wilson, I.B.H. Fucosyltransferase Substrate Specificity and the Order of Fucosylation in Invertebrates. Glycobiology 2005, 15, 463–474. [Google Scholar] [CrossRef] [Green Version]
- Titz, A.; Butschi, A.; Henrissat, B.; Fan, Y.-Y.; Hennet, T.; Razzazi-Fazeli, E.; Hengartner, M.O.; Wilson, I.B.H.; Künzler, M.; Aebi, M. Molecular Basis for Galactosylation of Core Fucose Residues in Invertebrates. J. Biol. Chem. 2009, 284, 36223–36233. [Google Scholar] [CrossRef] [Green Version]
- Eckmair, B.; Jin, C.; Abed-Navandi, D.; Paschinger, K. Multistep Fractionation and Mass Spectrometry Reveal Zwitterionic and Anionic Modifications of the N- and O-Glycans of a Marine Snail. Mol. Cell. Proteom. 2016, 15, 573–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoeva, S.; Rachev, R.; Severov, S.; Voelter, W.; Genov, N. Carbohydrate Content and Monosaccharide Composition of Rapana Thomasiana Grosse (Gastropoda) Hemocyanin and Its Structural Subunits. Comparison with Gastropodan Hemocyanins. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1995, 110, 761–765. [Google Scholar] [CrossRef] [PubMed]
- Khoo, K.-H. Structural Variations in Schistosomal Glycans. Trends Glycosci. Glycotechnol. 2001, 13, 493–506. [Google Scholar] [CrossRef]
- Smit, C.H.; van Diepen, A.; Nguyen, D.L.; Wuhrer, M.; Hoffmann, K.F.; Deelder, A.M.; Hokke, C.H. Glycomic Analysis of Life Stages of the Human Parasite Schistosoma Mansoni Reveals Developmental Expression Profiles of Functional and Antigenic Glycan Motifs *[S]. Mol. Cell. Proteom. 2015, 14, 1750–1769. [Google Scholar] [CrossRef] [Green Version]
- Hori, T.; Sugita, M.; Ando, S.; Kuwahara, M.; Kumauchi, K.; Sugie, E.; Itasaka, O. Characterization of a Novel Glycosphingolipid, Ceramide Nonasaccharide, Isolated from Spermatozoa of the Fresh Water Bivalve, Hyriopsis Schlegelii. J. Biol. Chem. 1981, 256, 10979–10985. [Google Scholar] [CrossRef]
- Spiro, R.G.; Bhoyroo, V.D. Occurrence of Sulfate in the Asparagine-Linked Complex Carbohydrate Units of Thyroglobulin. Identification and Localization of Galactose 3-Sulfate and N-Acetylglucosamine 6-Sulfate Residues in the Human and Calf Proteins. J. Biol. Chem. 1988, 263, 14351–14358. [Google Scholar] [CrossRef]
- Miyata, S.; Sato, C.; Kumita, H.; Toriyama, M.; Vacquier, V.D.; Kitajima, K. Flagellasialin: A Novel Sulfated 2,9-Linked Polysialic Acid Glycoprotein of Sea Urchin Sperm Flagella. Glycobiology 2006, 16, 1229–1241. [Google Scholar] [CrossRef]
- Yu, S.-Y.; Wu, S.-W.; Hsiao, H.-H.; Khoo, K.-H. Enabling Techniques and Strategic Workflow for Sulfoglycomics Based on Mass Spectrometry Mapping and Sequencing of Permethylated Sulfated Glycans. Glycobiology 2009, 19, 1136–1149. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.-Y.; Snovida, S.; Khoo, K.-H. Permethylation and Microfractionation of Sulfated Glycans for MS Analysis. Bio-Protocol 2020, 10, e3617. [Google Scholar] [CrossRef]
- Tan, M.; Jiang, X. Histo-Blood Group Antigens: A Common Niche for Norovirus and Rotavirus. Expert Rev. Mol. Med. 2014, 16, e5. [Google Scholar] [CrossRef]
- Taube, S.; Mallagaray, A.; Peters, T. Norovirus, Glycans and Attachment. Curr. Opin. Virol. 2018, 31, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Tenge, V.R.; Hu, L.; Prasad, B.V.V.; Larson, G.; Atmar, R.L.; Estes, M.K.; Ramani, S. Glycan Recognition in Human Norovirus Infections. Viruses 2021, 13, 2066. [Google Scholar] [CrossRef] [PubMed]
- Le Guyader, F.S.; Atmar, R.L.; Le Pendu, J. Transmission of Viruses through Shellfish: When Specific Ligands Come into Play. Curr. Opin. Virol. 2012, 2, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Fiege, B.; Rademacher, C.; Cartmell, J.; Kitov, P.I.; Parra, F.; Peters, T. Molecular Details of the Recognition of Blood Group Antigens by a Human Norovirus as Determined by STD NMR Spectroscopy. Angew. Chem. Int. Ed. Engl. 2012, 51, 928–932. [Google Scholar] [CrossRef]
- Fiege, B.; Leuthold, M.; Parra, F.; Dalton, K.P.; Meloncelli, P.J.; Lowary, T.L.; Peters, T. Epitope Mapping of Histo Blood Group Antigens Bound to Norovirus VLPs Using STD NMR Experiments Reveals Fine Details of Molecular Recognition. Glycoconj. J. 2017, 34, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Tseng, H.C.; Hsiao, C.T.; Yamakawa, N.; Guérardel, Y.; Khoo, K.H. Discovery Sulfoglycomics and Identification of the Characteristic Fragment Ions for High-Sensitivity Precise Mapping of Adult Zebrafish Brain-Specific Glycotopes. Front Mol Biosci. 2021, 8. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Trisaccharides | ||
|---|---|---|---|
| α-l-Fuc-(1→2)-[α-d-GalNAc-(1→3)]-β-D-Gal | α-l-Fuc-(1→2)-[α-d-Me3GalNAc-(1→3)]-β-D-Gal | α-l-Me4-Fuc-(1→2)-[α-d-Me3GalNAc-(1→3)]-β-D-Gal | |
| Norwalk |  |  |  |
| TV24 |  |  |  |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Auger, A.; Yu, S.-Y.; Guu, S.-Y.; Quéméner, A.; Euller-Nicolas, G.; Ando, H.; Desdouits, M.; Le Guyader, F.S.; Khoo, K.-H.; Le Pendu, J.; et al. Species-Specific N-Glycomes and Methylation Patterns of Oysters Crassostrea gigas and Ostrea edulis and Their Possible Consequences for the Norovirus–HBGA Interaction. Mar. Drugs 2023, 21, 342. https://doi.org/10.3390/md21060342
Auger A, Yu S-Y, Guu S-Y, Quéméner A, Euller-Nicolas G, Ando H, Desdouits M, Le Guyader FS, Khoo K-H, Le Pendu J, et al. Species-Specific N-Glycomes and Methylation Patterns of Oysters Crassostrea gigas and Ostrea edulis and Their Possible Consequences for the Norovirus–HBGA Interaction. Marine Drugs. 2023; 21(6):342. https://doi.org/10.3390/md21060342
Chicago/Turabian StyleAuger, Audrey, Shin-Yi Yu, Shih-Yun Guu, Agnès Quéméner, Gabriel Euller-Nicolas, Hiromune Ando, Marion Desdouits, Françoise S. Le Guyader, Kay-Hooi Khoo, Jacques Le Pendu, and et al. 2023. "Species-Specific N-Glycomes and Methylation Patterns of Oysters Crassostrea gigas and Ostrea edulis and Their Possible Consequences for the Norovirus–HBGA Interaction" Marine Drugs 21, no. 6: 342. https://doi.org/10.3390/md21060342
APA StyleAuger, A., Yu, S.-Y., Guu, S.-Y., Quéméner, A., Euller-Nicolas, G., Ando, H., Desdouits, M., Le Guyader, F. S., Khoo, K.-H., Le Pendu, J., Chirat, F., & Guerardel, Y. (2023). Species-Specific N-Glycomes and Methylation Patterns of Oysters Crassostrea gigas and Ostrea edulis and Their Possible Consequences for the Norovirus–HBGA Interaction. Marine Drugs, 21(6), 342. https://doi.org/10.3390/md21060342