
Molecular Networking Revealed Unique UV-Absorbing Phospholipids: Favilipids from the Marine Sponge Clathria faviformis

 ,
,  , ,
, ,  and
and
Abstract
:
1. Introduction
2. Results
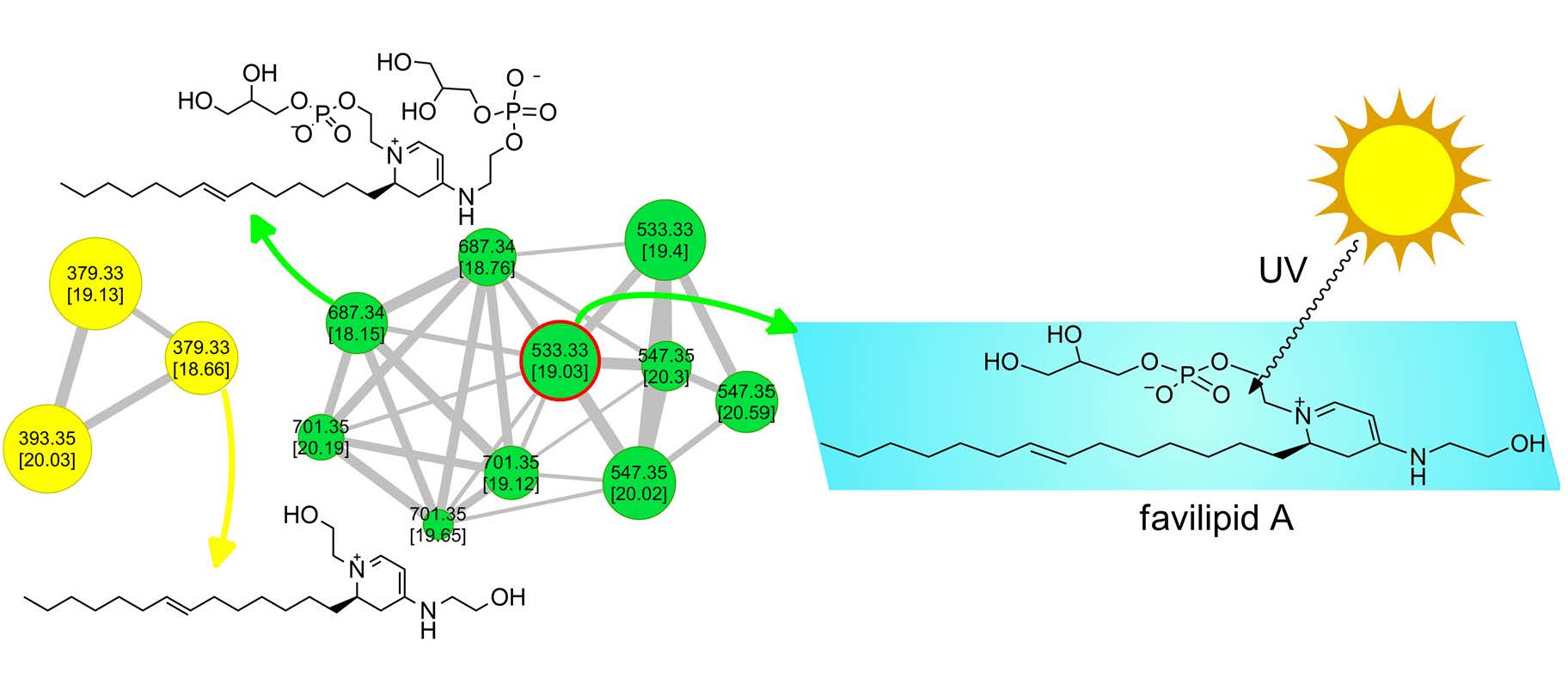
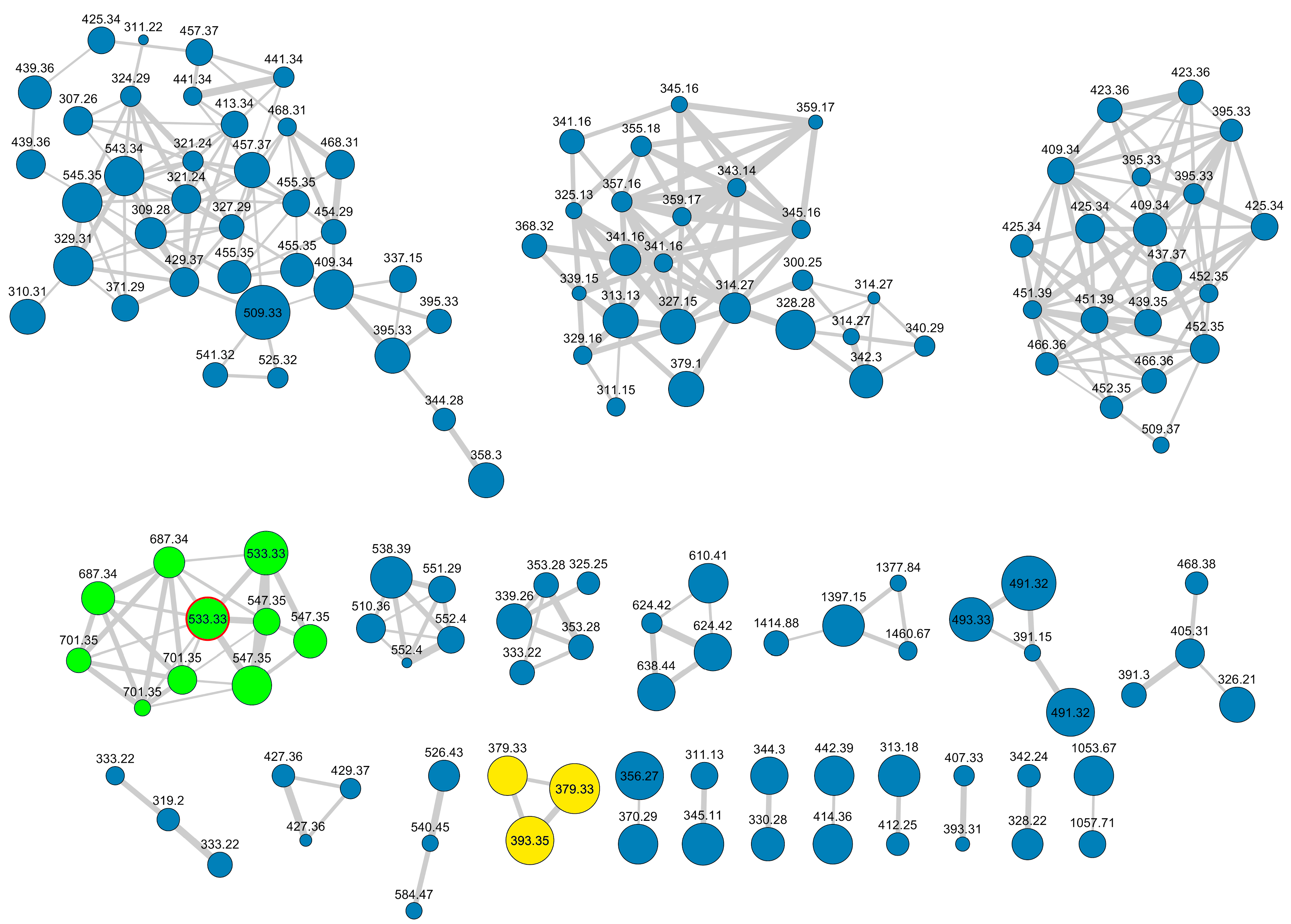
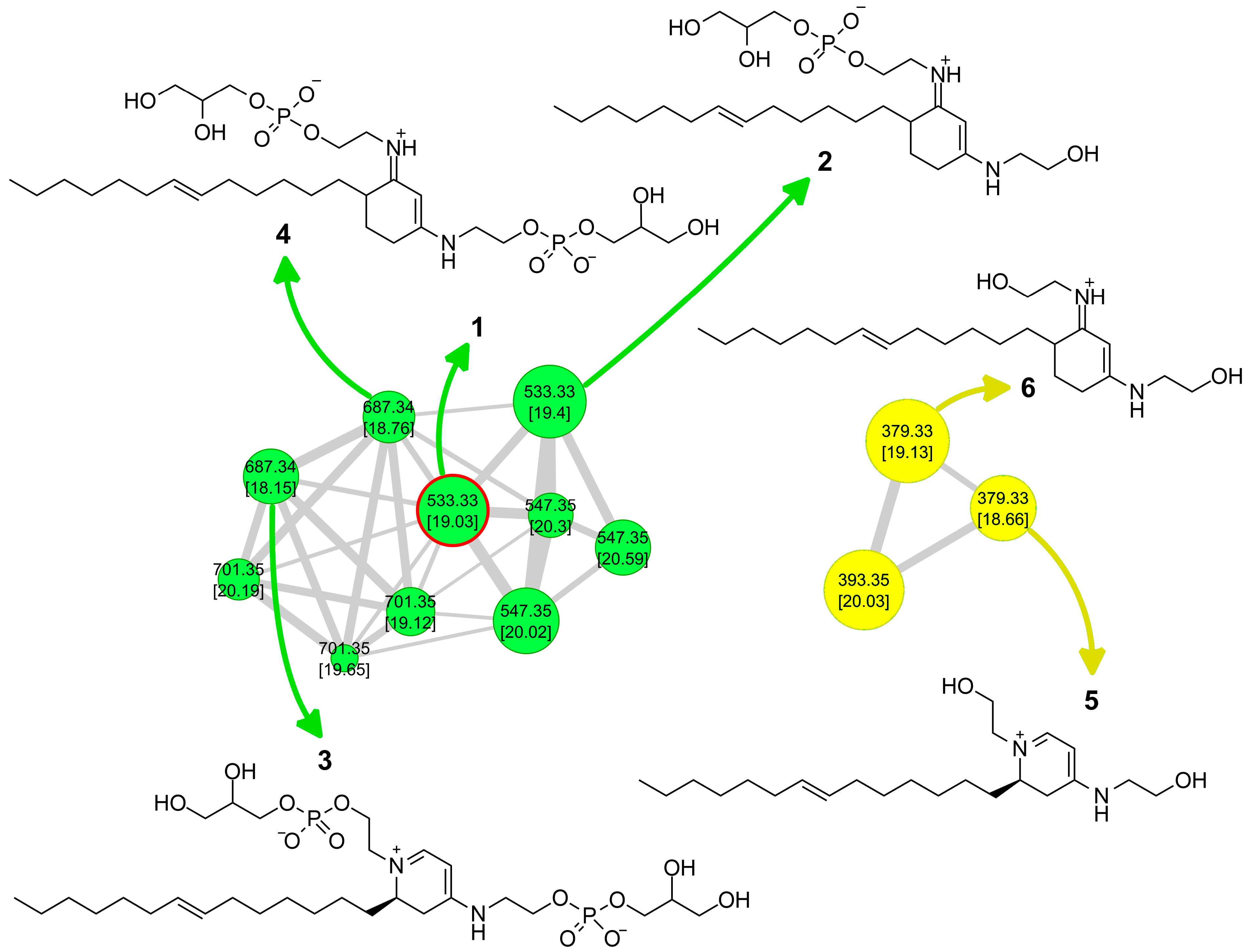
2.1. Extraction, LC-HRMS/MS Analysis, and Construction of the Molecular Network
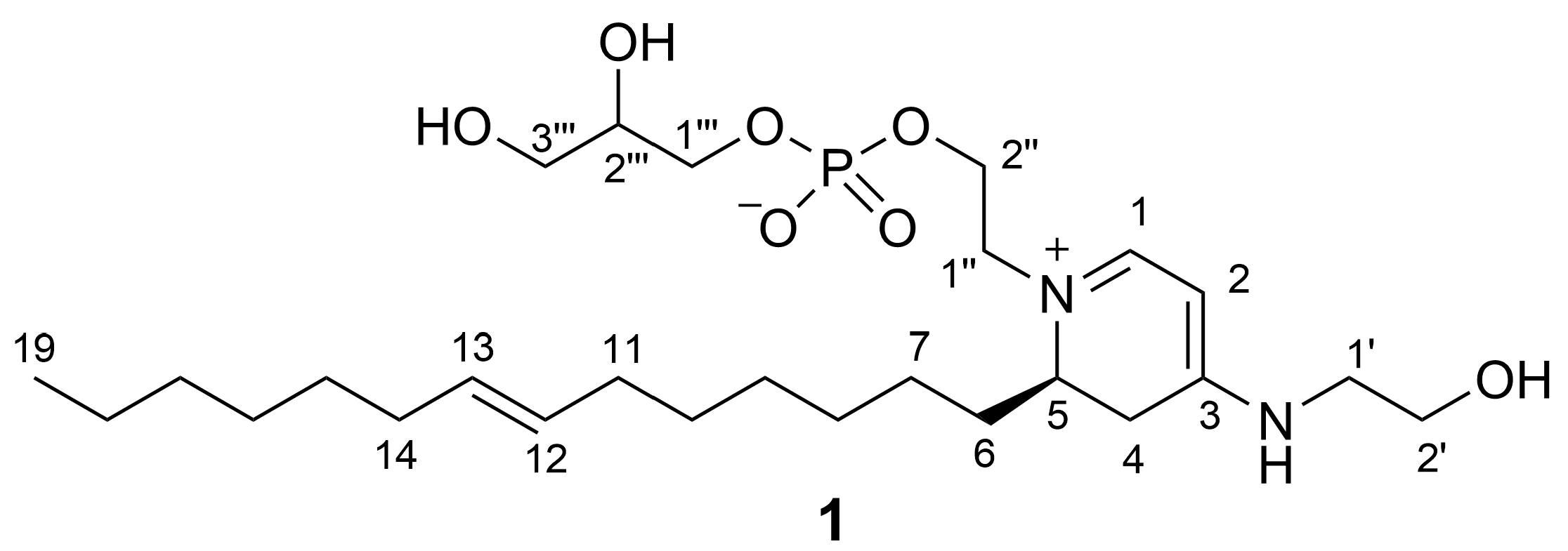
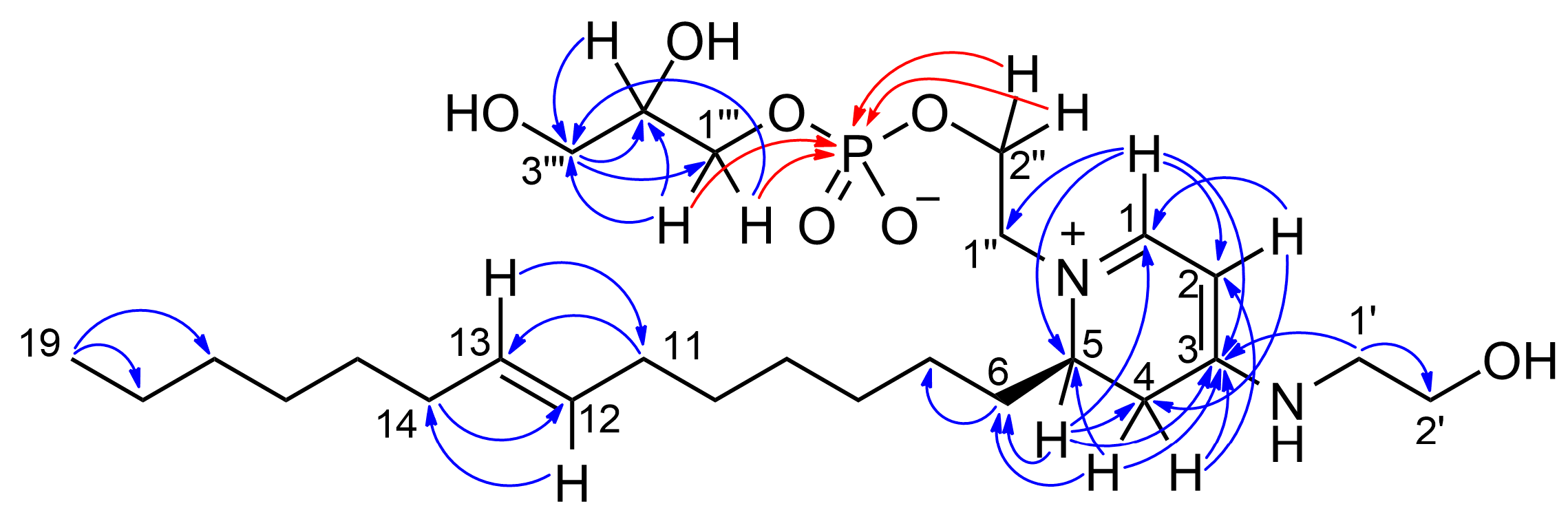
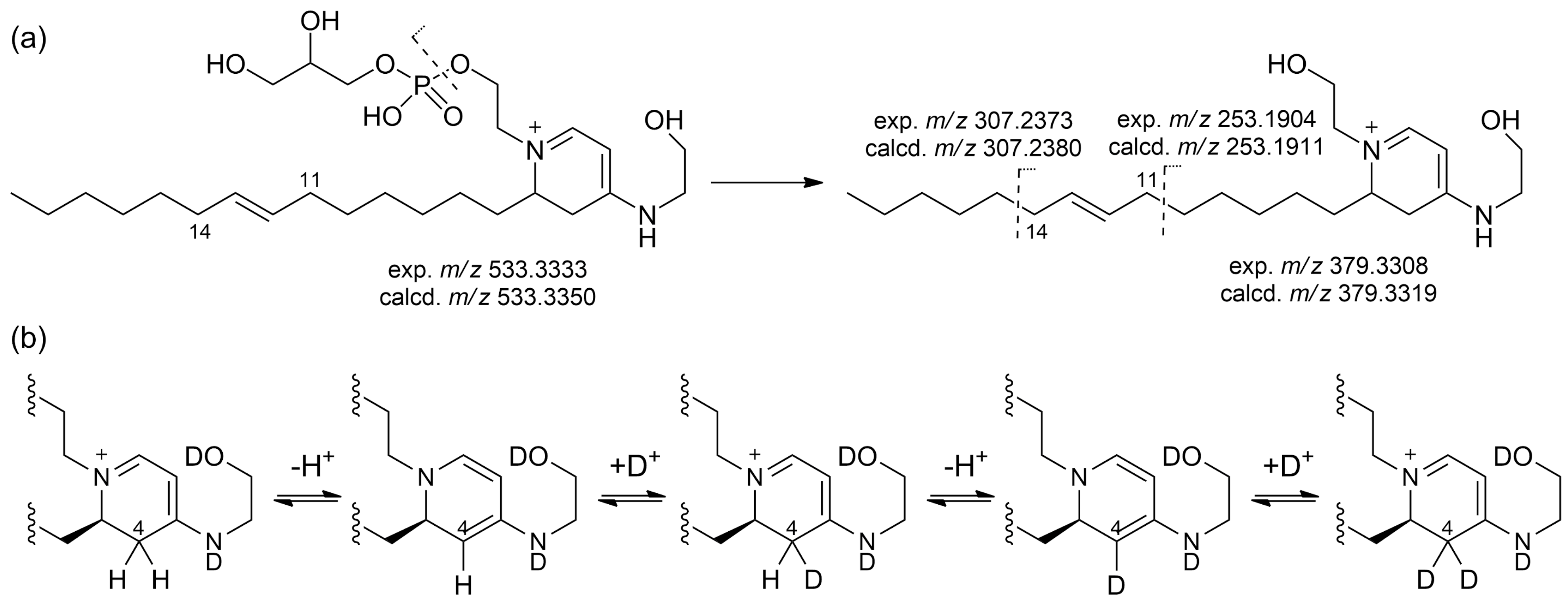
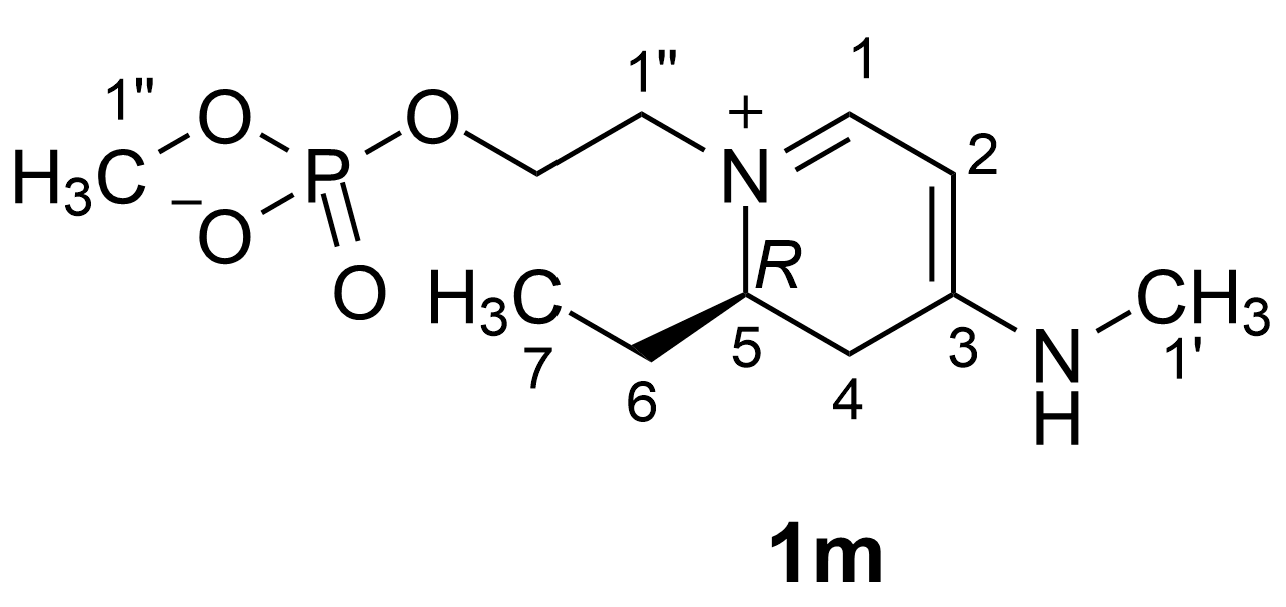
2.2. Isolation and Structure Elucidation of Favilipid A (1)
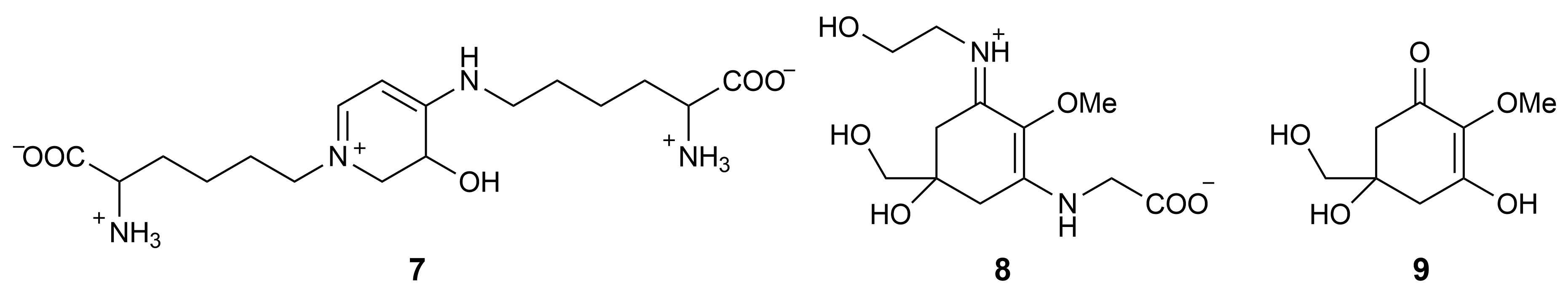
2.3. Other Favilipids
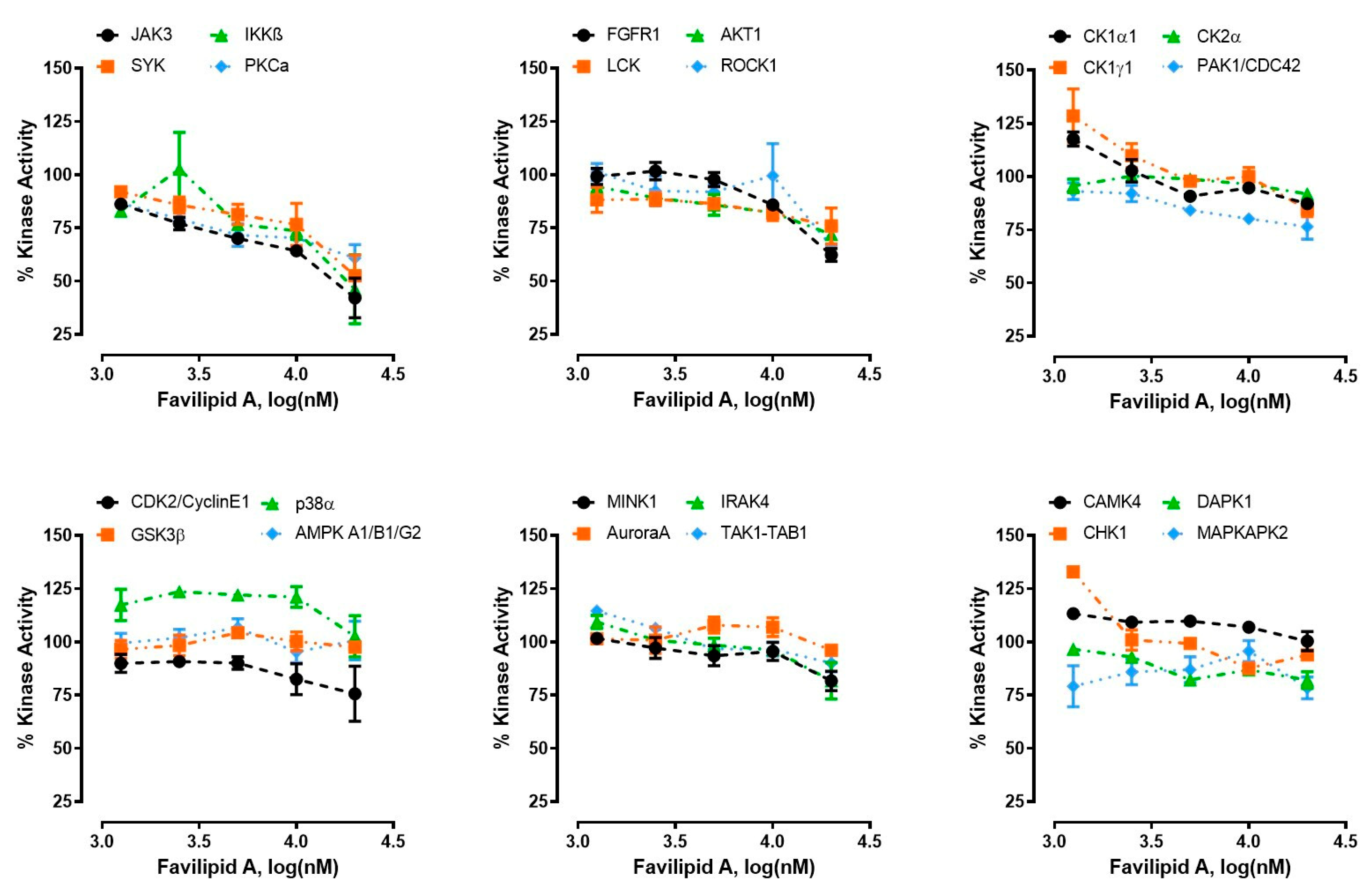
2.4. Kinase Activity Assays
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Collection, Extraction, and Isolation
4.3. LC-HRMS and LC-HRMS/MS
4.4. LC-HRMS/MS Data Processing and Molecular Networking
4.5. Computational Details
4.6. Kinase Activity Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shang, J.; Hu, B.; Wang, J.; Zhu, F.; Kang, Y.; Li, D.; Sun, H.; Kong, D.-X.; Hou, T. Cheminformatic Insight into the Differences between Terrestrial and Marine Originated Natural Products. J. Chem. Inf. Model. 2018, 58, 1182–1193. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luz-zatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [Green Version]
- Teta, R.; Della Sala, G.; Esposito, G.; Via, C.W.; Mazzoccoli, C.; Piccoli, C.; Bertin, M.J.; Costantino, V.; Mangoni, A. A joint molecular networking study of a Smenospongia sponge and a cyanobacterial bloom revealed new antiproliferative chlorinated polyketides. Org. Chem. Front. 2019, 6, 1762–1774. [Google Scholar] [CrossRef]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2022, 39, 1122–1171. [Google Scholar] [CrossRef]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2020, 37, 175–223. [Google Scholar] [CrossRef]
- Bodor, A.; Bounedjoum, N.; Vincze, G.E.; Erdeiné Kis, Á.; Laczi, K.; Bende, G.; Szilágyi, Á.; Kovács, T.; Perei, K.; Rákhely, G. Challenges of unculturable bacteria: Environmental perspectives. Rev. Environ. Sci. Biotechnol. 2020, 19, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Lehnert, H.; Soest, R.W. Van Noth Jamaican Deep Fore-reef Sponges. Beaufortia 1996, 46, 53–81. [Google Scholar]
- Scarpato, S.; Teta, R.; Della Sala, G.; Pawlik, J.R.; Costantino, V.; Mangoni, A. New Tricks with an Old Sponge: Feature-Based Molecular Networking Led to Fast Identification of New Stylissamide L from Stylissa caribica. Mar. Drugs 2020, 18, 443. [Google Scholar] [CrossRef]
- Olivon, F.; Grelier, G.; Roussi, F.; Litaudon, M.; Touboul, D. MZmine 2 Data-Preprocessing to Enhance Molecular Networking Reliability. Anal. Chem. 2017, 89, 7836–7840. [Google Scholar] [CrossRef]
- Olivon, F.; Elie, N.; Grelier, G.; Roussi, F.; Litaudon, M.; Touboul, D. MetGem Software for the Generation of Molecular Net-works Based on the t-SNE Algorithm. Anal. Chem. 2018, 90, 13900–13908. [Google Scholar] [CrossRef]
- Nothias, L.F.; Petras, D.; Schmid, R.; Dührkop, K.; Rainer, J.; Sarvepalli, A.; Protsyuk, I.; Ernst, M.; Tsugawa, H.; Fleischauer, M.; et al. Feature-based molecular networking in the GNPS analysis environment. Nat. Methods 2020, 17, 905–908. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software Environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Dalisay, D.S.; Molinski, T.F. NMR Quantitation of Natural Products at the Nanomole Scale. J. Nat. Prod. 2009, 72, 739–744. [Google Scholar] [CrossRef] [Green Version]
- Shindy, H.A. Fundamentals in the chemistry of cyanine dyes: A review. Dye. Pigment. 2017, 145, 505–513. [Google Scholar] [CrossRef]
- Anderson, A.G.; Daugs, E.D.; Kao, L.G.; Wang, J.F. Dicyclopenta[ef,kl]heptalene (azupyrene) chemistry. Jutz synthesis byproducts. Synthesis and thermal isomerization of 1-methylazupyrene. J. Org. Chem. 1986, 51, 2961–2965. [Google Scholar] [CrossRef]
- SciFinder, Chemical Abstracts Service, Columbus, OH, USA, Spectrum ID CNUC00015518, RN 52950-92-2. Available online: https://scifinder.cas.org (accessed on 28 December 2022).
- Malhotra, S.S.; Whiting, M.C. Researches on polyenes. Part VII. The preparation and electronic absorption spectra of ho-mologous series of simple cyanines, merocyanines, and oxonols. J. Chem. Soc. 1960, 3812–3822. [Google Scholar] [CrossRef]
- Badertscher, M.; Bühlmann, P.; Pretsch, E. Structure Determination of Organic Compounds, 4th ed.; Springer: Berlin/Heidelberg, Germany, 2009; ISBN 978-3-540-93809-5. [Google Scholar]
- Hsu, F. Electrospray ionization with higher-energy collision dissociation tandem mass spectrometry toward characterization of ceramides as [M+Li ]+ ions: Mechanisms of fragmentation and structural identification. Anal. Chim. Acta 2021, 1142, 221–234. [Google Scholar] [CrossRef]
- Grauso, L.; Teta, R.; Esposito, G.; Menna, M.; Mangoni, A. Computational prediction of chiroptical properties in structure elucidation of natural products. Nat. Prod. Rep. 2019, 36, 1005–1030. [Google Scholar] [CrossRef]
- Bruhn, T.; Schaumlöffel, A.; Hemberger, Y.; Pescitelli, G. SpecDis Version 1.71, Berlin, Germany. 2017. Available online: http:/specdis-software.jimdo.com (accessed on 28 December 2022).
- Grimme, S.; Neese, F. Double-hybrid density functional theory for excited electronic states of molecules. J. Chem. Phys. 2007, 127, 154116. [Google Scholar] [CrossRef]
- Cheng, R.; Feng, Q.; Argirov, O.K.; Ortwerth, B.J. Structure elucidation of a novel yellow chromophore from human lens protein. J. Biol. Chem. 2004, 279, 45441–45449. [Google Scholar] [CrossRef] [Green Version]
- Orfanoudaki, M.; Hartmann, A.; Alilou, M.; Gelbrich, T.; Planchenault, P.; Derbré, S.; Schinkovitz, A.; Richomme, P.; Hensel, A.; Ganzera, M. Absolute configuration of mycosporine-like amino acids, their wound healing properties and in vitro anti-aging effects. Mar. Drugs 2020, 18, 35. [Google Scholar] [CrossRef]
- Chen, M.; Rubin, G.M.; Jiang, G.; Raad, Z.; Ding, Y. Biosynthesis and Heterologous Production of Mycosporine-Like Amino Acid Palythines. J. Org. Chem. 2021, 86, 11160–11168. [Google Scholar] [CrossRef]
- Duong-Ly, K.C.; Peterson, J.R. The human kinome and kinase inhibition. Curr. Protoc. Pharmacol. 2013, 60, 2.9.1–2.9.14. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, D.M.; Bonelli, M.; Gadina, M.; O’Shea, J.J. Type I/II cytokines, JAKs, and new strategies for treating autoimmune diseases. Nat. Rev. Rheumatol. 2016, 12, 25–36. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Xia, Y.; Parker, A.S.; Verma, I.M. IKK biology. Immunol. Rev. 2012, 246, 239–253. [Google Scholar] [CrossRef] [Green Version]
- Geahlen, R.L. Getting Syk: Spleen tyrosine kinase as a therapeutic target. Trends Pharmacol. Sci. 2014, 35, 414–422. [Google Scholar] [CrossRef] [Green Version]
- Pei, H.; He, L.; Shao, M.; Yang, Z.; Ran, Y.; Li, D.; Zhou, Y.; Tang, M.; Wang, T.; Gong, Y.; et al. Discovery of a highly selective JAK3 inhibitor for the treatment of rheumatoid arthritis. Sci. Rep. 2018, 8, 5273. [Google Scholar] [CrossRef] [Green Version]
- Chambers, M.C.; Maclean, B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J.; et al. A cross-platform toolkit for mass spectrometry and proteomics. Nat. Biotechnol. 2012, 30, 918–920. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
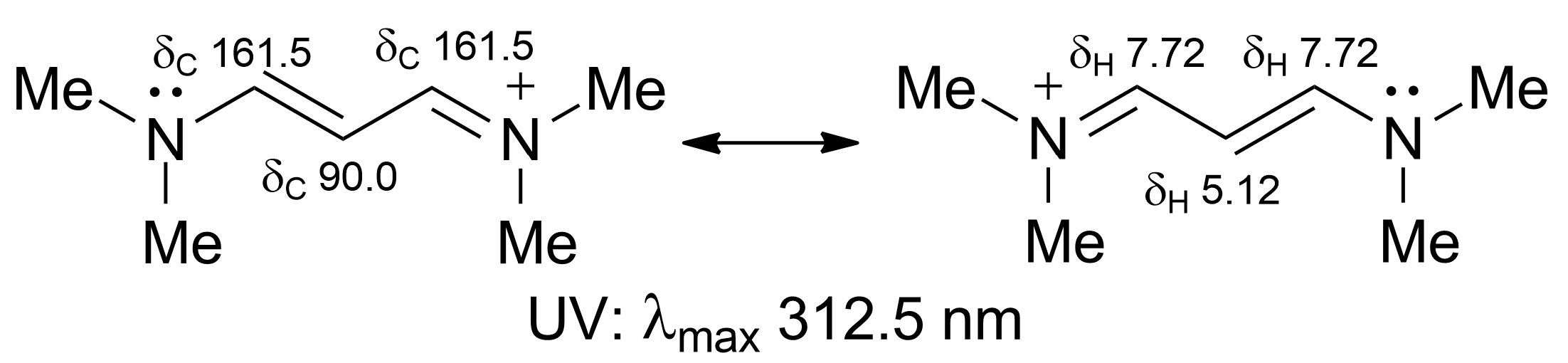{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pos. | δC, Type | δH, Mult (J in Hz) | ROESY | 13C and 31P HMBC | |
|---|---|---|---|---|---|
| 1 | 159.3, CH | 7.63, d (6.6) | H-2, H1″a | C-1, C-3, C-5, C-1″ | |
| 2 | 86.4, CH | 5.28, d (6.6) | H-1, H2-1′ | C-1, C-4 | |
| 3 | 165.4, C | - | |||
| 4 | 32.4, CH2 | a | 3.29, dd (17.0, 7.1) | H-4b | C-3, C-5, C-6 |
| b | 2.67, br. d (17.0) | H-4a, H-5 | C-2, C-3 | ||
| 5 | 56.1, CH | 3.87, br. quartet (7.1) | H-4b, H2-6 | C-1, C-3, C-4, C-6 | |
| 6 | 28.9, CH2 | 1.67, quartet (7.1) | H-5, H-1″b | C-4, C-7, C-8 | |
| 7 | 26.4, CH2 | a | 1.38, m | ||
| b | 1.32, m | ||||
| 8, 9, 16 | 30.1, CH2 | 1.34–1.29, m | |||
| 10 | 30.5, CH2 | 1.34, m | H-12 | ||
| 11 | 33.5, CH2 | 1.98, m | H-13 | C-10, C-12, C-13 | |
| 12 | 131.4, CH | 5.39, m | H2-10, H-14 | C-11, C-14 | |
| 13 | 131.4, CH | 5.39, m | H2-13, H-11 | C-11, C-14 | |
| 14 | 33.5, CH2 | 1.98, m | H-12 | C-12, C-13, C-15 | |
| 15 | 30.5, CH2 | 1.34, m | H-13 | ||
| 17 | 32.9, CH2 | 1.28, m | |||
| 18 | 23.6, CH2 | 1.31, m | H3-19 | ||
| 19 | 14.3, CH3 | 0.90, t (7.0) | H2-18 | C-17, C-18 | |
| 1′ | 47.4, CH2 | 3.46, t (5.3) | H-2, H2-2′ | C-3, C-2′ | |
| 2′ | 60.3, CH2 | 3.75, t (5.3) | H2-1′ | ||
| 1″ | 56.6, CH2 | a | 3.72, ddd (14.6, 7.8, 3.3) | H-1, H-1″b, H-2″b | |
| b | 3.66, ddd (14.6, 5.0, 3.6) | H2-6, H-1″a, H-2″b | |||
| 2″ | 64.6, CH2 | a | 4.10, m | H-2″b | P |
| b | 4.00, m | H-1″a, H-1″b, H-2″a | P | ||
| 1‴ | 67.7, CH2 | a | 3.89, m | H-1‴b | C-3‴, P |
| b | 3.84, m | H-1‴a | C-2‴, C-3‴, P | ||
| 2‴ | 72.6, CH | 3.75, m | H-3‴b | C-1‴, C-3‴ | |
| 3‴ | 63.9, CH2 | a | 3.59, dd (11.4, 5.3) | H-3‴b | C-1‴, C-2‴ |
| b | 3.54, dd (11.4, 5.7) | H-2‴, H-3‴a | C-1‴, C-2‴ |
| Kinase | % Inhibition | IC50 (μM) | 95% CI (μM) |
|---|---|---|---|
| JAK3 | 58% | 3.5 | 2.0–5.9 |
| IKKβ | 54% | 6.0 | 1.4–77.8 |
| SYK | 48% | 8.3 | 4.9–13.8 |
| PKCα | 40% | 3.3 | 2.1–5.0 |
| FGFR1 | 38% | 10.7 | 8.8–12.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scarpato, S.; Teta, R.; De Cicco, P.; Borrelli, F.; Pawlik, J.R.; Costantino, V.; Mangoni, A. Molecular Networking Revealed Unique UV-Absorbing Phospholipids: Favilipids from the Marine Sponge Clathria faviformis. Mar. Drugs 2023, 21, 58. https://doi.org/10.3390/md21020058
Scarpato S, Teta R, De Cicco P, Borrelli F, Pawlik JR, Costantino V, Mangoni A. Molecular Networking Revealed Unique UV-Absorbing Phospholipids: Favilipids from the Marine Sponge Clathria faviformis. Marine Drugs. 2023; 21(2):58. https://doi.org/10.3390/md21020058
Chicago/Turabian StyleScarpato, Silvia, Roberta Teta, Paola De Cicco, Francesca Borrelli, Joseph R. Pawlik, Valeria Costantino, and Alfonso Mangoni. 2023. "Molecular Networking Revealed Unique UV-Absorbing Phospholipids: Favilipids from the Marine Sponge Clathria faviformis" Marine Drugs 21, no. 2: 58. https://doi.org/10.3390/md21020058
APA StyleScarpato, S., Teta, R., De Cicco, P., Borrelli, F., Pawlik, J. R., Costantino, V., & Mangoni, A. (2023). Molecular Networking Revealed Unique UV-Absorbing Phospholipids: Favilipids from the Marine Sponge Clathria faviformis. Marine Drugs, 21(2), 58. https://doi.org/10.3390/md21020058