
Botryllin, a Novel Antimicrobial Peptide from the Colonial Ascidian Botryllus schlosseri




Abstract
:1. Introduction
2. Results
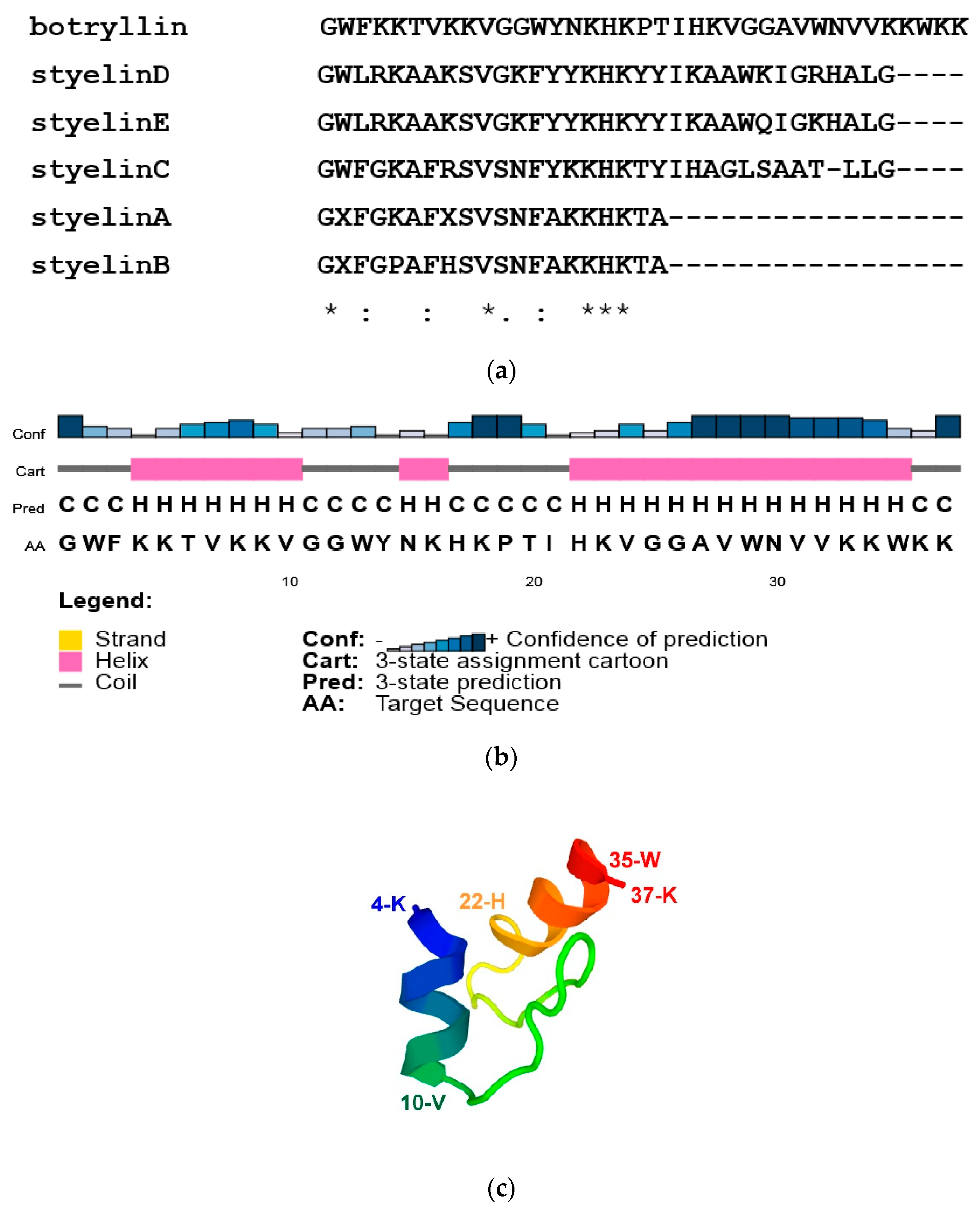
2.1. Botryllin Identification
2.2. Transcription Rate of the Botryllin Gene
2.3. Botryllin Has an Antimicrobial/Antimycotic Activity
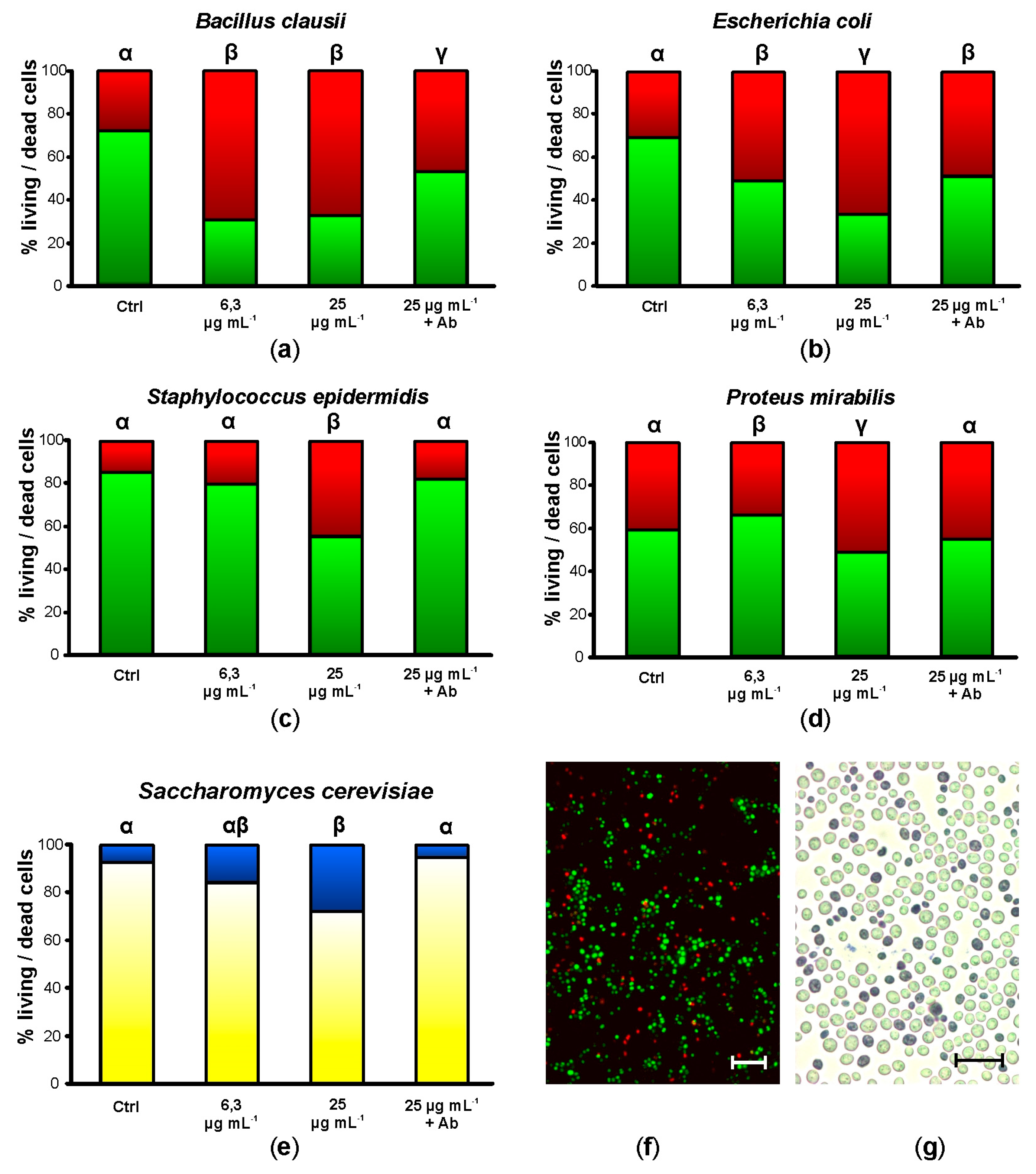
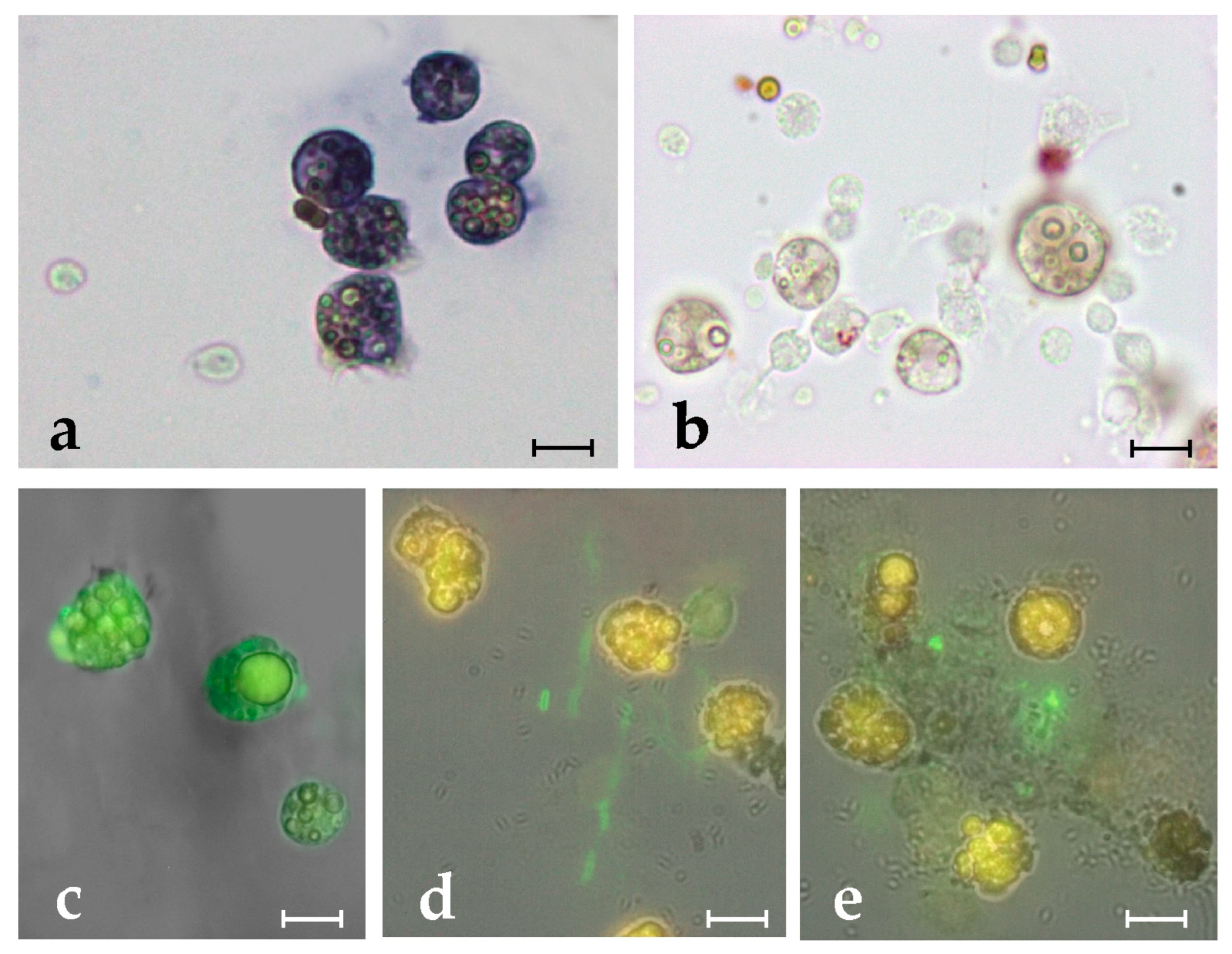
2.4. Botryllin Increases the Mortality of Bacterial and Yeast Cells
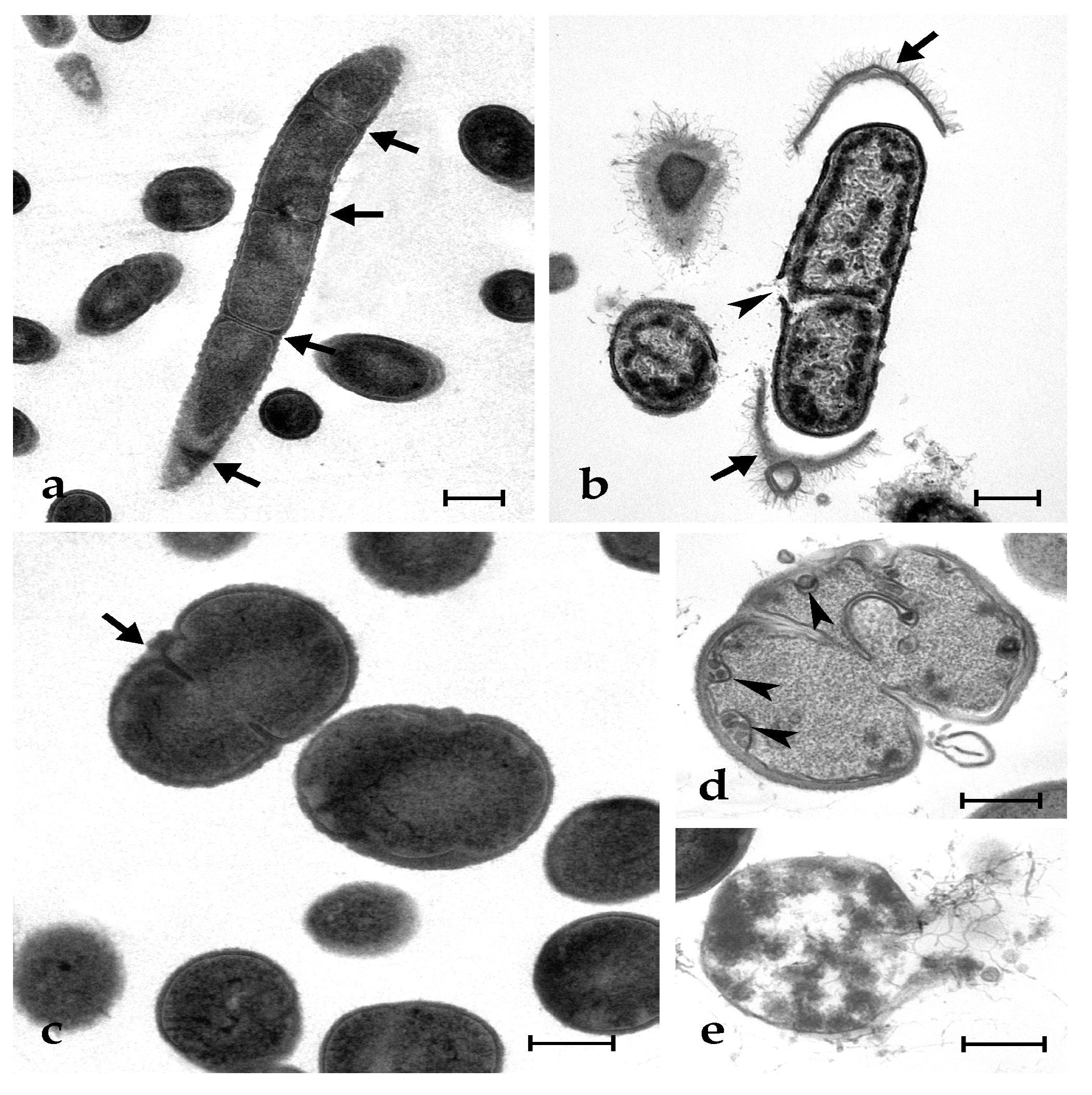
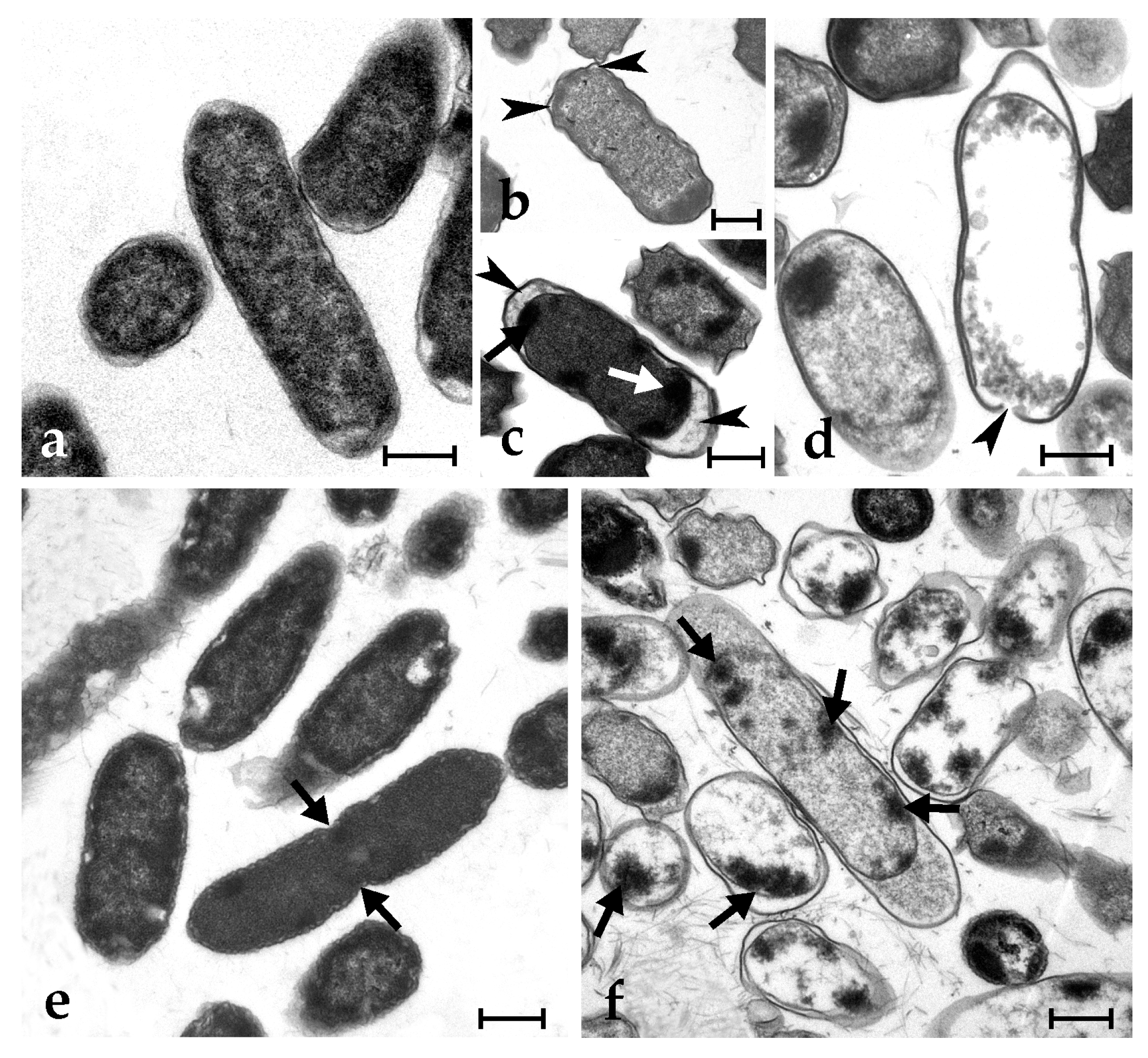
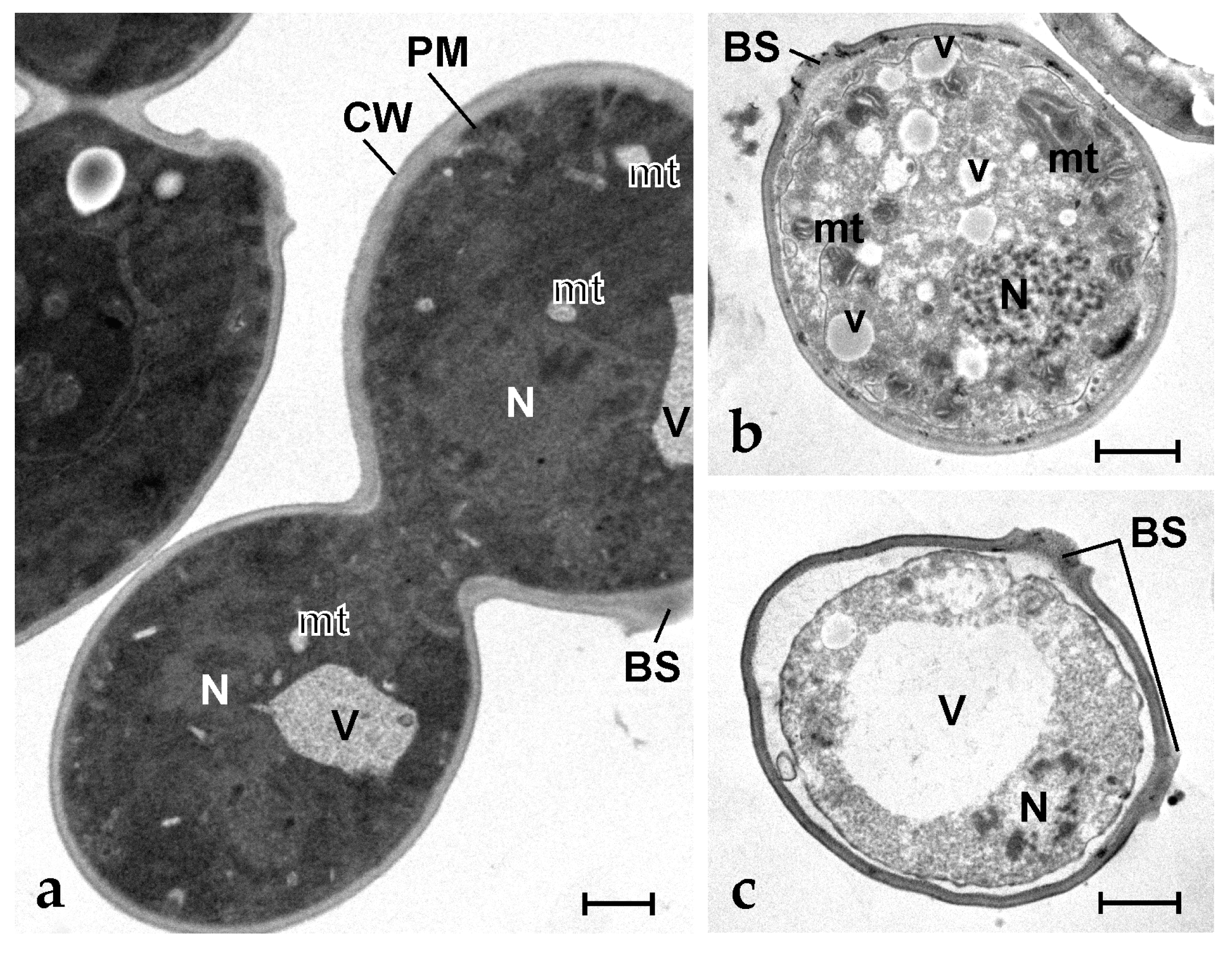
2.5. Botryllin Causes Irreversible Morphological Alterations of Microbial and Yeast Cells
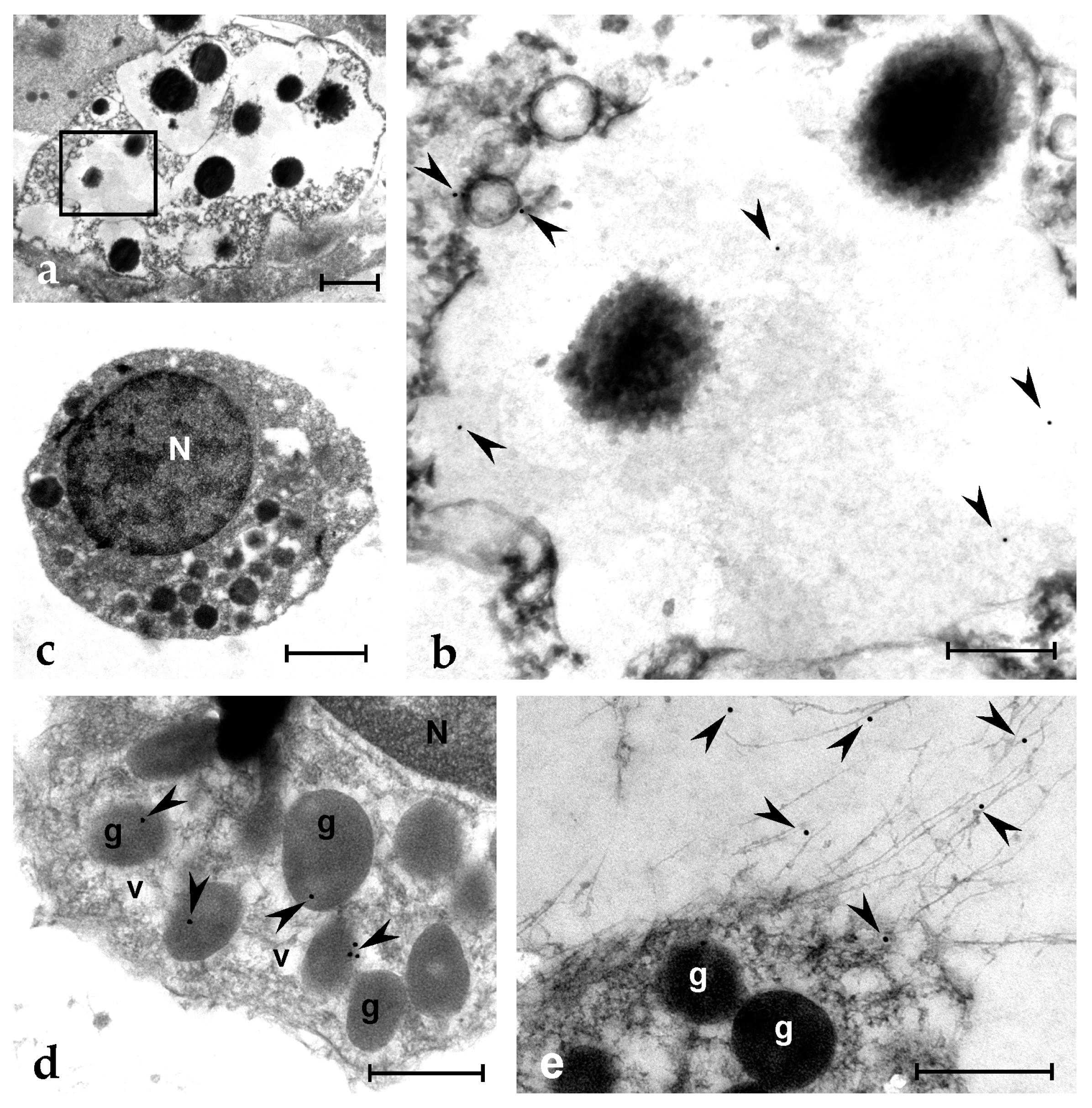
2.6. Botryllin Is Synthesised by MC, Stored Inside Their Granules and Released upon MC Degranulation
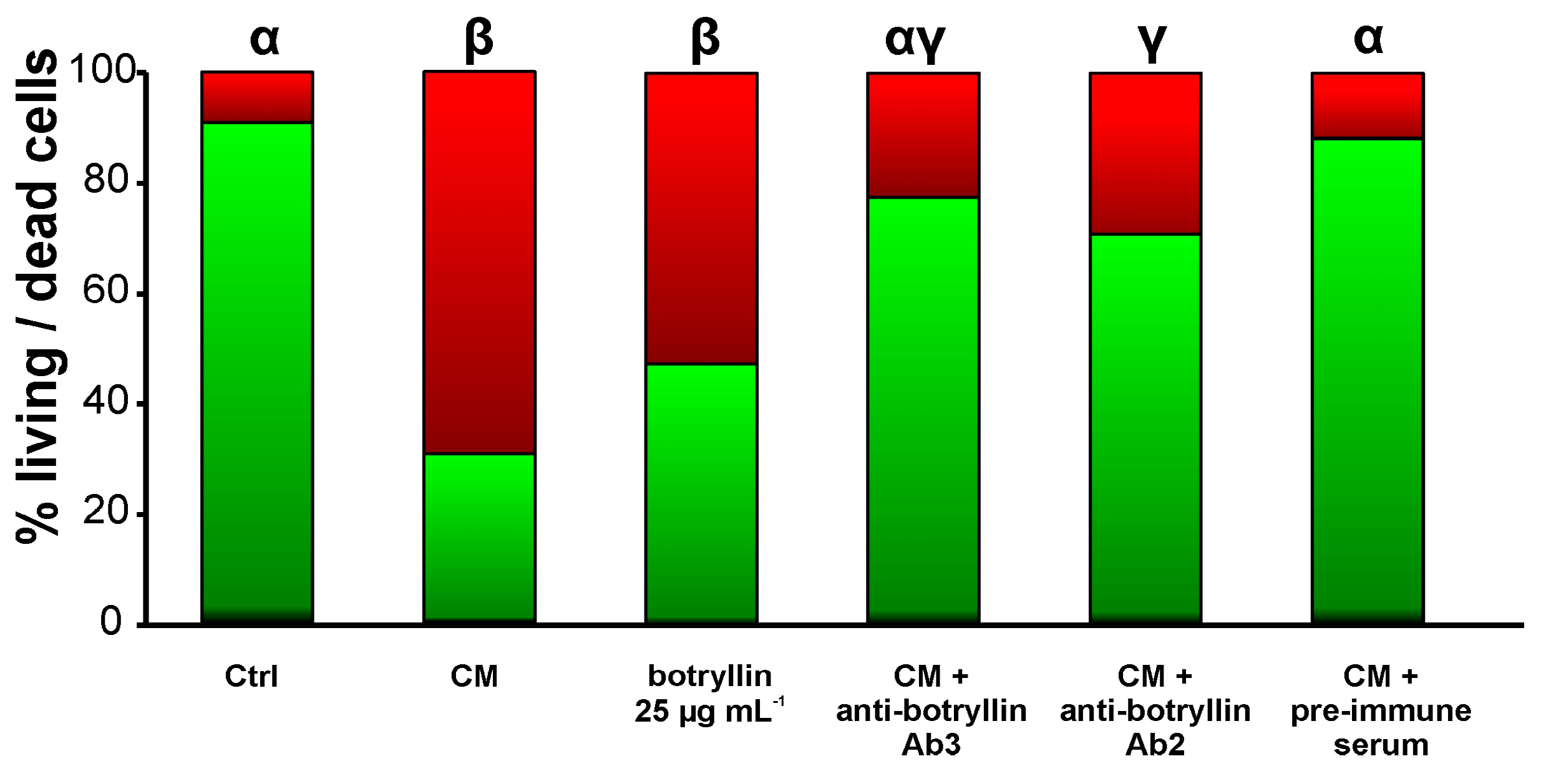
2.7. B. clausii-Conditioned Medium (CM) Has Biocidal Activity That Is Inhibited by Anti-Botryllin Antibodies
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Haemocyte Collection
4.3. Primer Design, RNA Extraction, cDNA Synthesis, Cloning, and Sequencing
4.4. Three-Dimensional Structure of Botryllin
4.5. Quantitative Real-Time PCT (qRT-PCR)
4.6. In Situ Hybridization (ISH)
4.7. Synthetic Peptide and Specific Antibodies
4.8. Immunocytochemistry
4.9. Minimal Inhibitory Concentration (MIC) and Minimal Bactericidal Concentration (MBC) Determination
4.10. Microbial Cell Mortality Assay
4.11. Temperature, pH and Salinity Stability Assays
4.12. Electron Microscopy
4.13. Preparation of Conditioned Medium (CM)
4.14. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gould, I.M. Stewardship of antibiotic use and resistance surveillance: The international scene. J. Hosp. Infect. 1999, 43, S253–S260. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, J. Review on Antimicrobial Resistance. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations; HM Government and Wellcome Trust: London, UK, 2016. [Google Scholar]
- Giuliani, A.; Pirri, G.; Nicoletto, S.F. Antimicrobial peptides: An overview of a promising class of therapeutics. Cent. Eur. J. Biol. 2007, 2, 1–33. [Google Scholar] [CrossRef]
- Bierbaum, G.; Sahl, H.G. Induction of autolysis of staphylococci by the basic peptide antibiotics Pep 5 and nisin and their influence on the activity of autolytic enzymes. Arch. Microbiol. 1985, 141, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Westerhoff, H.V.; Juretić, D.; Hendler, R.W.; Zasloff, M. Magainins and the disruption of membrane-linked free-energy transduction. Proc. Natl. Acad. Sci. USA 1989, 86, 6597–6601. [Google Scholar] [CrossRef] [Green Version]
- Matsuzaki, K. Why and how are peptide-lipid interactions utilized for self-defense? Magainins and tachyplesins as archetypes. Biochim. Biophys. Acta 1999, 1462, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Weiss, T.M.; Lehrer, R.I.; Huang, H.W. Crystallization of antimicrobial pores in membranes: Magainin and protegrin. Biophys J. 2000, 79, 2002–2009. [Google Scholar] [CrossRef] [Green Version]
- Kragol, G.; Lovas, S.; Varadi, G.; Condie, B.A.; Hoffmann, R.; Otvos, L., Jr. The antibacterial peptide pyrrhocoricin inhibits the ATPase actions of DnaK and prevents chaperone-assisted protein folding. Biochemistry 2001, 40, 3016–3026. [Google Scholar] [CrossRef]
- Lazzaro, B.P.; Zasloff, M.; Rolff, J. Antimicrobial peptides: Application informed by evolution. Science 2020, 368, eaau5480. [Google Scholar] [CrossRef]
- Shai, Y. Mechanism of the binding, insertion and destabilization of phospholipid bilayer membranes by alpha-helical antimicrobial and cell non-selective membrane-lytic peptides. BiochimBiophys Acta 1999, 1462, 55–70. [Google Scholar]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial peptides: Classification, design, application and research progress in multiple fields. Front. Microbiol. 2020, 11, 582779. [Google Scholar] [CrossRef]
- Rabel, D.; Charlet, M.; Ehret-Sabatier, L.; Cavicchioli, L.; Cudic, M.; Otvos, L., Jr.; Bulet, P. Primary structure and in vitro antibacterial properties of the Drosophila melanogaster attacin C pro-domain. J. Biol. Chem. 2004, 279, 14853–14859. [Google Scholar] [CrossRef] [PubMed]
- Fjell, C.D.; Hiss, J.A.; Hancock, R.E.; Schneider, G. Designing antimicrobial peptides: Form follows function. Nat. Rev. Drug Discov. 2011, 11, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Bahar, A.A.; Ren, D. Antimicrobial peptides. Pharmaceuticals 2013, 6, 1543–1575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E.; Sahl, H.G. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Deslouches, B.; Di, Y.P. Antimicrobial peptides with selective antitumor mechanisms: Prospect for anticancer applications. Oncotarget. 2017, 8, 46635–46651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bevins, C.L.; Salzman, N.H. Paneth cells, antimicrobial peptides and maintenance of intestinal homeostasis. Nat. Rev. Microbiol. 2011, 9, 356–368. [Google Scholar] [CrossRef]
- Login, F.H.; Balmand, S.; Vallier, A.; Vincent-Monégat, C.; Vigneron, A.; Weiss-Gayet, M.; Rochat, D.; Heddi, A. Antimicrobial peptides keep insect endosymbionts under control. Science 2011, 334, 362–365. [Google Scholar] [CrossRef]
- Franzenburg, S.; Walter, J.; Künzel, S.; Wang, J.; Baines, J.F.; Bosch, T.C.; Fraune, S. Distinct antimicrobial peptide expression determines host species-specific bacterial associations. Proc. Natl. Acad. Sci. USA 2013, 110, E3730–E3738. [Google Scholar] [CrossRef] [Green Version]
- Cullen, T.W.; Schofield, W.B.; Barry, N.A.; Putnam, E.E.; Rundell, E.A.; Trent, M.S.; Degnan, P.H.; Booth, C.J.; Yu, H.; Goodman, A.L. Gut microbiota. Antimicrobial peptide resistance mediates resilience of prominent gut commensals during inflammation. Science 2015, 347, 170–175. [Google Scholar] [CrossRef] [Green Version]
- Arnold, M.F.F.; Shabab, M.; Penterman, J.; Boehme, K.L.; Griffitts, J.S.; Walker, G.C. Genome-wide sensitivity analysis of the microsymbiont Sinorhizobiummeliloti to symbiotically important, defensin-like host peptides. mBio 2017, 8, e01060-17. [Google Scholar] [CrossRef] [PubMed]
- Leal, M.C.; Puga, J.; Serôdio, J.; Gomes, N.C.M.; Calado, R. Trends in the discovery of new marine natural products from invertebrates over the last two decades—Where and what are we bioprospecting? PLoS ONE 2012, 7, e30580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datta, D.; Talapatra, S.; Swarnakar, S. Bioactive compounds from marine invertebrates for potential medicines—An overview. Int. Lett. Nat. Sci. 2015, 34, 42–61. [Google Scholar] [CrossRef]
- Romano, G.; Almeida, M.; Varela Coelho, A.; Cutignano, A.; Gonçalves, L.G.; Hansen, E.; Khnykin, D.; Mass, T.; Ramšak, A.; Rocha, M.S.; et al. Biomaterials and bioactive natural products from marine invertebrates: From basic research to innovative applications. Mar. Drugs 2022, 20, 219. [Google Scholar] [CrossRef]
- Tincu, J.A.; Taylor, S.W. Antimicrobial peptides from marine invertebrates. Antimicrob. Agents Chemother. 2004, 48, 3645–3654. [Google Scholar] [CrossRef] [Green Version]
- Delsuc, F.; Brinkmann, H.; Chourrout, D.; Philippe, H. Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature 2006, 439, 965–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casertano, M.; Menna, M.; Imperatore, C. The ascidian-derived metabolites with antimicrobial properties. Antibiotics 2020, 9, 510. [Google Scholar] [CrossRef]
- Kwan, J.C.; Donia, M.S.; Han, A.W.; Hirose, E.; Haygood, M.G.; Schmidt, E.W. Genome streamlining and chemical defense in a coral reef symbiosis. Proc. Natl. Acad. Sci. USA 2012, 109, 20655–20660. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, E.W.; Donia, M.S.; McIntosh, J.A.; Fricke, W.F.; Ravel, J. Origin and variation of tunicate secondary metabolites. J. Nat Prod. 2012, 75, 295–304. [Google Scholar] [CrossRef] [Green Version]
- Lehrer, R.I.; Andrew Tincu, J.; Taylor, S.W.; Menzel, L.P.; Waring, A.J. Natural peptide antibiotics from tunicates: Structures, functions and potential uses. Integr. Comp. Biol. 2003, 43, 313–322. [Google Scholar] [CrossRef]
- Zhao, C.; Liaw, L.; Lee, I.H.; Lehrer, R.I. cDNA cloning of three cecropin-like antimicrobial peptides (styelins) from the tunicate, Styela clava. FEBS Lett. 1997, 412, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.H.; Zhao, C.; Nguyen, T.; Menzel, L.; Waring, A.J.; Sherman, M.A.; Lehrer, R.I. Clavaspirin, an antibacterial and haemolytic peptide from Styela Clava. J. Pept. Res. 2001, 58, 445–456. [Google Scholar] [PubMed]
- Menzel, L.P.; Lee, I.H.; Sjostrand, B.; Lehrer, R.I. Immunolocalization of clavanins in Styela clava hemocytes. Dev. Comp. Immunol. 2002, 26, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Tincu, J.A.; Menzel, L.P.; Azimov, R.; Sands, J.; Hong, T.; Waring, A.J.; Taylor, S.W.; Lehrer, R.I. Plicatamide, an antimicrobial octapeptide from Styela plicata hemocytes. J. Biol. Chem. 2003, 278, 13546–13553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azumi, K.; Yokosawa, H.; Ishii, S. Halocyamines: Novel antimicrobial tetrapeptide-like substances isolated from the hemocytes of the solitary ascidian Halocynthia Roretzi. Biochemistry 1990, 29, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Ballarin, L. Ascidian cytotoxic cells: State of the art and research perspectives. ISJ-Invertebr. Surviv. J. 2012, 9, 1–6. [Google Scholar]
- Fedders, H.; Michalek, M.; Grötzinger, J.; Leippe, M. An exceptional salt-tolerant antimicrobial peptide derived from a novel gene family of haemocytes of the marine invertebrate Ciona intestinalis. Biochem. J. 2008, 416, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Fedders, H.; Leippe, M. A Reverse search for antimicrobial peptides in Ciona intestinalis: Identification of a gene family expressed in hemocytes and evaluation of activity. Dev. Comp. Immunol. 2008, 32, 286–298. [Google Scholar] [CrossRef]
- Fedders, H.; Podschun, R.; Leippe, M. The antimicrobial peptide Ci-MAM-A24 is highly active against multidrug-resistant and anaerobic bacteria pathogenic for humans. Int. J. Antimicrob. Agents 2010, 36, 264–266. [Google Scholar] [CrossRef] [Green Version]
- Di Bella, M.D.; Fedders, H.; Leippe, M.; De Leo, G. A preliminary study on antimicrobial peptides in the naturally damaged tunic of Ciona intestinalis (Tunicata). In Science Against Microbial Pathogens: Communicating Current Research and Technological Advances; Méndez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2011; pp. 1003–1007. [Google Scholar]
- Tsukimoto, M.; Nagaoka, M.; Shishido, Y.; Fujimoto, J.; Nishisaka, F.; Matsumoto, S.; Harunari, E.; Imada, C.; Matsuzaki, T. Bacterial production of the tunicate-derived antitumor cyclic depsipeptidedidemnin B. J. Nat. Prod. 2011, 74, 2329–2331. [Google Scholar] [CrossRef]
- Xu, Y.; Kersten, R.D.; Nam, S.J.; Lu, L.; Al-Suwailem, A.M.; Zheng, H.; Fenical, W.; Dorrestein, P.C.; Moore, B.S.; Qian, P.Y. Bacterial biosynthesis and maturation of the didemnin anti-cancer agents. J. Am. Chem. Soc. 2012, 134, 8625–8632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramesh, C.; Tulasi, B.R.; Raju, M.; Thakur, N.; Dufossé, L. Marine natural products from tunicates and their associated microbes. Mar. Drugs 2021, 19, 308. [Google Scholar] [CrossRef] [PubMed]
- Franchi, N.; Ballarin, L. Immunity in Protochordates: The tunicate perspective. Front. Immunol. 2017, 8, 674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballarin, L.; Cima, F. Cytochemical properties of Botryllus schlosseri haemocytes: Indications for morpho-functional characterisation. Eur. J. Histochem. 2005, 49, 255–264. [Google Scholar]
- Franchi, N.; Ballarin, L. Preliminary characterization of complement in a colonial tunicate: C3, Bf and inhibition of C3 opsonic activity by compstatin. Dev. Comp. Immunol. 2014, 46, 430–438. [Google Scholar] [CrossRef]
- Franchi, N.; Ballarin, L. Morula cells as key hemocytes of the lectin pathway of complement activation in the colonial tunicate Botryllus schlosseri. Fish Shellfish Immunol. 2017, 63, 157–164. [Google Scholar]
- Peronato, A.; Franchi, N.; Ballarin, L. Insights into the complement system of tunicates: C3a/C5aR of the colonial ascidian Botryllus schlosseri. Biology 2020, 9, 263. [Google Scholar] [CrossRef]
- Menin, A.; Del Favero, M.; Cima, F.; Ballarin, L. Release of phagocytosis-stimulating factor (s) by morula cells in a colonial ascidian. Mar. Biol. 2005, 148, 225–230. [Google Scholar] [CrossRef]
- Menin, A.; Ballarin, L. Immunomodulatory molecules in the compound ascidian Botryllus schlosseri: Evidence from conditioned media. J. Invertebr. Pathol. 2008, 99, 275–280. [Google Scholar] [CrossRef]
- Vizioli, J.; Salzet, M. Antimicrobial peptides from animals: Focus on invertebrates. Trends Pharmacol. Sci. 2002, 23, 494–496. [Google Scholar] [CrossRef]
- Jenssen, H.; Hamill, P.; Hancock, E.W.R. Peptide antimicrobial agents. Clin. Microbiol. Rev. 2006, 19, 491–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballarin, L.; Cima, F.; Sabbadin, A. Phenoloxidase in the colonial ascidian Botryllus schlosseri (Urochordata: Ascidiacea). Anim. Biol. 1994, 3, 41–48. [Google Scholar]
- Ballarin, L.; Cima, F.; Sabbadin, A. Phenoloxidase and cytotoxicity in the compound ascidian Botryllus schlosseri. Dev. Comp. Immunol. 1998, 22, 479–492. [Google Scholar] [CrossRef]
- Frizzo, A.; Guidolin, L.; Ballarin, L.; Sabbadin, A. Purification and partial characterisation of phenoloxidase from the colonial ascidian Botryllus schlosseri. Mar. Biol. 1999, 135, 483–488. [Google Scholar] [CrossRef]
- Gasparini, F.; Franchi, N.; Spolaore, B.; Ballarin, L. Novel rhamnose-binding lectins from the colonial ascidian Botryllus schlosseri. Dev. Comp. Immunol. 2008, 32, 1177–1191. [Google Scholar] [CrossRef]
- Franchi, N.; Schiavon, F.; Carletto, M.; Gasparini, F.; Bertoloni, G.; Tosatto, S.C.E.; Ballarin, L. Immune roles of a rhamnose-binding lectin in the colonial ascidian Botryllus schlosseri. Immunobiology 2011, 216, 725–736. [Google Scholar] [CrossRef]
- Peronato, A.; Drago, L.; Rothbaecher, U.; Macor, P.; Ballarin, L.; Franchi, N. Complement system and phagocytosis in a colonial protochordate. Dev. Comp. Immunol. 2020, 103, 103530. [Google Scholar] [CrossRef]
- Ballarin, L.; Cima, F.; Sabbadin, A. Histoenzymatic staining and characterization of the colonial ascidian Botryllus schlosseri hemocytes. Boll. Zool. 1993, 60, 19–24. [Google Scholar] [CrossRef]
- Ballarin, L.; Franchini, A.; Ottaviani, E.; Sabbadin, A. Morula cells as the main immunomodulatory haemocytes in ascidians: Evidences from the colonial species Botryllus schlosseri. Biol. Bull. 2001, 201, 59–64. [Google Scholar] [CrossRef]
- Ballarin, L.; Cima, F.; Sabbadin, A. Morula cells and histocompatibility in the colonial ascidian Botryllus schlosseri. Zool. Sci. 1995, 12, 757–764. [Google Scholar] [CrossRef] [Green Version]
- Ballarin, L.; Cima, F.; Floreani, M.; Sabbadin, A. Oxidative stress induces cytotoxicity during rejection reaction in the compound ascidian Botryllus schlosseri. Comp. Biochem. Physiol. 2002, 133C, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Cima, F.; Sabbadin, A.; Ballarin, L. Cellular aspects of allorecognition in the compound ascidian Botryllus schlosseri. Dev. Comp. Immunol. 2004, 28, 881–889. [Google Scholar] [CrossRef] [PubMed]
- Franchi, N.; Ballarin, L.; Peronato, A.; Cima, F.; Grimaldi, A.; Girardello, R.; de Eguileor, M. Functional amyloidogenesis in immunocytes from the colonial ascidian Botryllus schlosseri: Evolutionary perspective. Dev. Comp. Immunol. 2019, 90, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.W.; Craig, A.G.; Fischer, W.H.; Park, M.; Lehrer, R.I. Styelin D, an extensively modified antimicrobial peptide from ascidian hemocytes. J. Biol. Chem. 2000, 275, 38417–38426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, I.H.; Cho, Y.; Lehrer, R.I. Styelins, broad-spectrum antimicrobial peptides from the solitary tunicate, Styela clava. Comp. Biochem. Physiol. 1997, 118B, 515–521. [Google Scholar] [CrossRef]
- Awuah, G.B.; Ramaswamy, H.S.; Economides, A. Thermal processing and quality: Principles and overview. Chem. Eng. Process. 2007, 46, 584–602. [Google Scholar] [CrossRef]
- Lappe, R.; Cladera-Olivera, F.; Dominguez, A.P.M.; Brandelli, A. Kinetics and thermodynamics of thermal inactivation of the antimicrobial peptide cerein 8A. J. Food Eng. 2009, 91, 223–227. [Google Scholar] [CrossRef]
- Goldman, M.J.; Anderson, G.M.; Stolzenberg, E.D.; Kari, U.P.; Zasloff, M.; Wilson, J.M. Human beta-defensin-1 is a salt-sensitive antibiotic in lung that is inactivated in cystic fibrosis. Cell 1997, 88, 553–560. [Google Scholar] [CrossRef] [Green Version]
- Sabbadin, A.; Zaniolo, G.; Ballarin, L. Genetic and cytological aspects of histocompatibility in ascidians. Boll. Zool. 1992, 59, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Gorman, A.; Mc Carty, J.; Finucane, D.; Reville, W.; Catter, T. Morphological assessment of apoptosis. In Techniques in Apoptosis—A User’s Guide; Catter, T.G., Martin, S.J., Eds.; Portland Press Ltd.: London, UK, 1996; pp. 1–20. [Google Scholar]
- Onofri, A. Routine statistical analyses of field experiments by using an Excel extension. In Proceedings of the 6th National Conference of Italian Biometric Society, Pisa, Italy, 20–22 June 2007; pp. 93–96. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | MIC (µg mL−1) | MBC (µg mL−1) |
|---|---|---|
| Bacillus clausii | 3.1 | >50.0 |
| Staphylococcus epidermidis | 3.1 | 12.5 |
| Escherichia coli | 50.0 | >50.0 |
| Proteus mirabilis | 6.3 | >50.0 |
| Saccharomyces cerevisiae | 6.3 | 12.5 |
| Temperature (°C) | pH | Ionic Strength [NaCl] (mM) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 60 (2 h) | 80 (2 h) | 100 (15 min) | 6.0 | 7.0 | 8.0 | 9.0 | 150 | 300 | 600 |
| 12.5 | 12.5 | 6.3 | 50.0 | 12.5 | 12.5 | 12.5 | 12.5 | 25.0 | 50.0 |
| Primer | Sequence |
|---|---|
| Botryllin forward | CTGGTTTCTCCAATAACG |
| Botryllin reverse1 | CAAGGTCATATTGGTGGCTA |
| Botryllin reverse2 | GTCGAAGCTGTGCAAGACAT |
| Botryllin-qRT PCR For | GGTCGGGGGATGGTATAAT |
| Botryllin-qRT PCR Rev | ACCATCGTATTCGTCCCG |
| BsEF1-alpha qPCR For | GCCGCCATACTCTGAAGC |
| BsEF1-alpha qPCR Rev | GTCCAACTGGCACTGTTCC |
| Botryllin ISH For | GGTCGGGGGATGGTATAAT |
| Botryllin ISH Rev | CAAGGTCATATTGGTGGCTA |
| Epitopes | Sequence Location | Antibody |
|---|---|---|
| GWYNKHKPTIHK | N-terminus of the mature peptide | Ab1 |
| VKGWGKDSDKEL | C-terminus of the mature peptide and beyond | Ab2 |
| KEHDRDEYDGAL | part of the propeptide beyond the C-terminus of the mature peptide | Ab3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Franchi, N.; Ballarin, L.; Cima, F. Botryllin, a Novel Antimicrobial Peptide from the Colonial Ascidian Botryllus schlosseri. Mar. Drugs 2023, 21, 74. https://doi.org/10.3390/md21020074
Franchi N, Ballarin L, Cima F. Botryllin, a Novel Antimicrobial Peptide from the Colonial Ascidian Botryllus schlosseri. Marine Drugs. 2023; 21(2):74. https://doi.org/10.3390/md21020074
Chicago/Turabian StyleFranchi, Nicola, Loriano Ballarin, and Francesca Cima. 2023. "Botryllin, a Novel Antimicrobial Peptide from the Colonial Ascidian Botryllus schlosseri" Marine Drugs 21, no. 2: 74. https://doi.org/10.3390/md21020074
APA StyleFranchi, N., Ballarin, L., & Cima, F. (2023). Botryllin, a Novel Antimicrobial Peptide from the Colonial Ascidian Botryllus schlosseri. Marine Drugs, 21(2), 74. https://doi.org/10.3390/md21020074