
The Genetic and Phenotypic Diversity of Bacillus spp. from the Mariculture System in China and Their Potential Function against Pathogenic Vibrio

Abstract
:1. Introduction
2. Results
2.1. Bacillus Isolates Information
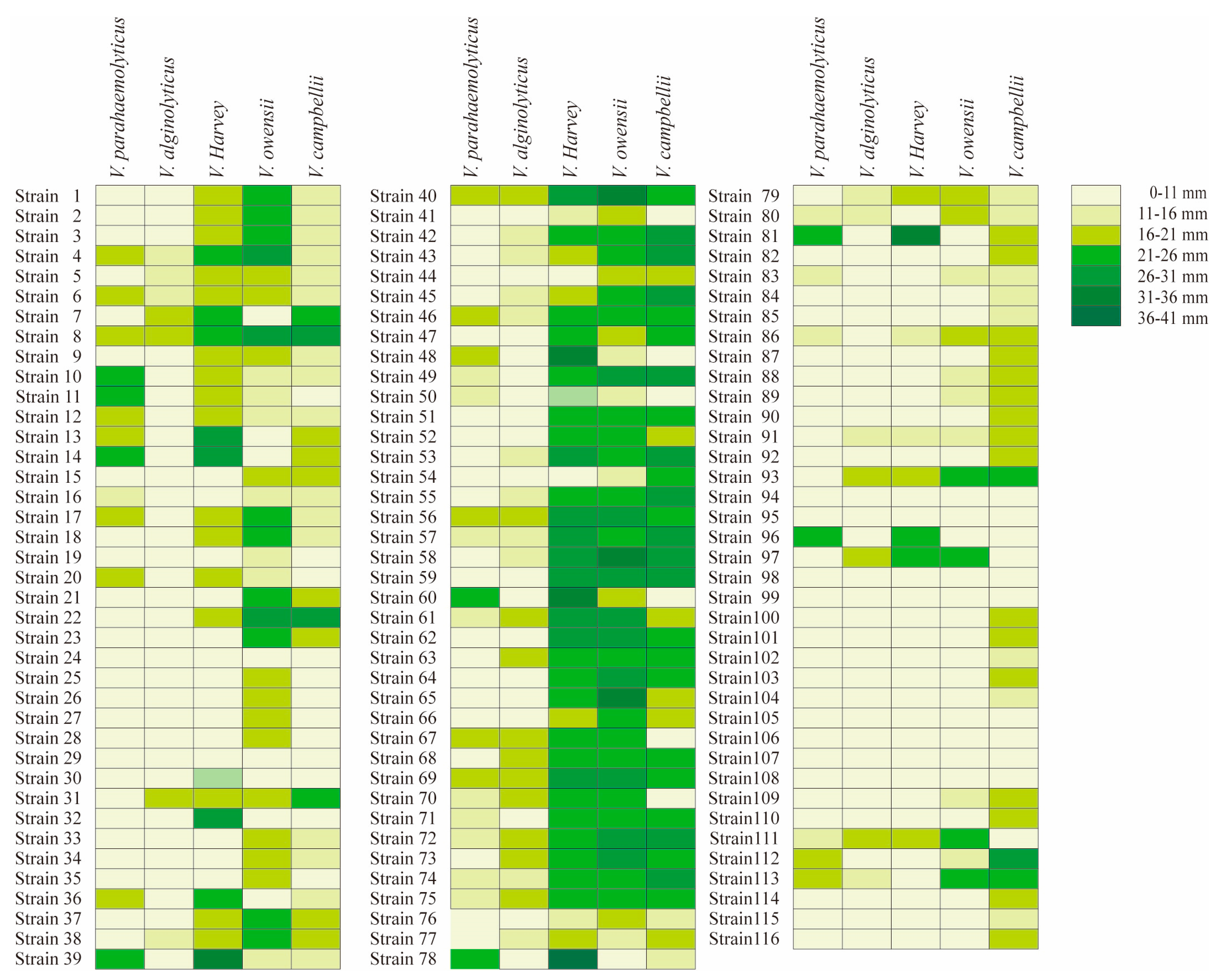
2.2. Antibacterial Activity Characterization of Bacillus Isolates against 5 Species of Vibrio
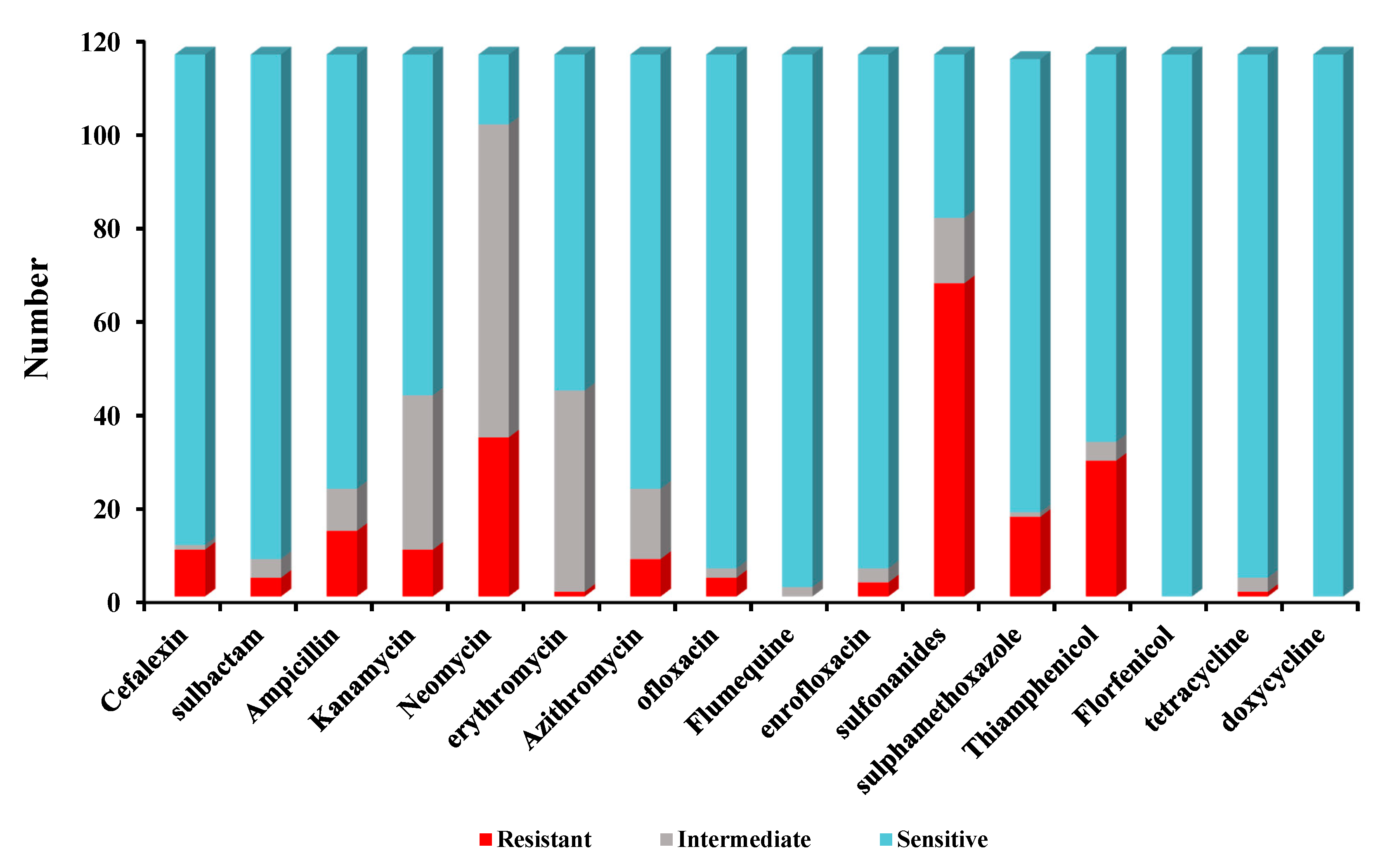
2.3. Antibiotic Susceptibility Characterization of Different Kinds of Bacillus Isolates
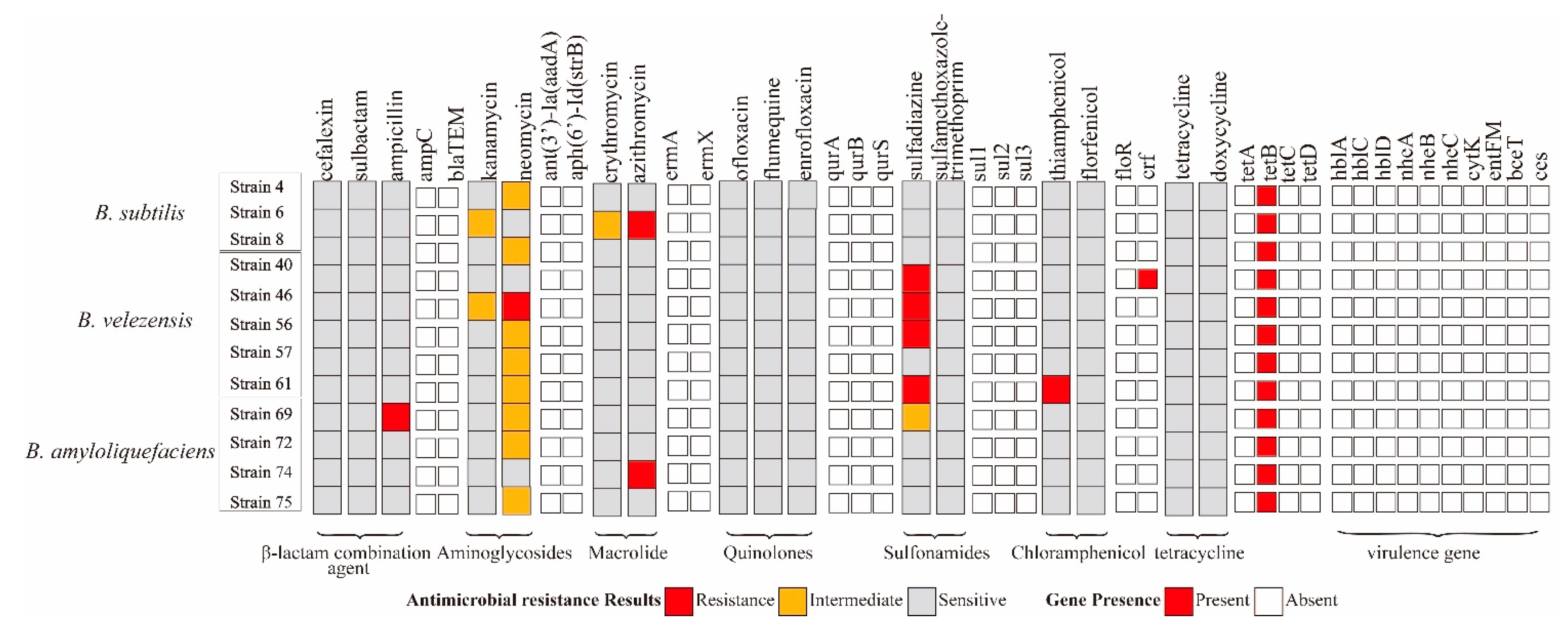
2.4. Distribution of Antimicrobial Resistance Genes among Bacillus Isolates
2.5. Distribution of Virulence Genes among Bacillus Isolates
2.6. Holistic Assessment of Potential Probiotics
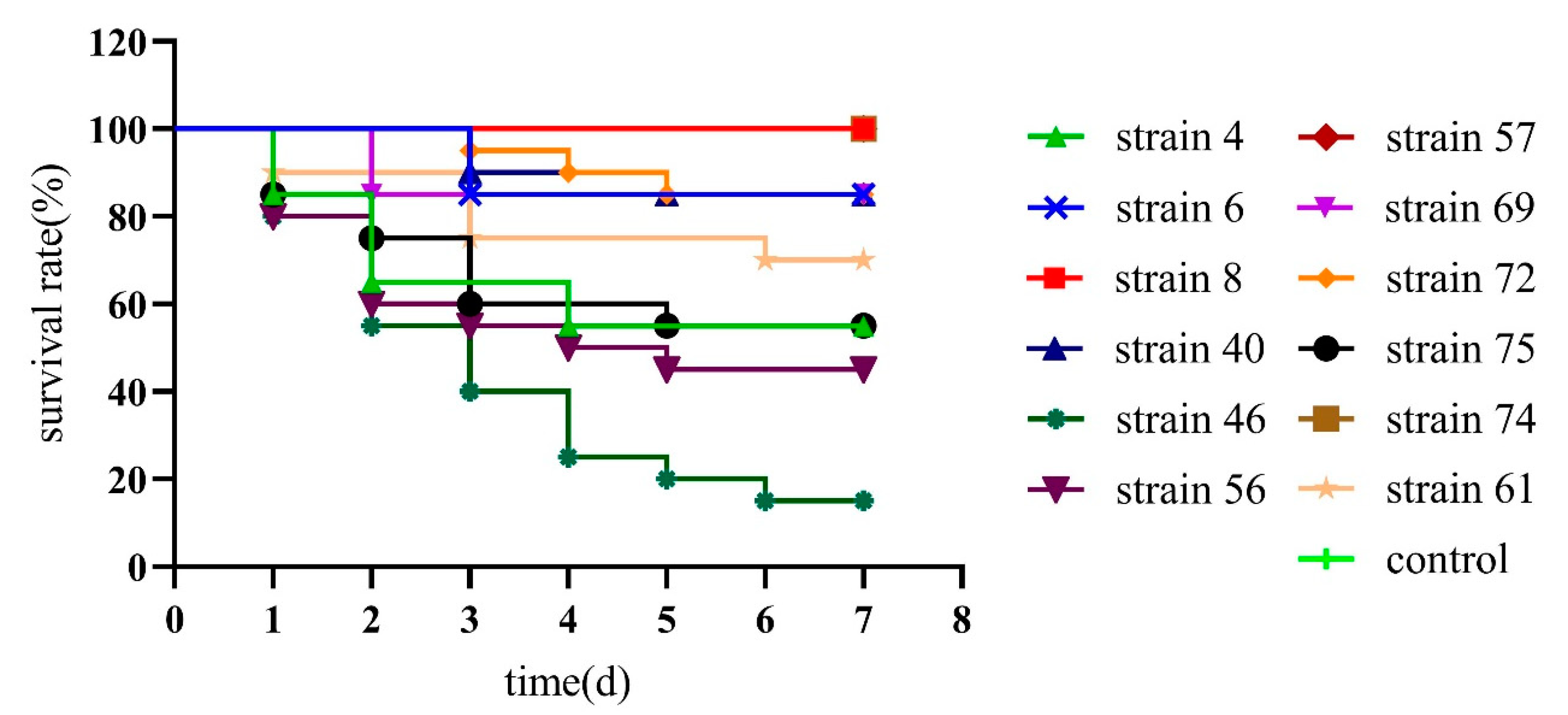
2.7. Safety Testing of Potential Probiotics Bacillus Isolates in L. vannamei
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Growth Condition
4.2. Antibacterial Ability of Bacillus Isolates against Vibrio spp.
4.3. Antibiotic Susceptibility Test
4.4. Antimicrobial Resistance Genes Detection among Bacillus Isolates
4.5. Molecular Detection of Potential Virulence Gene among Bacillus Isolates
4.6. Bio-Safety Testing in Litopenaeus vannamei
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Food and Agriculture Organization. The State of World Fisheries and Aquaculture; Food and Agriculture Organization of the United Nations: Rome, Italy, 2020. [Google Scholar]
- Abarike, E.D.; Jian, J.; Tang, J.; Cai, J.; Yu, H.; Lihua, C.; Jun, L. Influence of traditional Chinese medicine and Bacillus species (TCMBS) on growth, immune response and disease resistance in Nile tilapia, Oreochromis niloticus. Aquac. Res. 2018, 49, 2366–2375. [Google Scholar] [CrossRef]
- Flegel, T.W. Historic emergence, impact and current status of shrimp pathogens in Asia. J. Invertebr. Pathol. 2012, 110, 166–173. [Google Scholar] [CrossRef]
- Ghosh, A.K.; Panda, S.K.; Luyten, W. Anti-vibrio and immune-enhancing activity of medicinal plants in shrimp: A comprehensive review. Fish Shellfish Immunol. 2021, 117, 192–210. [Google Scholar] [CrossRef] [PubMed]
- Ishimaru, K.; Akagawa-Matsushita, M.; Muroga, K.; Microbiology, E. Vibrio penaeicida sp. nov., a pathogen of kuruma prawns (Penaeus japonicus). Int. J. Syst. Bacteriol. 1995, 45, 134–138. [Google Scholar] [CrossRef] [Green Version]
- Yi, Y.; Zhang, Z.; Zhao, F.; Liu, H.; Yu, L.; Zha, J.; Wang, G. Probiotic potential of Bacillus velezensis JW: Antimicrobial activity against fish pathogenic bacteria and immune enhancement effects on Carassius auratus. Fish Shellfish Immunol. 2018, 78, 322–330. [Google Scholar] [CrossRef]
- Gao, Y.; Han, G.; Qiang, L.; Zhang, L.; Tan, R.; Yu, Y. Hematological varieties, histological changes, and immune responses in the early stage of infection with Vibrio parahaemolyticus in Black rockfish Sebastes schlegelii. Aquacult Int. 2023, 31, 381–399. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Santos, R.A.; Monteiro, M.; Rangel, F.; Jerusik, R.; Saavedra, M.J.; Carvalho, A.P.; Oliva-Teles, A.; Serra, C.R. Bacillus spp. Inhibit Edwardsiella tarda Quorum-Sensing and Fish Infection. Mar. Drugs 2021, 19, 602. [Google Scholar] [CrossRef]
- Cha, J.-H.; Rahimnejad, S.; Yang, S.-Y.; Kim, K.-W.; Lee, K.-J. Evaluations of Bacillus spp. as dietary additives on growth performance, innate immunity and disease resistance of olive flounder (Paralichthys olivaceus) against Streptococcus iniae and as water additives. Aquaculture 2013, 402, 50–57. [Google Scholar] [CrossRef]
- Ramesh, D.; Souissi, S. Effects of potential probiotic Bacillus subtilis KADR1 and its subcellular components on immune responses and disease resistance in Labeo rohita. Aquac. Res. 2018, 49, 367–377. [Google Scholar] [CrossRef]
- Kavitha, M.; Raja, M.; Perumal, P. Evaluation of probiotic potential of Bacillus spp. isolated from the digestive tract of freshwater fish Labeo calbasu (Hamilton, 1822). Aquac. Rep. 2018, 11, 59–69. [Google Scholar] [CrossRef]
- Newaj-Fyzul, A.; Adesiyun, A.A.; Mutani, A.; Ramsubhag, A.; Brunt, J.; Austin, B. Bacillus subtilis AB1 controls Aeromonas infection in rainbow trout (Oncorhynchus mykiss, Walbaum). J. Appl. Microbiol. 2007, 103, 1699–1706. [Google Scholar] [CrossRef]
- Aly, S.M.; Abdel-Galil Ahmed, Y.; Abdel-Aziz Ghareeb, A.; Mohamed, M.F. Studies on Bacillus subtilis and Lactobacillus acidophilus, as potential probiotics, on the immune response and resistance of Tilapia nilotica (Oreochromis niloticus) to challenge infections. Fish Shellfish Immunol. 2008, 25, 128–136. [Google Scholar] [CrossRef]
- Tseng, D.Y.; Ho, P.L.; Huang, S.Y.; Cheng, S.C.; Shiu, Y.L.; Chiu, C.S.; Liu, C.H. Enhancement of immunity and disease resistance in the white shrimp, Litopenaeus vannamei, by the probiotic, Bacillus subtilis E20. Fish Shellfish Immunol. 2009, 26, 339–344. [Google Scholar] [CrossRef]
- Stenfors Arnesen, L.P.; Fagerlund, A.; Granum, P.E. From soil to gut: Bacillus cereus and its food poisoning toxins. FEMS Microbiol. Rev. 2008, 32, 579–606. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.G.; Lee, K.L.; Najiah, M.; Shariff, M.; Hassan, M.D. A new bacterial white spot syndrome (BWSS) in cultured tiger shrimp Penaeus monodon and its comparison with white spot syndrome (WSS) caused by virus. Dis. Aquat. Org. 2000, 41, 9–18. [Google Scholar] [CrossRef]
- Gueimonde, M.; Sánchez, B.; G de los Reyes-Gavilán, C.; Margolles, A. Antibiotic resistance in probiotic bacteria. Front. Microbiol. 2013, 4, 202. [Google Scholar] [CrossRef] [Green Version]
- Fu, S.; Yang, Q.; He, F.; Lan, R.; Hao, J.; Ni, P.; Liu, Y.; Li, R. National Safety Survey of Animal-use Commercial Probiotics and Their Spillover Effects From Farm to Humans: An Emerging Threat to Public Health. Clin. Infect. Dis. 2020, 70, 2386–2395. [Google Scholar] [CrossRef]
- Kumar, A.; Kumar, A.; Pratush, A. Molecular diversity and functional variability of environmental isolates of Bacillus species. Springerplus 2014, 3, 312. [Google Scholar] [CrossRef] [Green Version]
- Kanamoto, E.; Terashima, K.; Shiraki, Y.; Nishida, H. Diversity of Bacillus Isolates from the Sake Brewing Process at a Sake Brewery. Microorganisms 2021, 9, 1760. [Google Scholar] [CrossRef]
- Burkholder, P.R.; Pfister, R.M.; Leitz, F.H. Production of a pyrrole antibiotic by a marine bacterium. Appl. Microbiol. 1966, 14, 649–653. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Xia, Y.; Zhu, C.; Chu, W. Isolation of Marine Bacillus sp. with Antagonistic and Organic-Substances-Degrading Activities and Its Potential Application as a Fish Probiotic. Mar. Drugs 2018, 16, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.Y.; Liu, Y.; Miao, L.L.; Li, E.W.; Hou, T.T.; Liu, Z.P. Mechanism of anti-Vibrio activity of marine probiotic strain Bacillus pumilus H2, and characterization of the active substance. AMB Express 2017, 7, 23. [Google Scholar] [CrossRef] [Green Version]
- Uzun, E.; Ogut, H. The isolation frequency of bacterial pathogens from sea bass (Dicentrarchus labrax) in the Southeastern Black Sea. Aquaculture 2015, 437, 30–37. [Google Scholar] [CrossRef]
- Zhou, J.; Fang, W.; Yang, X.; Zhou, S.; Hu, L.; Li, X.; Qi, X.; Su, H.; Xie, L. A nonluminescent and highly virulent Vibrio harveyi strain is associated with “bacterial white tail disease” of Litopenaeus vannamei shrimp. PLoS ONE 2012, 7, e29961. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Wang, H.; Xie, G.; Zou, P.; Guo, C.; Liang, Y.; Huang, J. An isolate of Vibrio campbellii carrying the pir(VP) gene causes acute hepatopancreatic necrosis disease. Emerg. Microbes Infect. 2017, 6, e2. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.Y.; Xiao, J.Z.; Zhang, M.M.; Zhu, W.Y.; Xia, X.M.; Dai, X.L.; Pan, Y.J.; Yan, S.L.; Wang, Y.J. A Vibrio owensii strain as the causative agent of AHPND in cultured shrimp, Litopenaeus vannamei. J. Invertebr. Pathol. 2018, 153, 156–164. [Google Scholar] [CrossRef]
- Azimonti, G.; Bampidis, V.; Bastos MD, L.; Christensen, H.; Dusemund, B.; Fašmon Durjava, M.; Kouba, M.; López-Alonso, M.; López Puente, S.; Marcon, F.; et al. EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP). EFSA J. Eur. Food Saf. Auth. 2018, 20, e07268. [Google Scholar]
- Cui, Y.; Martlbauer, E.; Dietrich, R.; Luo, H.; Ding, S.; Zhu, K. Multifaceted toxin profile, an approach toward a better understanding of probiotic Bacillus cereus. Crit. Rev. Toxicol. 2019, 49, 342–356. [Google Scholar] [CrossRef]
- Cui, Y.; Wang, S.; Ding, S.; Shen, J.; Zhu, K. Toxins and mobile antimicrobial resistance genes in Bacillus probiotics constitute a potential risk for One Health. J. Hazard. Mater. 2020, 382, 121266. [Google Scholar] [CrossRef]
- Zhu, K.; Holzel, C.S.; Cui, Y.; Mayer, R.; Wang, Y.; Dietrich, R.; Didier, A.; Bassitta, R.; Martlbauer, E.; Ding, S. Probiotic Bacillus cereus Strains, a Potential Risk for Public Health in China. Front. Microbiol. 2016, 7, 718. [Google Scholar] [CrossRef]
- Nandi, A.; Dan, S.K.; Banerjee, G.; Ghosh, P.; Ghosh, K.; Ringo, E.; Ray, A.K. Probiotic Potential of Autochthonous Bacteria Isolated from the Gastrointestinal Tract of Four Freshwater Teleosts. Probiotics Antimicro. 2017, 9, 12–21. [Google Scholar] [CrossRef]
- Deng, F.; Chen, Y.; Sun, T.; Wu, Y.; Su, Y.; Liu, C.; Zhou, J.; Deng, Y.; Wen, J. Antimicrobial resistance, virulence characteristics and genotypes of Bacillus spp. from probiotic products of diverse origins. Food Res. Int. 2021, 139, 109949. [Google Scholar] [CrossRef]
- Hernando-Amado, S.; Coque, T.M.; Baquero, F.; Martinez, J.L. Antibiotic Resistance: Moving From Individual Health Norms to Social Norms in One Health and Global Health. Front. Microbiol. 2020, 11, 1914. [Google Scholar] [CrossRef]
- Patel, A.K.; Ahire, J.J.; Pawar, S.P.; Chaudhari, B.L.; Chincholkar, S.B. Comparative accounts of probiotic characteristics of Bacillus spp. isolated from food wastes. Food Res. Int. 2009, 42, 505–510. [Google Scholar] [CrossRef]
- Yaylaci, E.U. Isolation and characterization of Bacillus spp. from aquaculture cage water and its inhibitory effect against selected Vibrio spp. Arch. Microbiol. 2021, 204, 26. [Google Scholar] [CrossRef]
- Sorokulova, I.B.; Pinchuk, I.V.; Denayrolles, M.; Osipova, I.G.; Huang, J.M.; Cutting, S.M.; Urdaci, M.C. The safety of two Bacillus probiotic strains for human use. Dig. Dis. Sci. 2008, 53, 954–963. [Google Scholar] [CrossRef]
- Pall, E.; Niculae, M.; Brudasca, G.F.; Ravilov, R.K.; Sandru, C.D.; Cerbu, C.; Olah, D.; Zablau, S.; Potarniche, A.V.; Spinu, M.; et al. Assessment and Antibiotic Resistance Profiling in Vibrio Species Isolated from Wild Birds Captured in Danube Delta Biosphere Reserve, Romania. Antibiotics 2021, 10, 333. [Google Scholar] [CrossRef]
- Mohamad, N.; Amal, M.N.A.; Saad, M.Z.; Yasin, I.S.M.; Zulkiply, N.A.; Mustafa, M.; Nasruddin, N.S. Virulence-associated genes and antibiotic resistance patterns of Vibrio spp. isolated from cultured marine fishes in Malaysia. BMC Vet. Res. 2019, 15, 176. [Google Scholar] [CrossRef]
- Qiao, M.; Ying, G.G.; Singer, A.C.; Zhu, Y.G. Review of antibiotic resistance in China and its environment. Environ. Int. 2018, 110, 160–172. [Google Scholar] [CrossRef] [Green Version]
- Dang, H.; Zhang, X.; Song, L.; Chang, Y.; Yang, G. Molecular characterizations of oxytetracycline resistant bacteria and their resistance genes from mariculture waters of China. Mar. Pollut. Bull. 2006, 52, 1494–1503. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Yu, Y.; Liao, M.; Wang, Y.; Yang, G.; Zhang, Z.; Li, B.; Rong, X.; Wang, C. Physiology, metabolism, antibiotic resistance, and genetic diversity of Harveyi clade bacteria isolated from coastal mariculture system in China in the last two decades. Front. Mar. Sci. 2022, 9. [Google Scholar] [CrossRef]
- Van, C.N.; Zhang, L.J.; Thanh, T.V.T.; Son, H.P.H.; Ngoc, T.T.; Huang, Q.; Zhou, R. Association between the Phenotypes and Genotypes of Antimicrobial Resistance in Haemophilus parasuis Isolates from Swine in Quang Binh and Thua Thien Hue Provinces, Vietnam. Engineering 2020, 6, 40–48. [Google Scholar] [CrossRef]
- Monod, M.; Denoya, C.; Dubnau, D. Sequence and Properties of pIM13, a Macrolide-Lincosamide-Streptogramin B Resistance Plasmid from Bacillus subtilis. J. Bacteriol. 1986, 167, 138–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phelan, R.W.; Clarke, C.; Morrissey, J.P.; Dobson, A.D.; O’Gara, F.; Barbosa, T.M. Tetracycline resistance-encoding plasmid from Bacillus sp. strain #24, isolated from the marine sponge Haliclona simulans. Appl. Environ. Microbiol. 2011, 77, 327–329. [Google Scholar] [CrossRef] [Green Version]
- Roberts, A.P.; Pratten, J.; Wilson, M.; Mullany, P. Transfer of a conjugative transposon, Tn5397 in a model oral biofilm. FEMS Microbiol. Lett. 1999, 177, 63–66. [Google Scholar] [CrossRef] [PubMed]
- Neela, F.A.; Nonaka, L.; Rahman, M.H.; Suzuki, S. Transfer of the chromosomally encoded tetracycline resistance gene tet(M) from marine bacteria to Escherichia coli and Enterococcus faecalis. World J. Microbiol. Biotechnol. 2009, 25, 1095–1101. [Google Scholar] [CrossRef]
- Alvarez-Contreras, A.K.; Quinones-Ramirez, E.I.; Vazquez-Salinas, C. Prevalence, detection of virulence genes and antimicrobial susceptibility of pathogen Vibrio species isolated from different types of seafood samples at “La Nueva Viga” market in Mexico City. Antonie Leeuwenhoek 2021, 114, 1417–1429. [Google Scholar] [CrossRef]
- Bush, K.; Bradford, P.A. beta-Lactams and beta-Lactamase Inhibitors: An Overview. Cold Spring Harb. Perspect. Med. 2016, 6, a025247. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; CLSI: Wayne, PA, USA, 2018; Volume M100. [Google Scholar]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Krumperman, P.H. Multiple antibiotic resistance indexing of Escherichia coli to identify high-risk sources of fecal contamination of foods. Appl. Environ. Microbiol. 1983, 46, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Chon, J.W.; Kim, J.H.; Lee, S.J.; Hyeon, J.Y.; Song, K.Y.; Park, C.; Seo, K.H. Prevalence, phenotypic traits and molecular characterization of emetic toxin-producing Bacillus cereus strains isolated from human stools in Korea. J. Appl. Microbiol. 2012, 112, 1042–1049. [Google Scholar] [CrossRef] [PubMed]
- Heini, N.; Stephan, R.; Ehling-Schulz, M.; Johler, S. Characterization of Bacillus cereus group isolates from powdered food products. Int. J. Food Microbiol. 2018, 283, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Ehling-Schulz, M.; Vukov, N.; Schulz, A.; Shaheen, R.; Andersson, M.; Martlbauer, E.; Scherer, S. Identification and partial characterization of the nonribosomal peptide synthetase gene responsible for cereulide production in emetic Bacillus cereus. Appl. Environ. Microbiol. 2005, 71, 105–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawaz, M.; Sung, K.; Khan, S.A.; Khan, A.A.; Steele, R. Biochemical and molecular characterization of tetracycline-resistant Aeromonas veronii isolates from catfish. Appl. Environ. Microbiol. 2006, 72, 6461–6466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furlaneto-Maia, L.; Rocha, K.R.; Siqueira, V.L.; Furlaneto, M.C. Comparison between automated system and PCR-based method for identification and antimicrobial susceptibility profile of clinical Enterococcus spp. Rev. Inst. Med. Trop. São Paulo 2014, 56, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Kehrenberg, C.; Schwarz, S. Distribution of florfenicol resistance genes fexA and cfr among chloramphenicol-resistant Staphylococcus isolates. Antimicrob. Agents Chemother. 2006, 50, 1156–1163. [Google Scholar] [CrossRef] [Green Version]
- Sandvang, D.; Aarestrup, F.M.; Jensen, L.B. Characterisation of integrons and antibiotic resistance genes in Danish multiresistant Salmonella enterica Typhimurium DT104. Microbiol. Lett. 1997, 157, 177–181. [Google Scholar] [CrossRef]
- Li, Y.; Chen, L.; Wu, X.; Huo, S. Molecular characterization of multidrug-resistant avian pathogenic Escherichia coli isolated from septicemic broilers. Poult. Sci. 2015, 94, 601–611. [Google Scholar] [CrossRef]
- Bochniarz, M.; Adaszek, L.; Dziegiel, B.; Nowaczek, A.; Wawron, W.; Dabrowski, R.; Szczubial, M.; Winiarczyk, S. Factors responsible for subclinical mastitis in cows caused by Staphylococcus chromogenes and its susceptibility to antibiotics based on bap, fnbA, eno, mecA, tetK, and ermA genes. J. Dairy Sci. 2016, 99, 9514–9520. [Google Scholar] [CrossRef] [Green Version]
- Letchumanan, V.; Yin, W.F.; Lee, L.H.; Chan, K.G. Prevalence and antimicrobial susceptibility of Vibrio parahaemolyticus isolated from retail shrimps in Malaysia. Front. Microbiol. 2015, 6, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gxalo, O.; Digban, T.O.; Igere, B.E.; Olapade, O.A.; Okoh, A.I.; Nwodo, U.U. Virulence and Antibiotic Resistance Characteristics of Vibrio Isolates From Rustic Environmental Freshwaters. Front. Cell. Infect. Microbiol. 2021, 11, 732001. [Google Scholar] [CrossRef] [PubMed]
- Mohkam, M.; Nezafat, N.; Berenjian, A.; Mobasher, M.A.; Ghasemi, Y. Identification of Bacillus Probiotics Isolated from Soil Rhizosphere Using 16S rRNA, recA, rpoB Gene Sequencing and RAPD-PCR. Probiotics Antimicrob. Proteins 2016, 8, 8–18. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | Numbers | Isolates | Numbers |
|---|---|---|---|
| B. subtilis | 37 | B. tequilensis | 1 |
| B. velezensis | 28 | B.inaquosorum | 1 |
| B. amyloliquefaciens | 10 | B. stratosphericus | 1 |
| B.stercoris | 6 | B. koreensis | 1 |
| B. cereus | 6 | B. lehensis | 1 |
| B. thuringiensis | 3 | B. gibsonii | 1 |
| B. megaterium | 3 | B. methylotrophicus | 1 |
| B. flexus | 3 | B. licheniformis | 1 |
| B. nealsonii | 2 | B. pumilus | 1 |
| B. altitudinis | 2 | B. haikouensis | 1 |
| B. aryabhattai | 2 | B. circulans | 1 |
| B. atrophaeus | 2 | B. marisflavi | 1 |
| Pathogenic Bacterial | Bacillus Isolates Numbers |
|---|---|
| V. parahaemolyticus | 38 |
| V. alginolyticus | 35 |
| V. harvey | 70 |
| V. owensii | 81 |
| V. campbellii | 86 |
| Multiple Antibiotic Resistance | % of Occurrence |
|---|---|
| MAR (3) | 8.62 |
| MAR (4) | 4.31 |
| MAR (5) | 6.03 |
| MAR (6) | 1.72 |
| MAR (7) | 1.72 |
| Strains No. | Inhibition Zone (mm) * | ||||
|---|---|---|---|---|---|
| V. parahaemolyticus | V. alginolyticus | V. harvey | V. owensii | V. campbellii | |
| strain 4 | 16.0 ± 0.73 | 15.7 ± 0.41 | 21.3 ± 0.49 | 28.3 ± 0.98 | 14.5 ± 0.49 |
| strain 6 | 17.0 ± 0.90 | 14.0 ± 0.73 | 18.1 ± 0.57 | 19.3 ± 0.49 | 13.4 ± 0.65 |
| strain 8 | 18.5 ± 0.41 | 18.4 ± 0.49 | 22.6 ± 0.16 | 27.8 ± 0.65 | 28.7 ± 0.89 |
| strain 40 | 20.1 ± 0.73 | 18.5 ± 0.57 | 27.6 ± 0.24 | 33.4 ± 0.57 | 25.0 ± 0.57 |
| strain 46 | 16.2 ± 0.24 | 14.9 ± 0.65 | 25.1 ± 0.49 | 24.7 ± 0.49 | 23.6 ± 0.41 |
| strain 56 | 16.2 ± 0.98 | 17.4 ± 1.06 | 28.7 ± 0.33 | 30.9 ± 1.06 | 25.5 ± 0.65 |
| strain 57 | 15.3 ± 0.57 | 15.3 ± 0.98 | 26.9 ± 0.82 | 24.7 ± 1.14 | 27.7 ± 0.98 |
| strain 61 | 15.7 ± 0.65 | 16.4 ± 0.57 | 26.2 ± 0.98 | 30.7 ± 0.73 | 20.8 ± 0.90 |
| strain 69 | 19.2 ± 0.90 | 19.7 ± 0.65 | 26.8 ± 0.41 | 27.9 ± 0.82 | 23.1 ± 0.73 |
| strain 72 | 14.2 ± 0.73 | 17.7 ± 0.33 | 24.2 ± 0.65 | 27.5 ± 0.73 | 27.1 ± 1.22 |
| strain 74 | 14.3 ± 0.57 | 14.7 ± 0.98 | 22.7 ± 0.98 | 25.2 ± 0.98 | 26.0 ± 0.57 |
| strain 75 | 13.0 ± 0.98 | 16.6 ± 0.82 | 22.5 ± 0.49 | 24.3 ± 0.33 | 25.5 ± 0.73 |
| Primer | Sequence (5′-3′) | Gene | Amplicon Size (bp) | Reference |
|---|---|---|---|---|
| hblA-F | AAGCAATGGAATACAATGGG | hblA | 1154 | [54] |
| hblA-R | AGAATCTAAATCATGCCACTGC | |||
| hblC-F | GATACTCAATGTGGGAACTGC | hblC | 740 | |
| hblC-R | TTGAGACTGCTCGTCTAGTTG | |||
| hblD-F | ACCGGTAACACTATTCATGC | hblD | 829 | |
| hblD-R | GAGTCCATATGCTTAGATGC | |||
| nheA-F | GTTAGGATCACAATCACCGC | nheA | 755 | |
| nheA-R | ACGAATGTAATTTGAGTCGC | |||
| nheB-F | TTTAGTGGATCTGTACGC | nheB | 743 | |
| nheB-R | TTAATGTTCGTTAATCCTGC | |||
| nheC-F | TGGATTCCAAGATGTAACG | nheC | 683 | |
| nheC-R | ATTACGACTTCTGCTTGTGC | |||
| cytK-F | CGACGTCACAAGTTGTAACA | cytK | 565 | [55] |
| cytK-R | CGTGTGTAAATACCCAGTT | |||
| entFM-F | GTTCGTTCAGGTGCTGGTAC | entFM | 486 | |
| entFM-R | AGCTGGGCCTGTACGTACTT | |||
| bceT-F | TTACATTACCAGGACGTGCTT | bceT | 428 | [56] |
| bceT-R | TGTTTGTGATTGTAATTCAGG | |||
| ces-F | GGTGACACATTATCATATAAGGTG | ces | 1271 | |
| ces-R | GTAAGCGAACCTGTCTGTAACAACA | |||
| tetA-F | GCTACATCCTGCTTGCCTTC | tetA | 212 | [57] |
| tetA-R | GCATAGATCGCCGTGAAGAG | |||
| tetB-F | TACGTGAATTTATTGCTTCGG | tetB | 206 | |
| tetB-R | ATACAGCATCCAAAGCGCAC | |||
| tetD-F | TGTGCTGTGGATGTTGTATCTC | tetD | 844 | |
| tetD-R | CAGTGCCGTGCCAATCAG | |||
| qnrA-F | TTCAGCAAGAGGATTTCTCA | qnrA | 500 | [58] |
| qnrA-R | GGCAGCACTATTACTCCCAA | |||
| qnrB-F | CCTGAGCGGCACTGAATTTAT | qnrB | 617 | |
| qnrB-R | GTTTGCTGCTCGCCAGTCGA | |||
| qnrS-F | ACATAAAGACTTAAGTGATC | qnrS | 619 | |
| qnrS-R | CAATTAGTCAGGATAAAC | |||
| floR-F | CTGCTGATGGCTCCTTTC | flor | 650 | [59] |
| floR-R | GCCGTGGCGTAACAGAT | |||
| cfr-F | TGAAGTATAAAGCAGGTTGGGAGTCA | cfr | 746 | |
| cfr-R | ACCATATAATTGACCACAAGCAGC | |||
| Sul1-F | GTGACGGTGTTCGGCATTCT | sul1 | 800 | [60] |
| Sul1-R | TCCGAGAAGGTGATTGCGCT | |||
| Sul2-F | CATCATTTTCGGCATCGTC | sul2 | 793 | |
| Sul2-R | TCTTGCGGTTTCTTTCAGC | |||
| Sul3-F | GCAACAGTTGGTGCTAAACGAGA | sul3 | 578 | |
| Sul3-R | AGCAGATGTGATTGATTTGGGAG | |||
| ant(3′)-Ia(aadA)-F | ATCTGGCTATCTTGCTGACA | ant(3′)-Ia(aadA) | 388 | [61] |
| ant(3′)-Ia(aadA)-R | TTGGTGATCTCGCCTTTC | |||
| aph(6′)-Id(strB)-F | ATGTTCATGCCGCCTGTTTTT | aph(6′)-Id(strB) | 837 | |
| aph(6′)-Id(strB)-R | CTAGTATGACGTCTGTCGC | |||
| ermA-F | GTTCAAGAACAATCAATACAGAG | ermA | 421 | [62] |
| ermA-R | GGATCAGGAAAAGGACATTTTAC | |||
| ermX-F | GTTGCGCTCTAACCGCTAAGGC | ermX | 566 | |
| ermX-R | CCATGGGGACCACTGAGCCGTC | |||
| blaTEM | AAAGATGCTGAAGATCA | blaTEM | 425 | [63] |
| blaTEM | TTTGGTATGGCTTCATTC | |||
| ampC | GCGAAAGCCAGCTGTCGGGC | ampC | 550 | [64] |
| ampC | CCYTTTTATGTACCCAYGA | |||
| blaZ | ACTTCAACACCTGCTTTC | blaZ | 490 | |
| blaZ | TGACCACTTTTATCAGCAACC | |||
| 27F | AGAGTTTGATCCTGGCTCAG | 16S rDNA | 1542 | [65] |
| 1492R | TACGGCTACCTTGTTACGACTT | |||
| recA-F | GATCGTCAAGCAGCCTTAGAT | recA | 540 | |
| recA-R | TTACCGACCATAACGCCGAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Y.; Zhang, Y.; Wang, Y.; Liao, M.; Li, B.; Rong, X.; Wang, C.; Ge, J.; Wang, J.; Zhang, Z. The Genetic and Phenotypic Diversity of Bacillus spp. from the Mariculture System in China and Their Potential Function against Pathogenic Vibrio. Mar. Drugs 2023, 21, 228. https://doi.org/10.3390/md21040228
Yu Y, Zhang Y, Wang Y, Liao M, Li B, Rong X, Wang C, Ge J, Wang J, Zhang Z. The Genetic and Phenotypic Diversity of Bacillus spp. from the Mariculture System in China and Their Potential Function against Pathogenic Vibrio. Marine Drugs. 2023; 21(4):228. https://doi.org/10.3390/md21040228
Chicago/Turabian StyleYu, Yongxiang, Yang Zhang, Yingeng Wang, Meijie Liao, Bin Li, Xiaojun Rong, Chunyuan Wang, Jianlong Ge, Jinjin Wang, and Zheng Zhang. 2023. "The Genetic and Phenotypic Diversity of Bacillus spp. from the Mariculture System in China and Their Potential Function against Pathogenic Vibrio" Marine Drugs 21, no. 4: 228. https://doi.org/10.3390/md21040228
APA StyleYu, Y., Zhang, Y., Wang, Y., Liao, M., Li, B., Rong, X., Wang, C., Ge, J., Wang, J., & Zhang, Z. (2023). The Genetic and Phenotypic Diversity of Bacillus spp. from the Mariculture System in China and Their Potential Function against Pathogenic Vibrio. Marine Drugs, 21(4), 228. https://doi.org/10.3390/md21040228