
Alexandrium spp.: From Toxicity to Potential Biotechnological Benefits

 ,
,  ,
,  ,
,  and
and 
Abstract
:
1. Introduction
2. Alexandrium Secondary Metabolites and Possible Biotechnological Applications
2.1. Saxitoxin
2.1.1. Chemistry
2.1.2. Toxicity
2.1.3. Possible Applications
2.2. Spiroimines (Gymnodimines and Spirolides)
2.2.1. Chemistry
2.2.2. Toxicity
2.2.3. Possible Applications
2.3. Goniodomins
2.3.1. Chemistry
2.3.2. Toxicity
2.3.3. Possible Applications
2.4. Tetrodotoxin
2.4.1. Chemistry
2.4.2. Toxicity
2.4.3. Possible Applications
2.5. Other Molecules
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Gómez, F. A List of Dinoflagellates in the World’s Oceans. Acta Bot. Croat. 2005, 64, 129–212. [Google Scholar]
- Gómez, F. A Checklist and Classification of Living Dinoflagellates (Dinoflagellata, Alveolata). Cicimar Oceánides 2012, 27, 65–140. [Google Scholar] [CrossRef]
- Gómez, F.; Artigas, L.F. Redefinition of the Dinoflagellate Genus Alexandrium Based on Centrodinium: Reinstatement of Gessnerium and Protogonyaulax, and Episemicolon Gen. Nov. (Gonyaulacales, Dinophyceae). J. Mar. Biol. 2019, 2019, 1284104. [Google Scholar] [CrossRef]
- Taylor, F.J.R.; Hoppenrath, M.; Saldarriaga, J.F. Dinoflagellate Diversity and Distribution. Biodivers. Conserv. 2008, 17, 407–418. [Google Scholar] [CrossRef]
- Cohen, N.R.; McIlvin, M.R.; Moran, D.M.; Held, N.A.; Saunders, J.K.; Hawco, N.J.; Brosnahan, M.; DiTullio, G.R.; Lamborg, C.; McCrow, J.P.; et al. Dinoflagellates Alter Their Carbon and Nutrient Metabolic Strategies across Environmental Gradients in the Central Pacific Ocean. Nat. Microbiol. 2021, 6, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Smayda, T.J. Adaptive Ecology, Growth Strategies and the Global Bloom Expansion of Dinoflagellates. J. Oceanogr. 2002, 58, 281–294. [Google Scholar] [CrossRef]
- Schnepf, E.; Elbrächter, M. Nutritional Strategies in Dinoflagellates. Eur. J. Protistol. 1992, 28, 3–24. [Google Scholar] [CrossRef]
- Camacho, F.G.; Rodríguez, J.G.; Mirón, A.S.; García, M.C.C.; Belarbi, E.H.; Chisti, Y.; Grima, E.M. Biotechnological Significance of Toxic Marine Dinoflagellates. Biotechnol. Adv. 2007, 25, 176–194. [Google Scholar] [CrossRef]
- Matsuyama, Y. Impacts of the Harmful Dinoflagellate Heterocapsa circularisquama Bloom on Shellfish Aquaculture in Japan and Some Experimental Studies on Invertebrates. Harmful Algae 2012, 14, 144–155. [Google Scholar] [CrossRef]
- Sakamoto, S.; Lim, W.A.; Lu, D.; Dai, X.; Orlova, T.; Iwataki, M. Harmful Algal Blooms and Associated Fisheries Damage in East Asia: Current Status and Trends in China, Japan, Korea and Russia. Harmful Algae 2021, 102, 101787. [Google Scholar] [CrossRef]
- Bechard, A. Red Tide at Morning, Tourists Take Warning? County-Level Economic Effects of HABS on Tourism Dependent Sectors. Harmful Algae 2019, 85, 101689. [Google Scholar] [CrossRef] [PubMed]
- Berge, E.; Whiteley, W.; Audebert, H.; De Marchis, G.; Fonseca, A.C.; Padiglioni, C.; Pérez De La Ossa, N.; Strbian, D.; Tsivgoulis, G.; Turc, G. European Stroke Organisation (ESO) Guidelines on Intravenous Thrombolysis for Acute Ischaemic Stroke. Eur. Stroke J. 2021, 6, I–LXII. [Google Scholar] [CrossRef] [PubMed]
- Bricelj, V.M.; Haubois, A.-G.; Sengco, M.R.; Pierce, R.H.; Culter, J.K.; Anderson, D.M. Trophic Transfer of Brevetoxins to the Benthic Macrofaunal Community during a Bloom of the Harmful Dinoflagellate Karenia Brevis in Sarasota Bay, Florida. Harmful Algae 2012, 16, 27–34. [Google Scholar] [CrossRef]
- Alves, T.; Mafra, L. Diel Variations in Cell Abundance and Trophic Transfer of Diarrheic Toxins during a Massive Dinophysis Bloom in Southern Brazil. Toxins 2018, 10, 232. [Google Scholar] [CrossRef] [PubMed]
- Gibble, C.M.; Hoover, B.A. Interactions between Seabirds and Harmful Algal Blooms. In Harmful Algal Blooms; Shumway, S.E., Burkholder, J.M., Morton, S.L., Eds.; Wiley: Hoboken, NJ, USA, 2018; pp. 223–242. ISBN 978-1-118-99467-2. [Google Scholar]
- Reis Costa, P. Impact and Effects of Paralytic Shellfish Poisoning Toxins Derived from Harmful Algal Blooms to Marine Fish. Fish Fish. 2016, 17, 226–248. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. 30 March 2023. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. Available online: https://www.algaebase.org (accessed on 11 August 2023).
- Anderson, D.M.; Alpermann, T.J.; Cembella, A.D.; Collos, Y.; Masseret, E.; Montresor, M. The Globally Distributed Genus Alexandrium: Multifaceted Roles in Marine Ecosystems and Impacts on Human Health. Harmful Algae 2012, 14, 10–35. [Google Scholar] [CrossRef]
- Van Dolah, F.M. Marine Algal Toxins: Origins, Health Effects, and Their Increased Occurrence. Environ. Health Perspect. 2000, 108, 133–141. [Google Scholar] [CrossRef]
- Anderson, D.M.; Glibert, P.M.; Burkholder, J.M. Harmful Algal Blooms and Eutrophication: Nutrient Sources, Composition, and Consequences. Estuaries 2002, 25, 704–726. [Google Scholar] [CrossRef]
- John, U.; Litaker, R.W.; Montresor, M.; Murray, S.; Brosnahan, M.L.; Anderson, D.M. Formal Revision of the Alexandrium tamarense Species Complex (Dinophyceae) Taxonomy: The Introduction of Five Species with Emphasis on Molecular-Based (rDNA) Classification. Protist 2014, 165, 779–804. [Google Scholar] [CrossRef]
- Long, M.; Krock, B.; Castrec, J.; Tillmann, U. Unknown Extracellular and Bioactive Metabolites of the Genus Alexandrium: A Review of Overlooked Toxins. Toxins 2021, 13, 905. [Google Scholar] [CrossRef]
- Lewis, A.M.; Coates, L.N.; Turner, A.D.; Percy, L.; Lewis, J. A Review of the Global Distribution of Alexandrium minutum (Dinophyceae) and Comments on Ecology and Associated Paralytic Shellfish Toxin Profiles, with a Focus on Northern Europe. J. Phycol. 2018, 54, 581–598. [Google Scholar] [CrossRef] [PubMed]
- Casabianca, S.; Penna, A.; Pecchioli, E.; Jordi, A.; Basterretxea, G.; Vernesi, C. Population Genetic Structure and Connectivity of the Harmful Dinoflagellate Alexandrium minutum in the Mediterranean Sea. Proc. R. Soc. B 2012, 279, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Wang, F.; Zhang, W. Omics Analysis for Dinoflagellates Biology Research. Microorganisms 2019, 7, 288. [Google Scholar] [CrossRef] [PubMed]
- Vingiani, G.M.; Štālberga, D.; De Luca, P.; Ianora, A.; De Luca, D.; Lauritano, C. De Novo Transcriptome of the Non-Saxitoxin Producing Alexandrium tamutum Reveals New Insights on Harmful Dinoflagellates. Mar. Drugs 2020, 18, 386. [Google Scholar] [CrossRef]
- Meng, F.-Q.; Song, J.-T.; Zhou, J.; Cai, Z.-H. Transcriptomic Profile and Sexual Reproduction-Relevant Genes of Alexandrium minutum in Response to Nutritional Deficiency. Front. Microbiol. 2019, 10, 2629. [Google Scholar] [CrossRef]
- Jaeckisch, N.; Yang, I.; Wohlrab, S.; Glöckner, G.; Kroymann, J.; Vogel, H.; Cembella, A.; John, U. Comparative Genomic and Transcriptomic Characterization of the Toxigenic Marine Dinoflagellate Alexandrium ostenfeldii. PLoS ONE 2011, 6, e28012. [Google Scholar] [CrossRef]
- Brown, E.R.; Moore, S.G.; Gaul, D.A.; Kubanek, J. Differentiating Toxic and Nontoxic Congeneric Harmful Algae Using the Non-Polar Metabolome. Harmful Algae 2021, 110, 102129. [Google Scholar] [CrossRef]
- Galluzzi, L.; Penna, A.; Bertozzini, E.; Vila, M.; Garcés, E.; Magnani, M. Development of a Real-Time PCR Assay for Rapid Detection and Quantification of Alexandrium Minutum (a Dinoflagellate). Appl. Environ. Microbiol. 2004, 70, 1199–1206. [Google Scholar] [CrossRef]
- Galluzzi, L.; Bertozzini, E.; Penna, A.; Perini, F.; Garcés, E.; Magnani, M. Analysis of rRNA Gene Content in the Mediterranean Dinoflagellate Alexandrium catenella and Alexandrium taylori: Implications for the Quantitative Real-Time PCR-Based Monitoring Methods. J. Appl. Phycol. 2010, 22, 1–9. [Google Scholar] [CrossRef]
- Lauritano, C.; Andersen, J.H.; Hansen, E.; Albrigtsen, M.; Escalera, L.; Esposito, F.; Helland, K.; Hanssen, K.Ø.; Romano, G.; Ianora, A. Bioactivity Screening of Microalgae for Antioxidant, Anti-Inflammatory, Anticancer, Anti-Diabetes, and Antibacterial Activities. Front. Mar. Sci. 2016, 3, 68. [Google Scholar] [CrossRef]
- Balech, E. The Genus Alexandrium halim (Dinoflagellata); Sherkin Island Marine Station: Sherkin Island, Ireland, 1995. [Google Scholar]
- Lim, A.S.; Jeong, H.J.; Ok, J.H. Five Alexandrium Species Lacking Mixotrophic Ability. ALGAE 2019, 34, 289–301. [Google Scholar] [CrossRef]
- Wyatt, T.; Jenkinson, I.R. Notes on Alexandrium Population Dynamics. J. Plankton Res. 1997, 19, 551–575. [Google Scholar] [CrossRef]
- Anderson, D.M. Physiology and Bloom Dynamics of Toxic Alexandrium Species, with Emphasis on Life Cycle Transitions. Nato Asi. Ser. G Ecol. Sci. 1998, 41, 29–48. [Google Scholar]
- Saide, A.; Martínez, K.A.; Ianora, A.; Lauritano, C. Unlocking the Health Potential of Microalgae as Sustainable Sources of Bioactive Compounds. IJMS 2021, 22, 4383. [Google Scholar] [CrossRef] [PubMed]
- Schantz, E.J.; Ghazarossian, V.E.; Schnoes, H.K.; Strong, F.M.; Springer, J.P.; Pezzanite, J.O.; Clardy, J. Structure of Saxitoxin. J. Am. Chem. Soc. 1975, 97, 1238–1239. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.-Z. Neurotoxins from Marine Dinoflagellates: A Brief Review. Mar. Drugs 2008, 6, 349–371. [Google Scholar] [CrossRef] [PubMed]
- Llewellyn, L.E. Saxitoxin, a Toxic Marine Natural Product That Targets a Multitude of Receptors. Nat. Prod. Rep. 2006, 23, 200. [Google Scholar] [CrossRef]
- Kim, H.-S.; Park, H.; Wang, H.; Kim, T.; Ki, J.-S. Saxitoxins-Producing Potential of the Marine Dinoflagellate Alexandrium affine and Its Environmental Implications Revealed by Toxins and Transcriptome Profiling. Mar. Environ. Res. 2023, 185, 105874. [Google Scholar] [CrossRef]
- Ciminiello, P.; Fattorusso, E.; Forino, M.; Montresor, M. Saxitoxin and Neosaxitoxin as Toxic Principles of Alexandrium andersoni (Dinophyceae) from the Gulf of Naples, Italy. Toxicon 2000, 38, 1871–1877. [Google Scholar] [CrossRef]
- Sampedro, N.; Franco, J.M.; Zapata, M.; Riobó, P.; Garcés, E.; Penna, A.; Caillaud, A.; Diogène, J.; Cacho, E.; Camp, J. The Toxicity and Intraspecific Variability of Alexandrium andersonii Balech. Harmful Algae 2013, 25, 26–38. [Google Scholar] [CrossRef]
- Navarro, J.M.; Contreras, A.M. An Integrative Response by Mytilus Chilensis to the Toxic Dinoflagellate Alexandrium catenella. Mar. Biol. 2010, 157, 1967–1974. [Google Scholar] [CrossRef]
- Tatters, A.O.; Flewelling, L.J.; Fu, F.; Granholm, A.A.; Hutchins, D.A. High CO2 Promotes the Production of Paralytic Shellfish Poisoning Toxins by Alexandrium catenella from Southern California Waters. Harmful Algae 2013, 30, 37–43. [Google Scholar] [CrossRef]
- Anderson, D.M.; Fachon, E.; Pickart, R.S.; Lin, P.; Fischer, A.D.; Richlen, M.L.; Uva, V.; Brosnahan, M.L.; McRaven, L.; Bahr, F.; et al. Evidence for Massive and Recurrent Toxic Blooms of Alexandrium catenella in the Alaskan Arctic. Proc. Natl. Acad. Sci. USA 2021, 118, e2107387118. [Google Scholar] [CrossRef] [PubMed]
- Eckford-Soper, L.K.; Bresnan, E.; Lacaze, J.-P.; Green, D.H.; Davidson, K. The Competitive Dynamics of Toxic Alexandrium Fundyense and Non-Toxic Alexandrium tamarense: The Role of Temperature. Harmful Algae 2016, 53, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Okita, Y.; Matsuda, H.; Okino, T.; Yamaguchi, K. From the Dinoflagellate Alexandrium hiranoi. Phytochemistry 1998, 48, 85–88. [Google Scholar] [CrossRef]
- Espiña, B.; Cagide, E.; Louzao, M.C.; Vilariño, N.; Vieytes, M.R.; Takeda, Y.; Sasaki, M.; Botana, L.M. Cytotoxicity of Goniodomin A and B in Non Contractile Cells. Toxicol. Lett. 2016, 250–251, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Zhai, X.; Hao, S.; Shang, L.; Deng, Y.; Chai, Z.; Chen, J.; Hu, Z.; Tang, Y.Z. Novel Non-Paralytic Shellfish Toxin and Non-Spirolide Toxicity to Finfish, Brine Shrimp, and Rotifer Observed in a Culture of the Dinoflagellate Alexandrium insuetum Isolated From the Coastal Water of China. Front. Mar. Sci. 2021, 8, 735752. [Google Scholar] [CrossRef]
- Shang, L.; Xu, Y.; Leaw, C.P.; Lim, P.T.; Wang, J.; Chen, J.; Deng, Y.; Hu, Z.; Tang, Y.Z. Potent Allelopathy and Non-PSTs, Non-Spirolides Toxicity of the Dinoflagellate Alexandrium leei to Phytoplankton, Finfish and Zooplankton Observed from Laboratory Bioassays. Sci. Total Environ. 2021, 780, 146484. [Google Scholar] [CrossRef]
- Penna, A.; Perini, F.; Dell’Aversano, C.; Capellacci, S.; Tartaglione, L.; Giacobbe, M.G.; Casabianca, S.; Fraga, S.; Ciminiello, P.; Scardi, M. The Sxt Gene and Paralytic Shellfish Poisoning Toxins as Markers for the Monitoring of Toxic Alexandrium Species Blooms. Environ. Sci. Technol. 2015, 49, 14230–14238. [Google Scholar] [CrossRef]
- Hsia, M.H.; Morton, S.L.; Smith, L.L.; Beauchesne, K.R.; Huncik, K.M.; Moeller, P.D.R. Production of Goniodomin A by the Planktonic, Chain-Forming Dinoflagellate Alexandrium monilatum (Howell) Balech Isolated from the Gulf Coast of the United States. Harmful Algae 2006, 5, 290–299. [Google Scholar] [CrossRef]
- May, S.P.; Burkholder, J.M.; Shumway, S.E.; Hégaret, H.; Wikfors, G.H.; Frank, D. Effects of the Toxic Dinoflagellate Alexandrium monilatum on Survival, Grazing and Behavioral Response of Three Ecologically Important Bivalve Molluscs. Harmful Algae 2010, 9, 281–293. [Google Scholar] [CrossRef]
- Mackenzie, L.; White, D.; Oshima, Y.; Kapa, J. The Resting Cyst and Toxicity of Alexandrium ostenfeldii (Dinophyceae) in New Zealand. Phycologia 1996, 35, 148–155. [Google Scholar] [CrossRef]
- Cembella, A.D.; Lewis, N.I.; Quilliam, M.A. The Marine Dinoflagellate Alexandrium ostenfeldii (Dinophyceae) as the Causative Organism of Spirolide Shellfish Toxins. Phycologia 2000, 39, 67–74. [Google Scholar] [CrossRef]
- Gribble, K.E.; Keafer, B.A.; Quilliam, M.A.; Cembella, A.D.; Kulis, D.M.; Manahan, A.; Anderson, D.M. Distribution and Toxicity of Alexandrium ostenfeldii (Dinophyceae) in the Gulf of Maine, USA. Deep. Sea Res. Part. II Top. Stud. Oceanogr. 2005, 52, 2745–2763. [Google Scholar] [CrossRef]
- Borkman, D.G.; Smayda, T.J.; Tomas, C.R.; York, R.; Strangman, W.; Wright, J.L.C. Toxic Alexandrium peruvianum (Balech and de Mendiola) Balech and Tangen in Narragansett Bay, Rhode Island (USA). Harmful Algae 2012, 19, 92–100. [Google Scholar] [CrossRef]
- Tomas, C.R.; Van Wagoner, R.; Tatters, A.O.; White, K.D.; Hall, S.; Wright, J.L.C. Alexandrium peruvianum (Balech and Mendiola) Balech and Tangen a New Toxic Species for Coastal North Carolina. Harmful Algae 2012, 17, 54–63. [Google Scholar] [CrossRef]
- Blossom, H.E.; Daugbjerg, N.; Hansen, P.J. Toxic Mucus Traps: A Novel Mechanism That Mediates Prey Uptake in the Mixotrophic Dinoflagellate Alexandrium pseudogonyaulax. Harmful Algae 2012, 17, 40–53. [Google Scholar] [CrossRef]
- Zmerli Triki, H.; Laabir, M.; Moeller, P.; Chomérat, N.; Kéfi Daly-Yahia, O. First Report of Goniodomin a Production by the Dinoflagellate Alexandrium pseudogonyaulax Developing in Southern Mediterranean (Bizerte Lagoon, Tunisia). Toxicon 2016, 111, 91–99. [Google Scholar] [CrossRef]
- Collins, C.; Graham, J.; Brown, L.; Bresnan, E.; Lacaze, J.-P.; Turrell, E.A. Identification and toxicity of Alexandrium tamarense (dinophyceae) in scottish waters. J. Phycol. 2009, 45, 692–703. [Google Scholar] [CrossRef]
- Ichimi, K.; Suzuki, T.; Ito, A. Variety of PSP Toxin Profiles in Various Culture Strains of Alexandrium tamarense and Change of Toxin Profile in Natural A. tamarense Population. J. Exp. Mar. Biol. Ecol. 2002, 273, 51–60. [Google Scholar] [CrossRef]
- Hashimoto, T.; Matsuoka, S.; Yoshimatsu, S.; Miki, K.; Nishibori, N.; Nishio, S.; Noguchi, T. First Paralytic Shellfish Poison (PSP) Infestation of Bivalves Due to Toxic Dinoflagellate Alexandrium tamiyavanichii, in the Southeast Coasts of the Seto Inland Sea, Japan. J. Food Hyg. Soc. Jpn. 2002, 43, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Emura, A.; Matsuyama, Y.; Oda, T. Evidence for the Production of a Novel Proteinaceous Hemolytic Exotoxin by Dinoflagellate Alexandrium taylorii. Harmful Algae 2004, 3, 29–37. [Google Scholar] [CrossRef]
- Tillmann, U.; Krock, B.; Wietkamp, S.; Beran, A. A Mediterranean Alexandrium taylorii (Dinophyceae) Strain Produces Goniodomin A and Lytic Compounds but Not Paralytic Shellfish Toxins. Toxins 2020, 12, 564. [Google Scholar] [CrossRef] [PubMed]
- Cusick, K.; Sayler, G. An Overview on the Marine Neurotoxin, Saxitoxin: Genetics, Molecular Targets, Methods of Detection and Ecological Functions. Mar. Drugs 2013, 11, 991–1018. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, L.; Miller, A.; Kosatsky, T. Changing Trends in Paralytic Shellfish Poisonings Reflect Increasing Sea Surface Temperatures and Practices of Indigenous and Recreational Harvesters in British Columbia, Canada. Mar. Drugs 2021, 19, 568. [Google Scholar] [CrossRef] [PubMed]
- Van Wagoner, R.M.; Misner, I.; Tomas, C.R.; Wright, J.L.C. Occurrence of 12-Methylgymnodimine in a Spirolide-Producing Dinoflagellate Alexandrium peruvianum and the Biogenetic Implications. Tetrahedron Lett. 2011, 52, 4243–4246. [Google Scholar] [CrossRef]
- Hansen, P.J.; Cembella, A.D.; Moestrup, Ø. The marine dinoflagellate Alexandrium ostenfeldii: Paralytic shellfish toxin concentration, composition, and toxicity to a tintinnid ciliate. J. Phycol. 1992, 28, 597–603. [Google Scholar] [CrossRef]
- Harris, C.M.; Reece, K.S.; Stec, D.F.; Scott, G.P.; Jones, W.M.; Hobbs, P.L.M.; Harris, T.M. The Toxin Goniodomin, Produced by Alexandrium Spp., Is Identical to Goniodomin A. Harmful Algae 2020, 92, 101707. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. PubChem Compound Summary for CID:56947150, Saxitoxin. 2023. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/56947150#section=2D-Structure (accessed on 19 August 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID: 21590535, Spirolide A. 2023. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/21590535#section=2D-Structure (accessed on 12 December 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID: 101136678, Spirolide C. 2023. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/101136678#section=2D-Structure (accessed on 12 December 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID: 11649137, Gymnodimine A. 2023. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/11649137#section=2D-Structure (accessed on 13 December 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID: 101136679, 13-Desmethyl Spirolide C. 2023. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/101136679#section=2D-Structure (accessed on 29 August 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID: 6506119, Goniodomin A. 2023. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/6506119#section=2D-Structure (accessed on 30 August 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID: 11174599, Tetrodoxin. 2023. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/tetrodotoxin#section=2D-Structure (accessed on 11 December 2023).
- Wiese, M.; D’Agostino, P.M.; Mihali, T.K.; Moffitt, M.C.; Neilan, B.A. Neurotoxic Alkaloids: Saxitoxin and Its Analogs. Mar. Drugs 2010, 8, 2185–2211. [Google Scholar] [CrossRef]
- Bordner, J.; Thiessen, W.E.; Bates, H.A.; Rapoport, H. Structure of a Crystalline Derivative of Saxitoxin. Structure of Saxitoxin. J. Am. Chem. Soc. 1975, 97, 6008–6012. [Google Scholar] [CrossRef]
- Wang, D.-Z.; Zhang, S.-F.; Zhang, Y.; Lin, L. Paralytic Shellfish Toxin Biosynthesis in Cyanobacteria and Dinoflagellates: A Molecular Overview. J. Proteom. 2016, 135, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Durán-Riveroll, L.; Cembella, A. Guanidinium Toxins and Their Interactions with Voltage-Gated Sodium Ion Channels. Mar. Drugs 2017, 15, 303. [Google Scholar] [CrossRef] [PubMed]
- Mattei, C.; Legros, C. The Voltage-Gated Sodium Channel: A Major Target of Marine Neurotoxins. Toxicon 2014, 91, 84–95. [Google Scholar] [CrossRef] [PubMed]
- England, S.; De Groot, M.J. Subtype-selective Targeting of Voltage-gated Sodium Channels. Br. J. Pharmacol. 2009, 158, 1413–1425. [Google Scholar] [CrossRef]
- Su, Z.; Sheets, M.; Ishida, H.; Li, F.; Barry, W.H. Saxitoxin Blocks L-Type I Ca. J. Pharmacol. Exp. Ther. 2004, 308, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Salata, J.J.; Bennett, P.B. Saxitoxin Is a Gating Modifier of hERG K+ Channels. J. Gen. Physiol. 2003, 121, 583–598. [Google Scholar] [CrossRef]
- Zhang, F.; Xu, X.; Li, T.; Liu, Z. Shellfish Toxins Targeting Voltage-Gated Sodium Channels. Mar. Drugs 2013, 11, 4698–4723. [Google Scholar] [CrossRef]
- Kao, C.Y. Paralytic Shellfish Poisoning. In Algal Toxins in Seafood and Drinking Water; Elsevier: Amsterdam, The Netherlands, 1993; pp. 75–86. ISBN 978-0-12-247990-8. [Google Scholar]
- Kohane, D.; Lu, N.; Gokgolkline, A.; Shubina, M.; Kuang, Y.; Hall, S.; Strichartz, G.; Berde, C. The Local Anesthetic Properties and Toxicity of Saxitonin Homologues for Rat Sciatic Nerve Block in Vivo. Reg. Anesth. Pain. Med. 2000, 25, 52–59. [Google Scholar] [CrossRef]
- Xu, Y.; He, X.; Li, H.; Zhang, T.; Lei, F.; Gu, H.; Anderson, D.M. Molecular Identification and Toxin Analysis of Alexandrium Spp. in the Beibu Gulf: First Report of Toxic A. Tamiyavanichii in Chinese Coastal Waters. Toxins 2021, 13, 161. [Google Scholar] [CrossRef]
- Lattes, K.; Venegas, P.; Lagos, N.; Lagos, M.; Pedraza, L.; Rodriguez-Navarro, A.J.; García, C. Local Infiltration of Gonyautoxin Is Safe and Effective in Treatment of Chronic Tension-Type Headache. Neurol. Res. 2009, 31, 228–233. [Google Scholar] [CrossRef]
- Molgó, J.; Marchot, P.; Aráoz, R.; Benoit, E.; Iorga, B.I.; Zakarian, A.; Taylor, P.; Bourne, Y.; Servent, D. Cyclic Imine Toxins from Dinoflagellates: A Growing Family of Potent Antagonists of the Nicotinic Acetylcholine Receptors. J. Neurochem. 2017, 142, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.; Murphy, M.; Clausen, J.; Richard, D.; Quilliam, M.; MacKinnon, S.; LaBlanc, P.; Mueller, R.; Pulido, O. Neural Injury Biomarkers of Novel Shellfish Toxins, Spirolides: A Pilot Study Using Immunochemical and Transcriptional Analysis. NeuroToxicology 2003, 24, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Guéret, S.M.; Brimble, M.A. Spiroimine Shellfish Poisoning (SSP) and the Spirolide Family of Shellfish Toxins: Isolation, Structure, Biological Activity and Synthesis. Nat. Prod. Rep. 2010, 27, 1350. [Google Scholar] [CrossRef] [PubMed]
- Otero, A.; Chapela, M.-J.; Atanassova, M.; Vieites, J.M.; Cabado, A.G. Cyclic Imines: Chemistry and Mechanism of Action: A Review. Chem. Res. Toxicol. 2011, 24, 1817–1829. [Google Scholar] [CrossRef] [PubMed]
- Stivala, C.E.; Benoit, E.; Aráoz, R.; Servent, D.; Novikov, A.; Molgó, J.; Zakarian, A. Synthesis and Biology of Cyclic Imine Toxins, an Emerging Class of Potent, Globally Distributed Marine Toxins. Nat. Prod. Rep. 2015, 32, 411–435. [Google Scholar] [CrossRef] [PubMed]
- Stivala, C.E.; Zakarian, A. Studies Toward the Synthesis of Spirolides: Assembly of the Elaborated E-Ring Fragment. Org. Lett. 2009, 11, 839–842. [Google Scholar] [CrossRef] [PubMed]
- Zurhelle, C.; Nieva, J.; Tillmann, U.; Harder, T.; Krock, B.; Tebben, J. Identification of Novel Gymnodimines and Spirolides from the Marine Dinoflagellate Alexandrium ostenfeldii. Mar. Drugs 2018, 16, 446. [Google Scholar] [CrossRef]
- Albuquerque, E.X.; Pereira, E.F.R.; Alkondon, M.; Rogers, S.W. Mammalian Nicotinic Acetylcholine Receptors: From Structure to Function. Physiol. Rev. 2009, 89, 73–120. [Google Scholar] [CrossRef]
- Kharrat, R.; Servent, D.; Girard, E.; Ouanounou, G.; Amar, M.; Marrouchi, R.; Benoit, E.; Molgó, J. The Marine Phycotoxin Gymnodimine Targets Muscular and Neuronal Nicotinic Acetylcholine Receptor Subtypes with High Affinity. J. Neurochem. 2008, 107, 952–963. [Google Scholar] [CrossRef]
- Hauser, T.A.; Hepler, C.D.; Kombo, D.C.; Grinevich, V.P.; Kiser, M.N.; Hooker, D.N.; Zhang, J.; Mountfort, D.; Selwood, A.; Akireddy, S.R.; et al. Comparison of Acetylcholine Receptor Interactions of the Marine Toxins, 13-Desmethylspirolide C and Gymnodimine. Neuropharmacology 2012, 62, 2239–2250. [Google Scholar] [CrossRef]
- Munday, R.; Towers, N.R.; Mackenzie, L.; Beuzenberg, V.; Holland, P.T.; Miles, C.O. Acute Toxicity of Gymnodimine to Mice. Toxicon 2004, 44, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Munday, R.; Quilliam, M.A.; LeBlanc, P.; Lewis, N.; Gallant, P.; Sperker, S.A.; Ewart, H.S.; MacKinnon, S.L. Investigations into the Toxicology of Spirolides, a Group of Marine Phycotoxins. Toxins 2011, 4, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Burton, I.W.; Cembella, A.D.; Curtis, J.M.; Quilliam, M.A.; Walter, J.A.; Wright, J.L.C. Characterization of Spirolides A, C, and 13-Desmethyl C, New Marine Toxins Isolated from Toxic Plankton and Contaminated Shellfish. J. Nat. Prod. 2001, 64, 308–312. [Google Scholar] [CrossRef] [PubMed]
- Alonso, E.; Otero, P.; Vale, C.; Alfonso, A.; Antelo, A.; Gimenez-Llort, L.; Chabaud, L.; Guillou, C.; Botana, M.L. Benefit of 13-Desmethyl Spirolide C Treatment in Triple Transgenic Mouse Model of Alzheimer Disease: Beta-Amyloid and Neuronal Markers Improvement. CAR 2013, 10, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Makabe, K.; Yamaguchi, K.; Konosu, S.; Wälchli, M.R. Goniodomin a, a Novel Polyether Macrolide from the Dinoflagellate Goniodoma pseudogoniaulax. Tetrahedron Lett. 1988, 29, 1149–1152. [Google Scholar] [CrossRef]
- Furukawa, K.; Sakai, K.; Watanabe, S.; Maruyama, K.; Murakami, M.; Yamaguchi, K.; Ohizumi, Y. Goniodomin A Induces Modulation of Actomyosin ATPase Activity Mediated through Conformational Change of Actin. J. Biol. Chem. 1993, 268, 26026–26031. [Google Scholar] [CrossRef] [PubMed]
- Abe, M.; Inoue, D.; Matsunaga, K.; Ohizumi, Y.; Ueda, H.; Asano, T.; Murakami, M.; Sato, Y. Goniodomin A, an Antifungal Polyether Macrolide, Exhibits Antiangiogenic Activities via Inhibition of Actin Reorganization in Endothelial Cells. J. Cell. Physiol. 2002, 190, 109–116. [Google Scholar] [CrossRef]
- Mizuno, K.; Nakahata, N.; Ito, E.; Murakami, M.; Yamaguchi, K.; Ohizumi, Y. Goniodomin A, an Antifungal Polyether Macrolide, Increases the Filamentous Actin Content of 1321N1 Human Astrocytoma Cells. J. Pharm. Pharmacol. 1998, 50, 645–648. [Google Scholar] [CrossRef]
- Llewellyn, L.E. Sodium Channel Inhibiting Marine Toxins. In Marine Toxins as Research Tools; Fusetani, N., Kem, W., Eds.; Progress in Molecular and Subcellular Biology; Springer: Berlin/Heidelberg, Germany, 2009; Volume 46, pp. 67–97. ISBN 978-3-540-87892-6. [Google Scholar]
- Kodama, M.; Sato, S.; Sakamoto, S.; Ogata, T. Occurrence of Tetrodotoxin in Alexandrium tamarense, a Causative Dinoflagellate of Paralytic Shellfish Poisoning. Toxicon 1996, 34, 1101–1105. [Google Scholar] [CrossRef]
- Tsuda, K.; Ikuma, S.; Kawamura, M.; Tachikawa, R.; Sakai, K.; Tamura, C.; Amakasu, O. Tetrodotoxin. VII. On the Structures of Tetrodotoxin and Its Derivatives. Chem. Pharm. Bull. 1964, 12, 1357–1374. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Sugimoto, A.; Takai, A.; Yasumoto, T. Effects of Specific Modifications of Several Hydroxyls of Tetrodotoxin on Its Affinity to Rat Brain Membrane. J. Pharmacol. Exp. Ther. 1999, 289, 1688–1696. [Google Scholar] [PubMed]
- Bane, V.; Lehane, M.; Dikshit, M.; O’Riordan, A.; Furey, A. Tetrodotoxin: Chemistry, Toxicity, Source, Distribution and Detection. Toxins 2014, 6, 693–755. [Google Scholar] [CrossRef] [PubMed]
- Chau, R.; Kalaitzis, J.A.; Neilan, B.A. On the Origins and Biosynthesis of Tetrodotoxin. Aquat. Toxicol. 2011, 104, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Kao, C.Y. Actions of Chiriquitoxin on Frog Skeletal Muscle Fibers and Implications for the Tetrodotoxin/Saxitoxin Receptor. J. Gen. Physiol. 1992, 100, 609–622. [Google Scholar] [CrossRef] [PubMed]
- Denac, H.; Mevissen, M.; Scholtysik, G. Structure, Function and Pharmacology of Voltage-Gated Sodium Channels. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2000, 362, 453–479. [Google Scholar] [CrossRef] [PubMed]
- Hille, B. The Permeability of the Sodium Channel to Organic Cations in Myelinated Nerve. J. Gen. Physiol. 1971, 58, 599–619. [Google Scholar] [CrossRef] [PubMed]
- Catterall, W.A. Cellular and Molecular Biology of Voltage-Gated Sodium Channels. Physiol. Rev. 1992, 72, S15–S48. [Google Scholar] [CrossRef]
- Walker, J.R.; Novick, P.A.; Parsons, W.H.; McGregor, M.; Zablocki, J.; Pande, V.S.; Du Bois, J. Marked Difference in Saxitoxin and Tetrodotoxin Affinity for the Human Nociceptive Voltage-Gated Sodium Channel (Na v 1.7). Proc. Natl. Acad. Sci. USA 2012, 109, 18102–18107. [Google Scholar] [CrossRef]
- Lee, C.H.; Ruben, P.C. Interaction between Voltage-Gated Sodium Channels and the Neurotoxin, Tetrodotoxin. Channels 2008, 2, 407–412. [Google Scholar] [CrossRef]
- Hwang, D.; Noguchi, T. Tetrodotoxin Poisoning. In Advances in Food and Nutrition Research; Elsevier: Amsterdam, The Netherlands, 2007; Volume 52, pp. 141–236. ISBN 978-0-12-373711-3. [Google Scholar]
- Lago, J.; Rodríguez, L.; Blanco, L.; Vieites, J.; Cabado, A. Tetrodotoxin, an Extremely Potent Marine Neurotoxin: Distribution, Toxicity, Origin and Therapeutical Uses. Mar. Drugs 2015, 13, 6384–6406. [Google Scholar] [CrossRef]
- Noguchi, T.; Ebesu, J.S.M. Puffer Poisoing: Epidemiologi and Tretment. J. Toxicol. Toxin Rev. 2001, 20, 1–10. [Google Scholar] [CrossRef]
- Saoudi, M.; Abdelmouleh, A.; El Feki, A. Tetrodotoxin: A Potent Marine Toxin. Toxin Rev. 2010, 29, 60–70. [Google Scholar] [CrossRef]
- How, C.-K.; Chern, C.-H.; Huang, Y.-C.; Wang, L.-M.; Lee, C.-H. Tetrodotoxin Poisoning. Am. J. Emerg. Med. 2003, 21, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Sims, J.K.; Ostman, D.C. Pufferfish Poisoning: Emergency Diagnosis and Management of Mild Human Tetrodotoxication. Ann. Emerg. Med. 1986, 15, 1094–1098. [Google Scholar] [CrossRef]
- Lange, W.R. Puffer Fish Poisoning. Am. Fam. Physician 1990, 42, 1029–1033. [Google Scholar] [PubMed]
- Nieto, F.R.; Cobos, E.J.; Tejada, M.Á.; Sánchez-Fernández, C.; González-Cano, R.; Cendán, C.M. Tetrodotoxin (TTX) as a Therapeutic Agent for Pain. Mar. Drugs 2012, 10, 281–305. [Google Scholar] [CrossRef] [PubMed]
- Hagen, N.A.; Cantin, L.; Constant, J.; Haller, T.; Blaise, G.; Ong-Lam, M.; Du Souich, P.; Korz, W.; Lapointe, B. Tetrodotoxin for Moderate to Severe Cancer-Related Pain: A Multicentre, Randomized, Double-Blind, Placebo-Controlled, Parallel-Design Trial. Pain Res. Manag. 2017, 2017, 7212713. [Google Scholar] [CrossRef]
- Galasso, C.; Nuzzo, G.; Brunet, C.; Ianora, A.; Sardo, A.; Fontana, A.; Sansone, C. The Marine Dinoflagellate Alexandrium minutum Activates a Mitophagic Pathway in Human Lung Cancer Cells. Mar. Drugs 2018, 16, 502. [Google Scholar] [CrossRef]
- Yamasaki, Y.; Katsuo, D.; Nakayasu, S.; Salati, C.; Duan, J.; Zou, Y.; Matsuyama, Y.; Yamaguchi, K.; Oda, T. Purification and Characterization of a Novel High Molecular Weight Exotoxin Produced by Red Tide Phytoplankton, Alexandrium tamarense. J. Biochem. Mol. Toxicol. 2008, 22, 405–415. [Google Scholar] [CrossRef]
- Sansone, C.; Nuzzo, G.; Galasso, C.; Casotti, R.; Fontana, A.; Romano, G.; Ianora, A. The Marine Dinoflagellate Alexandrium andersoni Induces Cell Death in Lung and Colorectal Tumor Cell Lines. Mar. Biotechnol. 2018, 20, 343–352. [Google Scholar] [CrossRef]
- De Vera, C.; Díaz Crespín, G.; Hernández Daranas, A.; Montalvão Looga, S.; Lillsunde, K.-E.; Tammela, P.; Perälä, M.; Hongisto, V.; Virtanen, J.; Rischer, H.; et al. Marine Microalgae: Promising Source for New Bioactive Compounds. Mar. Drugs 2018, 16, 317. [Google Scholar] [CrossRef] [PubMed]
- Long, M.; Marie, D.; Szymczak, J.; Toullec, J.; Bigeard, E.; Sourisseau, M.; Le Gac, M.; Guillou, L.; Jauzein, C. Dinophyceae Can Use Exudates as Weapons against the Parasite Amoebophrya sp. (Syndiniales). ISME Commun. 2021, 1, 34. [Google Scholar] [CrossRef] [PubMed]
- Lassudrie, M.; Soudant, P.; Nicolas, J.-L.; Fabioux, C.; Lambert, C.; Miner, P.; Le Grand, J.; Petton, B.; Hégaret, H. Interaction between Toxic Dinoflagellate Alexandrium catenella Exposure and Disease Associated with Herpesvirus OsHV-1 μVar in Pacific Oyster Spat Crassostrea Gigas. Harmful Algae 2015, 45, 53–61. [Google Scholar] [CrossRef]
- Lauritano, C.; Martín, J.; De La Cruz, M.; Reyes, F.; Romano, G.; Ianora, A. First Identification of Marine Diatoms with Anti-Tuberculosis Activity. Sci. Rep. 2018, 8, 2284. [Google Scholar] [CrossRef]
- Chen, L.; Giesy, J.P.; Adamovsky, O.; Svirčev, Z.; Meriluoto, J.; Codd, G.A.; Mijovic, B.; Shi, T.; Tuo, X.; Li, S.-C.; et al. Challenges of Using Blooms of Microcystis Spp. in Animal Feeds: A Comprehensive Review of Nutritional, Toxicological and Microbial Health Evaluation. Sci. Total Environ. 2021, 764, 142319. [Google Scholar] [CrossRef]
- Bashir, F.; Bashir, A.; Bouaïcha, N.; Chen, L.; Codd, G.A.; Neilan, B.; Xu, W.-L.; Ziko, L.; Rajput, V.D.; Minkina, T.; et al. Cyanotoxins, Biosynthetic Gene Clusters, and Factors Modulating Cyanotoxin Biosynthesis. World J. Microbiol. Biotechnol. 2023, 39, 241. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
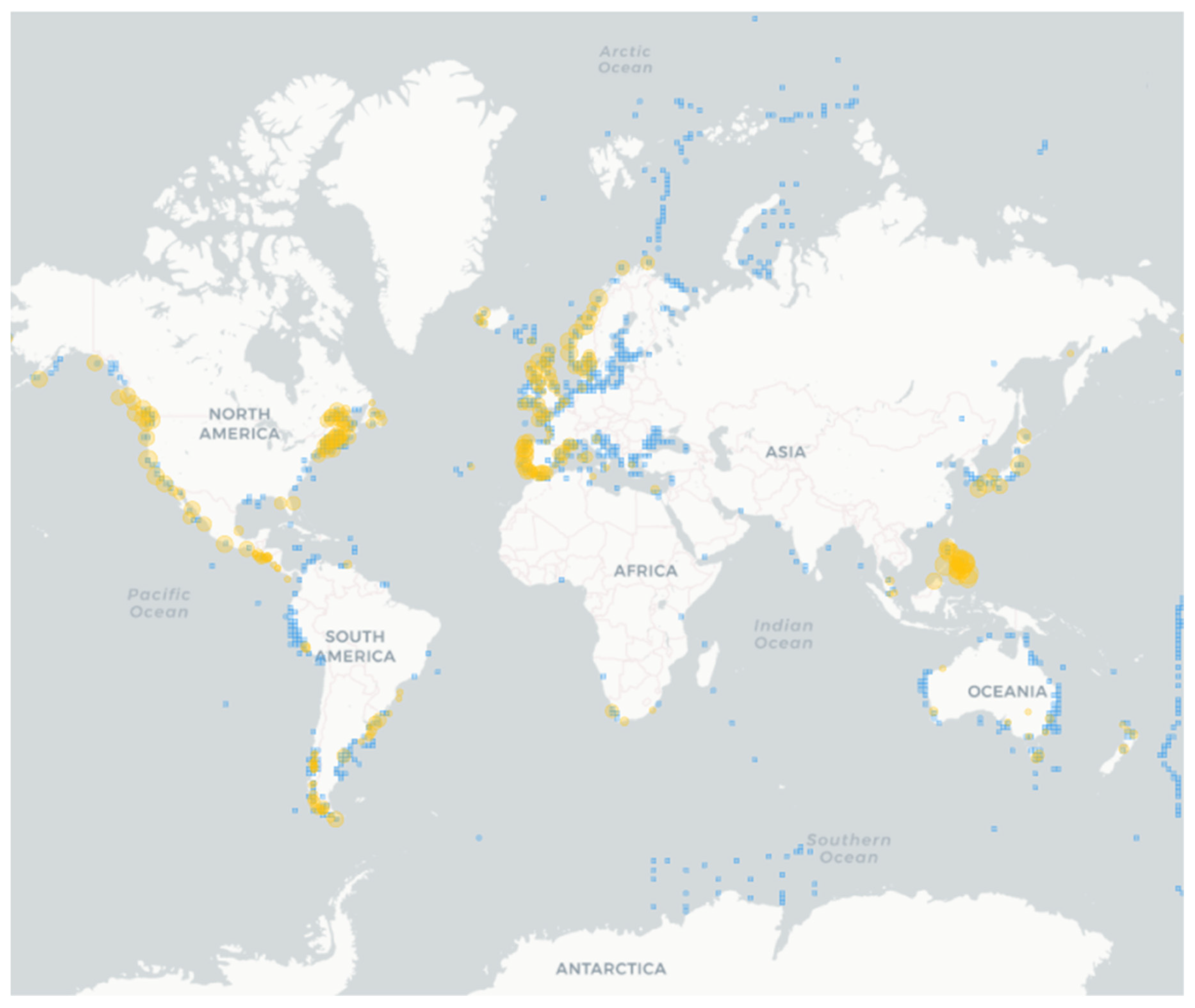{kind=link}
{kind=link}
| Species | Toxins | Effects | References |
|---|---|---|---|
| A. affine (H. Inoue and Y. Fukuyo) Balech | Saxitoxins (only genes found) | Not evaluated | [41] |
| A. andersonii Balech * | Saxitoxins (GTX2, NEO, STX) | Inhibition of voltage-gated sodium channels, hemolytic activity | [42,43] |
| A. catenella (Whedon and Kofoid) Balech | Saxitoxins (GTX1, GTX4, NEO, STX) | PSP | [44,45,46] |
| A. fundyense Balech | Saxitoxins (GTX3, Neo, STX) | PSP | [47] |
| A. hiranoi Kita and Fukuyo | Goniodomin A | Cytotoxic, disturbs the actomyosin ATPase activity in diverse cell types | [48,49] |
| A. insuetum Balech | Not characterized | PSP | [50] |
| A. leei Balech | Not characterized, but neither PSTs nor spirolides | Mortality of finfish and rotifers | [51] |
| A. minutum Halim * | Saxitoxins (C1, C2, NEO, GTX1, GTX2, GTX3, GTX4) | PSP | [23,52] |
| A. monilatum (Howell) Balech | Goniodomin A | Highly toxic to fish and shellfish | [53,54] |
| A. ostenfeldii (Paulsen) Balech and Tangen | Saxitoxins (GTX3, GTX5); spirolides (A, B, C2, D2) | PSP (saxitoxins); toxicity to shellfish (spirolides) | [55,56,57] |
| A. peruvianum (Balech and Mendiola) | Hemolytic substances, Saxitoxins (B1, C1, C2, GTX2, GTX3, B1) Spirolides (C and D) | PSP | [58,59] |
| A. pseudogonyaulax (Biecheler) Horiguchi ex K. Yuki and Y. Fukuyo | Goniodomin A | Fish mortality | [60,61] |
| A. tamarense (Lebour) Balech * | Saxitoxins (C2, NEO, GTX3, GTX4) | PSP | [62,63] |
| A. tamiyavanichii Balech | Saxitoxins (C1, C2, GTX1, GTX2, GTX3, GTX5, STX) | PSP | [64] |
| A. tamutum | Saxitoxins (only genes found) | Antiproliferative activity on melanoma (A2058) and normal human lung fibroblast (MRC-5) | [26,32] |
| A. taylori Balech | Goniodomin A, Hemolytic substances | Effects on oyster larvae, sea-monkey mortality | [65,66] |
| Species | Sampling Location and Culture Conditions | Active Compounds/Extracts/Fractions | Bioactivities | In Vivo/In Vitro | Mechanism | References |
|---|---|---|---|---|---|---|
| Alexandrium andersonii | Mediterranean Sea [137]. - | Fraction B (digalactosyl diacyl glycerol and sulfoquinovosyl diacyl glycerol, were the predominant metabolites) | Cytotoxic (Anti-cancer) | In vitro investigation on: A549 cells | Down-regulation of anti-apoptotic molecules (BIRC3 and TRAF2) and up-regulation of death receptors (CD27, DR3, TNF, and TNFS8) | [133] |
| Alexandrium andersonii | Mediterranean Sea. - | Fraction D (rich in sterols and triacylglycerols) | Cytotoxic (Anti-cancer) | In vitro investigation on: HT29 cells | Induction of cell death with over expression of FASLG and TNF genes Up-regulation of Gadd45 alpha and gamma together with Foxl1 genes | [133] |
| Alexandrium catenella | Thau lagoon, France. Cultivated in L1 medium | - | Anti-viral | Herpes virus OsHV-1 μvar Oyster affected | Antagonist relationship that is established between the virus and A. catenella | [136] |
| Alexandrium minutum | Gulf of Naples. Cultivated in Keller medium | Fraction 3B (rich in carbohydrates (94%) and protein (4%)) | Cytotoxic (Anti-cancer) | In vitro investigation on: PC-3, HT29, A549 cells | Up-regulation of autophagy-related protein Atg12 with a consequent increase in ATP6V1G2 and BNIP3 Up-regulation of mitophagy genes PINK1 and Parkin gene | [131] |
| Alexandrium minutum | Coastal marine waters of the NE Atlantic Ocean. Cultivated in F/2 medium, 27 of salinity (Guillard’s Marine Water Enrichment Solution) and Keller medium, 35 of salinity | A. minutum exudates | Anti-parasite | Anti-parasite activity against Amoebophrya sp. | Permeation of the membranes of Amoebophrya sp. dinospores, leading to the loss of dinofluorescence associated in turn with the loss of virulence of the parasite in culture | [135] |
| Alexandrium ostenfeldii | Coastal marine waters of the NE Atlantic Ocean. Cultivated in F/2 medium, 27 of salinity (Guillard’s Marine Water Enrichment Solution) and Keller medium, 35 of salinity | 13-Desmethyl Spirolide C | Anti-Alzheimer | In vivo investigation in mice 3xTg-AD | Increase in NAA levels; Increase in synaptophytin and decrease in intracellular amyloid beta in the hypothalamus | [105] |
| Alexandrium sp. | - | Goniodomin A | Anti-angiogenic | - | Induction of morphological changes in human brain astrocytoma cells. Inhibition of ATPase activities of atrial myofibrils, natural actomyosin, and reconstitution of actomyosin. | [107,109] |
| Alexandrium sp. | - | Saxitoxin, neosaxitoxin, decarbamoyl saxitoxin, and tetrodotoxin | Rat sciatic nerve block (local anesthesia) | In vivo; evaluation of lethality, onset and duration of action for thermal analgesia (hot-plate testing), and motor block (weight-bearing) | Analgesia for 60 min with neosaxitoxin at 34 ± 2 micromol/L, saxitoxin at 58 ± 3 micromol/L, TTX at 92 ± 5 micromol/L, and decarbamoyl saxitoxin at 268 ± 8 micromol/L | [89] |
| Alexandrium sp. | - | Gonyautoxins 2/3 epimers | Treatment of Chronic headache | In vivo: investigation on 27 patients affected of chronic headache | Reversible block of the voltage-gated sodium channels at axonal level. | [91] |
| Alexandrium sp. | - | Tetrodoxins | Analgesic effects. | In vivo: subcutaneous injection of 30 µg. | Blockage of one subclass of sodium channels, NaV 1.7. | [130] |
| Alexandrium tamarense | Hiroshima Bay, Japan. Cultivated in ESM (Erd-Schreiber modified) medium pH8.2 | Polysaccharide-based AT-toxin | Cytotoxic (Anti-cancer) | In vitro investigation on: HeLa, Vero, XC, CHO, and U937 cells | Induction of cell death in U937 by DNA fragmentation | [132] |
| Alexandrium tamarense | Cultivated in Keller medium | Total Extract | Anti-proliferative (Anti-cancer) | In vitro investigation on: MCF-7, MCF-10A, PC-3, and LNCaP cells | - | [134] |
| Alexandrium tamarense | Cultivated in Keller medium | Total Extract | Apoptotic (Anti-cancer) | In vitro investigation on: HepG2 cells. | - | [134] |
| Alexandrium tamutum, A. andersonii, A. minutum | Mediterranean Sea. Cultivated in Keller medium | Total Extract | Anti-proliferative (Anti-cancer) | In vitro on melanoma A2058 and MRC-5 cells | - | [32] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montuori, E.; De Luca, D.; Penna, A.; Stalberga, D.; Lauritano, C. Alexandrium spp.: From Toxicity to Potential Biotechnological Benefits. Mar. Drugs 2024, 22, 31. https://doi.org/10.3390/md22010031
Montuori E, De Luca D, Penna A, Stalberga D, Lauritano C. Alexandrium spp.: From Toxicity to Potential Biotechnological Benefits. Marine Drugs. 2024; 22(1):31. https://doi.org/10.3390/md22010031
Chicago/Turabian StyleMontuori, Eleonora, Daniele De Luca, Antonella Penna, Darta Stalberga, and Chiara Lauritano. 2024. "Alexandrium spp.: From Toxicity to Potential Biotechnological Benefits" Marine Drugs 22, no. 1: 31. https://doi.org/10.3390/md22010031
APA StyleMontuori, E., De Luca, D., Penna, A., Stalberga, D., & Lauritano, C. (2024). Alexandrium spp.: From Toxicity to Potential Biotechnological Benefits. Marine Drugs, 22(1), 31. https://doi.org/10.3390/md22010031