Targets and Effects of Yessotoxin, Okadaic Acid and Palytoxin: A Differential Review

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Yessotoxin (YTX)
2.1. General proprieties
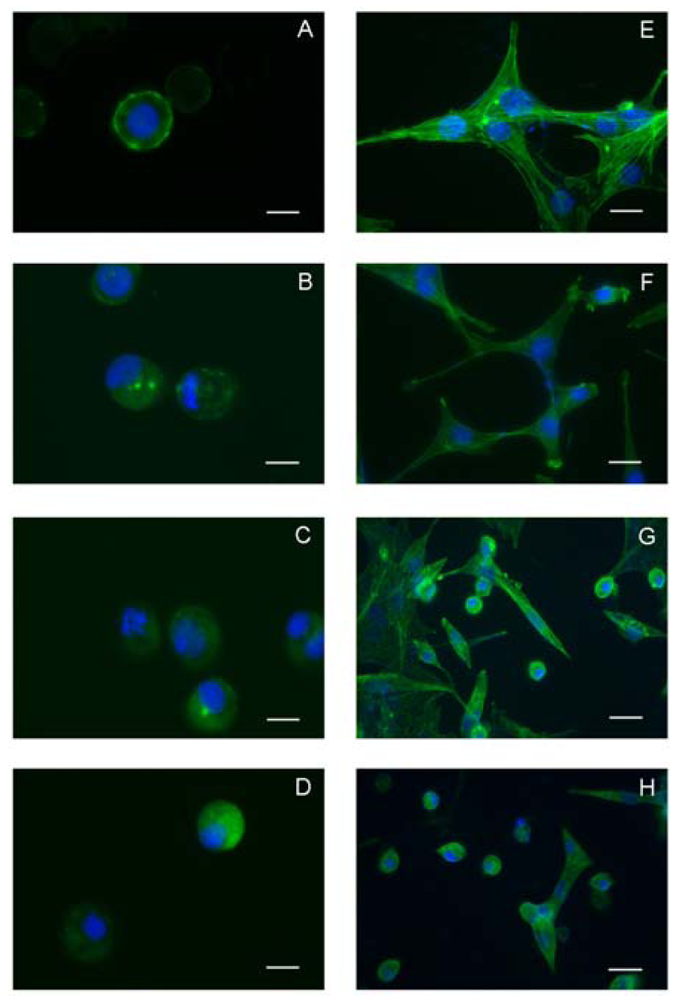
2.2. Data derived from experiments on invertebrate and vertebrate cell lines
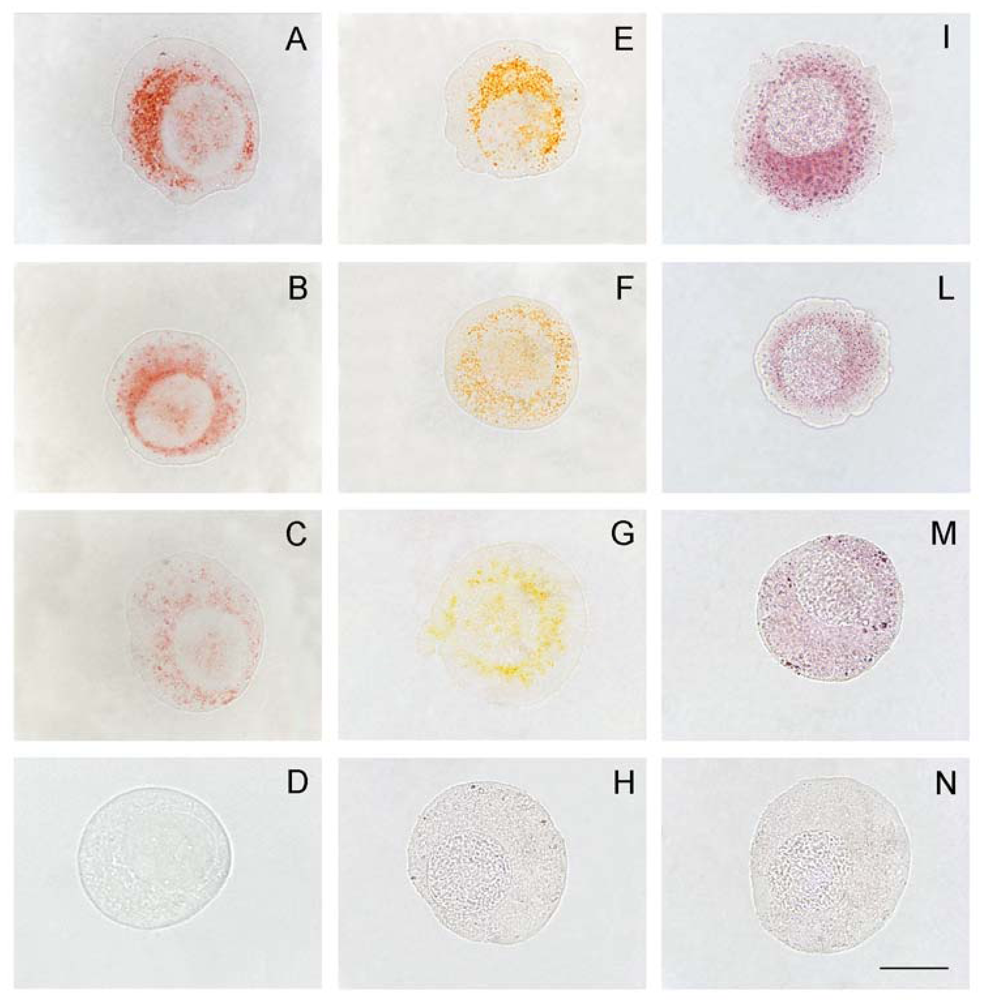
2.3. Data derived from experiments on the mussel Mytilus galloprovincialis
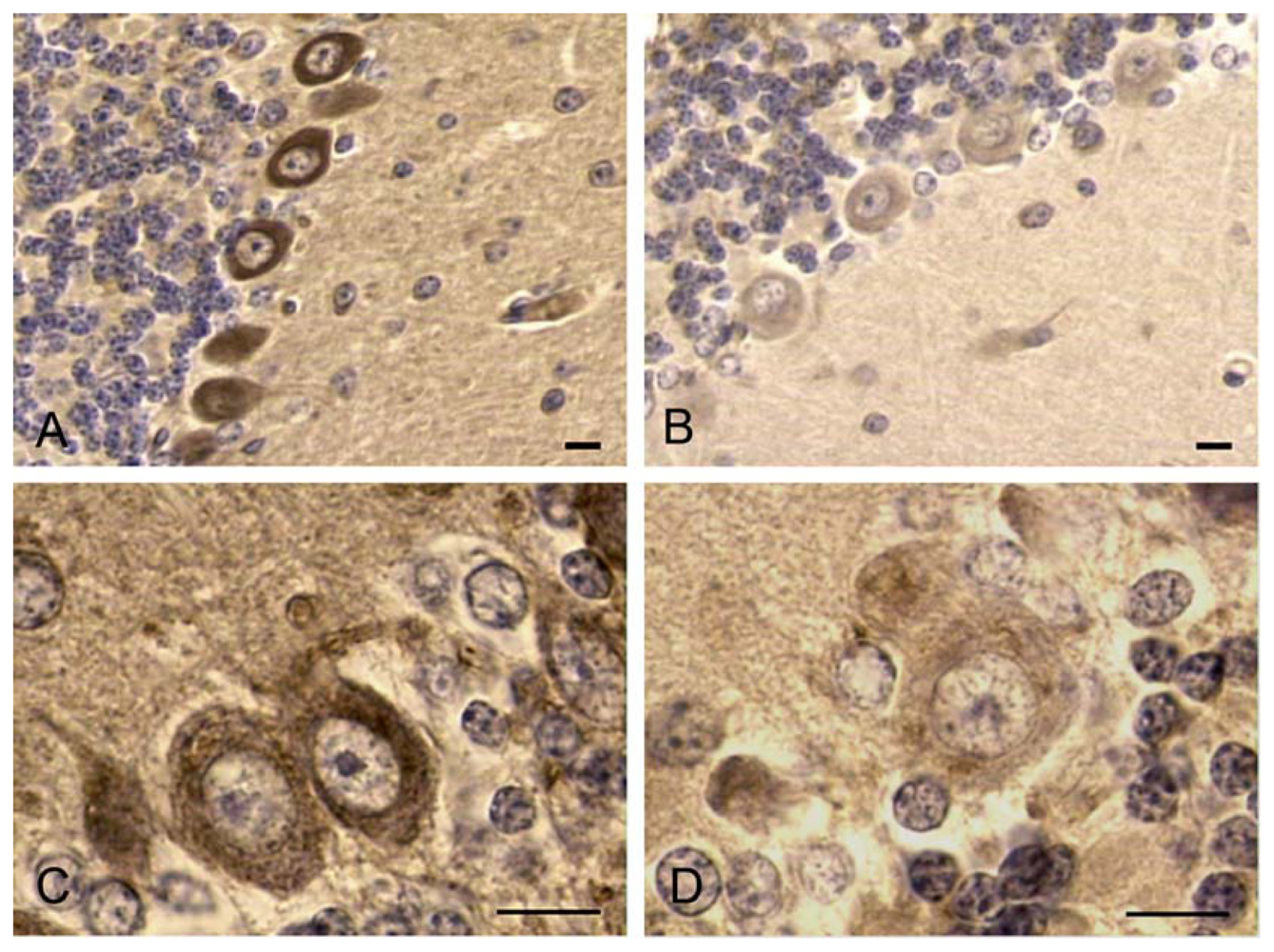
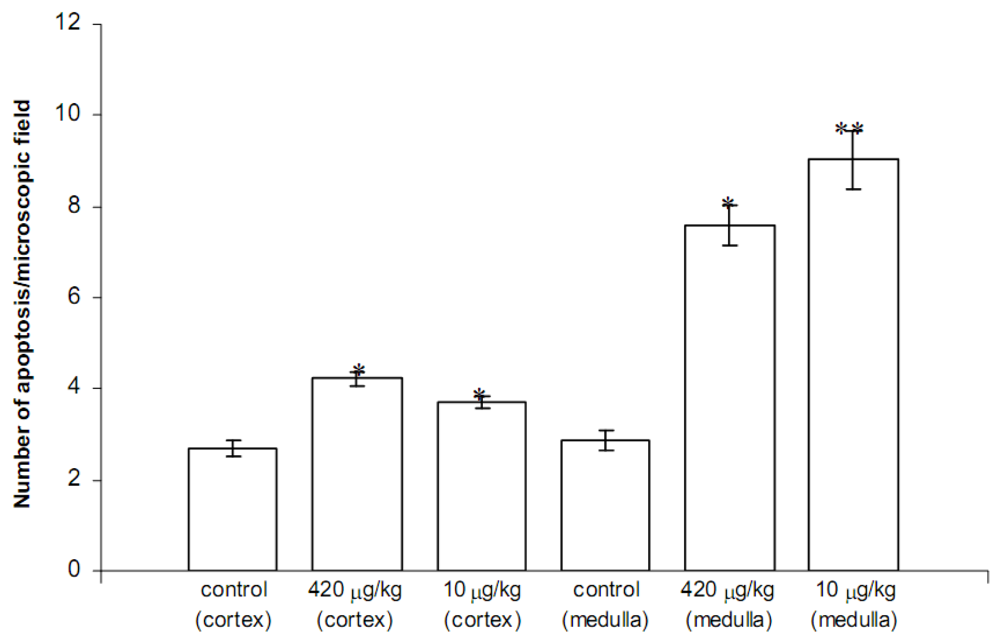
2.4. Data derived from experiments on Swiss CD1 mice
3. Okadaic Acid (OA) and Palytoxin (PTX)
3.1. General proprieties
3.2. Data derived from experiments on invertebrate models
3.3. Data derived from experiments on the amphibian X. laevis
4. Conclusions
Acknowledgements
- Samples Availability: Available from the authors.
References
- Wang, DZ. Neurotoxins from marine dinoflagellates: A brief review. Mar Drugs 2008, 6, 349–371. [Google Scholar]
- Geraci, JR; Anderson, DM; Timperi, RJ; St Aubin, DJ; Early, GA; Prescott, JH; Mayo, CA. Humpback whales (Megaoetera novaeangliae) fatally poisoned by dinoflagellate toxin. Can J Fish Aq Sc 1989, 46, 1895–1898. [Google Scholar]
- Landsberg, JH; Steidinger, K. A historical review of red tide events caused by Gymnodinium breve as related to mass mortalities of the endangered manatee (Trichechus manatus latirostris) in Florida, USA. In Harmful Microalgae; Reguera, B, Blanco, J, Fernandez, ML, Wyatt, T, Eds.; IOC-UNESCO: Xunta de Galicia, Spain, 1998; pp. 97–100. [Google Scholar]
- Scholin, CA; Gulland, F; Doucette, GJ; Benson, S; Busman, M; Chavez, FP; Cordaro, J; DeLong, R; De Vogelaere, A; Harvey, J; Haulena, M; Lefebvre, K; Lipscomb, T; Loscutoff, S; Lowenstine, LJ; Marin, R, III; Miller, PE; McLellan, WA; Moeller, PDR; Powell, CL; Rowles, T; Silvagni, P; Silver, M; Spraker, T; Trainer, V; Van Dolah, FM. Mortality of sea lions along the central California coast linked to a toxic diatom bloom. Nature 2000, 430, 80–84. [Google Scholar]
- Flewelling, LJ; Naar, JP; Abbott, JP; Baden, DG; Barros, NB; Bossart, GD; Bottein, MY; Hammond, DG; Haubold, EM; Heil, CA; Henry, MS; Jacocks, HM; Leighfield, TA; Pierce, RH; Pitchford, TD; Rommel, SA; Scott, PS; Steidinger, KA; Truby, EW; Van Dolah, FM; Landsberg, JH. Brevetoxicosis: Red tides and marine mammal mortalities. Nature 2005, 435, 755–756. [Google Scholar]
- Halstead, BW. Poisons and Venomous Animals of the World 1965, 3.
- Ade, P; Funari, E; Poletti, R. Risk to human health associated with marine toxic algae. Ann Ist Super Sanità 2003, 39, 53–68. [Google Scholar]
- Anderson, DM. Le maree rosse. Le Scienze 1994, 314, 74–81. [Google Scholar]
- Hallegraeff, GM. Harmful algal bloom: A global overview. In Manual on Harmful Marine Microalgae; Hallegraeff, GM, Anderson, DM, Cembella, AD, Eds.; IOC-UNESCO: Paris, France, 1995; pp. 1–22. [Google Scholar]
- Dounay, AB; Forsyth, CJ. Okadaic acid: The archetypal serine/threonine protein phosphatase inhibitor. Curr Med Chem 2002, 9, 1939–1980. [Google Scholar]
- Wattenberg, EV. Palytoxin: Exploiting a novel skin tumor promoter to explore signal transduction and carcinogenesis. Am J Physiol Cell Physiol 2007, 292, C24–32. [Google Scholar]
- Louzao, MC; Ares, IR; Cagide, E. Marine toxins and the cytoskeleton: A new view of palytoxin toxicity. FEBS J 2008, 275, 6067–6074. [Google Scholar]
- Vale, C; Botana, LM. Marine toxins and the cytoskeleton: Okadaic acid and dinophysistoxins. FEBS J 2008, 275, 6060–6066. [Google Scholar]
- Tubaro, A; Dell’Ovo, V; Sosa, S; Florio, C. Yessotoxins: A toxicological overview. Toxicon 2009. [Google Scholar] [CrossRef]
- Murata, M; Kumagai, M; Lee, JS; Yasumoto, T. Isolation and structure of yessotoxin, a novel polyether compound implicated in diarrhetic shellfish poisoning. Tetrahedron Lett 1987, 28, 5869–5872. [Google Scholar]
- Tubaro, A; Sosa, S; Carbonatto, M; Altinier, G; Vita, F; Melato, M; Satake, M; Yasumoto, T. Oral and intraperitoneal acute toxicity studies of yessotoxin and homoyessotoxins in mice. Toxicon 2003, 41, 783–792. [Google Scholar]
- Alfonso, A; de la Rosa, L; Vieytes, MR; Yasumoto, T; Botana, LM. Yessotoxin, a novel phycotoxin, activates phosphodiesterase activity. Effect of yessotoxin on cAMP levels in human lymphocytes. Biochem Pharmacol 2003, 65, 193–208. [Google Scholar]
- Terao, K; Ito, E; Oarada, M; Murata, M; Yasumoto, T. Histopathological studies on experimental marine toxin poisoning-5. The effects in mice of yessotoxin isolated from Patinopecten yessoensis and of a desulfated derivative. Toxicon 1990, 28, 1095–1104. [Google Scholar]
- Ogino, H; Kumagai, M; Yasumoto, T. Toxicological evaluation of yessotoxin. Nat Toxins 1997, 5, 255–259. [Google Scholar]
- Aune, T; Sørby, R; Yasumoto, T; Ramstad, H; Landsverk, T. Comparison of oral and intraperitoneal toxicity of yessotoxin towards mice. Toxicon 2002, 40, 77–82. [Google Scholar]
- Wolf, LW; LaRegina, MC; Tolbert, DL. A behavioral study of the development of hereditary cerebellar ataxia in the shaker rat mutant. In Behav Brain Res; 1996; Volume 75, pp. 67–81. [Google Scholar]
- Franchini, A; Marchesini, E; Poletti, R; Ottaviani, E. Acute toxic effect of the algal yessotoxin on Purkinje cells from the cerebellum of Swiss CD1 mice. Toxicon 2004, 43, 347–352. [Google Scholar]
- Malaguti, C; Ciminiello, P; Fattorusso, E; Rossini, GP. Caspase activation and death induced by yessotoxin in HeLa cells. Toxicol in Vitro 2002, 16, 357–363. [Google Scholar]
- Bianchi, C; Fato, R; Angelin, A; Trombetti, F; Ventrella, V; Borgatti, AR; Fattorusso, E; Ciminiello, P; Bernardi, P; Lenaz, G; Castelli, GP. Yessotoxin, a shellfish biotoxin, is a potent inducer of the permeability transition in isolated mitochondria and intact cells. Biochim Biophys Acta 2004, 1656, 139–147. [Google Scholar]
- Perez-Gomez, A; Ferrero-Gutierrez, A; Novelli, A; Franco, JM; Paz, B; Fernandez-Sanchez, MT. Potent neurotoxic action of the shellfish biotoxin yessotoxin on cultured cerebellar neurons. Toxicol Sci 2006, 90, 168–177. [Google Scholar]
- Orsi, CF; Colombari, B; Callegari, F; Todaro, AM; Ardizzoni, A; Rossini, GP; Blasi, E; Peppoloni, S. Yessotoxin inhibits phagocytic activity of macrophages. Toxicon 2010, 55, 265–273. [Google Scholar]
- Korsnes, MS; Hetland, DL; Espenes, A; Tranulis, M; Aune, T. Apoptotic events by yessotoxin in myoblast cell lines from rat and mouse. Toxicol in Vitro 2006, 20, 1077–1087. [Google Scholar]
- Lynn, DE; Dougherty, EM; McClintock, JT; Loeb, M. Development of cell lines from various tissues of Lepidoptera. In Invertebrate and Fish Tissue Culture; Kuroda, Y, Kurstak, E, Maramorosch, K, Eds.; Japan Scientific Societies Press: Berlin, Germany, 1988; pp. 239–242. [Google Scholar]
- Malagoli, D. Cell death in the IPLB-LdFB insect cell line: Facts and implications. Curr Phar Des 2008, 14, 126–130. [Google Scholar]
- Malagoli, D; Marchesini, E; Ottaviani, E. Lysosomes as the target of yessotoxin in invertebrate and vertebrate cell lines. Toxicol Lett 2006, 167, 75–83. [Google Scholar]
- de la Rosa, LA; Alfonso, A; Filarino, N; Vieytes, MR; Botana, LM. Modulation of cytosolic calcium levels of human lymphocytes by yessotoxin, a novel marine phycotoxin. Biochem Pharmacol 2001, 61, 827–833. [Google Scholar]
- Malagoli, D; Casarini, L; Ottaviani, E. Algal toxin yessotoxin signalling pathways involve immunocyte mussel calcium channels. Cell Biol Int 2006, 30, 721–726. [Google Scholar]
- Draisci, R; Ferretti, E; Palleschi, L; Marchiafava, C; Poletti, R; Milandri, A; Ceredi, A; Pompei, M. High levels of yessotoxin in mussels and presence of yessotoxin and homoyessotoxin in dinoflagellates of the Adriatic Sea. Toxicon 1999, 37, 1187–1193. [Google Scholar]
- Morton, SL; Vershinin, A; Leighfield, T; Smith, L; Quilliam, M. Identification of yessotoxin in mussels from the Caucasian Black Sea Coast of the Russian Federation. Toxicon 2007, 50, 581–584. [Google Scholar]
- Amzil, Z; Sibat, M; Royer, F; Savar, V. First report on azaspiracid and yessotoxin groups detection in French shellfish. Toxicon 2008, 52, 39–48. [Google Scholar]
- Franchini, A; Milandri, A; Poletti, R; Ottaviani, E. Immunolocalization of yessotoxins in the mussel Mytilus galloprovincialis. Toxicon 2003, 41, 967–970. [Google Scholar]
- Aasen, J; Samdal, IA; Miles, CO; Dahl, E; Briggs, LR; Aune, T. Yessotoxins in Norwegian blue mussels (Mytilus edulis): Uptake from Protoceratium reticulatum, metabolism and depuration. Toxicon 2005, 45, 265–272. [Google Scholar]
- Malagoli, D; Ottaviani, E. Yessotoxin affects fMLP-induced cell shape changes in Mytilus galloprovincialis immunocytes. Cell Biol Int 2004, 28, 57–61. [Google Scholar]
- Aune, T; Aasen, JA; Miles, CO; Larsen, S. Effect of mouse strain and gender on LD(50) of yessotoxin. Toxicon 2008, 52, 535–540. [Google Scholar]
- Franchini, A; Marchesini, E; Poletti, R; Ottaviani, E. Lethal and sub-lethal yessotoxin dose-induced morpho-functional alterations in intraperitoneal injected Swiss CD1 mice. Toxicon 2004, 44, 83–90. [Google Scholar]
- Ring, NP; Addis, BJ. Thymoma: An integrated clinicopathological and immunohistochemical study. J Pathol 1986, 149, 327–337. [Google Scholar]
- Fukai, I; Masaoka, A; Hashimoto, T; Yamakawa, Y; Mizuno, T; Tanamura, O. Cytokeratins in normal thymus and thymic epithelial tumors. Cancer 1993, 71, 99–105. [Google Scholar]
- Palestro, G; Novero, D; Geuna, M; Chiarle, R; Chiusa, L; Pagano, M; Pich, A. Role of the perivascular epithelium in the histogenesis of Hassall’s corpuscles. A morphologic and immunohistological study. Int J Surg Pathol 1998, 6, 213–222. [Google Scholar]
- Kuo, T. Cytokeratin profiles of the thymus and thymomas: Histogenetic correlations and proposal for a histological classification of thymomas. Histopathology 2000, 36, 403–414. [Google Scholar]
- Yasumoto, T; Murata, M; Oshima, Y; Sano, M; Matsumoto, GK; Clardy, J. Diarrhetic shellfish toxin. Tetrahedon 1985, 41, 1019–1025. [Google Scholar]
- Kumagai, M; Yamagi, T; Murata, M; Yasumoto, T; Kat, M; Lassus, P; Rodriguez-Vazquez, JA. Okadaic acid as the causative toxin of diarretic shellfish poisoning in Europe. Agric Biol Chem 1986, 50, 2857–2863. [Google Scholar]
- Bialojan, C; Takai, A. Inhibitory effect of a marine-sponge toxin, okadaic acid, on protein phosphatases. Biochem J 1988, 256, 283–290. [Google Scholar]
- Edebo, L; Lange, S; Li, XP; Allenmark, S. Toxic mussels and okadaic acid induce rapid hypersecretion in the rat small intestine. APMIS 1988, 96, 1029–1035. [Google Scholar]
- Ito, E; Terao, K. Injury and recovery process of intestine caused by okadaic acid and related compounds. Nat Toxins 1994, 2, 371–377. [Google Scholar]
- Ito, E; Yasumoto, T; Takai, A; Imanishi, S; Harada, K. Investigation of the distribution and excretion of okadaic acid in mice using immunostaining method. Toxicon 2002, 40, 159–165. [Google Scholar]
- Franchini, A; Marchesini, E; Poletti, R; Ottaviani, E. Swiss mice CD1 fed on mussels contaminated by okadaic acid and yessotoxins: effects on thymus and spleen. Eur J Histochem 2005, 49, 73–82. [Google Scholar]
- Berven, G; Saetre, F; Halvorsen, K; Seglen, PO. Effects of the diarrhetic shellfish toxin, okadaic acid, on cytoskeletal elements, viability and functionality of rat liver and intestinal cells. Toxicon 2001, 39, 349–362. [Google Scholar]
- Fernandez, MT; Zitko, V; Gascon, S; Novelli, A. The marine toxin okadaic acid is a potent neurotoxin for cultured cerebellar neurons. Life Sci 1991, 49, PL157–162. [Google Scholar]
- Rossini, GP; Sgarbi, N; Malaguti, C. The toxic responses induced by okadaic acid involve processing of multiple caspase isoforms. Toxicon 2001, 39, 763–770. [Google Scholar]
- Leira, F; Alvarez, C; Vieites, JM; Vieytes, MR; Botana, LM. Characterization of distinct apoptotic changes induced by okadaic acid and yessotoxin in the BE(2)-M17 neuroblastoma cell line. Toxicol in Vitro 2002, 16, 23–31. [Google Scholar]
- Fujiki, H; Suganuma, M. Tumor promotion by inhibitors of protein phosphatases 1 and 2A: The okadaic acid class of compounds. Adv Cancer Res 1993, 61, 143–194. [Google Scholar]
- Fujiki, H; Suganuma, M. Unique features of the okadaic acid activity class of tumor promoters. J. Cancer Res. Clin. Oncol. 1999, 125, 150–155. [Google Scholar]
- Fujiki, H; Suganuma, M. Carcinogenic aspects of protein phosphatase 1 and 2A inhibitors. Prog Mol Subcell Biol 2009, 46, 221–254. [Google Scholar]
- Moore, RE; Scheuer, PJ. Palytoxin: A new marine toxin from a coelenterate. Science 1971, 172, 495–498. [Google Scholar]
- Deeds, JR; Schwartz, MD. Human risk associated with palytoxin exposure. Toxicon 2009. [Google Scholar] [CrossRef]
- Kodama, AM; Hokama, Y; Yasumoto, T; Fukui, M; Manea, SJ; Sutherland, N. Clinical and laboratory findings implicating palytoxin as cause of ciguatera poisoning due to Decapterus macrosoma (mackerel). Toxicon 1989, 27, 1051–1053. [Google Scholar]
- Okano, H; Masuoka, H; Kamei, S; Seko, T; Koyabu, S; Tsuneoka, K; Tamai, T; Ueda, K; Nakazawa, S; Sugawa, M; Sukuki, H; Watanabe, M; Yatani, R; Nakano, T. Rhabdomyolysis and myocardial damage induced by palytoxin, a toxin of blue humphead parrotfish. Intern Med 1998, 37, 330–333. [Google Scholar]
- Taniyama, S; Mahmud, Y; Terada, M; Takatani, T. Occurrence of a food poisoning incident by palytoxin from a serranid Epinephelus sp. in Japan. J Nat Toxins 2002, 11, 277–282. [Google Scholar]
- Sosa, S; Del Bavero, G; De Bortoli, M; Vita, F; Soranzo, MR; Beltramo, D; Ardizzone, M; Tubaro, A. Palytoxin toxicity after acute oral administration in mice. Toxicol Lett 2009, 191, 253–259. [Google Scholar]
- Vale, C; Alfonso, A; Sunol, C; Vieytes, MR; Botana, LM. Modulation of calcium entry and glutamate release in cultured cerebellar granule cells by palytoxin. J Neurosci Res 2006, 83, 1393–1406. [Google Scholar]
- Vale, C; Gómez-Limia, B; Vieytes, MR; Botana, LM. Mitogen-activated protein kinases regulate palytoxin-induced calcium influx and cytotoxicity in cultured neurons. Br J Pharmacol 2007, 152, 256–266. [Google Scholar]
- Didden, W; Rombke, J. Enchytraeids as indicator organisms for chemical stress in terrestrial ecosystems. Ecotoxicol Environ Safety 2001, 50, 25–43. [Google Scholar]
- Franchini, A; Marchetti, M. The effects of okadaic acid on Enchytraeus crypticus (Annelida: Oligochaeta). Inv Surv J 2006, 3, 111–117. [Google Scholar]
- Cancio, I; ap Gwynn, I; Ireland, MP; Cajaraville, MP. Lysosomal origin of the chloragosomes in the chloragogenous tissue of the earthworm Eisenia foetida: Cytochemical demonstration of acid phosphatase activity. Histochem J 1995, 27, 591–596. [Google Scholar]
- Franchini, A; Ottaviani, E. Age-related toxic effects and recovery from okadaic acid treatment in Enchytraeus crypticus (Annelida: Oligochaeta). Toxicon 2008, 52, 115–121. [Google Scholar]
- Gleibs, S; Mebs, D. Distribution and sequestration of palytoxin in coral reef animals. Toxicon 1999, 37, 1521–1527. [Google Scholar]
- Malagoli, D; Casarini, L; Ottaviani, E. Effects of the marine toxins okadaic acid and palytoxin on mussel phagocytosis. Fish Shellfish Immunol 2008, 24, 180–186. [Google Scholar]
- Malagoli, D; Gobba, F; Ottaviani, E. Effects of 50 Hz magnetic fields on the signalling pathways of fMLP-induced shape changes in invertebrate immunocytes: The activation of an alternative “stress pathway”. Biochim Biophys Acta 2003, 1620, 185–190. [Google Scholar]
- Bantle, JA; Sabourin, TD. Standard Guide for Conducting the Frog Embryo Teratogenesis Assay-Xenopus (FETAX); ASTM: Philadelphia, PA, USA, 1991. [Google Scholar]
- Bantle, JA. FETAX-A developmental toxicity assay using frog embryos. In Fundamentals of Aquatic Toxicology, 2nd ed; Rand, GM, Ed.; Taylor and Francis: Washington DC, USA, 1995; pp. 207–228. [Google Scholar]
- Mann, RM; Bidwell, JR. Application of the FETAX protocol to assess the developmental toxicity of nonylphenol ethoxylate to Xenopus laevis and two Australian frogs. Aquat Toxicol 2000, 51, 19–29. [Google Scholar]
- Prati, M; Biganzoli, E; Boracchi, P; Tesauro, M; Monetti, C; Bernardini, G. Ecotoxicological soil evaluation by FETAX. Chemosphere 2000, 41, 1621–1628. [Google Scholar]
- Garber, EAE; Erb, JL; Magner, J; Larsen, G. Low levels of sodium and potassium in thewater from wetlands in Minnesota that contained malformed frogs affect the rate of Xenopus development. Environ Monit Assess 2004, 90, 45–64. [Google Scholar]
- Bernardini, G; Prati, M; Gornati, R. FETAX, a versatile tool in toxicology, can be conveniently integrated with molecular biology techniques. Appl Herpetol 2005, 2, 287–295. [Google Scholar]
- Casarini, L; Franchini, A; Malagoli, D; Ottaviani, E. Evaluation of the effects of the marine toxin okadaic acid by using FETAX assay. Toxicol Lett 2007, 145–151. [Google Scholar]
- Franchini, A; Casarini, L; Ottaviani, E. Toxicological effects of marine palytoxin evaluated by FETAX assay. Chemosphere 2008, 73, 267–271. [Google Scholar]
- Franchini, A; Casarini, L; Malagoli, D; Ottaviani, E. Expression of the genes siamois, engrailed-2, bmp4 and myf5 during Xenopus development in presence of the marine toxins okadaic acid and palytoxin. Chemosphere 2009, 77, 308–312. [Google Scholar]
- Nieuwkoop, PD; Faber, J. Normal Tables of Xenopus laevis (Daudin): A Systematic and Chronological Survey of the Development from the Fertilized Egg Till the End of Metamorphosis; North-Holland Publishing Company: Amsterdam, The Netherland, 1967. [Google Scholar]
- Hemmati-Brivanlou, A; de la Torre, JR; Holt, C; Harland, RM. Cephalic expression and molecular characterization of Xenopus En-2. Development 1991, 111, 715–724. [Google Scholar]
- Brannon, M; Kimelman, D. Activation of Siamois by the Wnt pathway. Dev Biol 1996, 180, 344–347. [Google Scholar]
- Dale, L; Wardle, FC. A gradient of BMP activity specifies dorsal-ventral fates in early Xenopus embryos. Semin Cell Dev Biol 1999, 10, 319–326. [Google Scholar]
- McGrew, LL; Takemaru, K; Bates, R; Moon, RT. Direct regulation of the Xenopus engrailed-2 promoter by the Wnt signaling pathway, and a molecular screen for Wnt-responsive genes, confirm a role for Wnt signaling during neural patterning in Xenopus. Mech Dev 1999, 87, 21–32. [Google Scholar]
- Chanoine, C; Hardy, S. Xenopus muscle development: From primary to secondary myogenesis. Dev Dyn 2003, 226, 12–23. [Google Scholar]
- Borello, U; Berarducci, B; Murphy, P; Bajard, L; Buffa, V; Piccolo, S; Buckingham, M; Cossu, G. The Wnt/b-catenin pathway regulates Gli-mediated Myf5 expression during somitogenesis. Development 2006, 133, 3723–3732. [Google Scholar]
- Ishibashi, H; Matsumura, N; Hanafusa, H; Matsumoto, K; De Robertis, EM; Kuroda, H. Expression of Siamois and Twin in the blastula Chordin/Noggin signaling center is required for brain formation in Xenopus laevis embryos. Mech Dev 2008, 125, 58–66. [Google Scholar]












© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Franchini, A.; Malagoli, D.; Ottaviani, E. Targets and Effects of Yessotoxin, Okadaic Acid and Palytoxin: A Differential Review. Mar. Drugs 2010, 8, 658-677. https://doi.org/10.3390/md8030658
Franchini A, Malagoli D, Ottaviani E. Targets and Effects of Yessotoxin, Okadaic Acid and Palytoxin: A Differential Review. Marine Drugs. 2010; 8(3):658-677. https://doi.org/10.3390/md8030658
Chicago/Turabian StyleFranchini, Antonella, Davide Malagoli, and Enzo Ottaviani. 2010. "Targets and Effects of Yessotoxin, Okadaic Acid and Palytoxin: A Differential Review" Marine Drugs 8, no. 3: 658-677. https://doi.org/10.3390/md8030658
APA StyleFranchini, A., Malagoli, D., & Ottaviani, E. (2010). Targets and Effects of Yessotoxin, Okadaic Acid and Palytoxin: A Differential Review. Marine Drugs, 8(3), 658-677. https://doi.org/10.3390/md8030658