Seaweed Polysaccharides and Derived Oligosaccharides Stimulate Defense Responses and Protection Against Pathogens in Plants

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Structure of Seaweed Polysaccharides and Derived Oligosaccharides
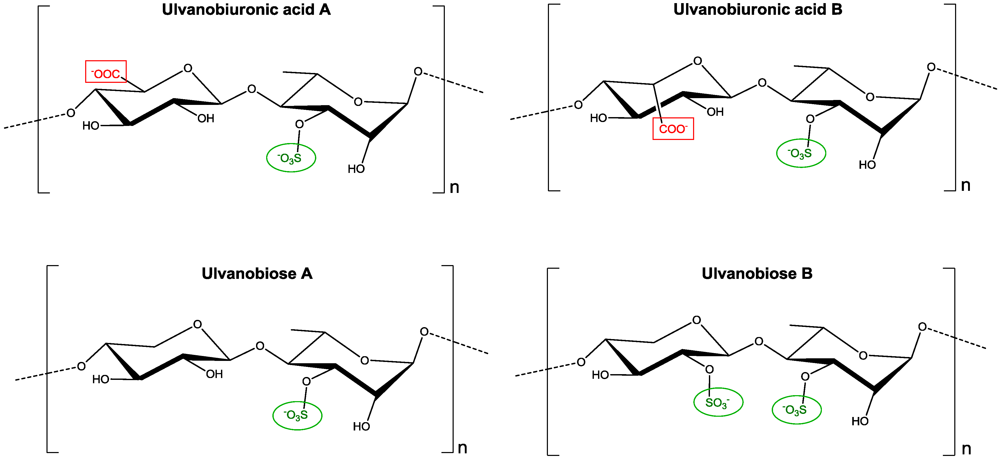
2.1. Ulvans and Oligo-Ulvans

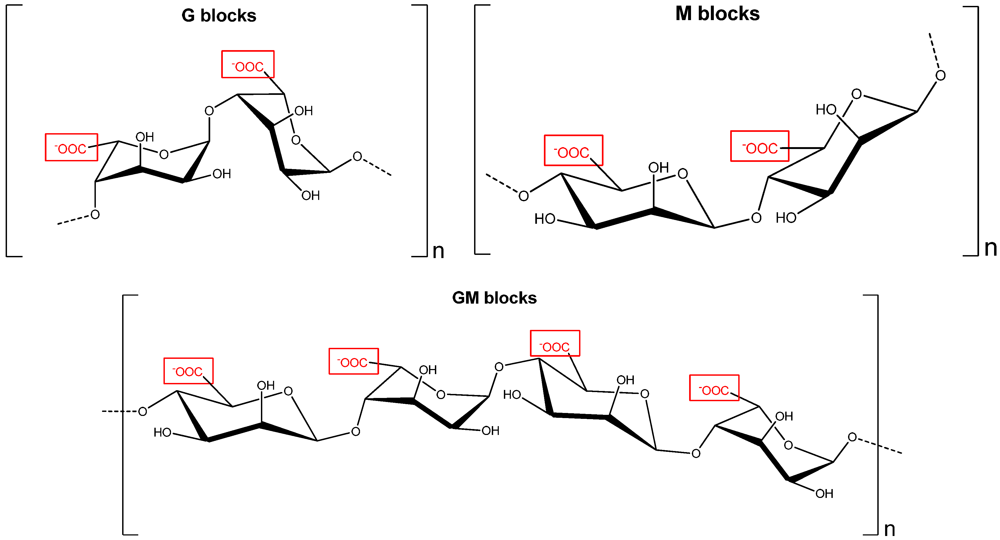
2.2. Alginates and Oligo-Alginates

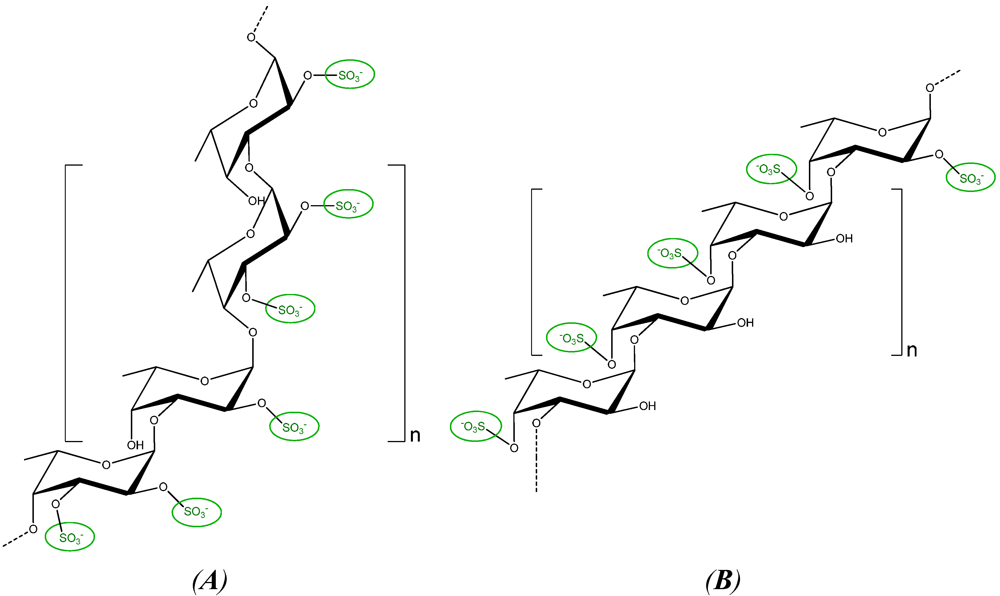
2.3. Fucans and Oligo-Fucans

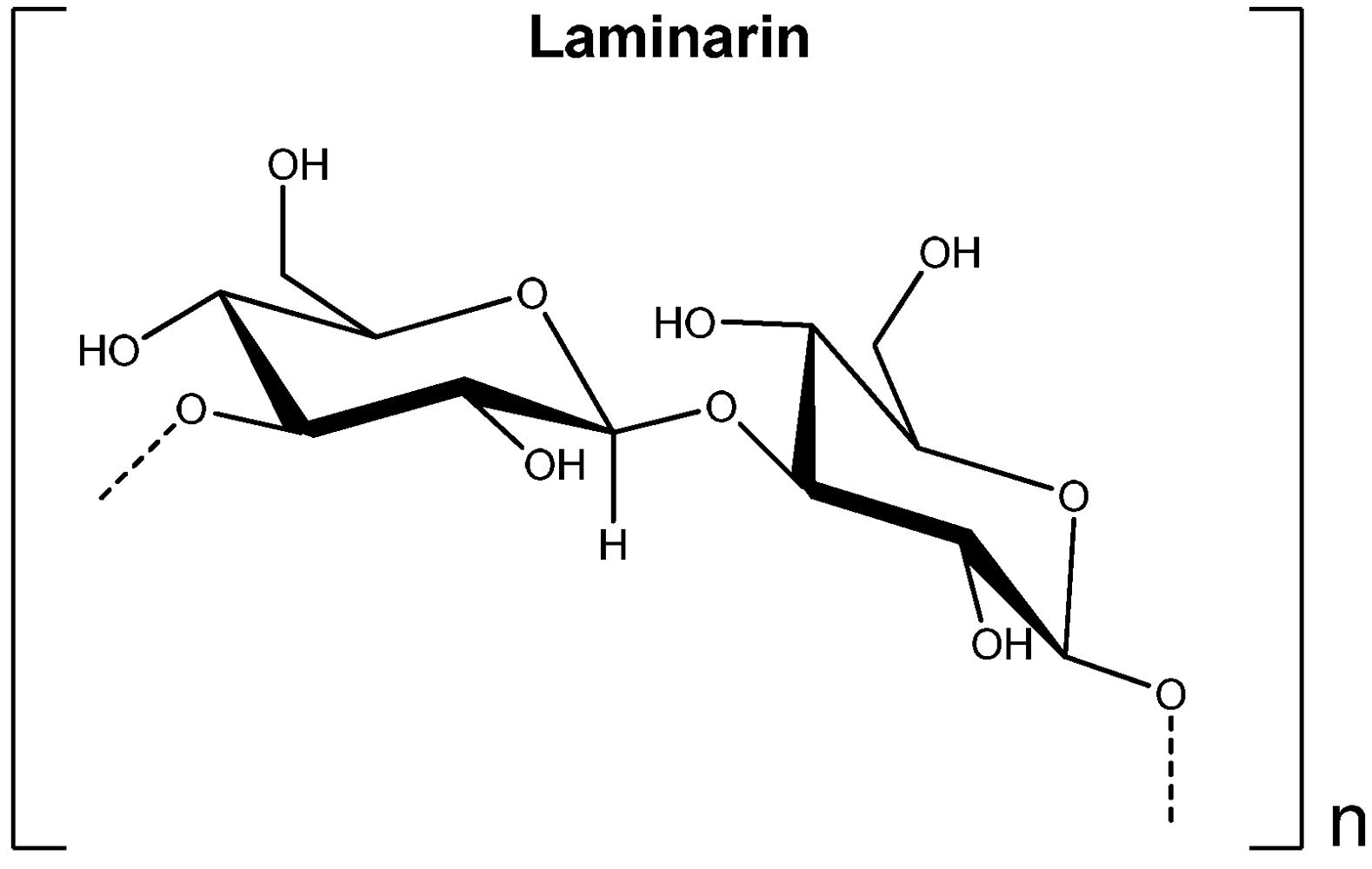
2.4. Laminarin

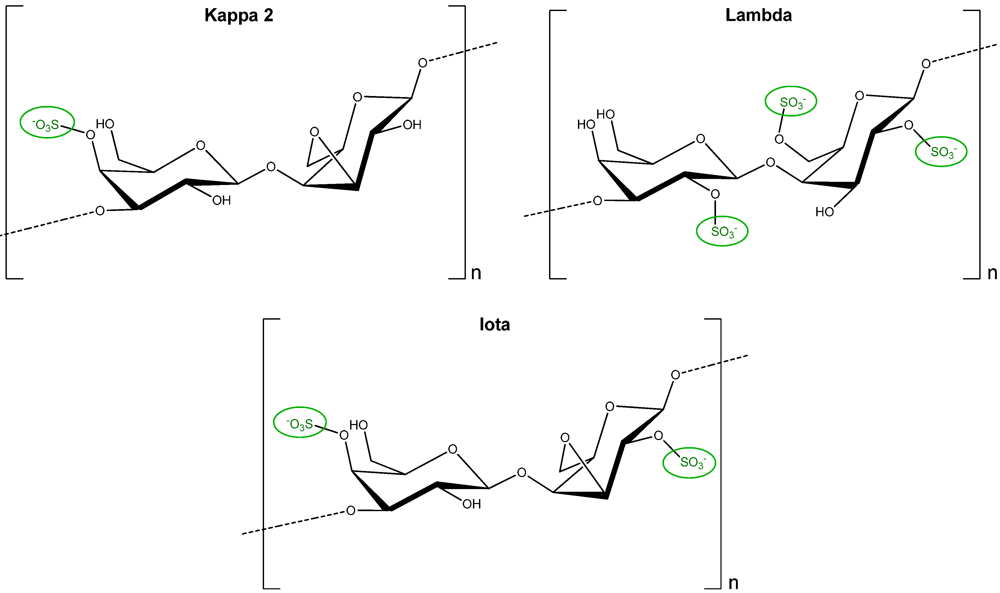
2.5. Carrageenans and Oligo-Carrageenans

3. Polysaccharides and Derived Oligosaccharides Stimulate Defense Responses and Protection Against Pathogens
3.1. Ulvans and Oligo-Ulvans
3.2. The Oligo-Alginate Poly-Ma
3.3. Fucans and Oligo-Fucans
3.4. Laminarin
3.5. Carrageenans and Oligo-Carrageenans
4. Conclusions
Acknowledgments
References
- Jiao, G.; Yu, G.; Zhang, J.; Ewart, S.H. Chemical structures and bioactivities of sulphated polysaccharides from marine algae. Mar. Drugs 2011, 9, 196–223. [Google Scholar]
- Rioux, L.E.; Turgeon, S.L.; Beaulieu, M. Characterization of polysaccharides extracted from brown seaweeds. Carbohydr. Polym. 2007, 69, 530–537. [Google Scholar]
- Patier, P.; Yvin, J.C.; Kloareg, B.; Liénart, Y.; Rochas, C. Seaweed liquid fertilizer from Ascophyllum nosodosum contains elicitors of plant glycanases. J. Appl. Phycol. 1993, 5, 343–349. [Google Scholar]
- Patier, P.; Potin, P.; Rochas, C.; Kloareg, B.; Yvin, J.C.; Liénart, Y. Free and silica-bound oligokappa-carrageenans elicit laminarase activity in Rubus cells and protoplasts. Plant Sci. 1995, 110, 27–35. [Google Scholar]
- Lahaye, M.; Robic, A. Structure and functional properties of ulvan, a polysaccharide from green seaweeds. Biomacromolecules 2007, 8, 1765–1774. [Google Scholar]
- Briand, X.; Cluzet, S.; Dumas, B.; Esquerré-Tugayé, M.-T.; Salamagne, S. Ulvans as Activators of Plant Defense and Resistance Reactions Against Biotic and Abiotic Stresses. US Patent 7,820,176, 26 October 2010. [Google Scholar]
- Briand, X.; Cluzet, S.; Dumas, B.; Esquerré-Tugayé, M.-T.; Salamagne, S. Use of Ulvans as Elicitors of Mechanisms for Nitrogen Absorption and Protein Synthesis. US Patent 7,892,311, 22 February 2011. [Google Scholar]
- Jaulneau, V.; Lafitte, C.; Jacquet, C.; Fournier, S.; Salamagne, S.; Briand, X.; Esquerré-Tugayé, M.-T.; Dumas, B. Ulvan, a sulphated polysaccharide from green algae, activates plant immunity through the jasmonic acid signaling pathway. J. Biomed. Biotechnol. 2010, 2010, 525291:1–525291:11. [Google Scholar]
- Mabeau, S.; Kloareg, B. Isolation and analysis of the cell walls of brown algae: Fucus spiralis, F. ceranoides, F. vesiculosus, F. serratus, Bifurcaria bifurcata and Laminaria digitata. J. Exp. Bot. 1986, 38, 1573–1580. [Google Scholar] [CrossRef]
- Chandía, N.P.; Matsuhiro, B.; Vázquez, A.E. Alginic acids in Lessonia trabeculata: Characterization by formic cid hydrolysis and FT-IR spectroscopy. Carbohydr. Polym. 2001, 46, 81–87. [Google Scholar]
- Laporte, D.; Vera, J.; Chandía, N.P.; Zúñiga, E.A.; Matsuhiro, B.; Moenne, A. Structurally unrelated algal oligosaccharides differentially stimulate growth and defense against tobacco mosaic virus in tobacco plants. J. Appl. Phycol. 2007, 19, 79–88. [Google Scholar]
- Lu, F.; Wei, X.; Zhao, R. Fucoidans: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar]
- Chevolot, L.; Mulloy, B.; Ratiskol, J.; Foucault, A.; Colliec-Jouault, S. A dissacharide repeat unit is the major structure in fucoidans from two species of brown algae. Carbohydr. Res. 2001, 330, 529–535. [Google Scholar]
- Chandía, N.P.; Matsuhiro, B. Characterization of a fucoidan from Lessonia vadosa (Phaeophyta) and its anticoagulant and elicitor properties. Int. J. Biol. Macromol. 2008, 42, 235–240. [Google Scholar]
- Klarzynski, O.; Descamps, V.; Plesse, B.; Yvin, J.C.; Kloareg, B.; Fritig, B. Sulphated fucan oligosaccharides elicit defense responses in tobacco and local and systemic resistance against tobacco mosaic virus. Mol. Plant Microbe Interact. 2003, 16, 1156–1122. [Google Scholar]
- O’Sullivan, L.; Murphy, B.; McLoughlin, P.; Duggan, P.; Lawlor, P.G.; Hughes, H.; Gardiner, G.E. Prebiotics from marine macroalgae for human and animal health applications. Mar. Drugs 2010, 8, 2038–2064. [Google Scholar]
- Chizhov, A.O.; Dell, A.; Morris, H.R.; Reason, A.J.; Haslam, S.M.; McDowell, R.A.; Chizhov, O.S.; Usov, A.I. Structural analysis of laminarans by MALDI and FAB mass spectrometry. Carbohydr. Res. 1998, 310, 203–210. [Google Scholar]
- McCandless, E.L.; Craigie, J.S. Sulphated polysaccharides in red and brown algae. Planta 1979, 112, 201–212. [Google Scholar]
- Moenne, A. Composition and Method to Stimulate Growth and Defense Against Pathogens in Plants. US Patent 12,666,700, 8 July 2009. [Google Scholar]
- Paulert, R.; Ebbinghaus, D.; Urlass, C.; Moerschbacher, M. Priming of the oxidative burst in rice and wheat cell cultures by ulvan, a polysaccharide from green macroalgae, and enhanced resistance against powdery mildew in wheat and barley plants. Plant Pathol. 2010, 59, 634–642. [Google Scholar]
- Cluzet, S.; Torregrosa, C.; Jacquet, C.; Lafitte, C.; Fournier, J.; Mercier, L.; Salamagne, S.; Briand, X.; Esquerré-Tugayé, M.T.; Dumas, B. Gene expression profiling and protection of Medicago truncatula against a fungal infection in response to an elicitor from green algae Ulva sp. Plant Cell Environ. 2004, 27, 917–928. [Google Scholar]
- Jaulneau, V.; Lafitte, C.; Corio-Costet, M.F.; Stadnik, M.J.; Salamagne, S.; Briand, X.; Esquerré-Tugayé, M.T.; Dumas, B. An Ulva armoricana extract protects plants against three powdery mildew pathogens. Eur. J. Plant Pathol. 2011, 131, 393–401. [Google Scholar]
- Paulert, R.; Talamini, V.; Cassolato, J.E.F.; Duarte, M.E.R.; Noseda, M.D.; Smania, A.; Stadnik, M.J. Effects of sulphated polysaccharide and alcoholic extracts from green seaweeds Ulva fasciata on anthracnose severity and growth of common bean (Phaseolus vulgaris L.). J. Plant Dis. Prot. 2009, 116, 263–270. [Google Scholar]
- Chandía, N.P.; Matsuhiro, B.; Mejías, E.; Moenne, A. Alginic acids in Lessonia vadosa: Partial hydrolysis and elicitor properties of the polymannuronic acid fraction. J. Appl. Phycol. 2004, 16, 127–133. [Google Scholar]
- Sels, J.; Mathys, J.; de Coninck, B.M.; Cammue, B.P.; de Bolle, M.F. Plant pathogenesis-related (PR) proteins: A focus on PR peptides. Plant Physiol. Biochem. 2008, 46, 941–950. [Google Scholar]
- Chong, J.; Baltz, R.; Schmitt, C.; Belfa, R.; Fritig, B.; Saindrenan, P. Down-regulation of a pathogen-responsive tobacco UDP-glucose:phenylpropanoid glucosyltransferase reduces scopoletin glucoside accumulation, enhances oxidative stress, and weakens virus resistance. Plant Cell 2002, 14, 1093–1107. [Google Scholar]
- Klarzynski, O.; Plesse, B.; Joubert, J.M.; Yvin, J.C.; Kopp, M.; Kloareg, B.; Fritig, B. Linear β-1,3 glucans are elicitors of defense responses in tobacco. Plant Physiol. 2000, 124, 1027–1037. [Google Scholar]
- Aziz, A.; Poinssot, B.; Daire, X.; Adrian, M.; Bézier, A.; Lambert, B.; Joubert, J.M.; Pugin, A. Laminarin elicits defense responses in grapevine and induces protection against Botrytis cinerea and Plasmopara viticola. Mol. Plant Microbe Interact. 2003, 16, 1118–1128. [Google Scholar]
- Ménard, R.; Alban, S.; de Ruffray, P.; Jamois, F.; Franz, G.; Fritig, B.; Yvin, J.C.; Kauffmann, S. β-1,3 glucan sulfate, induces salicylic acid signaling pathway in tobacco and Arabidopsis. Plant Cell 2004, 16, 3020–3032. [Google Scholar]
- Trouvelot, S.; Varnier, A.L.; Allègre, M.; Mercier, L.; Baillieul, F.; Arnould, C.; Gianinazzi-Pearson, V.; Klarzynski, O.; Joubert, J.M.; Pugin, A.; et al. A β-1,3 glucan sulfate induces resistance in grapevine against Plasmopara viticola through priming of defense responses, including HR-like cell death. Mol. Plant Microbe Interact. 2008, 21, 232–243. [Google Scholar]
- Mercier, L.; Lafitte, C.; Borderies, G.; Briand, X.; Esquerré-Tugayé, M.T.; Fournier, J. The algal polysaccharide carrageenans can act as an elicitor of plant defence. New Phytol. 2001, 149, 43–51. [Google Scholar]
- Vera, J. University of Santiago, Santiago, Chile. Depolymerization of carrageenan iota is required to induce protection against TMV infection in tobacco plants. 2011. Unpublished work. [Google Scholar]
- Vera, J.; Castro, J.; González, A.; Barrientos, H.; Matsuhiro, B.; Arce, P.; Zúñiga, G.; Moenne, A. Long term protection against tobacco mosaic virus induced by the marine alga oligo-sulphated-galactan Poly-Ga in tobacco plants. Mol. Plant Pathol. 2011, 12, 437–447. [Google Scholar]
- Vera, J.; Castro, J.; Contreras, R.A.; González, A.; Moenne, A. Oligo-carrageenans induced a long-term and broad-range protection against pathogens and the reversion of infections in tobacco plants (var. Xanthi). Physiol. Mol. Plant Pathol. 2011, in press. [Google Scholar]
- Castro, J.; Vera, J.; González, A.; Moenne, A. Oligo-carrageenans stimulate growth by enhancing photosynthesis, basal metabolism and cell cycle in tobacco plants (var. Burley). J. Plant Growth Regul. 2011. [Google Scholar]
- Moenne, A.; Castro, J.; Vera, J. Method to Stimulate Carbon Fixation in Plants with an Aqueous Solution of Oligo-Carrageenans Selected from Kappa1, Kappa2, Lambda and Iota. US Patent 12,911,790, 5 May 2011. [Google Scholar]
- Gonzalez, A. University of Santiago, Santiago, Chile. Oligo-carrageenans stimulate growth by enhancing photosynthesis and basal metabolism in eucalyptus trees (Eucalyptus globulus). 2011. Unpublished work. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Vera, J.; Castro, J.; Gonzalez, A.; Moenne, A. Seaweed Polysaccharides and Derived Oligosaccharides Stimulate Defense Responses and Protection Against Pathogens in Plants. Mar. Drugs 2011, 9, 2514-2525. https://doi.org/10.3390/md9122514
Vera J, Castro J, Gonzalez A, Moenne A. Seaweed Polysaccharides and Derived Oligosaccharides Stimulate Defense Responses and Protection Against Pathogens in Plants. Marine Drugs. 2011; 9(12):2514-2525. https://doi.org/10.3390/md9122514
Chicago/Turabian StyleVera, Jeannette, Jorge Castro, Alberto Gonzalez, and Alejandra Moenne. 2011. "Seaweed Polysaccharides and Derived Oligosaccharides Stimulate Defense Responses and Protection Against Pathogens in Plants" Marine Drugs 9, no. 12: 2514-2525. https://doi.org/10.3390/md9122514
APA StyleVera, J., Castro, J., Gonzalez, A., & Moenne, A. (2011). Seaweed Polysaccharides and Derived Oligosaccharides Stimulate Defense Responses and Protection Against Pathogens in Plants. Marine Drugs, 9(12), 2514-2525. https://doi.org/10.3390/md9122514